
Modulation of GLO1 Expression Affects Malignant Properties of Cells

 and
and 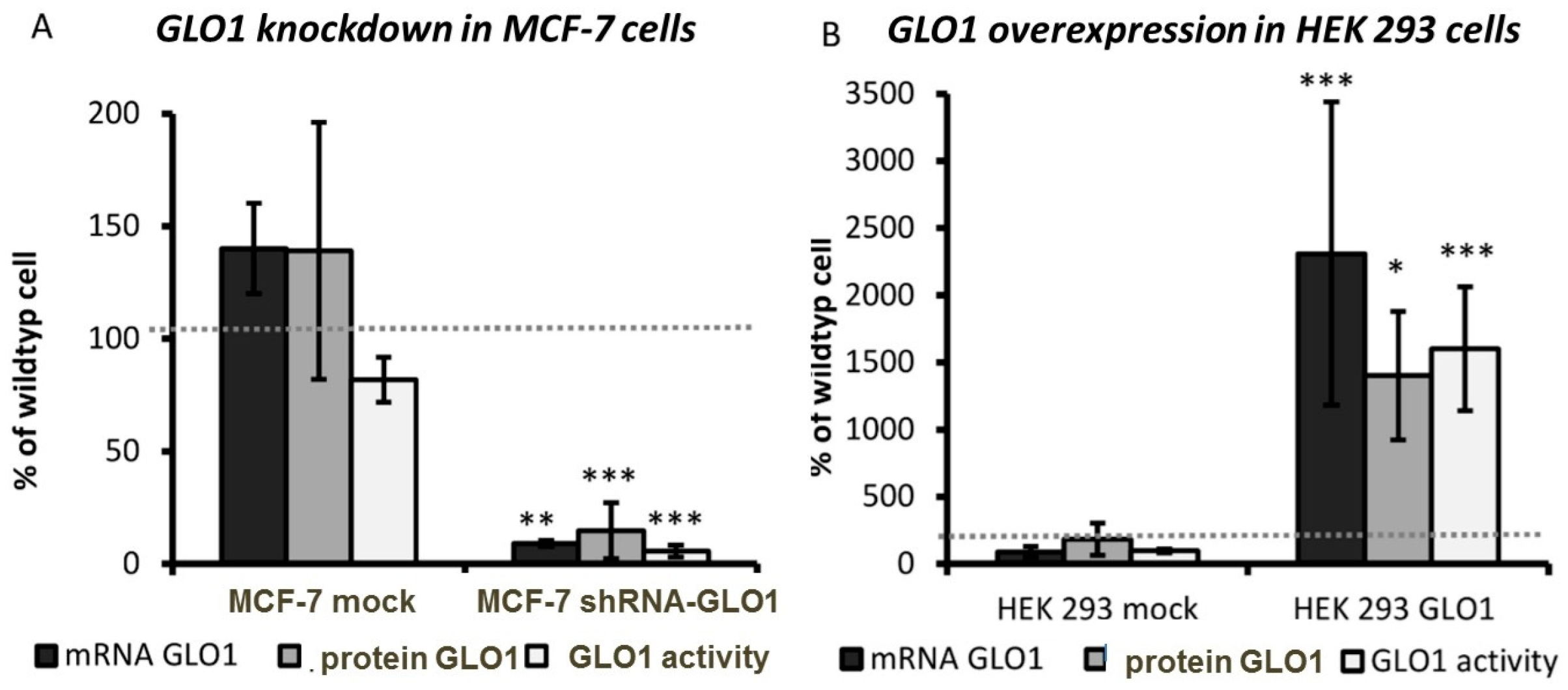{kind=link}
{kind=link}
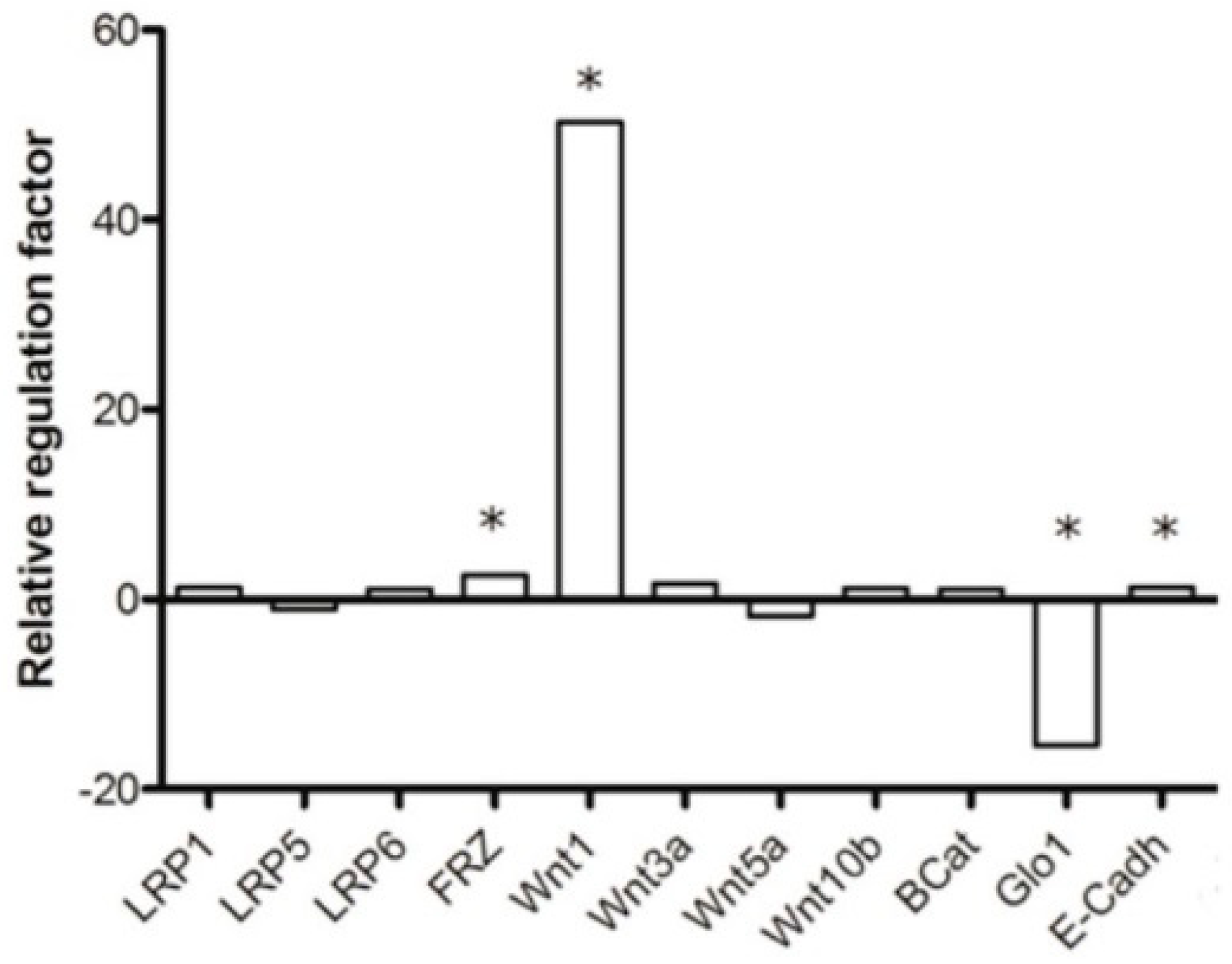
{kind=link}
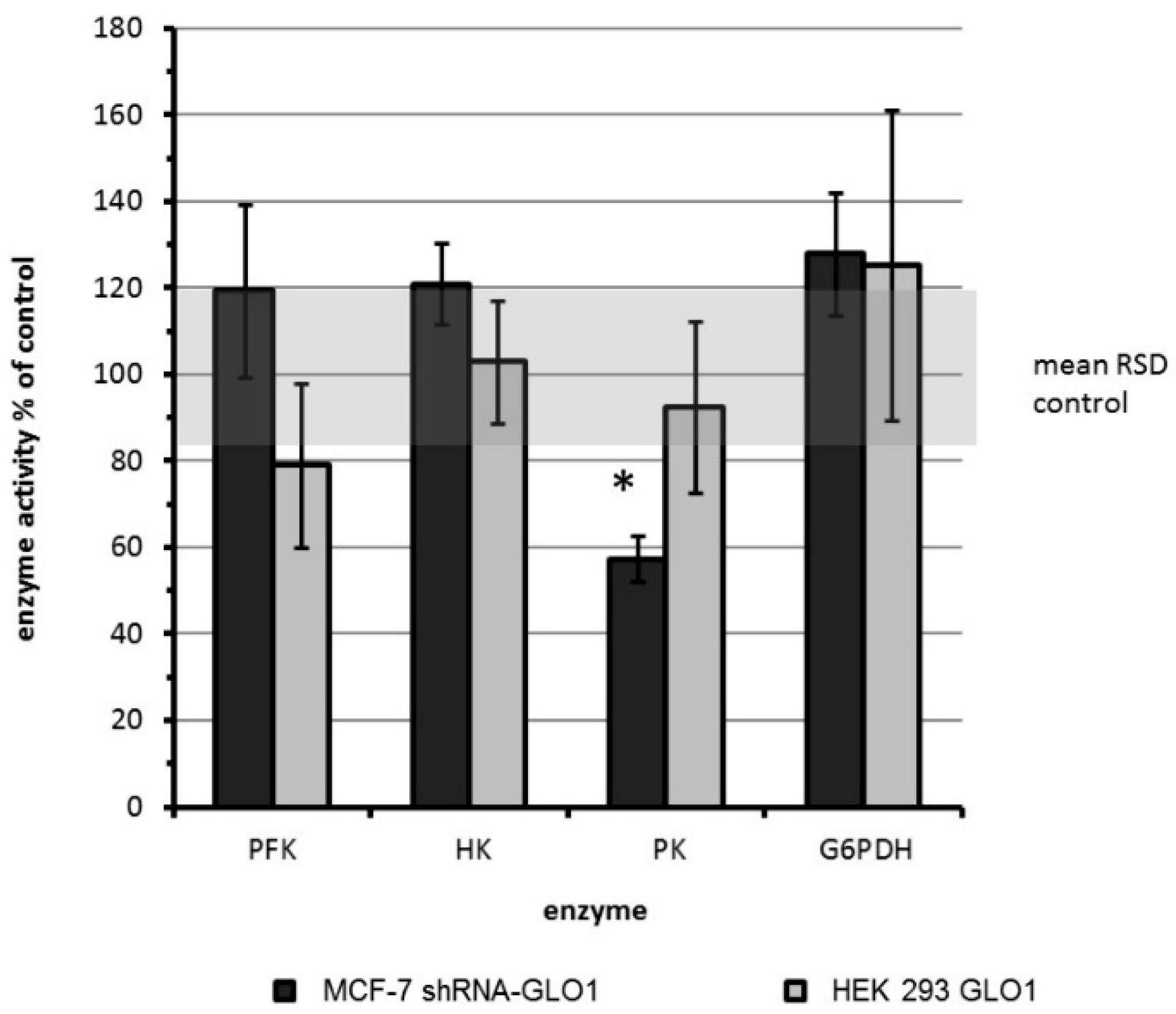
{kind=link}
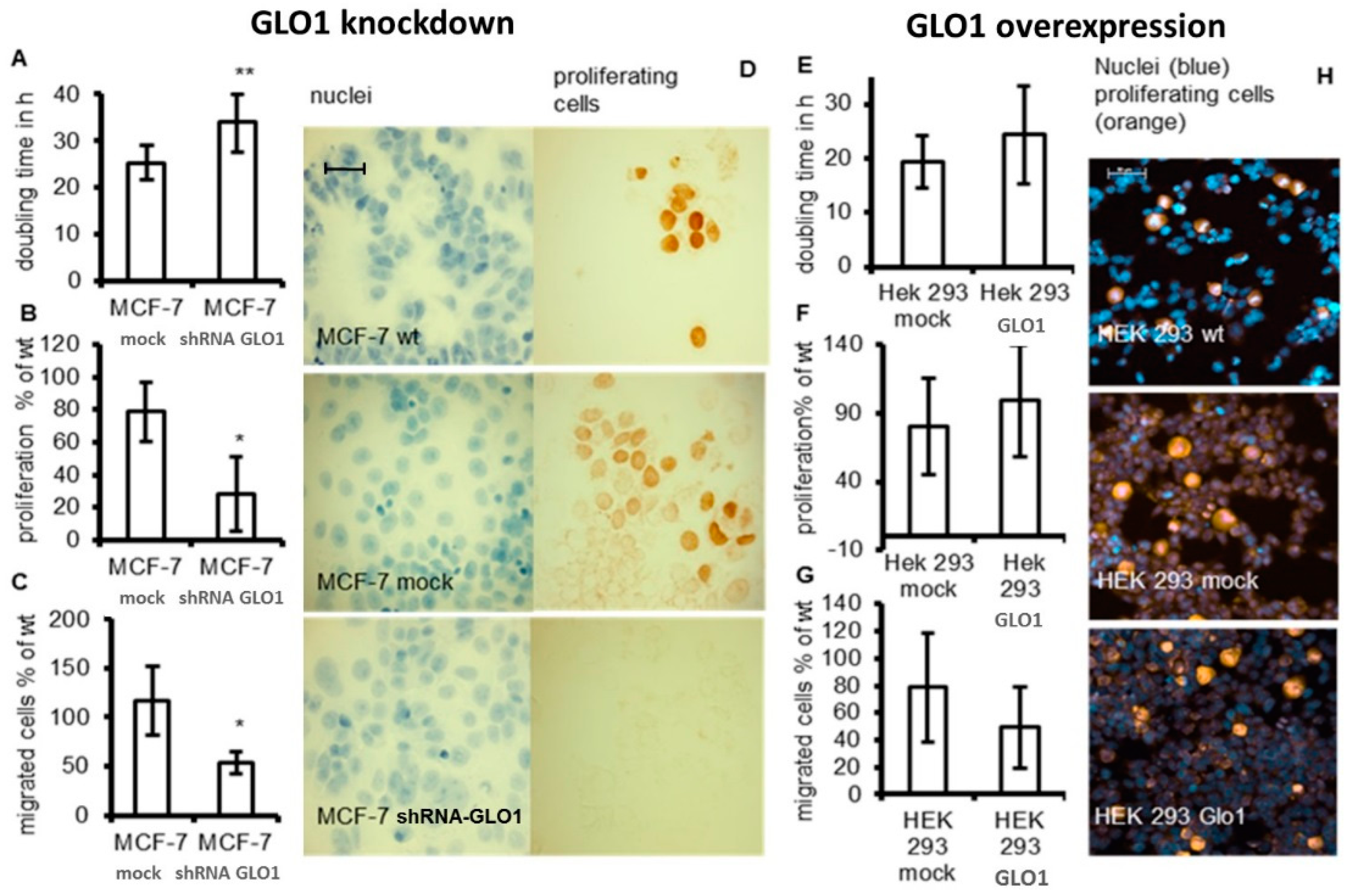
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Hutschenreuther, A.; Bigl, M.; Hemdan, N.Y.A.; Debebe, T.; Gaunitz, F.; Birkenmeier, G. Modulation of GLO1 Expression Affects Malignant Properties of Cells. Int. J. Mol. Sci. 2016, 17, 2133. https://doi.org/10.3390/ijms17122133
Hutschenreuther A, Bigl M, Hemdan NYA, Debebe T, Gaunitz F, Birkenmeier G. Modulation of GLO1 Expression Affects Malignant Properties of Cells. International Journal of Molecular Sciences. 2016; 17(12):2133. https://doi.org/10.3390/ijms17122133
Chicago/Turabian StyleHutschenreuther, Antje, Marina Bigl, Nasr Y. A. Hemdan, Tewodros Debebe, Frank Gaunitz, and Gerd Birkenmeier. 2016. "Modulation of GLO1 Expression Affects Malignant Properties of Cells" International Journal of Molecular Sciences 17, no. 12: 2133. https://doi.org/10.3390/ijms17122133
APA StyleHutschenreuther, A., Bigl, M., Hemdan, N. Y. A., Debebe, T., Gaunitz, F., & Birkenmeier, G. (2016). Modulation of GLO1 Expression Affects Malignant Properties of Cells. International Journal of Molecular Sciences, 17(12), 2133. https://doi.org/10.3390/ijms17122133