
Cerebellar Expression of the Neurotrophin Receptor p75 in Naked-Ataxia Mutant Mouse


 and
and 
Abstract
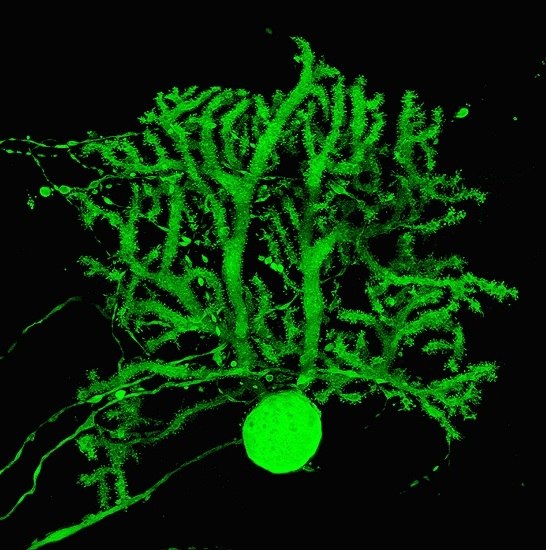
:
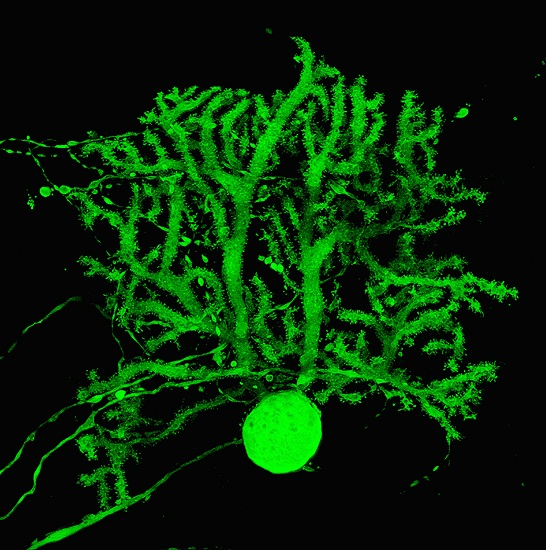{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animal Maintenance and Tissue Processing
2.2. Antibodies
2.3. Immunohistochemistry
2.4. Western Blotting
2.5. Preparation of the Figures
2.6. Statistical Methods
3. Results
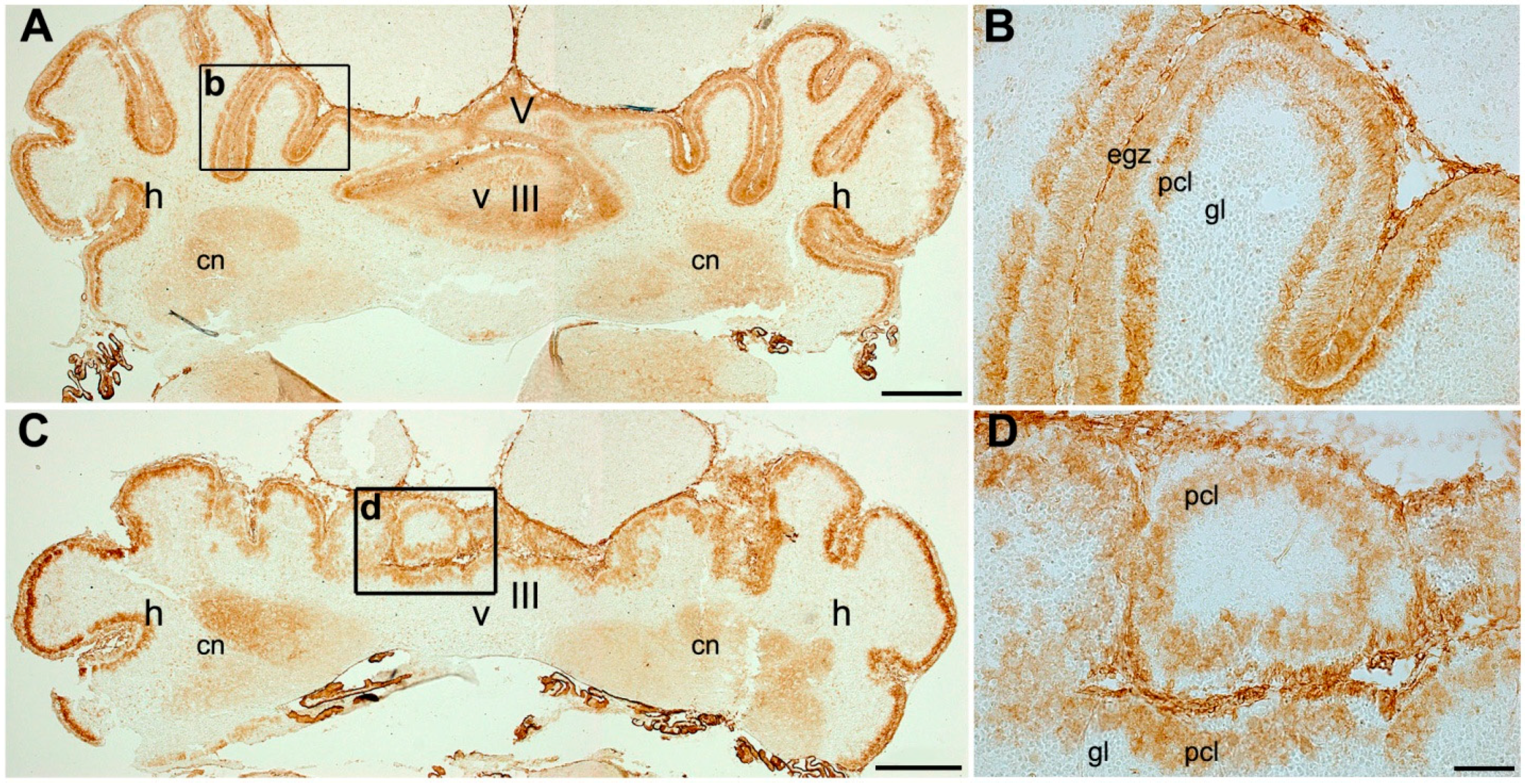
3.1. Expression Pattern of p75NTR in Adult Mouse Cerebellum
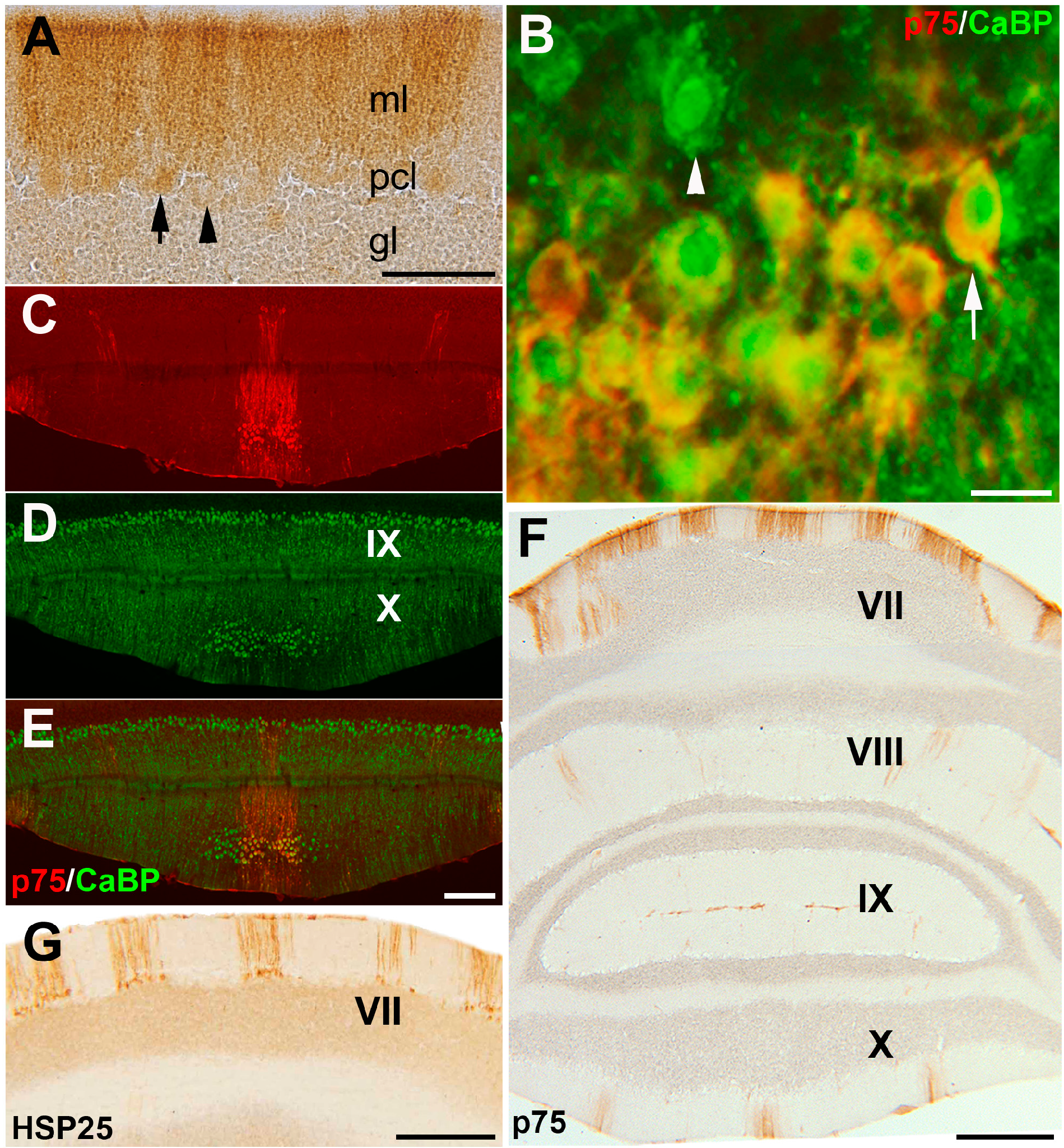
3.2. Pattern Expression of p75NTR in wt and nax Cerebellum at P4 and P6
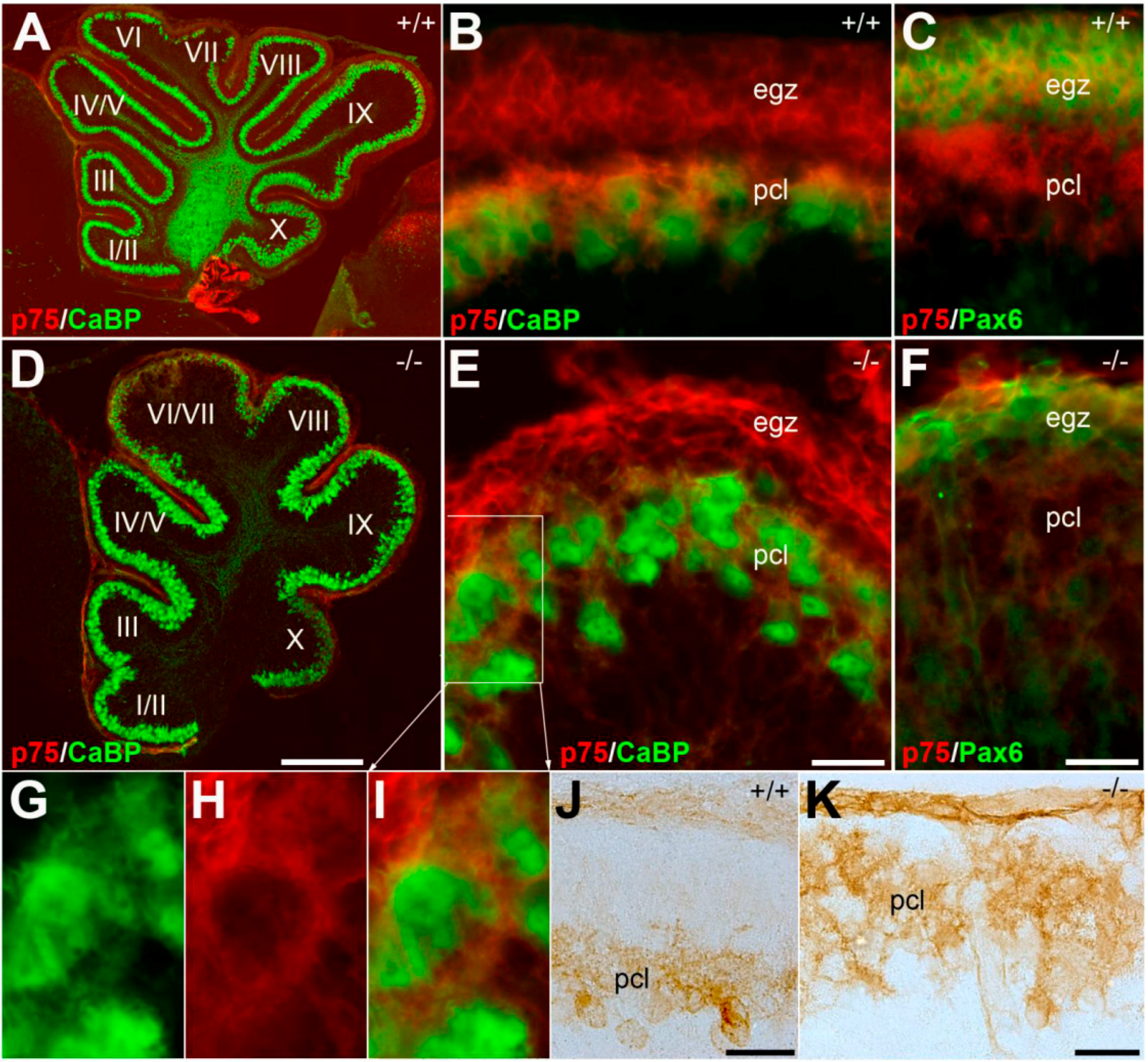
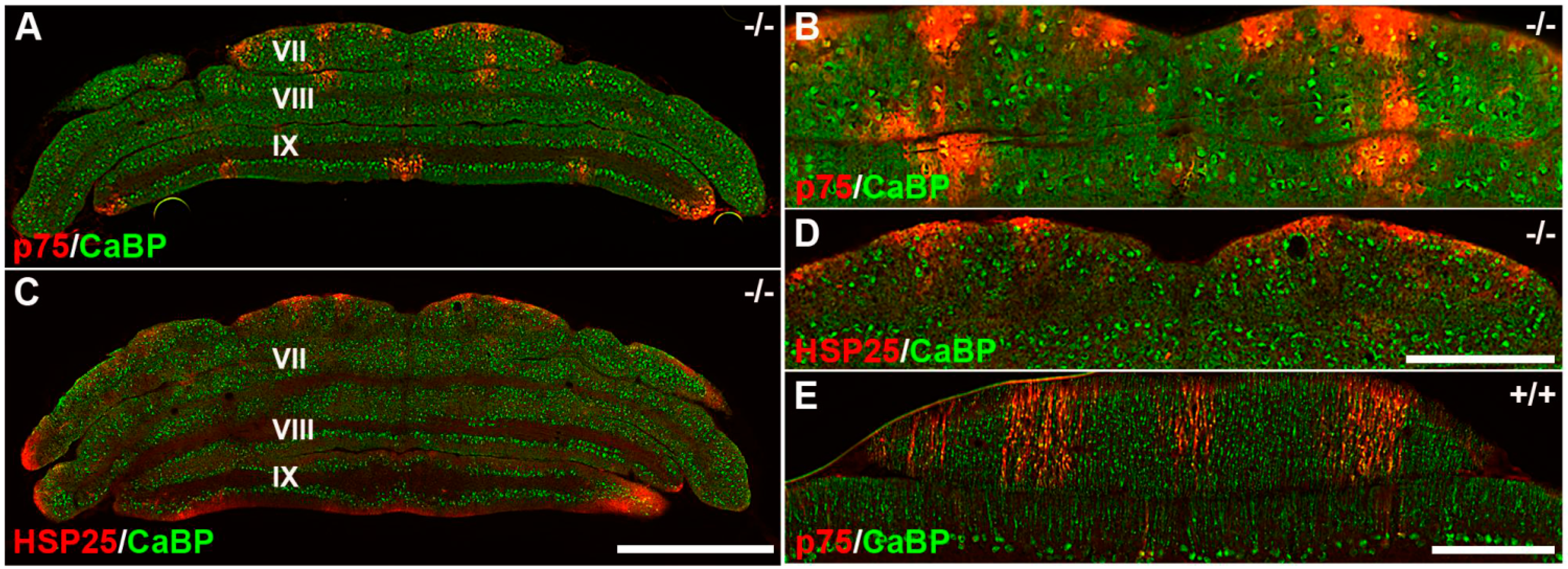
3.3. Pattern Expression of p75NTR in wt and nax at P22
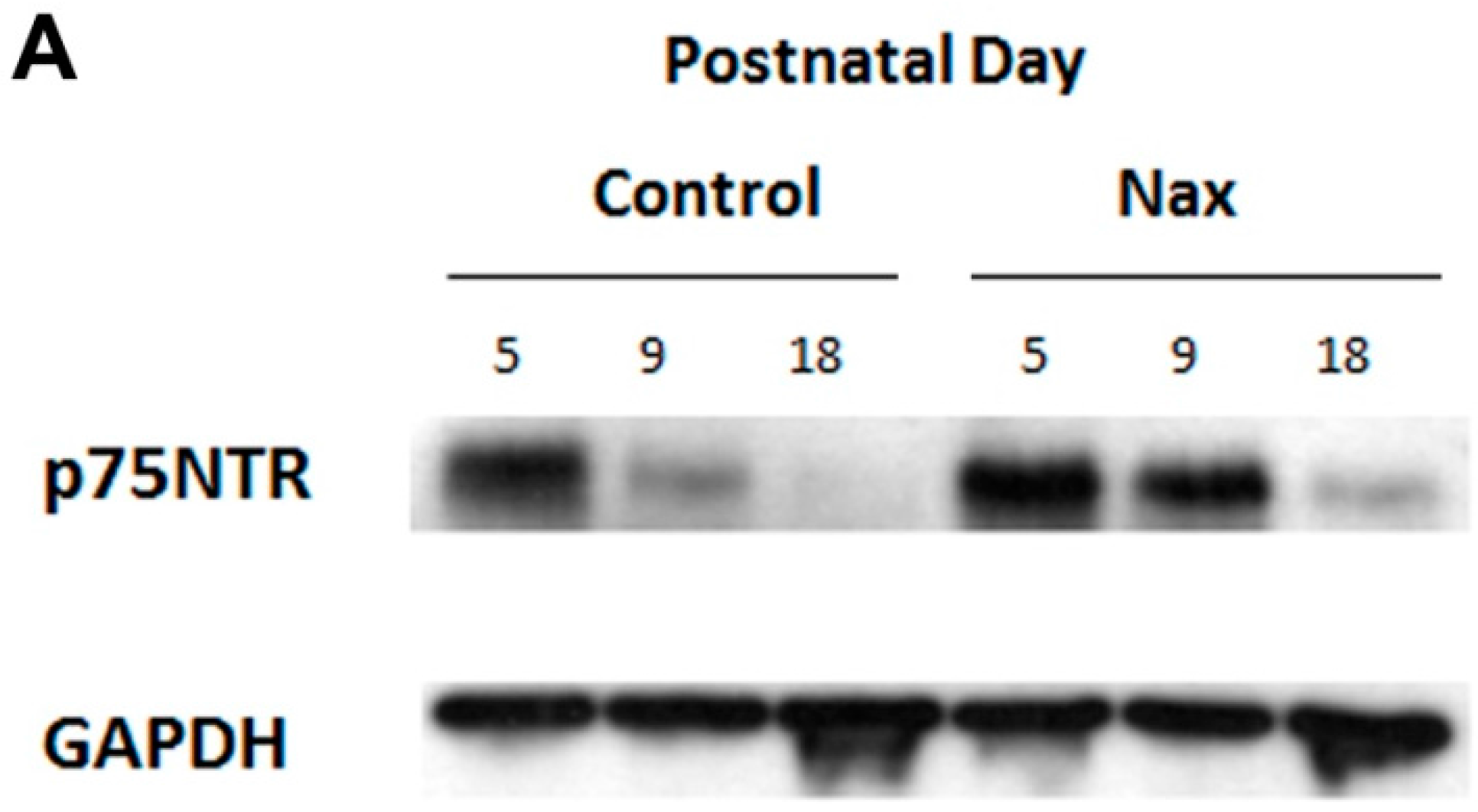
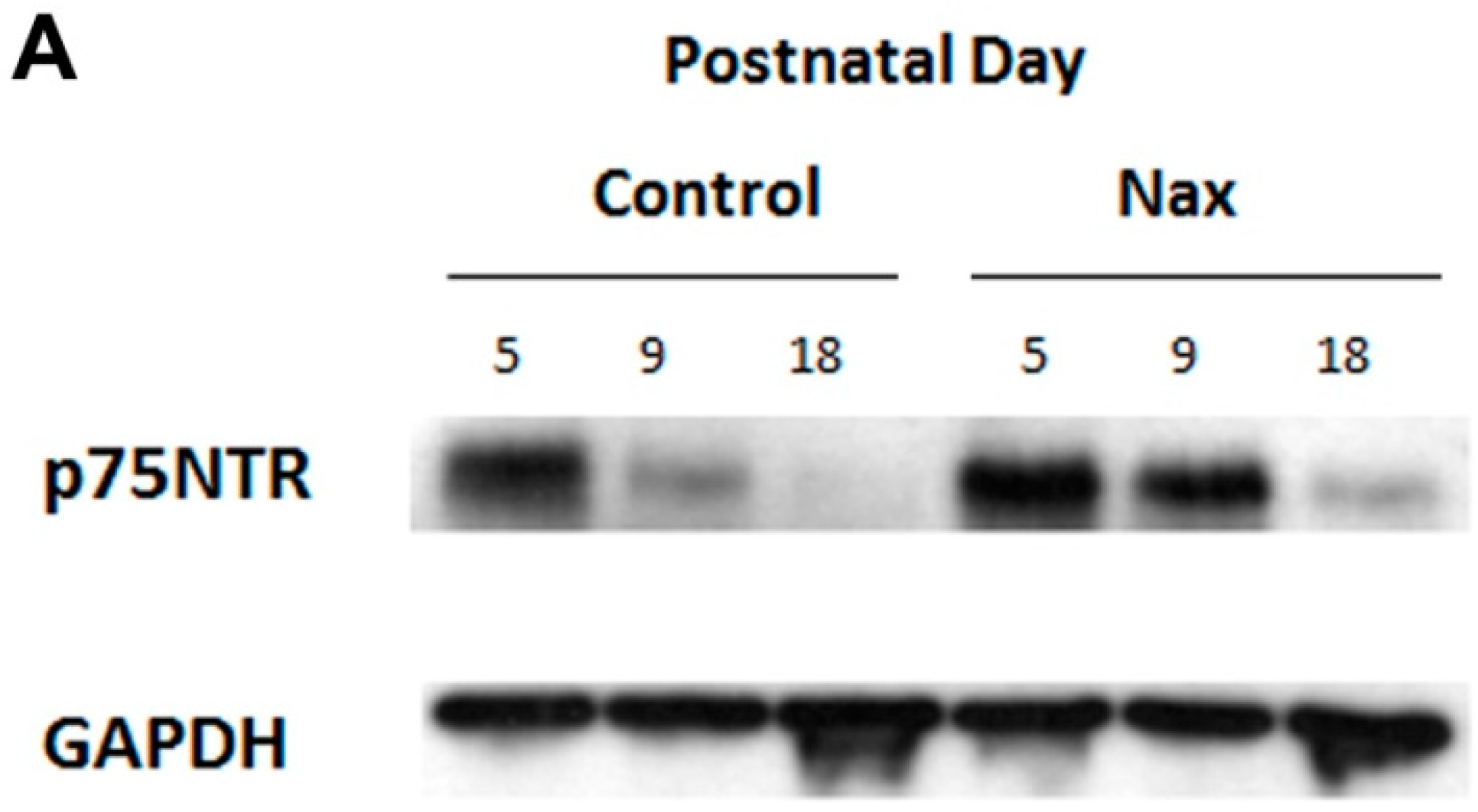
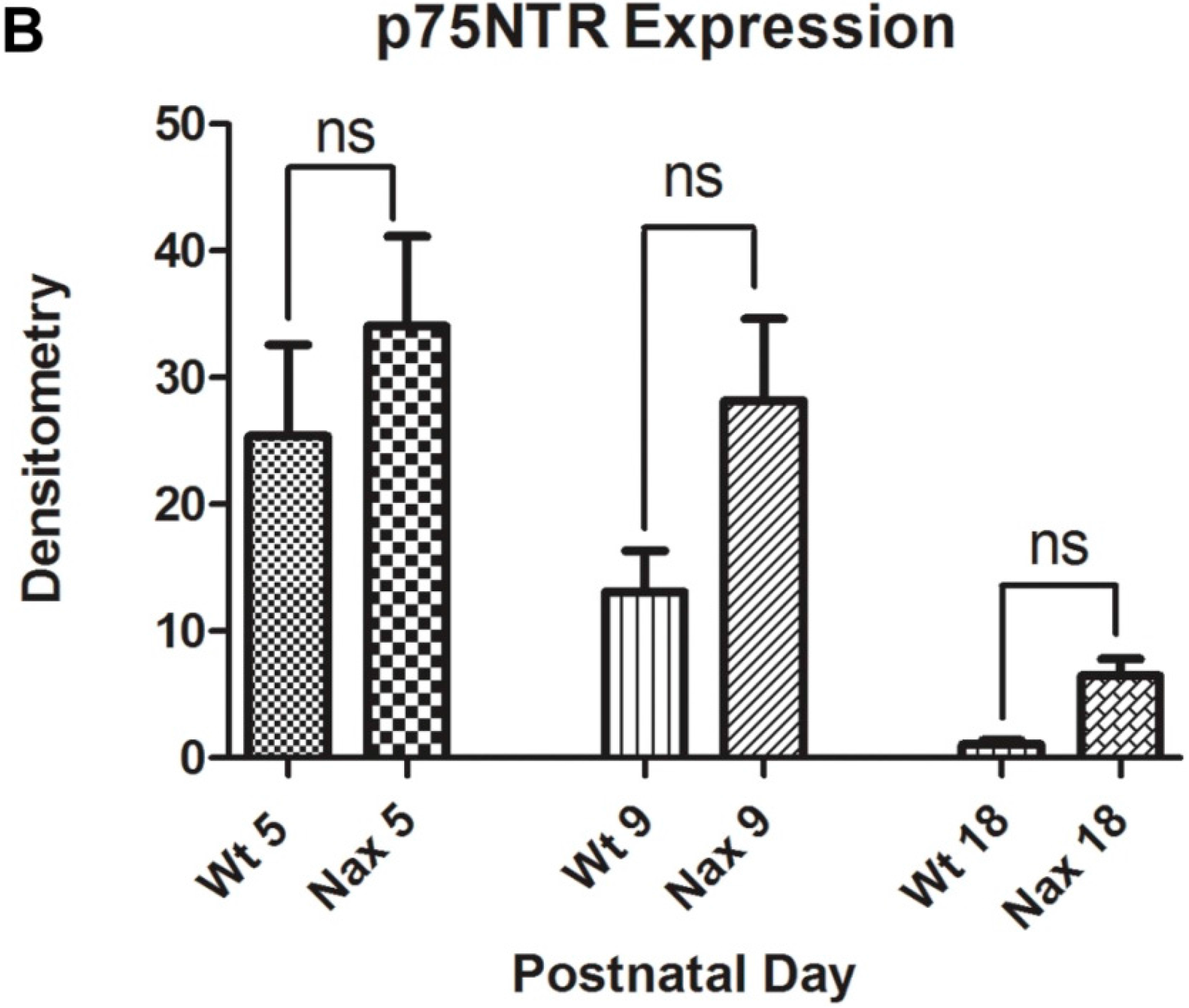
3.4. Protein Expression of p75NTR in wt and nax Cerebellum
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bailey, K.; Rahimi Balaei, M.; Mehdizadeh, M.; Marzban, H. Spatial and temporal expression of lysosomal acid phosphatase 2 (Acp2) reveals dynamic patterning of the mouse cerebellar cortex. Cerebellum 2013, 12, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.; Rahimi Balaei, M.; Mannan, A.; del Bigio, M.R.; Marzban, H. Purkinje cell compartmentation in the cerebellum of the lysosomal acid phosphatase 2 mutant mouse (nax—naked-ataxia mutant mouse). PLoS ONE 2014, 9, e94327. [Google Scholar] [CrossRef] [PubMed]
- Mannan, A.U.; Roussa, E.; Kraus, C.; Rickmann, M.; Maenner, J.; Nayernia, K.; Krieglstein, K.; Reis, A.; Engel, W. Mutation in the gene encoding lysosomal acid phosphatase (Acp2) causes cerebellum and skin malformation in mouse. Neurogenetics 2004, 5, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Melquist, S.; Craig, D.W.; Huentelman, M.J.; Crook, R.; Pearson, J.V.; Baker, M.; Zismann, V.L.; Gass, J.; Adamson, J.; Szelinger, S.; et al. Identification of a novel risk locus for progressive supranuclear palsy by a pooled genomewide scan of 500,288 single-nucleotide polymorphisms. Am. J. Hum. Genet. 2007, 80, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Pohl, S.; Mitchison, H.M.; Kohlschutter, A.; van Diggelen, O.; Braulke, T.; Storch, S. Increased expression of lysosomal acid phosphatase in CLN3-defective cells and mouse brain tissue. J. Neurochem. 2007, 103, 2177–2188. [Google Scholar] [CrossRef] [PubMed]
- Saftig, P.; Hartmann, D.; Lullmann-Rauch, R.; Wolff, J.; Evers, M.; Koster, A.; Hetman, M.; von Figura, K.; Peters, C. Mice deficient in lysosomal acid phosphatase develop lysosomal storage in the kidney and central nervous system. J. Biol. Chem. 1997, 272, 18628–18635. [Google Scholar] [CrossRef] [PubMed]
- Makrypidi, G.; Damme, M.; Muller-Loennies, S.; Trusch, M.; Schmidt, B.; Schluter, H.; Heeren, J.; Lubke, T.; Saftig, P.; Braulke, T. Mannose 6 dephosphorylation of lysosomal proteins mediated by acid phosphatases Acp2 and Acp5. Mol. Cell. Biol. 2012, 32, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Marzban, H.; Kim, C.T.; Doorn, D.; Chung, S.H.; Hawkes, R. A novel transverse expression domain in the mouse cerebellum revealed by a neurofilament-associated antigen. Neuroscience 2008, 153, 1190–1201. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Fukui, Y.; Hawkes, R. Spatial distribution of corticotropin-releasing factor immunopositive climbing fibers in the mouse cerebellum: Analysis by whole mount immunohistochemistry. Brain Res. 2008, 1222, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Ozol, K.; Hayden, J.M.; Oberdick, J.; Hawkes, R. Transverse zones in the vermis of the mouse cerebellum. J. Comp. Neurol. 1999, 412, 95–111. [Google Scholar] [CrossRef]
- Marzban, H.; Hawkes, R. On the architecture of the posterior zone of the cerebellum. Cerebellum 2011, 10, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Apps, R.; Hawkes, R. Cerebellar cortical organization: A one-map hypothesis. Nat. Rev. Neurosci. 2009, 10, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Consalez, G.G.; Hawkes, R. The compartmental restriction of cerebellar interneurons. Front. Neural Circuits 2012, 6, 123. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.H.; Marzban, H.; Hawkes, R. Compartmentation of the cerebellar nuclei of the mouse. Neuroscience 2009, 161, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Marzban, H.; Chung, S.H.; Pezhouh, M.K.; Feirabend, H.; Watanabe, M.; Voogd, J.; Hawkes, R. Antigenic compartmentation of the cerebellar cortex in the chicken (Gallus domesticus). J. Comp. Neurol. 2010, 518, 2221–2239. [Google Scholar] [CrossRef] [PubMed]
- Marzban, H.; Chung, S.; Watanabe, M.; Hawkes, R. Phospholipase Cβ4 expression reveals the continuity of cerebellar topography through development. J. Comp. Neurol. 2007, 502, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, L.; Eng, J.; Ivanov, N.; Garden, G.A.; La Spada, A.R. Autophagy activation and enhanced mitophagy characterize the Purkinje cells of pcd mice prior to neuronal death. Mol. Brain 2009, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Fayaz, S.M.; Suvanish Kumar, V.S.; Rajanikant, G.K. Necroptosis: Who knew there were so many interesting ways to die? CNS Neurol. Disorders Drug Targets 2014, 13, 42–51. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Yankner, B.A. Apoptosis in the nervous system. Nature 2000, 407, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Ichim, G.; Tauszig-Delamasure, S.; Mehlen, P. Neurotrophins and cell death. Exp. Cell Res. 2012, 318, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Mamidipudi, V.; Wooten, M.W. Dual role for p75(NTR) signaling in survival and cell death: Can intracellular mediators provide an explanation? J. Neurosci. Res. 2002, 68, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Florez-McClure, M.L.; Linseman, D.A.; Chu, C.T.; Barker, P.A.; Bouchard, R.J.; Le, S.S.; Laessig, T.A.; Heidenreich, K.A. The p75 neurotrophin receptor can induce autophagy and death of cerebellar purkinje neurons. J. Neurosci. 2004, 24, 4498–4509. [Google Scholar] [CrossRef] [PubMed]
- Meeker, R.B.; Williams, K.S. The p75 neurotrophin receptor: At the crossroad of neural repair and death. Neural Regen. Res. 2015, 10, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Tano, D.; Napieralski, J.A.; Eisenman, L.M.; Messer, A.; Plummer, J.; Hawkes, R. Novel developmental boundary in the cerebellum revealed by zebrin expression in the Lurcher (Lc/+) mutant mouse. J. Comp. Neurol. 1992, 323, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Baimbridge, K.G.; Miller, J.J. Immunohistochemical localization of calcium-binding protein in the cerebellum, hippocampal formation and olfactory bulb of the rat. Brain Res. 1982, 245, 223–229. [Google Scholar] [CrossRef]
- De Camilli, P.; Miller, P.E.; Levitt, P.; Walter, U.; Greengard, P. Anatomy of cerebellar Purkinje cells in the rat determined by a specific immunohistochemical marker. Neuroscience 1984, 11, 761–817. [Google Scholar] [CrossRef]
- Lotta, L.T.; Conrad, K.; Cory-Slechta, D.; Schor, N.F. Cerebellar Purkinje cell p75 neurotrophin receptor and autistic behavior. Transl. Psychiatry 2014, 4, e416. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.L.; Krueger-Naug, A.M.; Currie, R.W.; Hawkes, R. Constitutive expression of the 25-KDA heat shock protein HSP25 reveals novel parasagittal bands of purkinje cells in the adult mouse cerebellar cortex. J. Comp. Neurol. 2000, 416, 383–397. [Google Scholar] [CrossRef]
- Engelkamp, D.; Rashbass, P.; Seawright, A.; van Heyningen, V. Role of Pax6 in development of the cerebellar system. Development 1999, 126, 3585–3596. [Google Scholar] [PubMed]
- Sarna, J.R.; Hawkes, R. Patterned Purkinje cell death in the cerebellum. Prog. Neurobiol. 2003, 70, 473–507. [Google Scholar] [CrossRef]
- Sarna, J.R.; Larouche, M.; Marzban, H.; Sillitoe, R.V.; Rancourt, D.E.; Hawkes, R. Patterned Purkinje cell degeneration in mouse models of Niemann-Pick type C disease. J. Comp. Neurol. 2003, 456, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Duffin, C.A.; McFarland, R.; Sarna, J.R.; Vogel, M.W.; Armstrong, C.L. Heat shock protein 25 expression and preferential Purkinje cell survival in the Lurcher mutant mouse cerebellum. J. Comp. Neurol. 2010, 518, 1892–1907. [Google Scholar] [CrossRef] [PubMed]
- Sarna, J.R.; Hawkes, R. Patterned Purkinje cell loss in the ataxic sticky mouse. Eur. J. Neurosci. 2011, 34, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Sarna, J.R.; Marzban, H.; Watanabe, M.; Hawkes, R. Complementary stripes of phospholipase Cβ3 and Cβ4 expression by purkinje cell subsets in the mouse cerebellum. J. Comp. Neurol. 2006, 496, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Marzban, H.; Zahedi, S.; Sanchez, M.; Hawkes, R. Antigenic compartmentation of the cerebellar cortex in the syrian hamster mesocricetus auratus. Brain Res. 2003, 974, 176–183. [Google Scholar] [CrossRef]
- Marzban, H.; Khanzada, U.; Shabir, S.; Hawkes, R.; Langnaese, K.; Smalla, K.H.; Bockers, T.M.; Gundelfinger, E.D.; Gordon-Weeks, P.R.; Beesley, P.W. Expression of the immunoglobulin superfamily neuroplastin adhesion molecules in adult and developing mouse cerebellum and their localisation to parasagittal stripes. J. Comp. Neurol. 2003, 462, 286–301. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.L.; Vogel, M.W.; Hawkes, R. Development of HSP25 expression compartments is not constrained by Purkinje cell defects in the lurcher mouse mutant. J. Comp. Neurol. 2005, 491, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Jakob, U.; Gaestel, M.; Engel, K.; Buchner, J. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993, 268, 1517–1520. [Google Scholar] [PubMed]
- Lavoie, J.N.; Gingras-Breton, G.; Tanguay, R.M.; Landry, J. Induction of chinese hamster HSP27 gene expression in mouse cells confers resistance to heat shock. HSP27 stabilization of the microfilament organization. J. Biol. Chem. 1993, 268, 3420–3429. [Google Scholar] [PubMed]
- Mehlen, P.; Kretz-Remy, C.; Preville, X.; Arrigo, A.P. Human HSP27, drosophila HSP27 and human alphab-crystallin expression-mediated increase in glutathione is essential for the protective activity of these proteins against tnfalpha-induced cell death. EMBO J. 1996, 15, 2695–2706. [Google Scholar] [PubMed]
- Roux, P.P.; Barker, P.A. Neurotrophin signaling through the p75 neurotrophin receptor. Prog. Neurobiol. 2002, 67, 203–233. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahimi Balaei, M.; Jiao, X.; Ashtari, N.; Afsharinezhad, P.; Ghavami, S.; Marzban, H. Cerebellar Expression of the Neurotrophin Receptor p75 in Naked-Ataxia Mutant Mouse. Int. J. Mol. Sci. 2016, 17, 115. https://doi.org/10.3390/ijms17010115
Rahimi Balaei M, Jiao X, Ashtari N, Afsharinezhad P, Ghavami S, Marzban H. Cerebellar Expression of the Neurotrophin Receptor p75 in Naked-Ataxia Mutant Mouse. International Journal of Molecular Sciences. 2016; 17(1):115. https://doi.org/10.3390/ijms17010115
Chicago/Turabian StyleRahimi Balaei, Maryam, Xiaodan Jiao, Niloufar Ashtari, Pegah Afsharinezhad, Saeid Ghavami, and Hassan Marzban. 2016. "Cerebellar Expression of the Neurotrophin Receptor p75 in Naked-Ataxia Mutant Mouse" International Journal of Molecular Sciences 17, no. 1: 115. https://doi.org/10.3390/ijms17010115
APA StyleRahimi Balaei, M., Jiao, X., Ashtari, N., Afsharinezhad, P., Ghavami, S., & Marzban, H. (2016). Cerebellar Expression of the Neurotrophin Receptor p75 in Naked-Ataxia Mutant Mouse. International Journal of Molecular Sciences, 17(1), 115. https://doi.org/10.3390/ijms17010115