
Zinc Up-Regulates Insulin Secretion from β Cell-Like Cells Derived from Stem Cells from Human Exfoliated Deciduous Tooth (SHED)



Abstract
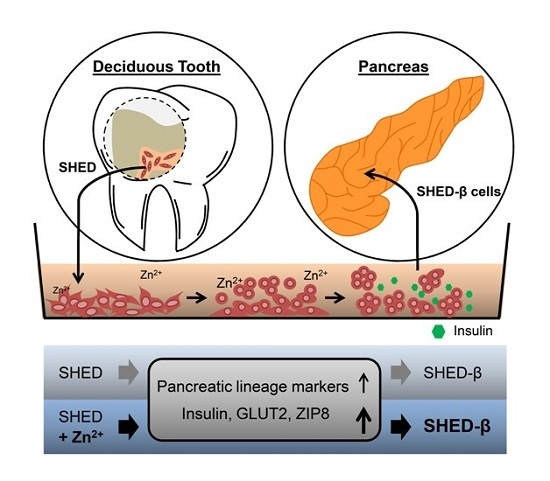
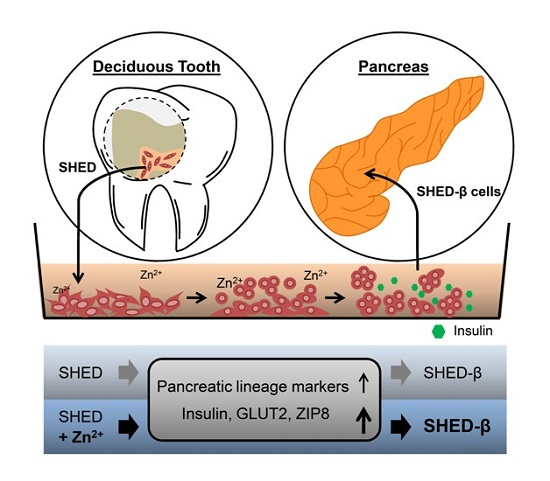
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
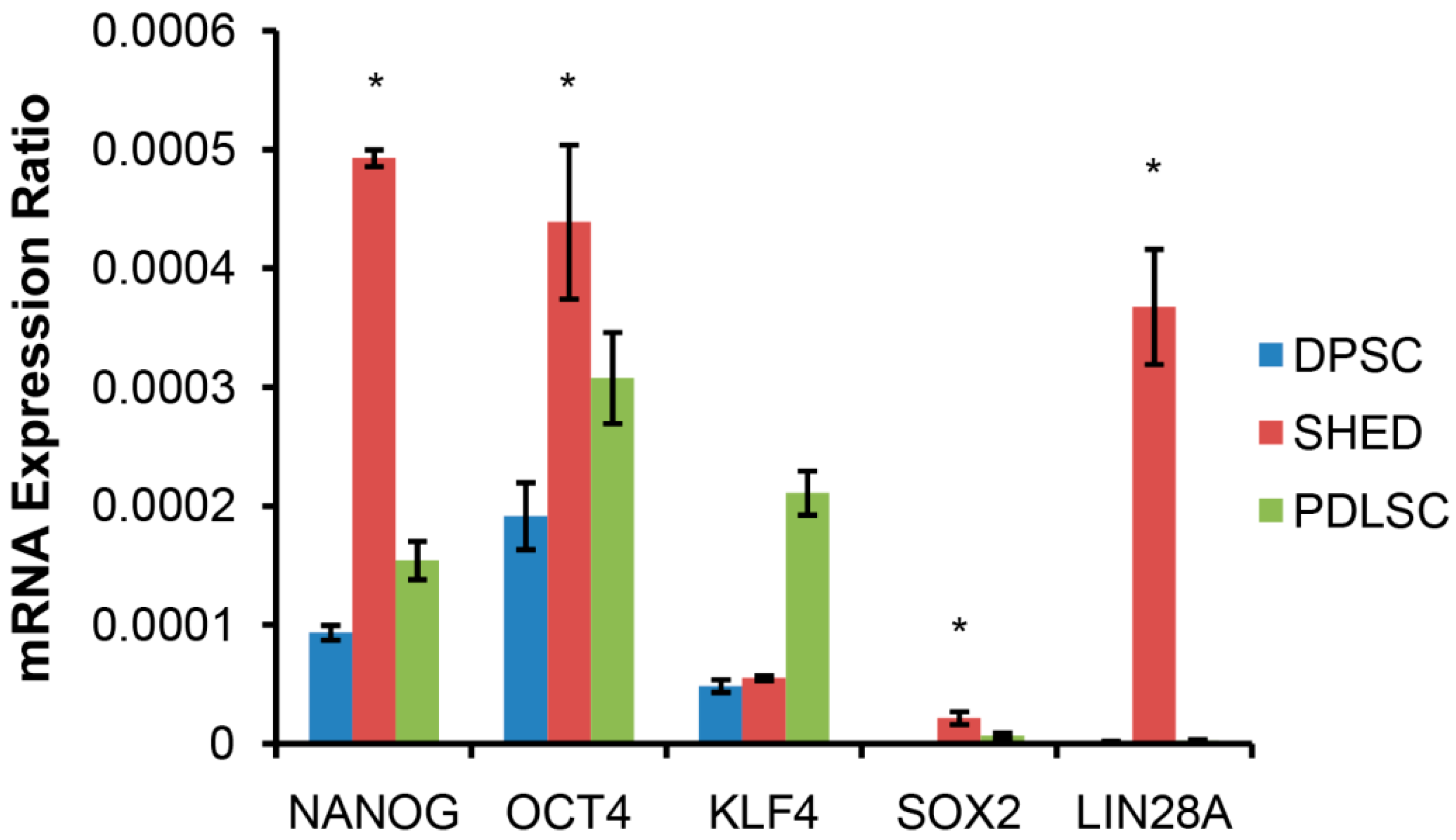
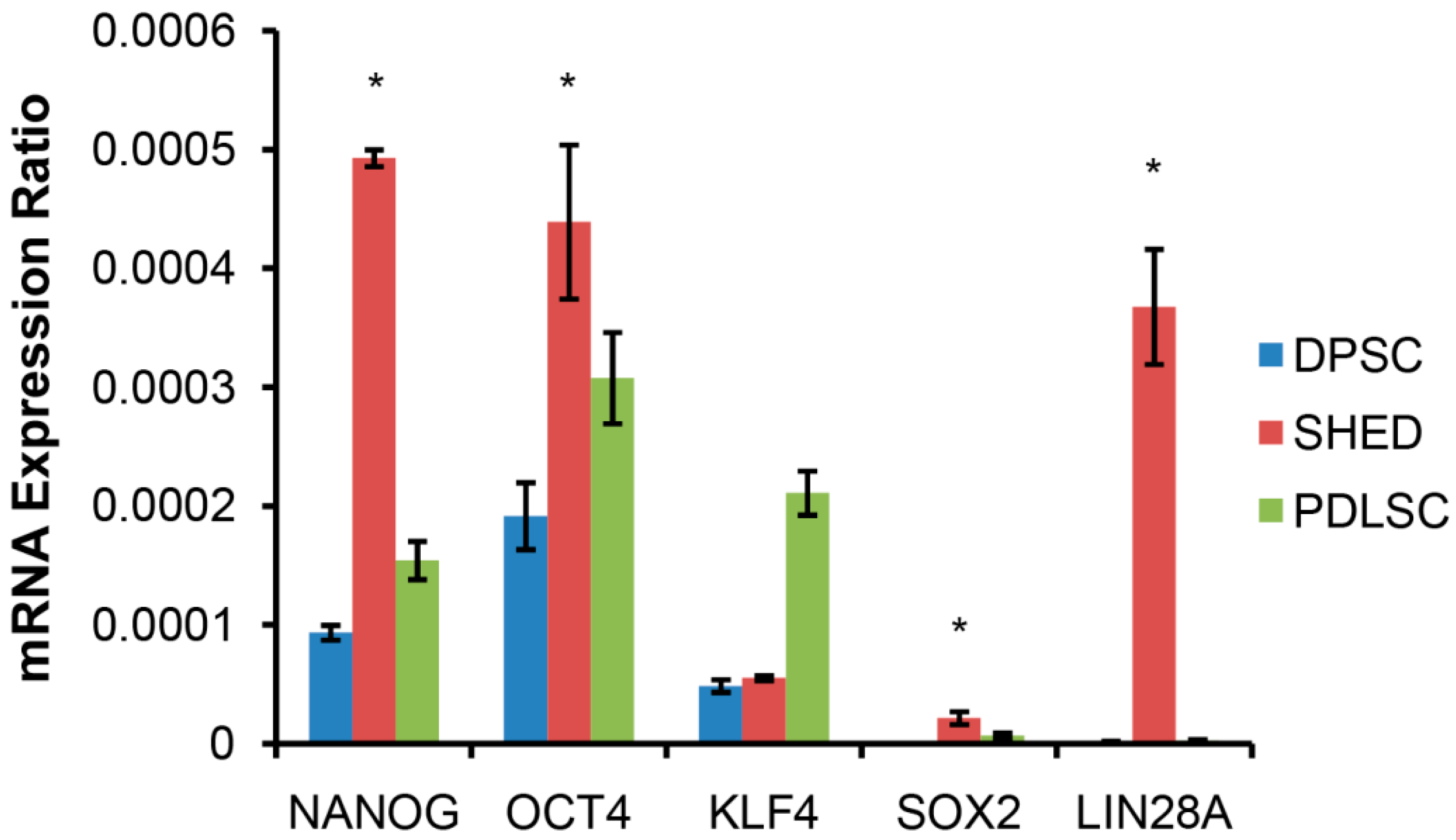
2.1. SHED Express a Strong “Stemness” Compared to Dental Pulp and Periodontal Ligament Originated Stem Cells (DPSC and PDLSC)
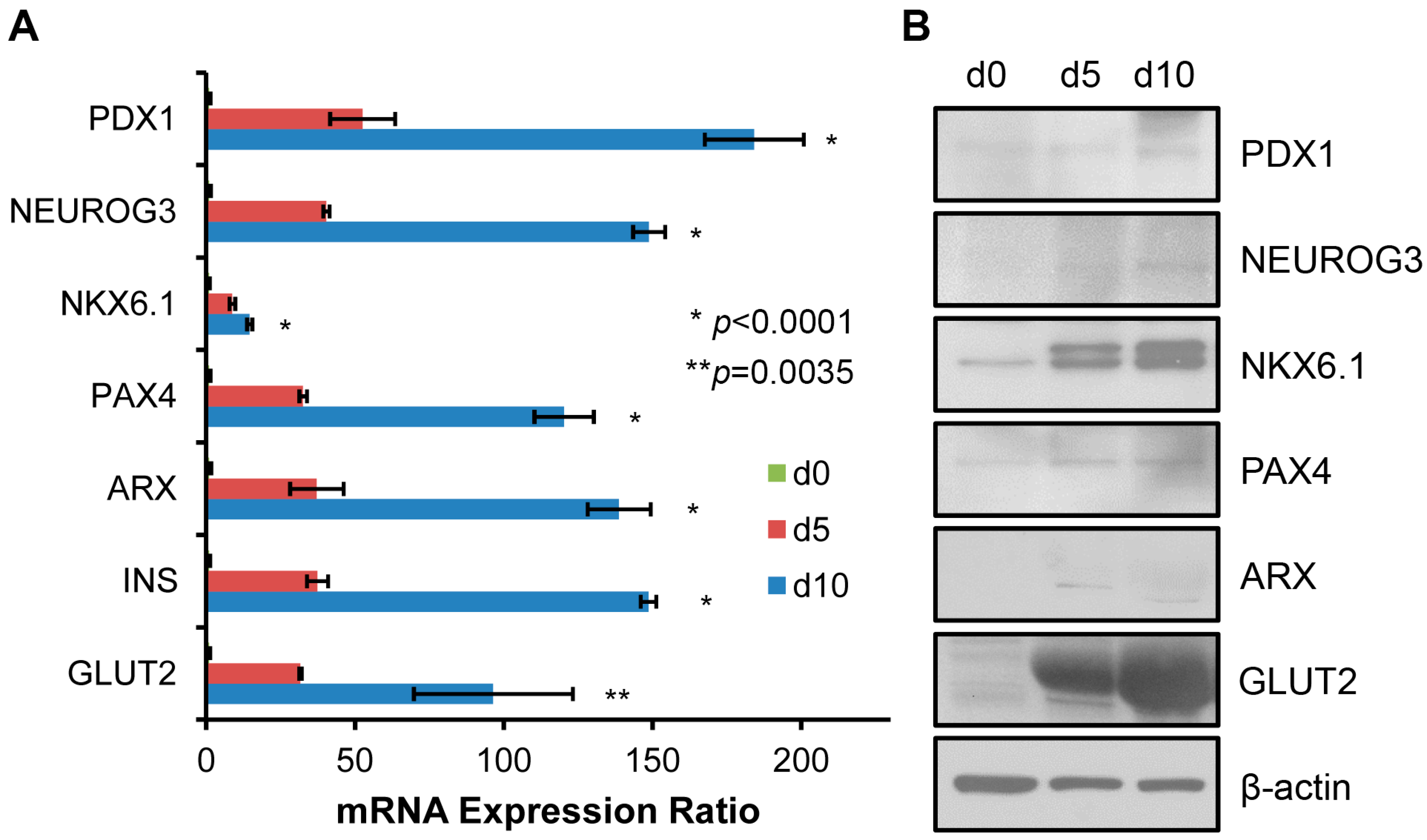
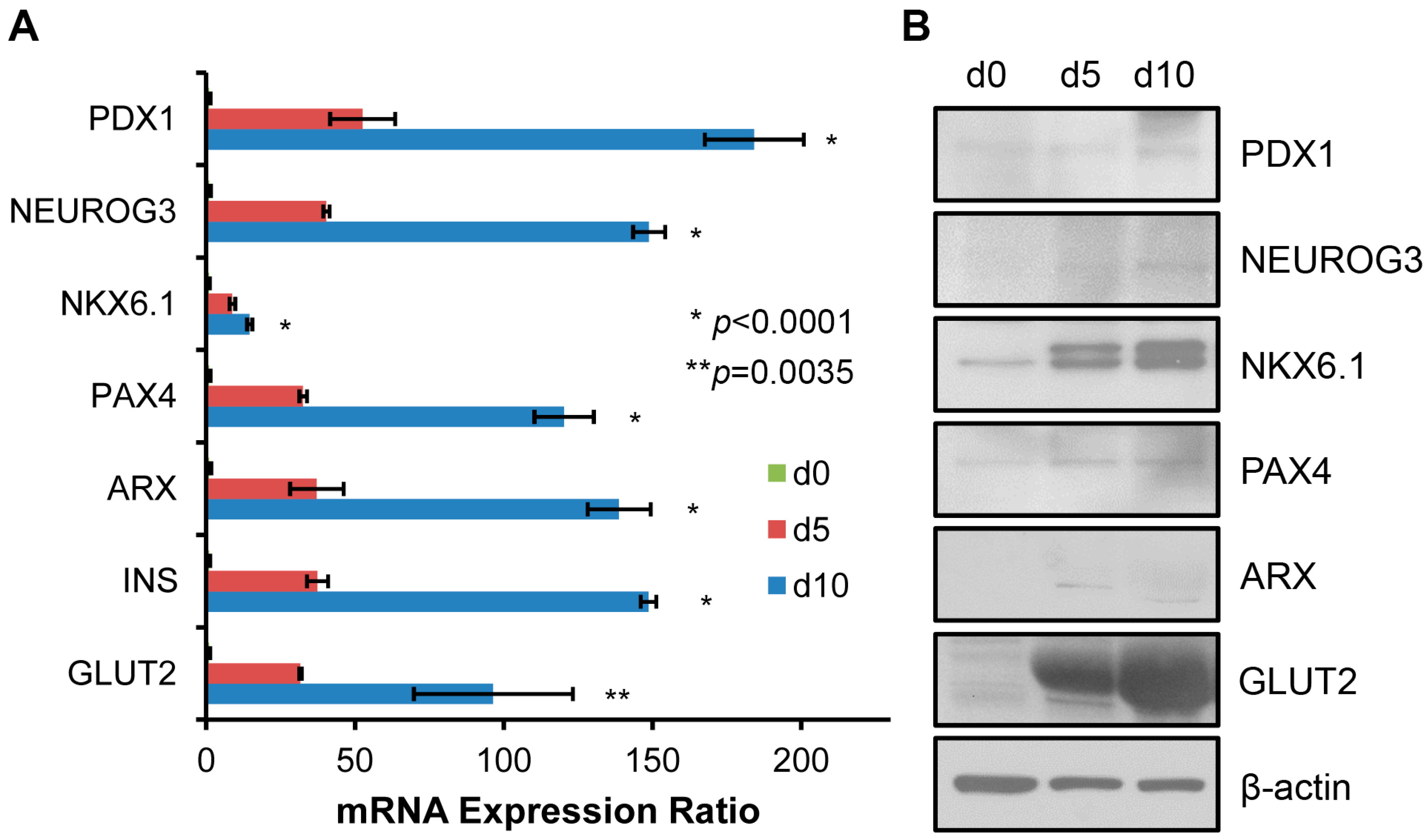
2.2. Genetic Markers in Pancreatic β Cell Lineage Expressed in SHED Indicate a Successful Conversion of SHED to SHED-β Cells
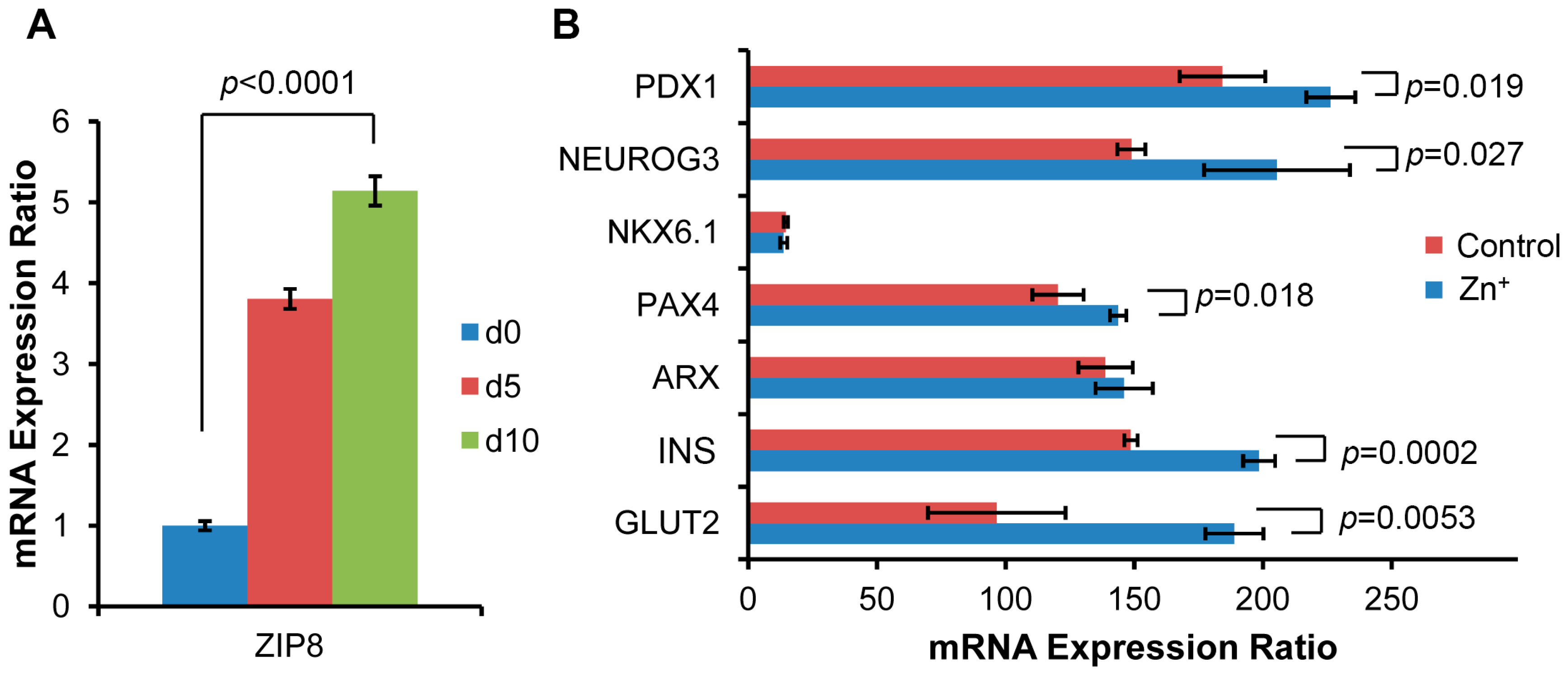
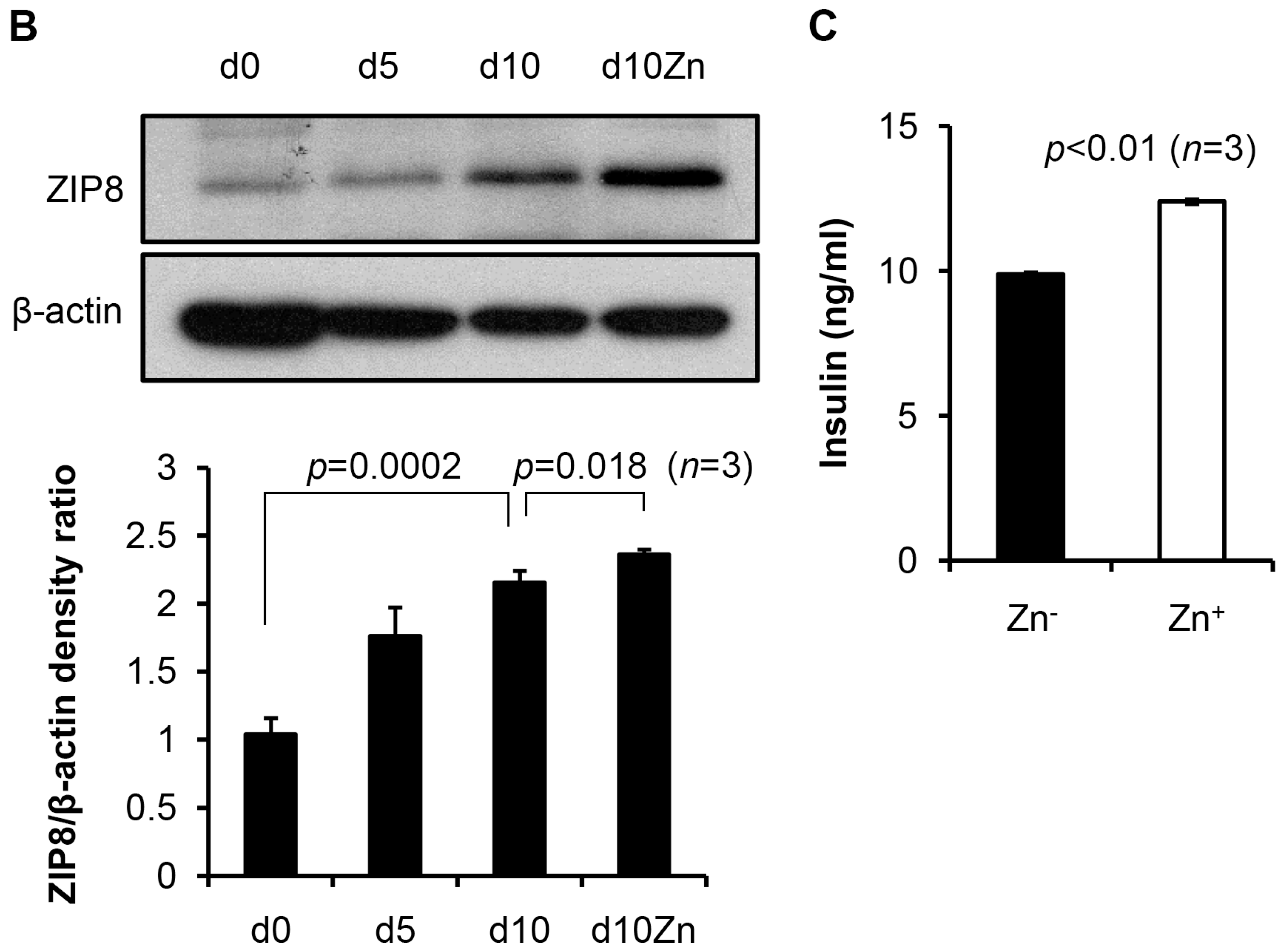
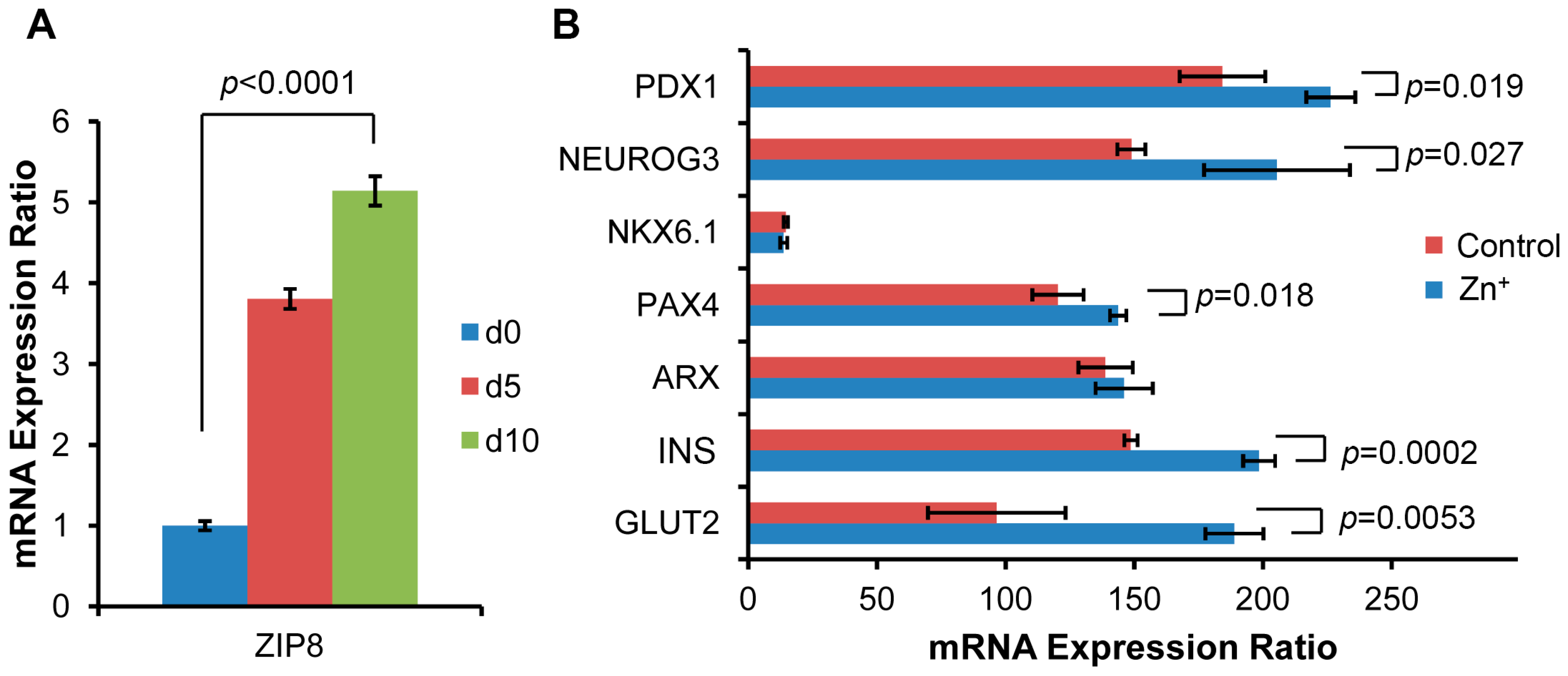
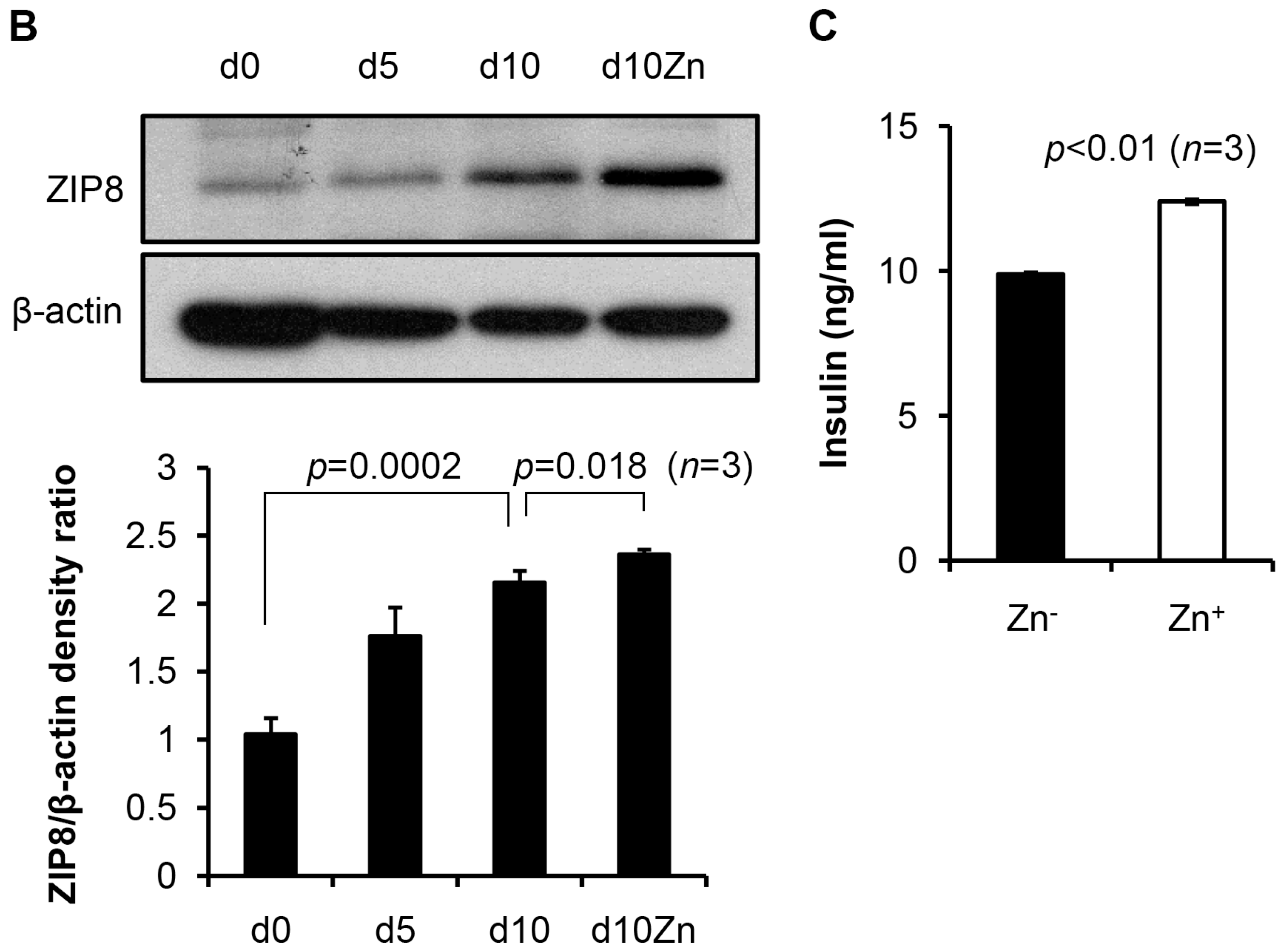
2.3. ZIP8 Expression and Zinc Supplementation Effects on the Markers in β Cell Lineage
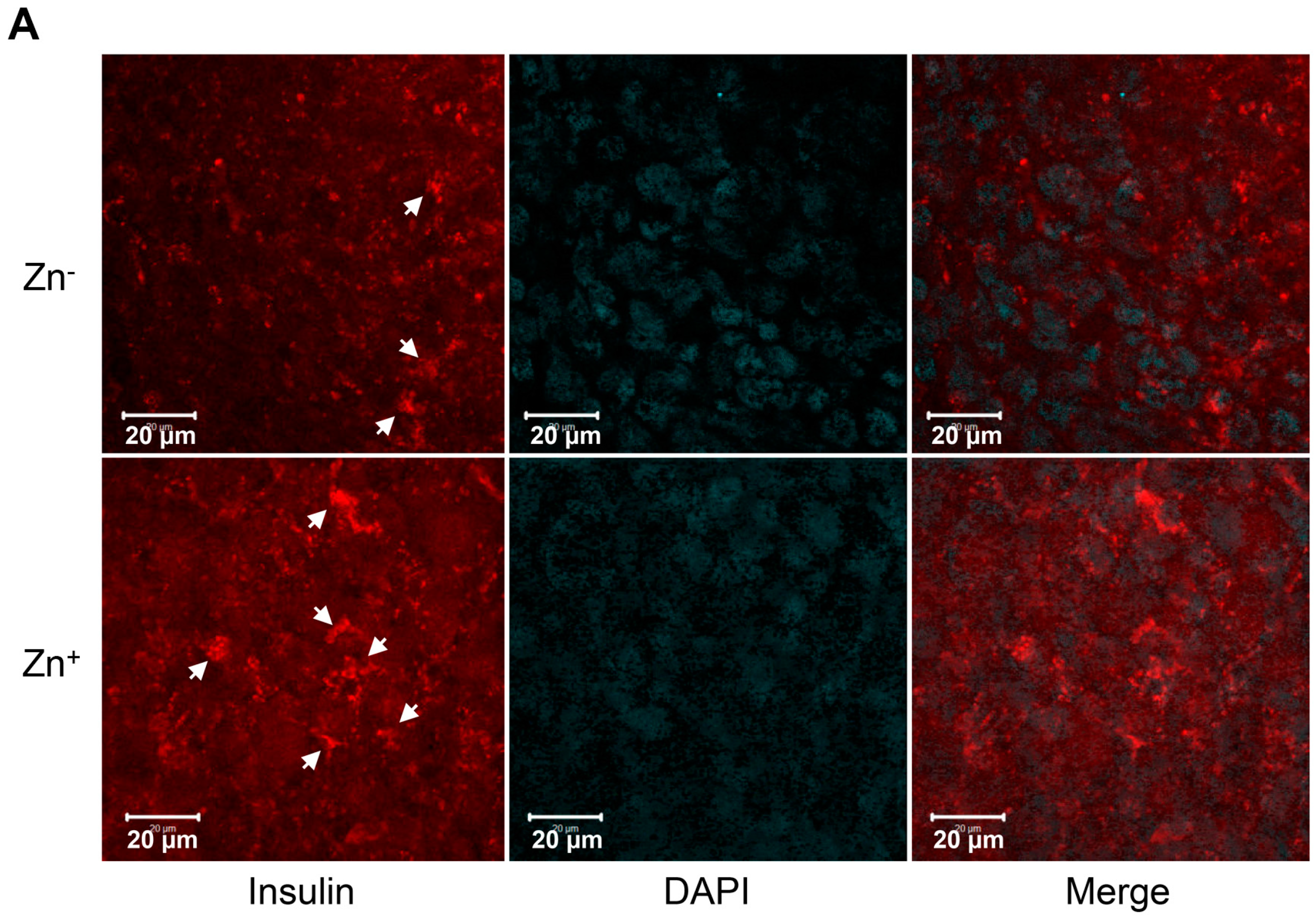
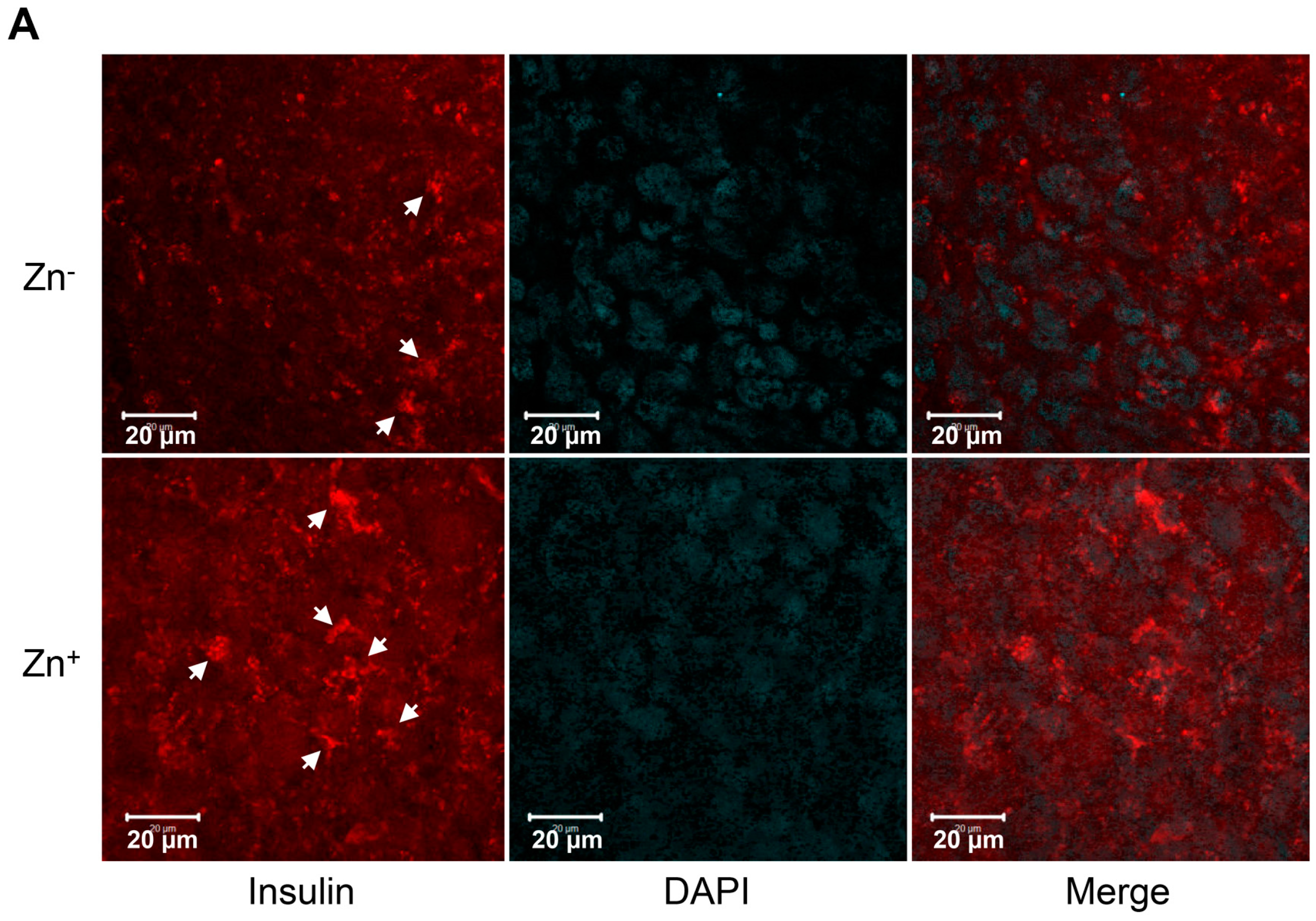
2.4. Levels of Zinc in Conjunction with ZIP8 in the Cytosol of SHED-β Cells Modulate Insulin Secretion
3. Discussion
4. Methods
4.1. Cell Cultures
4.2. In Vitro SHED-β Cell Differentiation
4.3. Real-Time qPCR
- NANOG, F: AGATGCCTCACACGGAGACT; R: TCTCTGCAGAAGTGGGTTGTT.
- OCT4, F: GAAAACCCACACTGCAGATCA; R: CGGTTACAGAACCACACTCG.
- KLF4, F: GGGAGAAGACACTGCGTCA; R: GGAAGCACTGGGGGAAGT.
- SOX2, F: TCTCATGATGTTCAACCATTCAC; R: CACATTTACATTCAAAGCACCAG.
- LIN28A, F: GAAGCGCAGATCAAAAGGAG; R: GCTGATGCTCTGGCAGAAGT.
- GAPDH, F: GGTGTGAACCATGAGAAGTATGA; R: GAGTCCTTCCACGATACCAAAG.
- PDX1, F: GGGTGACCACTAAACCAAAGA; R: GGTCATACTGGCTCGTGAATAG.
- NEUROG3, F: GCTGCTCATCGCTCTCTATTC; R: GGCAGGTCACTTCGTCTTC.
- NKX6.1, F: GAAGAGGACGACGACTACAATAAG; R: CTGCTGGACTTGTGCTTCT.
- PAX4, F: TGGGAAGGAGATGGCATAGA; R: ATCACAGGAAGGAGGAAGGA.
- ARX, F: GGCAAGGAGGTGTGCTAAA; R: GCTGGTCCTCTGTTTCCATT.
- INS, F: CTGGAGAACTACTGCAACTAGAC; R: TGCTGGTTCAAGGGCTTTAT.
- GLUT2, F: CCGCTGAGAAGATTAGACTTGG; R: GACTAGCTCCTGCCTGTTTATT.
- SLC39A8, F: GCTGGCTATTGGGACTCTTT; R: GCAACTGCCTTCTCAACATAAC.
- ACTB, F: GGATCAGCAAGCAGGAGTATG; R: AGAAAGGGTGTAACGCAACTAA.
4.4. Western-Blots
4.5. Immunofluorescence Assay
4.6. ELISA Assay for Insulin
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Vogel, G. Biomedicine. Stem cell recipe offers diabetes hope. Science 2014. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, M.A. The pathogenesis and natural history of type 1 diabetes. Cold Spring Harb. Perspect. Med. 2012, 2, a007641. [Google Scholar] [CrossRef] [PubMed]
- Kanafi, M.M.; Mamidi, M.K.; Sureshbabu, S.K.; Shahani, P.; Bhawna, C.; Warrier, S.R.; Bhonde, R. Generation of islet-like cell aggregates from human non-pancreatic cancer cell lines. Biotechnol. Lett. 2015, 37, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, G.; Riccio, M.; Pisciotta, A.; Beretti, F.; Maraldi, T.; Zavatti, M.; Cavallini, G.M.; la Sala, G.B.; Ferrari, A.; de Pol, A. In vitro differentiation into insulin-producing β-cells of stem cells isolated from human amniotic fluid and dental pulp. Dig. Liver Dis. 2013, 45, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Govindasamy, V.; Ronald, V.S.; Abdullah, A.N.; Nathan, K.R.; Ab Aziz, Z.A.; Abdullah, M.; Musa, S.; Kasim, N.H.; Bhonde, R.R. Differentiation of dental pulp stem cells into islet-like aggregates. J. Dent. Res. 2011, 90, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sha, X.J.; Li, G.H.; Yang, F.S.; Ji, K.; Wen, L.Y.; Liu, S.Y.; Chen, L.; Ding, Y.; Xuan, K. Comparative characterization of stem cells from human exfoliated deciduous teeth and dental pulp stem cells. Arch. Oral Biol. 2012, 57, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Bhonde, R.R.; Sheshadri, P.; Sharma, S.; Kumar, A. Making surrogate β-cells from mesenchymal stromal cells: Perspectives and future endeavors. Int. J. Biochem. Cell Biol. 2014, 46, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Pae, E.K.; Ahuja, B.; Kim, M.; Kim, G. Impaired glucose homeostasis after a transient intermittent hypoxic exposure in neonatal rats. Biochem. Biophys. Res. Commun. 2013, 441, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, M.K.; Helminen, O.; Matomäki, J.; Aspholm, S.; Mykkänen, J.; Mäkinen, M.; Simell, V.; Vähä-Mäkilä, M.; Simell, T.; Ilonen, J.; et al. Reduced β-cell function in early preclinical type 1 diabetes. Eur. J. Endocrinol. 2016, 174, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Pae, E.K.; Kim, G. Insulin production hampered by intermittent hypoxia via impaired zinc homeostasis. PLoS ONE 2014, 9, e90192. [Google Scholar] [CrossRef] [PubMed]
- King, J.C. Zinc: An essential but elusive nutrient. Am. J. Clin. Nutr. 2011, 94, 679S–684S. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Auld, D.S. Zinc coordination, function, and structure of zinc enzymes and other proteins. Biochemistry 1990, 29, 5647–5659. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Elnabawi, O.; Shin, D.; Pae, E.K. Transient Intermittent Hypoxia Exposure Disrupts Neonatal Bone Strength. Front. Pediatr. 2016, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Doménech, E.; Díaz-Gómez, N.M.; Barroso, F.; Cortabarria, C. Zinc and perinatal growth. Early Hum. Dev. 2001, 65, S111–S117. [Google Scholar] [CrossRef]
- Iitsuka, N.; Hie, M.; Tsukamoto, I. Zinc supplementation inhibits the increase in osteoclastogenesis and decrease in osteoblastogenesis in streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 2013, 714, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Yang, M.; Guo, B.; Cao, J.; Yang, L.; Guo, X. Zinc upregulates the expression of osteoprotegerin in mouse osteoblasts MC3T3-E1 through PKC/MAPK pathways. Biol. Trace Elem. Res. 2012, 146, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Barbieri, D.; Davison, N.; Yan, Y.; de Bruijn, J.D.; Yuan, H. Zinc in calcium phosphate mediates bone induction: In vitro and in vivo model. Acta Biomater. 2014, 10, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M.; Lönnerdal, B. Role of zinc in cellular zinc trafficking and mineralization in a murine osteoblast-like cell line. J. Nutr. Biochem. 2011, 22, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Marumo, K. Bone quality in diabetes. Front. Endocrinol. 2013, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.J.; Rakic, V.; Davis, W.A.; Chubb, S.A.; Kamber, N.; Prince, R.L.; Davis, T.M. Prevalence and predictors of osteopenia and osteoporosis in adults with Type 1 diabetes. Diabet. Med. 2009, 26, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.M.; Lee, E.H. Transcriptional regulatory cascades in Runx2-dependent bone development. Tissue Eng. B Rev. 2013, 19, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Hojyo, S.; Furuichi, T. Zinc signal: A new player in osteobiology. J. Bone Miner. Metab. 2013, 31, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, E.H.; Lomeda, R.A.; Feldmann, J.; Nixon, G.F.; Beattie, J.H.; Kwun, I.S. Zinc deprivation inhibits extracellular matrix calcification through decreased synthesis of matrix proteins in osteoblasts. Mol. Nutr. Food Res. 2011, 55, 1552–1560. [Google Scholar] [CrossRef] [PubMed]
- Lowe, N.M.; Fekete, K.; Decsi, T. Methods of assessment of zinc status in humans: A systematic review. Am. J. Clin. Nutr. 2009, 89, 2040S–2051S. [Google Scholar] [CrossRef] [PubMed]
- Dodson, G.; Steiner, D. The role of assembly in insulin's biosynthesis. Curr. Opin. Struct. Biol. 1998, 8, 189–194. [Google Scholar] [CrossRef]
- Chausmer, A.B. Zinc, insulin and diabetes. J. Am. Coll. Nutr. 1998, 17, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.; Mukai, E.; Inagaki, N. Role of endogenous ROS production in impaired metabolism-secretion coupling of diabetic pancreatic β cells. Prog. Biophys. Mol. Biol. 2011, 107, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Hardy, A.B.; Prentice, K.J.; Froese, S.; Liu, Y.; Andrews, G.K.; Wheeler, M.B. Zip4 mediated zinc influx stimulates insulin secretion in pancreatic beta cells. PLoS ONE 2015, 10, e0119136. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Batchuluun, B.; Ho, L.; Zhu, D.; Prentice, K.J.; Bhattacharjee, A.; Zhang, M.; Pourasgari, F.; Hardy, A.B.; Taylor, K.M.; et al. Characterization of Zinc Influx Transporters (ZIPs) in Pancreatic β Cells: Roles in regulating cytosolic zinc homeostasis and insulin secretion. J. Biol. Chem. 2015, 290, 18757–18769. [Google Scholar] [CrossRef] [PubMed]
- Potter, K.J.; Westwell-Roper, C.Y.; Klimek-Abercrombie, A.M.; Warnock, G.L.; Verchere, C.B. Death and dysfunction of transplanted β-cells: Lessons learned from type 2 diabetes? Diabetes 2014, 63, 12–19. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.; Shin, K.-H.; Pae, E.-K. Zinc Up-Regulates Insulin Secretion from β Cell-Like Cells Derived from Stem Cells from Human Exfoliated Deciduous Tooth (SHED). Int. J. Mol. Sci. 2016, 17, 2092. https://doi.org/10.3390/ijms17122092
Kim G, Shin K-H, Pae E-K. Zinc Up-Regulates Insulin Secretion from β Cell-Like Cells Derived from Stem Cells from Human Exfoliated Deciduous Tooth (SHED). International Journal of Molecular Sciences. 2016; 17(12):2092. https://doi.org/10.3390/ijms17122092
Chicago/Turabian StyleKim, Gyuyoup, Ki-Hyuk Shin, and Eung-Kwon Pae. 2016. "Zinc Up-Regulates Insulin Secretion from β Cell-Like Cells Derived from Stem Cells from Human Exfoliated Deciduous Tooth (SHED)" International Journal of Molecular Sciences 17, no. 12: 2092. https://doi.org/10.3390/ijms17122092
APA StyleKim, G., Shin, K.-H., & Pae, E.-K. (2016). Zinc Up-Regulates Insulin Secretion from β Cell-Like Cells Derived from Stem Cells from Human Exfoliated Deciduous Tooth (SHED). International Journal of Molecular Sciences, 17(12), 2092. https://doi.org/10.3390/ijms17122092