
Tocopherols and Tocotrienols in Common and Emerging Dietary Sources: Occurrence, Applications, and Health Benefits



Abstract
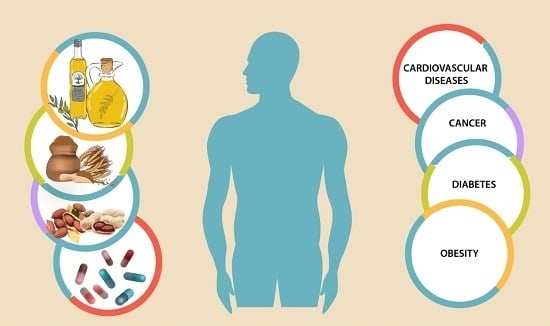
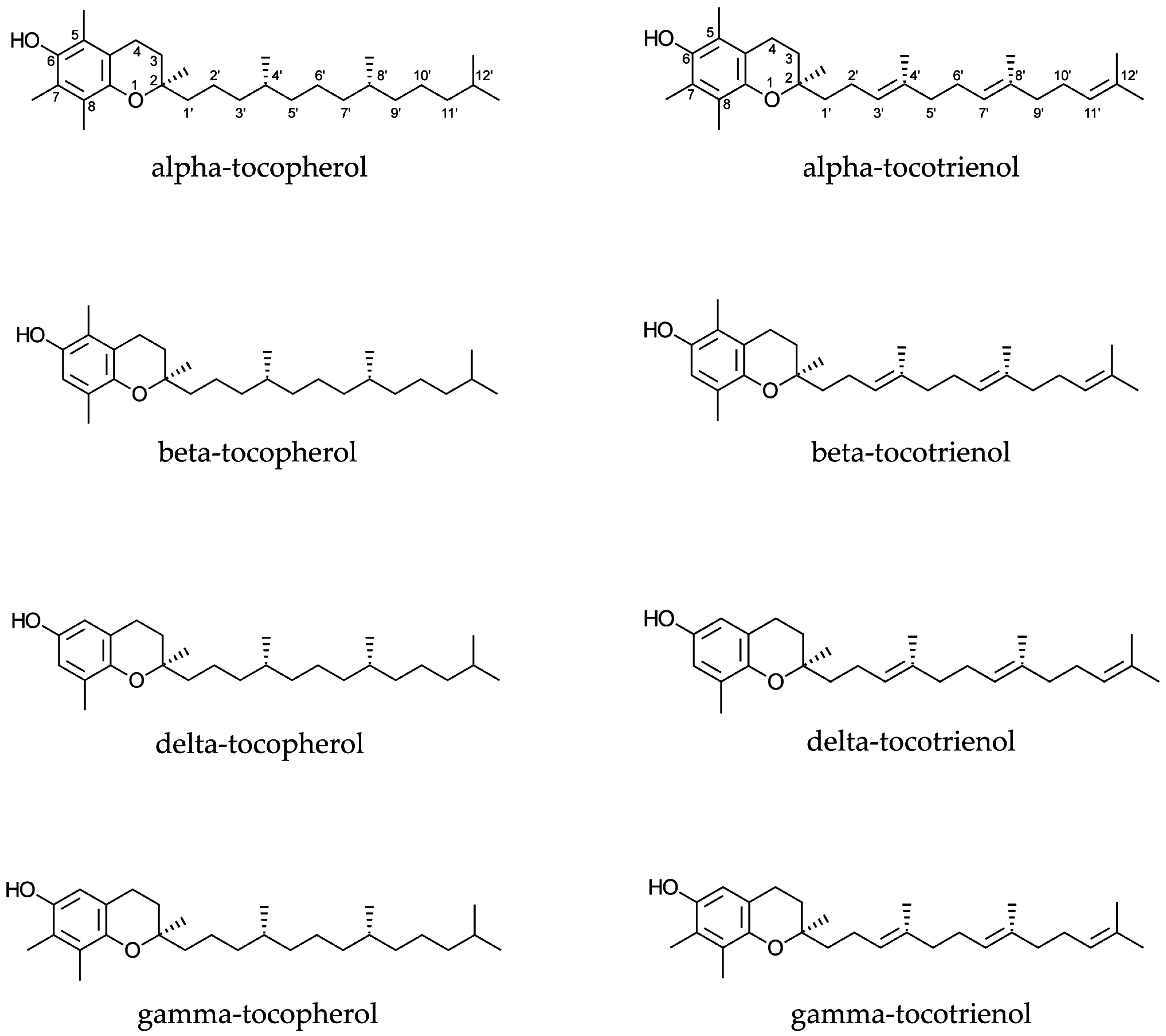
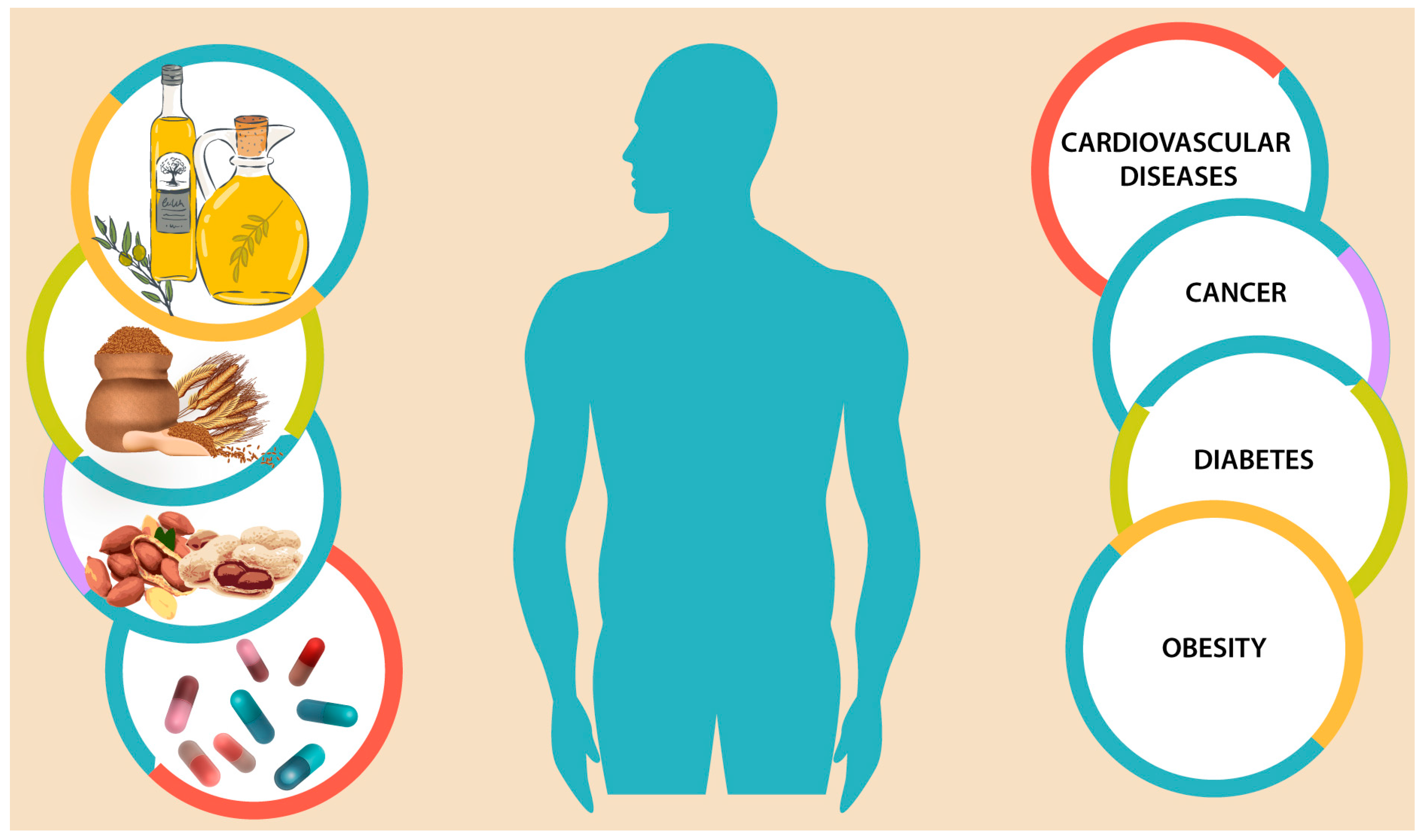
:
1. Introduction
2. Edible Oils as Sources of Tocopherols and Tocotrienols
2.1. Common Edible Oils
2.2. Specialty and Underutilized Edible Oils
3. Impact of New Cultivars on the Content of Tocopherols and Tocotrienols
4. Current and Potential Applications of Tocopherols and Tocotrienols
5. Stability and Bioavailability of Tocopherol and Tocotrienols
6. Tocopherols and Tocotrienols in Health Promotion
6.1. Tocopherols and Tocotrienols in the Prevention of Cardiovascular Diseases
6.2. Tocopherols and Tocotrienols as Adjuvants in Cancer Treatment and Prevention
6.3. Diabetes
6.4. Obesity
6.5. The Role of Tocopherols and Tocotrienols against Endogenous Formation of Trans Fatty Acids
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Zingg, J.M. Modulation of signal transduction by vitamin E. Mol. Asp. Med. 2007, 28, 481–506. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Shukla, V.K.S. Nontriacylglycerol constituents of fats, oils. Inf. Int. News Fats Oils Relat. Mater. 1996, 7, 1227–1232. [Google Scholar]
- Shin, E.-C.; Huang, Y.-Z.; Pegg, R.B.; Phillips, R.D.; Eitenmiller, R.R. Commercial Runner peanut cultivars in the United States: Tocopherol composition. J. Agric. Food Chem. 2009, 57, 10289–10295. [Google Scholar] [CrossRef] [PubMed]
- De Camargo, A.C.; Vieira, T.M.F.S.; Regitano-d’Arce, M.A.B.; de Alencar, S.M.; Calori-Domingues, M.A.; Canniatti-Brazaca, S.G. Gamma radiation induced oxidation and tocopherols decrease in in-shell, peeled and blanched peanuts. Int. J. Mol. Sci. 2012, 13, 2827–2845. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Alamprese, C.; Ratti, S. Tocopherols and tocotrienols as free radical-scavengers in refined vegetable oils and their stability during deep-fat frying. Food Chem. 2007, 102, 812–817. [Google Scholar] [CrossRef]
- Kornsteiner, M.; Wagner, K.H.; Elmadfa, I. Tocopherols and total phenolics in 10 different nut types. Food Chem. 2006, 98, 381–387. [Google Scholar] [CrossRef]
- Caporaso, N.; Savarese, M.; Paduano, A.; Guidone, G.; de Marco, E.; Sacchi, R. Nutritional quality assessment of extra virgin olive oil from the Italian retail market: Do natural antioxidants satisfy EFSA health claims? J. Food Compos. Anal. 2015, 40, 154–162. [Google Scholar] [CrossRef]
- Karmowski, J.; Hintze, V.; Kschonsek, J.; Killenberg, M.; Böhm, V. Antioxidant activities of tocopherols/tocotrienols and lipophilic antioxidant capacity of wheat, vegetable oils, milk and milk cream by using photochemiluminescence. Food Chem. 2015, 175, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Grilo, E.C.; Costa, P.N.; Gurgel, C.S.S.; Beserra, A.F.L.; Almeida, F.N.S.; Dimenstein, R. Alpha-tocopherol and gamma-tocopherol concentration in vegetable oils. Food Sci. Technol. (Campinas) 2014, 34, 379–385. [Google Scholar] [CrossRef]
- Schwartz, H.; Ollilainen, V.; Piironen, V.; Lampi, A.-M. Tocopherol, tocotrienol and plant sterol contents of vegetable oils and industrial fats. J. Food Compos. Anal. 2008, 21, 152–161. [Google Scholar] [CrossRef]
- Chun, J.; Lee, J.; Ye, L.; Exler, J.; Eitenmiller, R.R. Tocopherol and tocotrienol contents of raw and processed fruits and vegetables in the United States diet. J. Food Compos. Anal. 2006, 19, 196–204. [Google Scholar] [CrossRef]
- De Camargo, A.C.; Regitano-d’Arce, M.A.B.; Gallo, C.R.; Shahidi, F. Gamma-irradiation induced changes in microbiological status, phenolic profile and antioxidant activity of peanut skin. J. Funct. Foods 2015, 12, 129–143. [Google Scholar] [CrossRef]
- Alshikh, N.; de Camargo, A.C.; Shahidi, F. Phenolics of selected lentil cultivars: Antioxidant activities and inhibition of low-density lipoprotein and DNA damage. J. Funct. Foods 2015, 18, 1022–1038. [Google Scholar] [CrossRef]
- Ayoub, M.; de Camargo, A.C.; Shahidi, F. Antioxidants and bioactivities of free, esterified and insoluble-bound phenolics from berry seed meals. Food Chem. 2016, 197, 221–232. [Google Scholar] [CrossRef] [PubMed]
- De Camargo, A.C.; Regitano-d’Arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Low molecular weight phenolics of grape juice and winemaking byproducts: Antioxidant activities and inhibition of oxidation of human low-density lipoprotein cholesterol and DNA strand breakage. J. Agric. Food Chem. 2014, 62, 12159–12171. [Google Scholar] [CrossRef] [PubMed]
- Pryor, W.A. Vitamin E and heart disease: Basic science to clinical intervention trials. Free Radic. Biol. Med. 2000, 28, 141–164. [Google Scholar] [CrossRef]
- Pryor, W.A. The antioxidant nutrients and disease prevention—What do we know and what do we need to find out? Am. J. Clin. Nutr. 1991, 53, 391S–393S. [Google Scholar] [PubMed]
- Burton, G.W.; Traber, M.G.; Acuff, R.V.; Walters, D.N.; Kayden, H.; Hughes, L.; Ingold, K.U. Human plasma and tissue alpha-tocopherol concentrations in response to supplementation with deuterated natural and synthetic vitamin E. Am. J. Clin. Nutr. 1998, 67, 669–684. [Google Scholar] [PubMed]
- Carter, C.; Finley, W.; Fry, J.; Jackson, D.; Willis, L. Palm oil markets and future supply. Eur. J. Lipid Sci. Technol. 2007, 109, 307–314. [Google Scholar] [CrossRef]
- Syväoja, E.-L.; Pilronen, V.; Varo, P.; Koivistoinen, P.; Salminen, K. Tocopherols and tocotrienols in finnish foods: Oils and fats. J. Am. Oil Chem. Soc. 1986, 63, 328–329. [Google Scholar] [CrossRef]
- Velasco, L.; Perez-Vich, B.; Fernandez-Martinez, J.M. Novel variation for the tocopherol profile in a sunflower created by mutagenesis and recombination. Plant Breed. 2004, 123, 490–492. [Google Scholar] [CrossRef]
- Moreau, R.A.; Flores, R.A.; Hicks, K.B. Composition of functional lipids in hulled and hulless barley in fractions obtained by scarification and in barley oil. Cereal Chem. 2007, 84, 1–5. [Google Scholar] [CrossRef]
- Wang, L.; Newman, R.K.; Newman, C.W.; Jackson, L.L.; Hofer, P.J. Tocotrienol and fatty acid composition of barley oil and their effects on lipid metabolism. Plant Foods Hum. Nutr. 1993, 43, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Desai, I.D.; Bhagavan, H.; Salkeld, R.; Dutra de Oliveira, J.E. Vitamin E content of crude and refined vegetable oils in southern Brazil. J. Food Compos. Anal. 1988, 1, 231–238. [Google Scholar] [CrossRef]
- Carpenter, A.P. Determination of tocopherols in vegetable oils. J. Am. Oil Chem. Soc. 1979, 56, 668–671. [Google Scholar] [CrossRef]
- Corsini, M.S.; Silva, M.G.; Jorge, N. Loss in tocopherols and oxidative stability during the frying of frozen cassava chips. Grasas Aceites 2009, 60, 77–81. [Google Scholar] [CrossRef]
- Ng, M.H.; Choo, Y.M.; Ma, A.N.; Chuah, C.H.; Hashim, M.A. Separation of vitamin E (tocopherol, tocotrienol, and tocomonoenol) in palm oil. Lipids 2004, 39, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Goufo, P.; Trindade, H. Rice antioxidants: Phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid. Food Sci. Nutr. 2014, 2, 75–104. [Google Scholar] [CrossRef] [PubMed]
- Aued-Pimentel, S.; Takemoto, E.; Antoniassi, R.; Badolato, E.S.G. Composition of tocopherols in sesame seed oil: An indicative of adulteration. Grasas Aceites 2006, 57, 205–210. [Google Scholar] [CrossRef]
- Warner, K.; Mounts, T.L. Analysis of tocopherols and phytosterols in vegetable-oils by HPLC with evaporative light-scattering detection. J. Am. Oil Chem. Soc. 1990, 67, 827–831. [Google Scholar] [CrossRef]
- Sofi, F.; Macchi, C.; Abbate, R.; Gensini, G.F.; Casini, A. Mediterranean diet and health. Biofactors 2013, 39, 335–342. [Google Scholar] [CrossRef] [PubMed]
- De Camargo, A.C.; Canniatti-Brazaca, S.G. Peanuts as a source of protein, unsaturated fatty acids, tocopherol and polyphenols. In Peanuts: Production, Nutritional Content and Health Implications; Cook, R.W., Ed.; Nova Science Publishers: New York, NY, USA, 2014; pp. 69–80. [Google Scholar]
- Da Silva, A.C.; Jorge, N. Bioactive compounds of the lipid fractions of agro-industrial waste. Food Res. Int. 2014, 66, 493–500. [Google Scholar] [CrossRef]
- Malacrida, C.R.; Jorge, N. Yellow passion fruit seed oil (Passiflora edulis F. flavicarpa): Physical and chemical characteristics. Braz. Arch. Biol. Technol. 2012, 55, 127–134. [Google Scholar] [CrossRef]
- Malacrida, C.R.; Kimura, M.; Jorge, N. Characterization of a high oleic oil extracted from papaya (Carica papaya L.) seeds. Ciencia Tecnol. Aliment. 2011, 31, 929–934. [Google Scholar] [CrossRef]
- Malacrida, C.R.; Kimura, M.; Jorge, N. Phytochemicals and antioxidant activity of citrus seed oils. Food Sci. Technol. Res. 2012, 18, 399–404. [Google Scholar] [CrossRef]
- Seppanen, C.M.; Song, Q.; Csallany, A.S. The antioxidant functions of tocopherol and tocotrienol homologues in oils, fats, and food systems. J. Am. Oil Chem. Soc. 2010, 87, 469–481. [Google Scholar] [CrossRef]
- Li, Q.; Wang, J.; Shahidi, F. Chemical characteristics of cold-pressed blackberry, black raspberry, and blueberry seed oils and the role of the minor components in their oxidative stability. J. Agric. Food Chem. 2016, 64, 5410–5416. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, M.C.; Jorge, N. Fatty acids and bioactive compounds of the pulps and kernels of Brazilian palm species, guariroba (Syagrus oleraces), jerivá (Syagrus romanzoffiana) and macaúba (Acrocomia aculeata). J. Sci. Food Agric. 2012, 92, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.S.; Luzia, D.M.M.; Jorge, N. Physicochemical and bioactive properties of Hymenaea courbaril L. pulp and seed lipid fraction. Ind. Crops Prod. 2013, 49, 610–618. [Google Scholar] [CrossRef]
- Luzia, D.M.M.; Jorge, N. Bioactive substance contents and antioxidant capacity of the lipid fraction of Annona crassiflora Mart. seeds. Ind. Crops Prod. 2013, 42, 231–235. [Google Scholar] [CrossRef]
- Luzia, D.M.M.; Jorge, N. Antioxidant activity, fatty acid profile and tocopherols of Tamarindus indica L. Seeds. Ciencia Tecnol. Aliment. 2011, 31, 497–501. [Google Scholar] [CrossRef]
- Inada, K.O.P.; Oliveira, A.A.; Revoredo, T.B.; Martins, A.B.N.; Lacerda, E.C.Q.; Freire, A.S.; Braz, B.F.; Santelli, R.E.; Torres, A.G.; Perrone, D.; et al. Screening of the chemical composition and occurring antioxidants in jabuticaba (Myrciaria jaboticaba) and jussara (Euterpe edulis) fruits and their fractions. J. Funct. Foods 2015, 17, 422–433. [Google Scholar] [CrossRef]
- Da Silva, P.P.M.; Casemiro, R.C.; Zillo, R.R.; de Camargo, A.C.; Prospero, E.T.P.; Spoto, M.H.F. Sensory descriptive quantitative analysis of unpasteurized and pasteurized juçara pulp (Euterpe edulis) during long-term storage. Food Sci. Nutr. 2014, 2, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Barros, L.; Dias, M.I.; Santos-Buelga, C.; Ferreira, I.C.F.R.; Asquieri, E.R.; Berrios, J.D.J. Non-fermented and fermented jabuticaba (Myrciaria cauliflora Mart.) pomaces as valuable sources of functional ingredients. Food Chem. 2016, 208, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Darnet, S.; Serra, J.L.; da Cruz Rodrigues, A.M.; Meller da Silva, L.H. A high-performance liquid chromatography method to measure tocopherols in assai pulp (Euterpe oleracea). Food Res. Int. 2011, 44, 2107–2111. [Google Scholar] [CrossRef]
- Coimbra, M.C.; Jorge, N. Characterization of the pulp and kernel oils from Syagrus oleracea, Syagrus romanzoffiana, and Acrocomia aculeata. J. Food Sci. 2011, 76, C1156–C1161. [Google Scholar] [CrossRef] [PubMed]
- Montoya, C.; Cochard, B.; Flori, A.; Cros, D.; Lopes, R.; Cuellar, T.; Espeout, S.; Syaputra, I.; Villeneuve, P.; Pina, M.; et al. Genetic architecture of palm oil fatty acid composition in cultivated oil palm (Elaeis guineensis jacq.) compared to its wild relative E. oleifera (HBK) cortes. PLoS ONE 2014, 9, e95412. [Google Scholar] [CrossRef] [PubMed]
- Warner, K.; Gupta, M. Potato chip quality and frying oil stability of high oleic acid soybean oil. J. Food Sci. 2005, 70, S395–S400. [Google Scholar] [CrossRef]
- Merrill, L.I.; Pike, O.A.; Ogden, L.V.; Dunn, M.L. Oxidative stability of conventional and high-oleic vegetable oils with added antioxidants. J. Am. Oil Chem. Soc. 2008, 85, 771–776. [Google Scholar] [CrossRef]
- Shin, E.-C.; Craft, B.D.; Pegg, R.B.; Phillips, R.D.; Eitenmiller, R.R. Chemometric approach to fatty acid profiles in Runner-type peanut cultivars by principal component analysis (PCA). Food Chem. 2010, 119, 1262–1270. [Google Scholar] [CrossRef]
- Wilkin, J.D.; Ashton, I.P.; Fielding, L.M.; Tatham, A.S. Storage stability of whole and nibbed, conventional and high oleic peanuts (Arachis hypogeae L.). Food Bioprocess Technol. 2014, 7, 105–113. [Google Scholar] [CrossRef]
- Janila, P.; Pandey, M.K.; Shasidhar, Y.; Variath, M.T.; Sriswathi, M.; Khera, P.; Manohar, S.S.; Nagesh, P.; Vishwakarma, M.K.; Mishra, G.P.; et al. Molecular breeding for introgression of fatty acid desaturase mutant alleles (ahFAD2A and ahFAD2B) enhances oil quality in high and low oil containing peanut genotypes. Plant Sci. 2016, 242, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Abidi, S.L.; List, G.R.; Rennick, K.A. Effect of genetic modification on the distribution of minor constituents in canola oil. J. Am. Oil Chem. Soc. 1999, 76, 463–467. [Google Scholar] [CrossRef]
- Dionisi, F.; Prodolliet, J.; Tagliaferri, E. Assessment of olive oil adulteration by reversed-phase high-performance liquid chromatography amperometric detection of tocopherols and tocotrienols. J. Am. Oil Chem. Soc. 1995, 72, 1505–1511. [Google Scholar] [CrossRef]
- Clemente, T.E.; Cahoon, E.B. Soybean oil: Genetic approaches for modification of functionality and total content. Plant Physiol. 2009, 151, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lee, Y.Y.; Choi, M.S.; Jeong, K.H.; Lee, S.K.; Seo, M.J.; Yun, H.T.; Lee, C.K.; Kim, W.H.; Lee, S.C.; et al. Antioxidant activity and inhibition of lipid peroxidation in germinating seeds of transgenic soybean expressing OsHGGT. J. Agric. Food Chem. 2011, 59, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, X.H.; Chen, P.X.; Zhang, B.; Hernandez, M.; Zhang, H.; Marcone, M.F.; Liu, R.H.; Tsao, R. Characterisation of fatty acid, carotenoid, tocopherol/tocotrienol compositions and antioxidant activities in seeds of three Chenopodium quinoa Wild genotypes. Food Chem. 2015, 174, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Nakai, S.; Suenaga, M.; Sugihara, A.; Kitano, M.; Tominaga, Y. Facile purification of tocopherols from soybean oil deodorizer distillate in high yield using lipase. J. Am. Oil Chem. Soc. 2000, 77, 1009–1013. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Lipid oxidation and improving the oxidative stability. Chem. Soc. Rev. 2010, 39, 4067–4079. [Google Scholar] [CrossRef] [PubMed]
- Pestana-Bauer, V.R.; Zambiazi, R.C.; Mendonca, C.R.B.; Beneito-Cambra, M.; Ramis-Ramos, G. Gamma-oryzanol and tocopherol contents in residues of rice bran oil refining. Food Chem. 2012, 134, 1479–1483. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.M. Oat tocols—Concentration and stability in oat products and distribution within the kernel. Cereal Chem. 1995, 72, 21–24. [Google Scholar]
- Moreau, R.A.; Wayns, K.E.; Flores, R.A.; Hicks, K.B. Tocopherols and tocotrienols in barley oil prepared from germ and other fractions from scarification and sieving of hulless barley. Cereal Chem. 2007, 84, 587–592. [Google Scholar] [CrossRef]
- Kumar, G.S.; Krishna, A.G.G. Studies on the nutraceuticals composition of wheat derived oils wheat bran oil and wheat germ oil. J. Food Sci. Technol. 2015, 52, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Akoh, C.C. Antioxidant activities of annatto and palm tocotrienol-rich fractions in fish oil and structured lipid-based infant formula emulsion. Food Chem. 2015, 168, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Shammugasamy, B.; Ramakrishnan, Y.; Manan, F.; Muhammad, K. Rapid reversed-phase chromatographic method for determination of eight vitamin E isomers and γ-oryzanols in rice bran and rice bran oil. Food Anal. Methods 2015, 8, 649–655. [Google Scholar] [CrossRef]
- Chen, B.; McClements, D.J.; Decker, E.A. Minor components in food oils: A critical review of their roles on lipid oxidation chemistry in bulk oils and emulsions. Crit. Rev. Food Sci. Nutr. 2011, 51, 901–916. [Google Scholar] [CrossRef] [PubMed]
- Nagy, K.; Kerrihard, A.L.; Beggio, M.; Craft, B.D.; Pegg, R.B. Modeling the impact of residual fat-soluble vitamin (FSV) contents on the oxidative stability of commercially refined vegetable oils. Food Res. Int. 2016, 84, 26–32. [Google Scholar] [CrossRef]
- Huang, S.W.; Frankel, E.N.; German, J.B. Antioxidant activity of α- and γ-tocopherols in bulk oils and in oil-in-water emulsions. J. Agric. Food Chem. 1994, 42, 2108–2114. [Google Scholar] [CrossRef]
- Dolde, D.; Wang, T. Oxidation of corn oils with spiked tocols. J. Am. Oil Chem. Soc. 2011, 88, 1759–1765. [Google Scholar] [CrossRef]
- Wagner, K.H.; Wotruba, F.; Elmadfa, I. Antioxidative potential of tocotrienols and tocopherols in coconut fat at different oxidation temperatures. Eur. J. Lipid Sci. Tech. 2001, 103, 746–751. [Google Scholar] [CrossRef]
- Top, A.G.M.; Ong, A.S.H.; Kato, A.; Watanabe, H.; Kawada, T. Antioxidant activities of palm vitamin E with special reference to tocotrienols. Elaeis 1989, 1, 63–67. [Google Scholar]
- Melo, P.S.; Arrivetti, L.D.O.R.; de Alencar, S.M.; Skibsted, L.H. Antioxidative and prooxidative effects in food lipids and synergism with α-tocopherol of açaí seed extracts and grape rachis extracts. Food Chem. 2016, 213, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Shahidi, F. Tocopherols and phospholipids enhance the oxidative stability of borage and evening primrose triacylglycerols. J. Food Lipids 2000, 7, 143–150. [Google Scholar] [CrossRef]
- Let, M.B.; Jacobsen, C.; Meyer, A.S. Ascorbyl palmitate, gamma-tocopherol, and edta affect lipid oxidation in fish oil enriched salad dressing differently. J. Agric. Food Chem. 2007, 55, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Akoh, C.C. Oxidative stability of structured lipid-based infant formula emulsion: Effect of antioxidants. Food Chem. 2015, 178, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, Y.; Prinyawiwatkul, W.; King, J.M.; Xu, Z. Comparison of the activities of hydrophilic anthocyanins and lipophilic tocols in black rice bran against lipid oxidation. Food Chem. 2013, 141, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.D.; Harvey, K.A.; Pavlina, T.M.; Zaloga, G.P.; Siddiqui, R.A. Tocopherol and tocotrienol homologs in parenteral lipid emulsions. Eur. J. Lipid Sci. Tech. 2015, 117, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.D.; Harvey, K.A.; Pavlina, T.M.; Zaloga, G.P.; Siddiqui, R.A. Distribution of tocopherols and tocotrienols in guinea pig tissues following parenteral lipid emulsion infusion. J. Parenter. Enter. Nutr. 2016, 40, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Decker, E.A.; Goddard, J.M. Controlling lipid oxidation of food by active packaging technologies. Food Funct. 2013, 4, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Horn, A.F.; Nielsen, N.S.; Jacobsen, C. Additions of caffeic acid, ascorbyl palmitate or γ-tocopherol to fish oil-enriched energy bars affect lipid oxidation differently. Food Chem. 2009, 112, 412–420. [Google Scholar] [CrossRef]
- Kanatt, S.R.; Paul, P.; D’Souza, S.F.; Thomas, P. Lipid peroxidation in chicken meat during chilled storage as affected by antioxidants combined with low-dose gamma irradiation. J. Food Sci. 1998, 63, 198–200. [Google Scholar] [CrossRef]
- Barbosa-Pereira, L.; Cruz, J.M.; Sendón, R.; Rodríguez Bernaldo de Quirós, A.; Ares, A.; Castro-López, M.; Abad, M.J.; Maroto, J.; Paseiro-Losada, P. Development of antioxidant active films containing tocopherols to extend the shelf life of fish. Food Control 2013, 31, 236–243. [Google Scholar] [CrossRef]
- Chen, X.; Lee, D.S.; Zhu, X.; Yam, K.L. Release kinetics of tocopherol and quercetin from binary antioxidant controlled-release packaging films. J. Agric. Food Chem. 2012, 60, 3492–3497. [Google Scholar] [CrossRef] [PubMed]
- Marcos, B.; Sárraga, C.; Castellari, M.; Kappen, F.; Schennink, G.; Arnau, J. Development of biodegradable films with antioxidant properties based on polyesters containing α-tocopherol and olive leaf extract for food packaging applications. Food Packag. Shelf Life 2014, 1, 140–150. [Google Scholar] [CrossRef]
- Tricker, A.R.; Preussmann, R. Carcinogenic N-nitrosamines in the diet: Occurrence, formation, mechanisms and carcinogenic potential. Mutat. Res./Genet. Toxicol. 1991, 259, 277–289. [Google Scholar] [CrossRef]
- Yang, H.; Meng, P.; Xiong, Y.L.; Ma, L.; Wang, C.; Zhu, Y. Oxidation in HiOx-packaged pork Longissimus muscle predisposes myofibrillar and sarcoplasmic proteins to N-nitrosamine formation in nitrite-curing solution. Meat Sci. 2013, 95, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.I.; Skrypec, D.J.; Mandagere, A.K.; Booren, A.M.; Pearson, A.M. Further factors influencing N-nitrosamine formation in bacon. IARC Sci. Publ. 1984, 57, 301–309. [Google Scholar] [PubMed]
- Kurechi, T.; Kikugawa, K.; Ozawa, M. Effect of malondialdehyde on nitrosamine formation. Food Cosmet. Toxicol. 1980, 18, 119–122. [Google Scholar] [CrossRef]
- Fiddler, W.; Pensabene, J.W.; Piotrowski, E.G.; Phillips, J.G.; Keating, J.; Mergens, W.J.; Newmark, H.L. Inhibition of formation of volatile nitrosamines in fried bacon by the use of cure-solubilized α-tocopherol. J. Agric. Food Chem. 1978, 26, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, F.; Zhuang, H.; Chen, X.; Li, L.; Qiao, W.; Zhang, J. Effects of plant polyphenols and α-tocopherol on lipid oxidation, residual nitrites, biogenic amines, and N-nitrosamines formation during ripening and storage of dry-cured bacon. LWT—Food Sci. Technol. 2015, 60, 199–206. [Google Scholar] [CrossRef]
- Pourazrang, H.; Moazzami, A.A.; Bazzaz, B.S.F. Inhibition of mutagenic N-nitroso compound formation in sausage samples by using L-ascorbic acid and α-tocopherol. Meat Sci. 2002, 62, 479–483. [Google Scholar] [CrossRef]
- Pegg, R.B.; Shahidi, F. Unraveling the chemical identity of meat pigments. Crit. Rev. Food Sci. Nutr. 1997, 37, 561–589. [Google Scholar] [CrossRef] [PubMed]
- Djenane, D.; Sánchez-Escalante, A.; Beltrán, J.A.; Roncalés, P. Ability of α-tocopherol, taurine and rosemary, in combination with vitamin C, to increase the oxidative stability of beef steaks packaged in modified atmosphere. Food Chem. 2002, 76, 407–415. [Google Scholar] [CrossRef]
- Zhong, Y.; Lall, S.P.; Shahidi, F. Effects of oxidized dietary oil and vitamin E supplementation on lipid profile and oxidation of muscle and liver of juvenile atlantic cod (Gadus morhua). J. Agric. Food Chem. 2007, 55, 6379–6386. [Google Scholar] [CrossRef] [PubMed]
- Saberi, A.H.; Fang, Y.; McClements, D.J. Fabrication of vitamin E-enriched nanoemulsions: Factors affecting particle size using spontaneous emulsification. J. Colloid Interface Sci. 2013, 391, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Hensley, K.; Benaksas, E.J.; Bolli, R.; Comp, P.; Grammas, P.; Hamdheydari, L.; Mou, S.; Pye, Q.N.; Stoddard, M.F.; Wallis, G.; et al. New perspectives on vitamin E: γ-tocopherol and carboxyethylhydroxychroman metabolites in biology and medicine. Free Radic. Biol. Med. 2004, 36, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Reiter, E.; Jiang, Q.; Christen, S. Anti-inflammatory properties of α- and γ-tocopherol. Mol. Asp. Med. 2007, 28, 668–691. [Google Scholar] [CrossRef] [PubMed]
- Etminan, M.; Gill, S.S.; Samii, A. Intake of vitamin E, vitamin C, and carotenoids and the risk of Parkinson’s disease: A meta-analysis. Lancet Neurol. 2005, 4, 362–365. [Google Scholar] [CrossRef]
- Li, F.J.; Shen, L.; Ji, H.F. Dietary intakes of vitamin E, vitamin C, and beta-carotene and risk of Alzheimer’s disease: A meta-analysis. J. Alzheimers Dis. 2012, 31, 253–258. [Google Scholar] [PubMed]
- De Camargo, A.C.; Regitano-d’Arce, M.A.B.; de Alencar, S.M.; Canniatti-Brazaca, S.G.; de Souza Vieira, T.M.F.; Shahidi, F. Chemical changes and oxidative stability of peanuts as affected by the dry-blanching. J. Am. Oil Chem. Soc. 2016, 8, 1101–1109. [Google Scholar] [CrossRef]
- Orem, A.; Yucesan, F.B.; Orem, C.; Akcan, B.; Kural, B.V.; Alasalvar, C.; Shahidi, F. Hazelnut-enriched diet improves cardiovascular risk biomarkers beyond a lipid-lowering effect in hypercholesterolemic subjects. J. Clin. Lipidol. 2013, 7, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, V.; Pitonzo, R.; Bartolotta, A.; D’Oca, M.C.; Fuochi, P. Effects of γ-irradiation on the α-tocopherol and fatty acids content of raw unpeeled almond kernels (Prunus dulcis). LWT—Food Sci. Technol. 2014, 59, 572–576. [Google Scholar] [CrossRef]
- De Camargo, A.C.; Regitano-d’Arce, M.A.B.; de Souza, T.M.F.; de Alencar, S.M.; Gallo, C.R.; Spoto, M.H.F.; da Silva, P.P.M.; Canniatti-Brazaca, S.G. Gamma irradiation to improve microbiological safety of peanuts: Effects on peroxide value, HS-SPME-GC/MS hexanal content and sensory acceptance. In Peanuts: Production, Nutritional Content and Health Implications; Cook, R.W., Ed.; Nova Science Publishers: New York, NY, USA, 2014; pp. 51–68. [Google Scholar]
- Sabliov, C.M.; Fronczek, C.; Astete, C.E.; Khachaturyan, M.; Khachatryan, L.; Leonardi, C. Effects of temperature and UV light on degradation of α-tocopherol in free and dissolved form. J. Am. Oil Chem. Soc. 2009, 86, 895–902. [Google Scholar] [CrossRef]
- Altunkaya, A.; Gökmen, V.; Skibsted, L.H. pH dependent antioxidant activity of lettuce (L. sativa) and synergism with added phenolic antioxidants. Food Chem. 2016, 190, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Thoo, Y.Y.; Abas, F.; Lai, O.-M.; Ho, C.W.; Yin, J.; Hedegaard, R.V.; Skibsted, L.H.; Tan, C.P. Antioxidant synergism between ethanolic centella asiatica extracts and α-tocopherol in model systems. Food Chem. 2013, 138, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.C.; Jorge, N. Influence of Lentinus edodes and Agaricus blazei extracts on the prevention of oxidation and retention of tocopherols in soybean oil in an accelerated storage test. J. Food Sci. Technol. 2014, 51, 1208–1212. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, A.; Shahidi, F. Determination of antioxidant activity in free and hydrolyzed fractions of millet grains and characterization of their phenolic profiles by HPLC-DAD-ESI-MSn. J. Funct. Foods 2011, 3, 144–158. [Google Scholar] [CrossRef]
- De Camargo, A.C.; Regitano-d’Arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Enzyme-assisted extraction of phenolics from winemaking by-products: Antioxidant potential and inhibition of alpha-glucosidase and lipase activities. Food Chem. 2016, 212, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Nishio, K.; Akazawa, Y.O.; Yamanaka, K.; Miyama, A.; Yoshida, Y.; Noguchi, N.; Niki, E. Cytoprotective effects of vitamin E homologues against glutamate-induced cell death in immature primary cortical neuron cultures: Tocopherols and tocotrienols exert similar effects by antioxidant function. Free Radic. Biol. Med. 2010, 49, 1542–1549. [Google Scholar] [CrossRef] [PubMed]
- Terasawa, Y.; Ladha, Z.; Leonard, S.W.; Morrow, J.D.; Newland, D.; Sanan, D.; Packer, L.; Traber, M.G.; Farese, R.V. Increased atherosclerosis in hyperlipidemic mice deficient in α-tocopherol transfer protein and vitamin E. Proc. Natl. Acad. Sci. USA 2000, 97, 13830–13834. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, A.; Arita, M.; Sato, Y.; Kiyose, C.; Ueda, T.; Igarashi, O.; Arai, H.; Inoue, K. Affinity for α-tocopherol transfer protein as a determinant of the biological activities of vitamin E analogs. FEBS Lett. 1997, 409, 105–108. [Google Scholar] [CrossRef]
- Mah, E.; Sapper, T.N.; Chitchumroonchokchai, C.; Failla, M.L.; Schill, K.E.; Clinton, S.K.; Bobe, G.; Traber, M.G.; Bruno, R.S. Alpha-tocopherol bioavailability is lower in adults with metabolic syndrome regardless of dairy fat co-ingestion: A randomized, double-blind, crossover trial. Am. J. Clin. Nutr. 2015, 102, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Drotleff, A.M.; Bohnsack, C.; Schneider, I.; Hahn, A.; Ternes, W. Human oral bioavailability and pharmacokinetics of tocotrienols from tocotrienol-rich (tocopherol-low) barley oil and palm oil formulations. J. Funct. Foods 2014, 7, 150–160. [Google Scholar] [CrossRef]
- Madhujith, T.; Shahidi, F. Antioxidative and antiproliferative properties of selected barley (Hordeum vulgarae L.) cultivars and their potential for inhibition of low-density lipoprotein (LDL) cholesterol oxidation. J. Agric. Food Chem. 2007, 55, 5018–5024. [Google Scholar] [CrossRef] [PubMed]
- Salah, N.; Miller, N.J.; Paganga, G.; Tijburg, L.; Bolwell, G.P.; Riceevans, C. Polyphenolic flavanols as scavengers of aqueous phase radicals and as chain-breaking antioxidants. Arch. Biochem. Biophys. 1995, 322, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Ma, C.-M.; Shahidi, F. Antioxidant and antiviral activities of lipophilic epigallocatechin gallate (EGCG) derivatives. J. Funct. Foods 2012, 4, 87–93. [Google Scholar] [CrossRef]
- Krumova, K.; Friedland, S.; Cosa, G. How lipid unsaturation, peroxyl radical partitioning, and chromanol lipophilic tail affect the antioxidant activity of α-tocopherol: Direct visualization via high-throughput fluorescence studies conducted with fluorogenic α-tocopherol analogues. J. Am. Oil Chem. Soc. 2012, 134, 10102–10113. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Jialal, I.; Fuller, C.J.; Huet, B.A. The effect of α-tocopherol supplementation on LDL oxidation: A dose-response study. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Roberts Ii, L.J.; Oates, J.A.; Linton, M.F.; Fazio, S.; Meador, B.P.; Gross, M.D.; Shyr, Y.; Morrow, J.D. The relationship between dose of vitamin E and suppression of oxidative stress in humans. Free Radic. Biol. Med. 2007, 43, 1388–1393. [Google Scholar] [CrossRef] [PubMed]
- Loffredo, L.; Perri, L.; di Castelnuovo, A.; Iacoviello, L.; de Gaetano, G.; Violi, F. Supplementation with vitamin E alone is associated with reduced myocardial infarction: A meta-analysis. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 354–363. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Cardiovascular Disease. Available online: http://www.who.int/cardiovascular_diseases/en/ (accessed on 9 August 2016).
- Christen, S.; Woodall, A.A.; Shigenaga, M.K.; SouthwellKeely, P.T.; Duncan, M.W.; Ames, B.N. Gamma-tocopherol traps mutagenic electrophiles such as NOx and complements alpha-tocopherol: Physiological implications. Proc. Natl. Acad. Sci. USA 1997, 94, 3217–3222. [Google Scholar] [CrossRef] [PubMed]
- Abid-Essefi, S.; Baudrimont, I.; Hassen, W.; Ouanes, Z.; Mobio, T.A.; Anane, R.; Creppy, E.E.; Bacha, H. DNA fragmentation, apoptosis and cell cycle arrest induced by zearalenone in cultured DOK, Vero and Caco-2 cells: Prevention by vitamin E. Toxicology 2003, 192, 237–248. [Google Scholar] [CrossRef]
- Campbell, S.E.; Stone, W.L.; Lee, S.; Whaley, S.; Yang, H.S.; Qui, M.; Goforth, P.; Sherman, D.; McHaffie, D.; Krishnan, K. Comparative effects of RRR-alpha- and RRR-gamma-tocopherol on proliferation and apoptosis in human colon cancer cell lines. BMC Cancer 2006, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.-P.; Lee, K.-J.; Choi, J.-W.; Yun, C.-O.; Nah, J.-W. Targeting delivery of tocopherol and doxorubicin grafted-chitosan polymeric micelles for cancer therapy: In vitro and in vivo evaluation. Colloids Surf. B 2015, 133, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Drotleff, A.M.; Büsing, A.; Willenberg, I.; Empl, M.T.; Steinberg, P.; Ternes, W. HPLC separation of vitamin E and its oxidation products and effects of oxidized tocotrienols on the viability of MCF-7 breast cancer cells in vitro. J. Agric. Food Chem. 2015, 63, 8930–8939. [Google Scholar] [CrossRef] [PubMed]
- Pierre, F.H.F.; Martin, O.C.B.; Santarelli, R.L.; Tache, S.; Naud, N.; Gueraud, F.; Audebert, M.; Dupuy, J.; Meunier, N.; Attaix, D.; et al. Calcium and alpha-tocopherol suppress cured-meat promotion of chemically induced colon carcinogenesis in rats and reduce associated biomarkers in human volunteers. Am. J. Clin. Nutr. 2013, 98, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.F.; Cai, Q.Q.; Geng, Q.R.; Wang, J.; Lan, Y.D.; Zhan, Y.Q.; Xu, D.Z. Vitamin intake reduce the risk of gastric cancer: Meta-analysis and systematic review of randomized and observational studies. PLoS ONE 2014, 9, e116060. [Google Scholar] [CrossRef] [PubMed]
- Virtamo, J.; Taylor, P.R.; Kontto, J.; Männistö, S.; Utriainen, M.; Weinstein, S.J.; Huttunen, J.; Albanes, D. Effects of α-tocopherol and β-carotene supplementation on cancer incidence and mortality: 18-year postintervention follow-up of the alpha-tocopherol, beta-carotene cancer prevention study. Int. J. Cancer 2014, 135, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Menkes, M.S.; Comstock, G.W.; Vuilleumier, J.P.; Helsing, K.J.; Rider, A.A.; Brookmeyer, R. Serum beta-carotene, vitamins A and E, selenium, and the risk of lung cancer. N. Engl. J. Med. 1986, 315, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Simmons-Menchaca, M.; Gapor, A.; Sanders, B.G.; Kline, K. Induction of apoptosis in human breast cancer cells by tocopherols and tocotrienols. Nutr. Cancer 1999, 33, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Wada, S.; Satomi, Y.; Murakoshi, M.; Noguchi, N.; Yoshikawa, T.; Nishino, H. Tumor suppressive effects of tocotrienol in vivo and in vitro. Cancer Lett. 2005, 229, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Tam, K.-W.; Ho, C.-T.; Lee, W.-J.; Tu, S.-H.; Huang, C.-S.; Chen, C.-S.; Lee, C.-H.; Wu, C.-H.; Ho, Y.-S. Alteration of α-tocopherol-associated protein (TAP) expression in human breast epithelial cells during breast cancer development. Food Chem. 2013, 138, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Wu, Z.; Li, G.; Teng, C.; Liu, Y.; Wang, F.; Zhao, Y.; Pang, D. The plasma level of retinol, vitamins A, C and α-tocopherol could reduce breast cancer risk? A meta-analysis and meta-regression. J. Cancer Res. Clin. Oncol. 2015, 141, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Kalender, Y.; Yel, M.; Kalender, S. Doxorubicin hepatotoxicity and hepatic free radical metabolism in rats: The effects of vitamin E and catechin. Toxicology 2005, 209, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, N.; Gapor, A.; Chambers, A.F.; Carroll, K.K. Inhibition of proliferation of estrogen receptor-negative MDA-MB-435 and -positive MCF-7 human breast cancer cells by palm oil tocotrienols and tamoxifen, alone and in combination. J. Nutr. 1997, 127, S544–S548. [Google Scholar]
- Pathak, R.; Bachri, A.; Ghosh, S.P.; Koturbash, I.; Boerma, M.; Binz, R.K.; Sawyer, J.R.; Hauer-Jensen, M. The vitamin E analog gamma-tocotrienol (Gt3) suppresses radiation-induced cytogenetic damage. Pharm. Res. 2016, 33, 2117–2125. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Kang, Z.; Wong, C. Vitamin E tocotrienols improve insulin sensitivity through activating peroxisome proliferator-activated receptors. Mol. Nutr. Food Res. 2010, 54, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Bharti, S.K.; Kumar, A.; Sharma, N.K.; Prakash, O.; Jaiswal, S.K.; Krishnan, S.; Gupta, A.K.; Kumar, A. Tocopherol from seeds of Cucurbita pepo against diabetes: Validation by in vivo experiments supported by computational docking. J. Formos. Med. Assoc. 2013, 112, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Reunanen, A.; Marniemi, J.; Leino, A.; Aromaa, A. Low vitamin E status is a potential risk factor for insulin-dependent diabetes mellitus. J. Intern. Med. 1999, 245, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Gosho, M.; Yamamoto, T.; Kobayashi, Y.; Ishii, N.; Ohashi, T.; Nakade, Y.; Ito, K.; Fukuzawa, Y.; Yoneda, M. Vitamin E has a beneficial effect on nonalcoholic fatty liver disease: A meta-analysis of randomized controlled trials. Nutrition 2015, 31, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Botella-Carretero, J.I.; Balsa, J.A.; Vazquez, C.; Peromingo, R.; Diaz-Enriquez, M.; Escobar-Morreale, H.F. Retinol and alpha-tocopherol in morbid obesity and nonalcoholic fatty liver disease. Obes. Surg. 2010, 20, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Strauss, R.S. Comparison of serum concentrations of α-tocopherol and β-carotene in a cross-sectional sample of obese and nonobese children (NHANES III). J. Pediatr. 1999, 134, 160–165. [Google Scholar] [CrossRef]
- Ima-Nirwana, S.; Suhaniza, S. Effects of tocopherols and tocotrienols on body composition and bone calcium content in adrenalectomized rats replaced with dexamethasone. J. Med. Food. 2004, 7, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Uto-Kondo, H.; Ohmori, R.; Kiyose, C.; Kishimoto, Y.; Saito, H.; Igarashi, O.; Kondo, K. Tocotrienol suppresses adipocyte differentiation and Akt phosphorylation in 3t3-L1 preadipocytes. J. Nutr. 2009, 139, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Chatgilialoglu, C.; Zambonin, L.; Altieri, A.; Ferreri, C.; Mulazzani, Q.G.; Landi, L. Geometrical isomerism of monounsaturated fatty acids: Thiyl radical catalysis and influence of antioxidant vitamins. Free Radic. Biol. Med. 2002, 33, 1681–1692. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Katan, M.B.; Ascherio, A.; Stampfer, M.J.; Willett, W.C. Trans fatty acids and cardiovascular disease. N. Engl. J. Med. 2006, 354, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.-L.; Sun Hwang, L.; Shahidi, F.; Pan, M.-H.; Wang, Y.; Ho, C.-T. Endogenous formation of trans fatty acids: Health implications and potential dietary intervention. J. Funct. Foods 2016, 25, 14–24. [Google Scholar] [CrossRef]
- Zhao, M.L.; Tang, L.; Zhu, X.M.; Hu, J.N.; Li, H.Y.; Luo, L.P.; Lei, L.; Deng, Z.Y. Enzymatic production of zero-trans plastic fat rich in α-linolenic acid and medium-chain fatty acids from highly hydrogenated soybean oil, cinnamomum camphora seed oil, and perilla oil by lipozyme TL IM. J. Agric. Food Chem. 2013, 61, 1189–1195. [Google Scholar] [CrossRef] [PubMed]
- Kemény, Z.; Recseg, K.; Hénon, G.; Kovári, K.; Zwobada, F. Deodorization of vegetable oils: Prediction of trans polyunsaturated fatty acid content. J. Am. Oil Chem. Soc. 2001, 78, 973–979. [Google Scholar] [CrossRef]
- Milstien, S.; Katusic, Z. Oxidation of tetrahydrobiopterin by peroxynitrite: Implications for vascular endothelial function. Biochem. Biophys. Res. Commun. 1999, 263, 681–684. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Gamma-tocopherol may promote effective no synthase function by protecting tetrahydrobiopterin from peroxynitrite. Med. Hypotheses 2007, 69, 1367–1370. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC). World Cancer Factsheet. Available online: http://publications.cancerresearchuk.org/downloads/Product/CS_REPORT_WORLD.pdf (accessed on 9 August 2016).
- Calori-Domingues, M.A.; Teles, D.; Ponce, G.H.; de Camargo, A.C.; Dias, C.T.S.; da Gloria, E.M.; Aguila, L.S.H.; de Godoy, I.J. A non-toxigenic strain of Aspergillus flavus as a biological control against aflatoxin production in peanuts. In Peanuts: Production, Nutritional Content and Health Implications; Cook, R.W., Ed.; Nova Science Publishers: New York, NY, USA, 2014; pp. 81–93. [Google Scholar]
- Santili, A.B.N.; de Camargo, A.C.; Nunes, R.S.R.; Gloria, E.M.d.; Machado, P.F.; Cassoli, L.D.; Dias, C.T.S.; Calori-Domingues, M.A. Aflatoxin M1 in raw milk from different regions of São Paulo state—Brazil. Food Addit. Contam. B 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Calori-Domingues, M.A.; Bernardi, C.M.G.; Nardin, M.S.; de Souza, G.V.; dos Santos, F.G.R.; Stein, M.A.; da Gloria, E.M.; Dias, C.T.S.; de Camargo, A.C. Co-occurrence and distribution of deoxynivalenol, nivalenol and zearalenone in wheat from Brazil. Food Addit. Contam. B 2016, 9, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Miranda, D.D.C.; Arçari, D.P.; Ladeira, M.S.P.; Calori-Domingues, M.A.; Romero, A.C.; Salvadori, D.M.F.; Gloria, E.M.; Pedrazzoli, J., Jr.; Ribeiro, M.L. Analysis of DNA damage induced by aflatoxin B1 in Dunkin-Hartley guinea pigs. Mycopathologia 2007, 163, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; He, Y.; Cui, X.-X.; Goodin, S.; Wang, H.; Du, Z.Y.; Li, D.; Zhang, K.; Tony Kong, A.-N.; DiPaola, R.S.; et al. Potent inhibitory effect of δ-tocopherol on prostate cancer cells cultured in vitro and grown as xenograft tumors in vivo. J. Agric. Food Chem. 2014, 62, 10752–10758. [Google Scholar] [CrossRef] [PubMed]
- Corpet, D.E. Red meat and colon cancer: Should we become vegetarians, or can we make meat safer? Meat Sci. 2011, 89, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Cheung-Ong, K.; Giaever, G.; Nislow, C. DNA-damaging agents in cancer chemotherapy: Serendipity and chemical biology. Chem. Biol. 2013, 20, 648–659. [Google Scholar] [CrossRef] [PubMed]
- Šimůnek, T.; Štěrba, M.; Popelová, O.; Adamcová, M.; Hrdina, R.; Geršl, V. Anthracycline-induced cardiotoxicity: Overview of studies examining the roles of oxidative stress and free cellular iron. Pharmacol. Rep. 2009, 61, 154–171. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; de Camargo, A.C.; Shahidi, F. Phenolic compounds of pomegranate by-products (outer skin, mesocarp, divider membrane) and their antioxidant activities. J. Agric. Food Chem. 2016, 64, 6584–6604. [Google Scholar] [CrossRef] [PubMed]
- Mayer, P.J.; Lange, C.S.; Bradley, M.O.; Nichols, W.W. Age-dependent decline in rejoining of X-ray-induced DNA double-strand breaks in normal human lymphocytes. Mutat. Res. 1989, 219, 95–100. [Google Scholar] [CrossRef]
- Singh, N.P.; Danner, D.B.; Tice, R.R.; Brant, L.; Schneider, E.L. DNA damage and repair with age in individual human lymphocytes. Mutat. Res. 1990, 237, 123–130. [Google Scholar] [CrossRef]
- Green, M.H.L.; Lowe, J.E.; Waugh, A.P.W.; Aldridge, K.E.; Cole, J.; Arlett, C.F. Effect of diet and vitamin C on DNA strand breakage in freshly-isolated human white blood cells. Mutat. Res. 1994, 316, 91–102. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Ramdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Kadouh, H.C.; Zhu, W.; Zhou, K. Bioactivity-guided isolation and purification of α-glucosidase inhibitor, 6-O-D-glycosides, from tinta Cão grape pomace. J. Funct. Foods 2016, 23, 573–579. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Jiang, W.J.; Xie, Z.T.; Wu, W.L.; Zhang, D.F. Vitamin E and risk of age-related cataract: A meta-analysis. Public Health Nutr. 2015, 18, 2804–2814. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Demleitner, M.F.; Song, L.; Rychlik, M.; Huang, D. Oligomeric proanthocyanidins are the active compounds in Abelmoschus esculentus Moench for its α-amylase and α-glucosidase inhibition activity. J. Funct. Foods 2016, 20, 463–471. [Google Scholar] [CrossRef]
- Mayer-Davis, E.J.; Costacou, T.; King, I.; Zaccaro, D.J.; Bell, R.A. Plasma and dietary vitamin E in relation to incidence of type 2 diabetes—The insulin resistance and atherosclerosis study (IRAS). Diabetes Care 2002, 25, 2172–2177. [Google Scholar] [CrossRef] [PubMed]
- Rösen, P.; Nawroth, P.P.; King, G.; Möller, W.; Tritschler, H.J.; Packer, L. The role of oxidative stress in the onset and progression of diabetes and its complications: Asummary of a congress series sponsored by UNESCO-MCBN, the American Diabetes Association and the German Diabetes Society. Diabetes Metab. Res. Rev. 2001, 17, 189–212. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Obesity. Available online: http://www.who.int/topics/obesity/en/ (accessed on 9 August 2016).
- Wang, Y.F.; Monteiro, C.; Popkin, B.M. Trends of obesity and underweight in older children and adolescents in the United States, Brazil, China, and Russia. Am. J. Clin. Nutr. 2002, 75, 971–977. [Google Scholar] [PubMed]
- Schmidt, M.I.; Duncan, B.B.; Silva, G.A.E.; Menezes, A.M.; Monteiro, C.A.; Barreto, S.M.; Chor, D.; Menezes, P.R. Chronic non-communicable diseases in Brazil: Burden and current challenges. Lancet 2011, 377, 1949–1961. [Google Scholar] [CrossRef]
- Mendez, M.A.; Monteiro, C.A.; Popkin, B.M. Overweight exceeds underweight among women in most developing countries. Am. J. Clin. Nutr. 2005, 81, 714–721. [Google Scholar] [PubMed]
- Monteiro, C.A.; Conde, W.L.; Popkin, B.M. Independent effects of income and education on the risk of obesity in the Brazilian adult population. J. Nutr. 2001, 131, 881S–886S. [Google Scholar] [PubMed]
- Adams, L.A.; Lindor, K.D. Nonalcoholic fatty liver disease. Ann. Epidemiol. 2007, 17, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Fang, X.; Marshall, M.R.; Chung, S. Regulation of obesity and metabolic complications by gamma and delta tocotrienols. Molecules 2016, 21, 344. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Shimomitsu, T.; Takanami, Y.; Murase, N.; Katsumura, T.; Maruyama, C. Vitamin E level changes in serum and red blood cells due to acute exhaustive exercise in collegiate women. J. Nutr. Sci. Vitaminol. 2000, 46, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Balazy, M.; Chemtob, S. Trans-arachidonic acids: New mediators of nitro-oxidative stress. Pharmacol. Ther. 2008, 119, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Zambonin, L.; Ferreri, C.; Cabrini, L.; Prata, C.; Chatgilialoglu, C.; Landi, L. Occurrence of trans fatty acids in rats fed a trans-free diet: A free radical-mediated formation? Free Radic. Biol. Med. 2006, 40, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Zambonin, L.; Prata, C.; Cabrini, L.; Maraldi, T.; Fiorentini, D.; Sega, F.V.D.; Hakim, G.; Landi, L. Effect of radical stress and ageing on the occurrence of trans fatty acids in rats fed a trans-free diet. Free Radic. Biol. Med. 2008, 44, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Costantino, C.; Chatgilialoglu, C.; Landi, L.; Mulazzani, Q.G. The thiyl radical-mediated isomerization of cis-monounsaturated fatty acid residues in phospholipids: A novel path of membrane damage? Chem. Commun. 1999, 407–408. [Google Scholar] [CrossRef]
- Ferreri, C.; Costantino, C.; Perrotta, L.; Landi, L.; Mulazzani, Q.G.; Chatgilialoglu, C. Cis-trans isomerization of polyunsaturated fatty acid residues in phospholipids catalyzed by thiyl radicals. J. Am. Oil Chem. Soc. 2001, 123, 4459–4468. [Google Scholar] [CrossRef]
- Konings, A.W.T.; Damen, J.; Trieling, W.B. Protection of liposomal lipids against radiation-induced oxidative damage. Int. J. Radiat. Biol. 1979, 35, 343–350. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Oil [Reference] | Alpha-T | Beta-T | Gamma-T | Delta-T | Alpha-T3 | Beta-T3 | Gamma-T3 | Delta-T3 |
|---|---|---|---|---|---|---|---|---|
| Barley [23,24] | 14.2–20.1 | 0.60–1.90 | 3.50–15.1 | 0.90–4.60 | 46.5–76.1 | nd–12.4 | 8.50–18.6 | 0.50–2.6 |
| Coconut [11,21] | 0.20–1.82 | tr–0.25 | tr–0.12 | nd–0.39 | 1.09–3.00 | nd–0.17 | 0.33–0.64 | nd–0.10 |
| Corn [11,21] | 18.0–25.7 | 0.95–1.10 | 44.0–75.2 | 2.20–3.25 | 0.94–1.50 | nd | 1.30–2.00 | nd–0.26 |
| Cottonseed [25,26] | 30.5–57.3 | 0.04–0.30 | 10.5–31.7 | tr | nr | nr | nr | nr |
| Linseed [11,21] | 0.54–1.20 | nd–tr | 52.0–57.3 | 0.75–0.95 | nd | nd | nd | nd |
| Olive [11,21] | 11.9–17.0 | nd–0.27 | 0.89–1.34 | nd–tr | nd–tr | nd | nd | nd–tr |
| Palm [21,27,28] | 6.05–42.0 | nd–0.42 | tr–0.02 | nd–0.02 | 5.70–26.0 | nr–0.82 | 11.3–36.0 | 3.33–8.00 |
| Peanut [21,26] | 8.86–30.4 | nd–0.38 | 3.50–19.2 | 0.85–3.10 | nd | nd | nd | nd |
| Rapeseed [11,21] | 18.9–24.0 | nd–tr | 37–51 | 0.98–1.90 | nd | nd | nd | nd |
| Rice bran [25,29] | 0.73–15.9 | 0.19–2.5 | 0.26–8.00 | 0.03–2.70 | 0.84–13.8 | tr–2.6 | 1.74–23.1 | 0.14–2.53 |
| Safflower [21,26] | 36.7–47.7 | nd–1.20 | tr–2.56 | tr–0.65 | nd | nd | nd | nd |
| Sesame [11,30] | 0.24–36.0 | 0.28–0.80 | 16.0–57.0 | 0.17–13.0 | tr | nd | 0.34 | nr |
| Soybean [21,31] | 9.53–12.0 | 1.00–1.31 | 61.0–69.9 | 23.9–26.0 | nd | nd | nd | nd |
| Sunflower [11,25] | 32.7–59.0 | tr–2.40 | 1.40–4.50 | 0.27–0.50 | 0.11 | nd | tr | tr |
| Wheat germ [11,21] | 151–192 | 31.2–65.0 | tr–52.3 | nd–0.55 | 2.5–3.6 | nd–8.2 | nd–1.85 | nd–0.24 |
| Oil [Reference] | Alpha-T | Beta-T | Gamma-T | Delta-T |
|---|---|---|---|---|
| Camelina [11] | 3.80 | 0.09 | 72.0 | 1.50 |
| Grape [34] | 11.8 | nr | 60.1 | nr |
| Guariroba [40] | 1.19 | nd | nd | 0.78 |
| Guava [34] | 45.8 | nr | 93.1 | nr |
| Jatoba [41] | 88.6 | nr | 16.9 | nr |
| Jerivá [40] | 1.10 | 0.10 | nd | 0.78 |
| Lemon [37] | 102 | 2.20 | 1.33 | 19.0 |
| Macaúba [40] | 1.44 | 0.08 | nd | 0.79 |
| Melon [34] | 20.5 | nr | 250 | nr |
| Orange [37] | 300 | nd | nd | 18.6 |
| Papaya [36] | 5.18 | 0.21 | 0.18 | 1.89 |
| Passion fruit [35] | nd | 5.40 | 16.7 | 27.9 |
| Pinha [42] | 1.20 | 0.33 | 12.3 | 0.02 |
| Pumpkin [34] | 7.30 | nr | 294.5 | nr |
| Soursop [34] | 22.1 | nr | 7.10 | nr |
| Tamarind [43] | 1.24 | 0.70 | 3.82 | 0.02 |
| Tangerine [37] | 116 | 2.22 | nd | 19.2 |
| Tomato [34] | nd | nr | 32.9 | nr |
| Test Material | Main Outcomes | Disease/Function | Reference |
|---|---|---|---|
| Alpha-T | Synergism of alpha-tocopherol with hydrophilic phenolic extracts from Centella asiatica, açaí, and grape by-products in liposome model systems was dependent on the mixture ratio | Potential preventive oxidation in lipid membranes | [74,108] |
| Alpha-T | Healthy men who received at least 400 IU/day showed lower levels of oxidized LDL-cholesterol | Cardiovascular diseases | [122] |
| Alpha-T | Supplementation with alpha-tocopherol alone (400–800 IU/daily), but not in combination with other antioxidants, decreases fatal myocardial infarction by about 20% according to a meta-analysis evaluating randomized clinical trials | Cardiovascular diseases | [124] |
| Gamma-T | Generation of stable carbon-centered adducts through the nucleophilic 5-position thus trapping membrane-soluble electrophilic nitrogen oxides as evaluated in liposome model system and human LDL-cholesterol in vitro | Cardiovascular diseases | [126] |
| Alpha-T | Reduction of zearalenone-induced DNA fragmentation and apoptotic body formation using Vero, Caco-2 and DOK cell lines | Cancer | [127] |
| Alpha- and gamma-T | Gamma-T, but not alpha-T, reduced malignant colon cancer cell proliferation (SW480, HT-29, HCT-116, and HCT-15 cells) and the effectivity was dependent on the molecular characteristics of the cells | Cancer | [128] |
| Alpha-T | Polymeric micelles (targeting ligand-modified TOC-DOX) containing alpha-T (TOC) and doxorubicin (DOX) reduced proliferating cell nuclear antigen expression in tumor tissues of HER2/neu-positive SK-BR-3 tumor-bearing mice | Cancer | [129] |
| Alpha- and gamma-T3 | Both oxidized and nonoxidized gamma-T3 decreased the viability of MCF‑7 breast cancer cells in vitro and the former was more effective | Cancer | [130] |
| Alpha-T | Alpha-T reduced the number of mucin-depleted foci (a precancer lesion) in rats fed a diet containing cure meat | Cancer | [131] |
| Alpha-T | Alpha-T (10 mg/day) decreases the risk of gastric cancer development by 24% as evaluated by a meta-analysis | Cancer | [132] |
| Alpha-T | Alpha-T (50 mg/day) prevented the onset of prostate cancer and its related mortality showed long-term decrease (18-year post-trial) | Cancer | [133] |
| Alpha-T | Lower levels of alpha-T were associated with lung cancer development in humans | Cancer | [134] |
| Delta-T, alpha-, beta- and gamma-T3 | The mentioned tocols, but not alpha-, beta-, and gamma-T, were apoptotic inducers in both estrogen-responsive MCF7 and estrogen-nonresponsive MDA-MB-435 human breast cancer cell lines in vitro | Cancer | [135] |
| Alpha-, beta-, delta-, and gamma-T3 | All tocotrienols showed anticarcinogenic effects in lung and liver cancer in mice | Cancer | [136] |
| Alpha-, beta-, delta-, and gamma-T3 | Delta-T3 showed higher antiproliferative effect in human hepatocellular carcinoma HepG2 cells in vitro but increased CYP1A1 gene expression raising a concern of potential carcinogenic effect in some cases | Cancer | [136] |
| Alpha-T | Alpha-T-associated protein (TAP) expression was associated with the reduction of alpha-T levels in human breast tumour samples | Cancer | [137] |
| Alpha-T | Severe deficiency of alpha-T was associated with higher risk of breast cancer development as evaluated through a meta-analysis | Cancer | [138] |
| Alpha-T | Alpha-T (200 IU/kg/week) reduced the doxorubicin-induced hepatotoxicity in rats and decreased malondialdehyde levels in their liver | Cancer | [139] |
| Gamma- and delta-T3 | Both gamma- and delta-T3 showed synergistic effects against MCF-7 human breast cancer cells when combined with tamoxifen in proportion 1:1 | Cancer | [140] |
| Tocotrienol-rich fraction (TRF) from palm oil, alpha-T, alpha-, gamma-, and delta-T3 | All of them combined with tamoxifen (1:1) showed synergistic effects towards MDA-MB-435 cells proliferation | Cancer | [140] |
| Gamma-T3 | Gamma-T3 inhibited DNA double-strand breaks in gamma-irradiated human umbilical vein endothelial cells, positively influenced the expression of the DNA-repair gene RAD50 and diminished radiation-induced RAD50 suppression | Cancer | [141] |
| Alpha-, delta- and gamma-T3 | Peroxisome proliferator-activated receptors (PPAR) were modulated by alpha-T3 (PPARα), gamma-T3 (PPARα), and delta-T3 (PPARα, PPARγ, and PPARδ) | Diabetes | [142] |
| Tocotrienol-rich fraction (TRF) of palm oil | PPAR target genes of diabetic mice were regulated by TRF of palm oil, improved their glucose levels and sensitivity to insulin | Diabetes | [142] |
| Alpha-, beta-, delta-, and gamma-T | Antidiabetic properties tocopoherols are related to their docking to dipeptidyl peptidase IV and peroxisome proliferator-activated receptor gamma and the interaction nature (hydrogen bond, hydrophobic, or Pi-Pi) is dependent on the interacting residue | Diabetes | [143] |
| Alpha-T | Insulin-dependent diabetes mellitus (IDDM) with lower levels of alpha-T showed higher prevalence of IDDM after 4–14 years baseline examination | Diabetes | [144] |
| Alpha-T | Liver function and histologic changes in patients with nonalcoholic fatty liver disease, including nonalcoholic steatohepatitis were improved by alpha-T as evaluated by a meta-analysis of randomized controlled trials | Obesity | [145] |
| Alpha-T | Body mass index of morbidly obese patients was inversely correlated with their alpha-T serum concentration | Obesity | [146] |
| Alpha-T | Obese children (6–19 years old) showed lower serum concentration of alpha-T as demonstrated by the National Health and Examination Survey (NHANES III) | Obesity | [147] |
| Alpha-T and gamma-T3 | Gamma-T3 (60 mg/kg of body weight/day), but not alpha-T, reduced the body fat mass of rats treated with glucocorticoid | Obesity | [148] |
| Alpha-T, alpha- and gamma-T3 | Alpha- and gamma-T3, but not alpha-T, prevented differentiation of preadipocytes into adipocytes in 3T3-L1 cells | Obesity | [149] |
| Alpha-T | Cis-trans isomerization of phospholipid bilayers induced by thiyl radicals was inhibited by alpha-T | Cell membrane function | [150] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahidi, F.; De Camargo, A.C. Tocopherols and Tocotrienols in Common and Emerging Dietary Sources: Occurrence, Applications, and Health Benefits. Int. J. Mol. Sci. 2016, 17, 1745. https://doi.org/10.3390/ijms17101745
Shahidi F, De Camargo AC. Tocopherols and Tocotrienols in Common and Emerging Dietary Sources: Occurrence, Applications, and Health Benefits. International Journal of Molecular Sciences. 2016; 17(10):1745. https://doi.org/10.3390/ijms17101745
Chicago/Turabian StyleShahidi, Fereidoon, and Adriano Costa De Camargo. 2016. "Tocopherols and Tocotrienols in Common and Emerging Dietary Sources: Occurrence, Applications, and Health Benefits" International Journal of Molecular Sciences 17, no. 10: 1745. https://doi.org/10.3390/ijms17101745
APA StyleShahidi, F., & De Camargo, A. C. (2016). Tocopherols and Tocotrienols in Common and Emerging Dietary Sources: Occurrence, Applications, and Health Benefits. International Journal of Molecular Sciences, 17(10), 1745. https://doi.org/10.3390/ijms17101745