
Inhibition of Hepatocyte Apoptosis: An Important Mechanism of Corn Peptides Attenuating Liver Injury Induced by Ethanol

Abstract
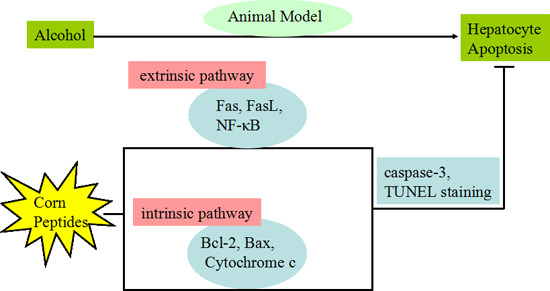
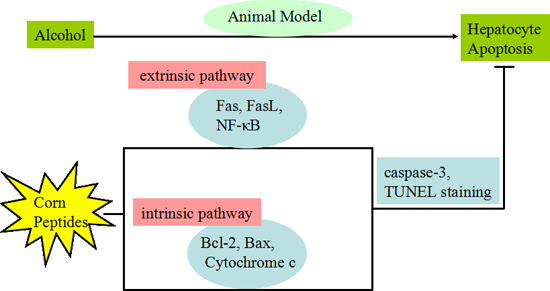
:
1. Introduction
2. Results
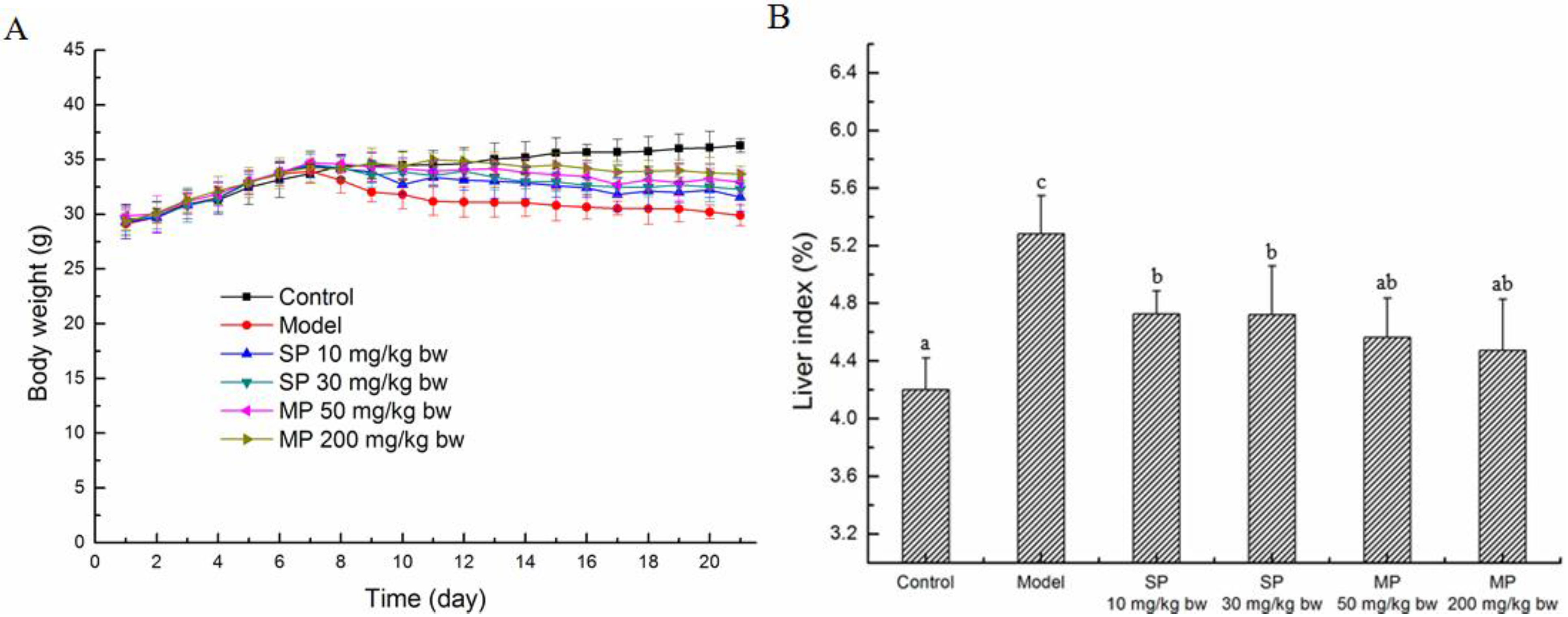
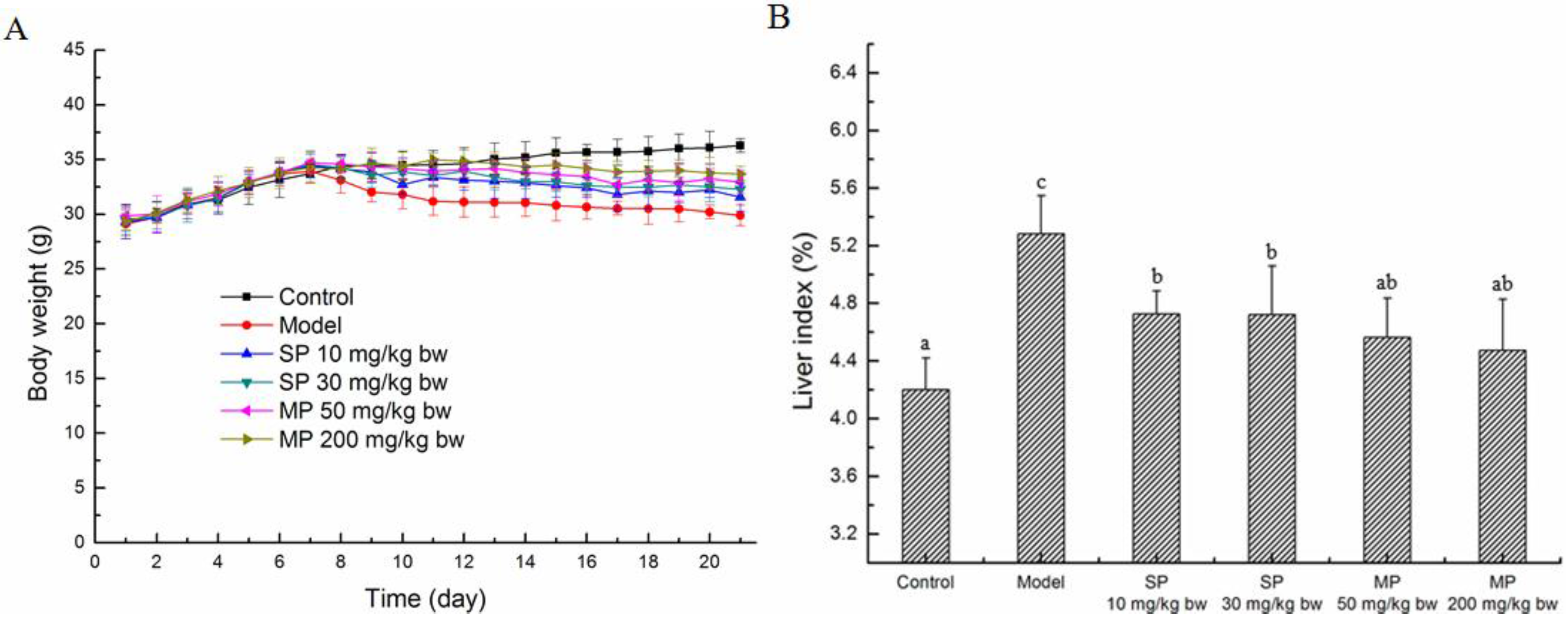
2.1. Effects of the Peptides on the Body Weights and Liver Index
2.2. Effects of the Peptides on the Activities of Serum Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST), the Levels of Hepatic Malonaldehyde (MDA) and Glutathione (GSH)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | ALT (IU/l) | AST (IU/l) | MDA (nmol/mg pro) | GSH (μmol/g pro) |
|---|---|---|---|---|
| Control | 7.77 ± 3.52 a | 10.66 ± 2.03 a | 2.51 ± 0.25 a | 2.85 ± 0.35 b |
| Model | 12.75 ± 4.40 b | 17.21 ± 8.97 b | 5.08 ± 0.45 d | 2.27 ± 0.16 a |
| SP 10 mg/kg bw | 10.45 ± 3.52 ab | 15.21 ± 2.77 ab | 3.47 ± 0.34 c | 3.09 ± 0.45 bc |
| SP 30 mg/kg bw | 9.56 ± 2.22 ab | 12.42 ± 3.52 ab | 3.40 ± 0.32 c | 3.50 ± 0.39 c |
| MP 50 mg/kg bw | 8.93 ± 2.34 ab | 11.68 ± 2.05 a | 3.15 ± 0.40 bc | 3.31 ± 0.33 bc |
| MP 200 mg/kg bw | 8.53 ± 2.03 ab | 11.53 ± 1.54 a | 2.84 ± 0.44 ab | 3.54 ± 0.27 c |
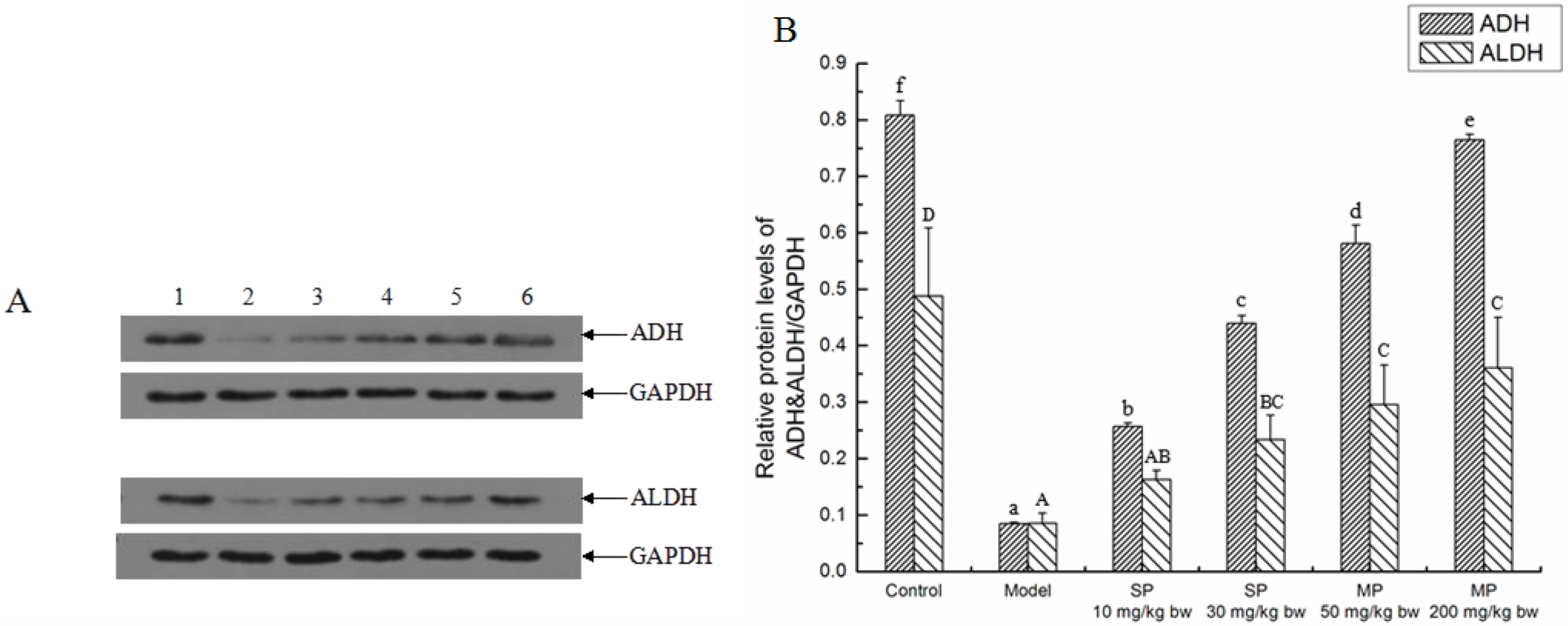
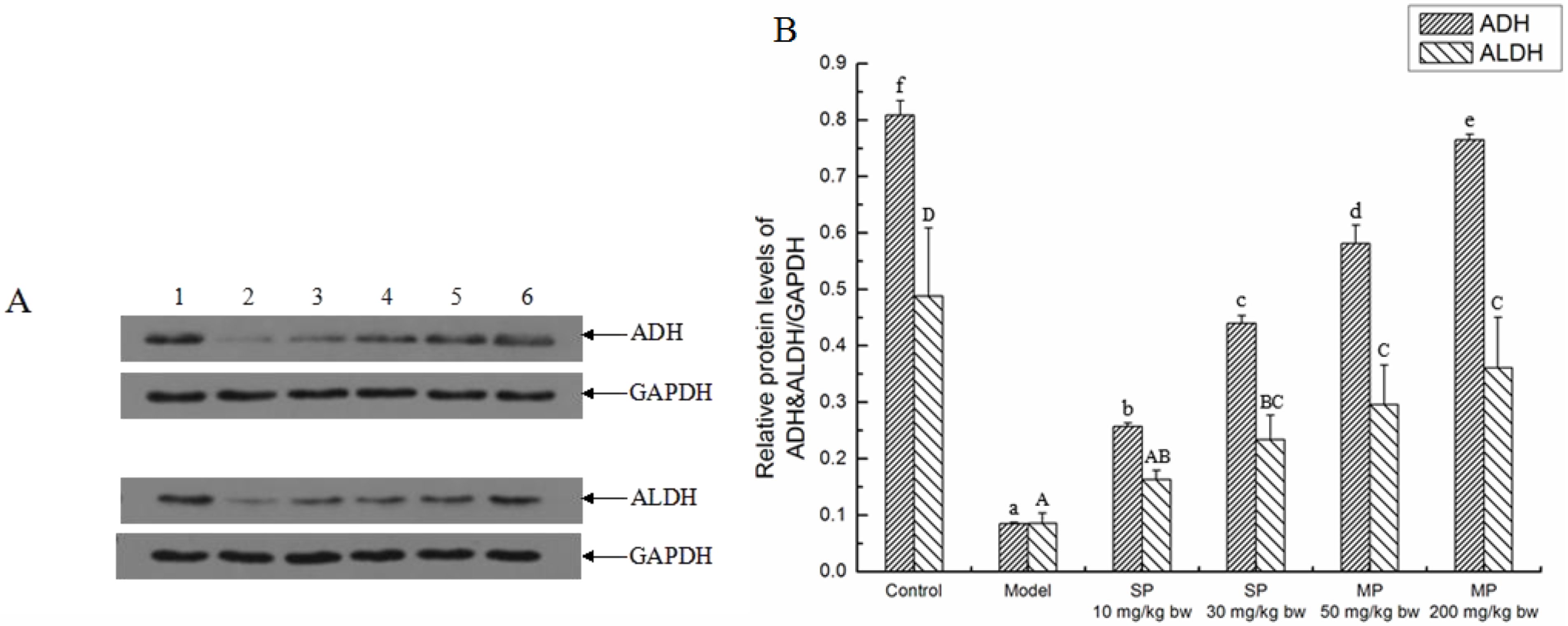
2.3. Effects of the Peptides on Key Ethanol Metabolizing Enzymes
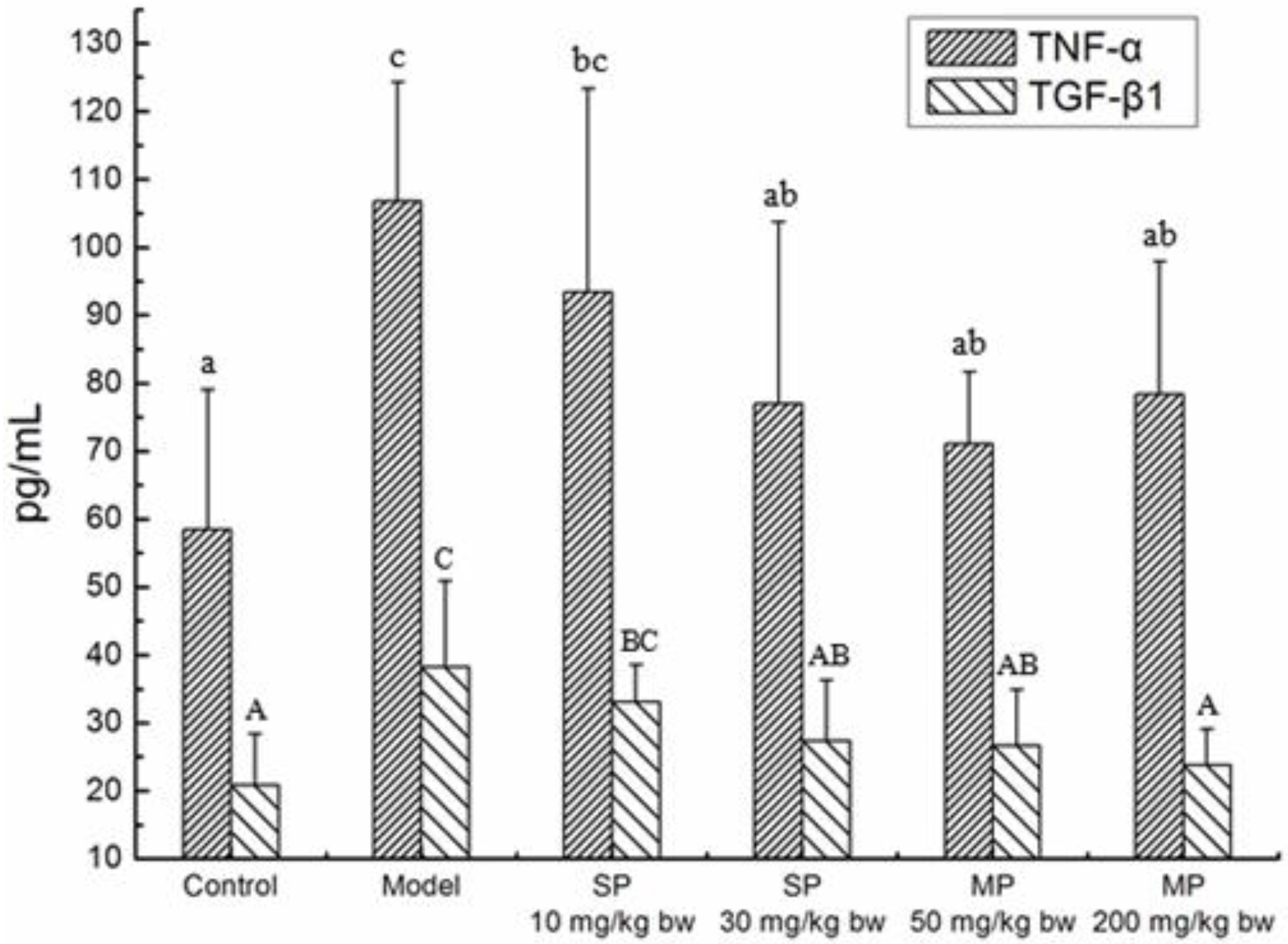
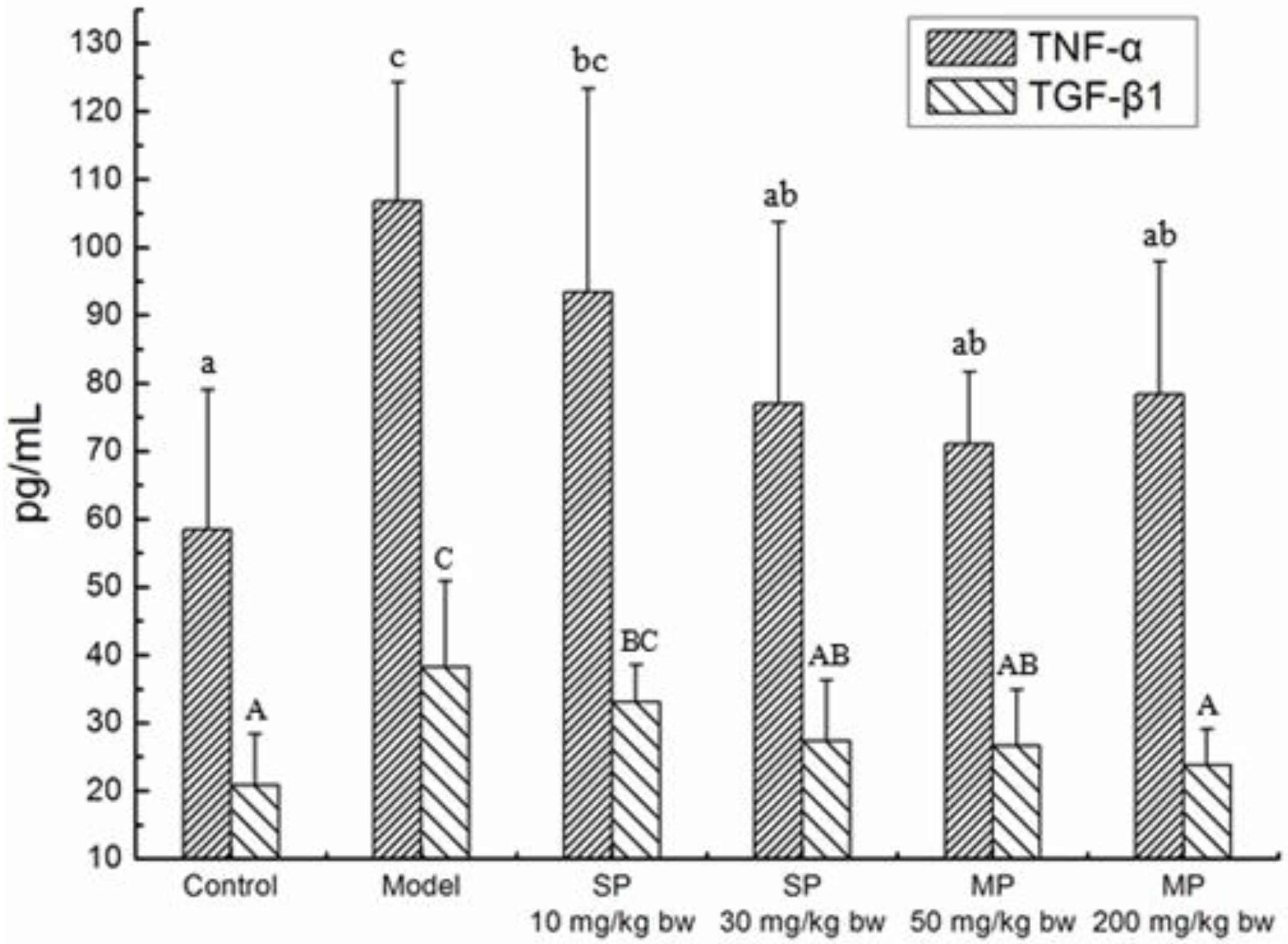
2.4. Effects of the Peptides on Cytokines TNF-α, TGF-β1
2.5. Effects of the Peptides on Apoptotic Pathway in Ethanol-Treated Mice
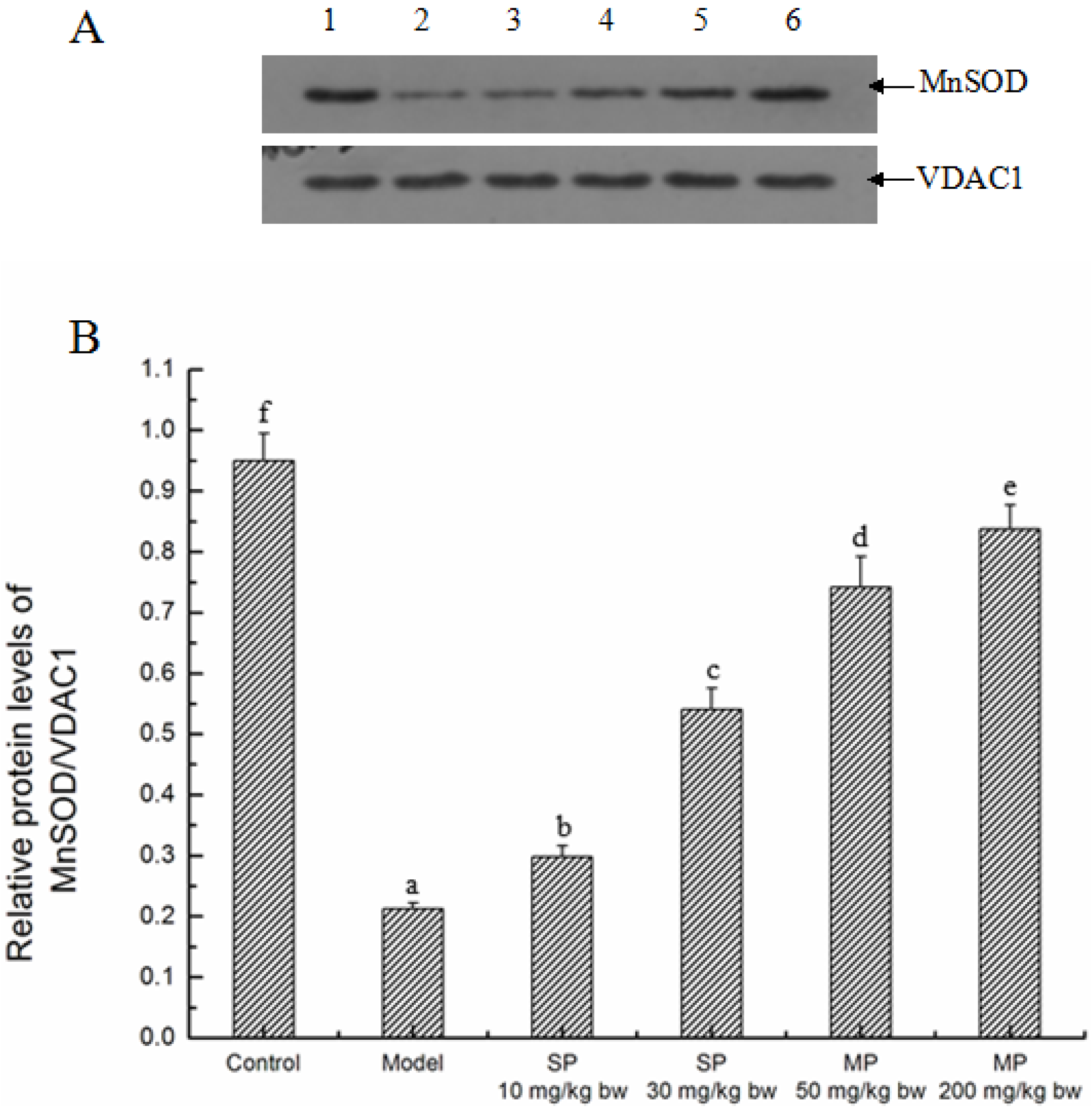
2.5.1. Effects of the Peptides on MnSOD
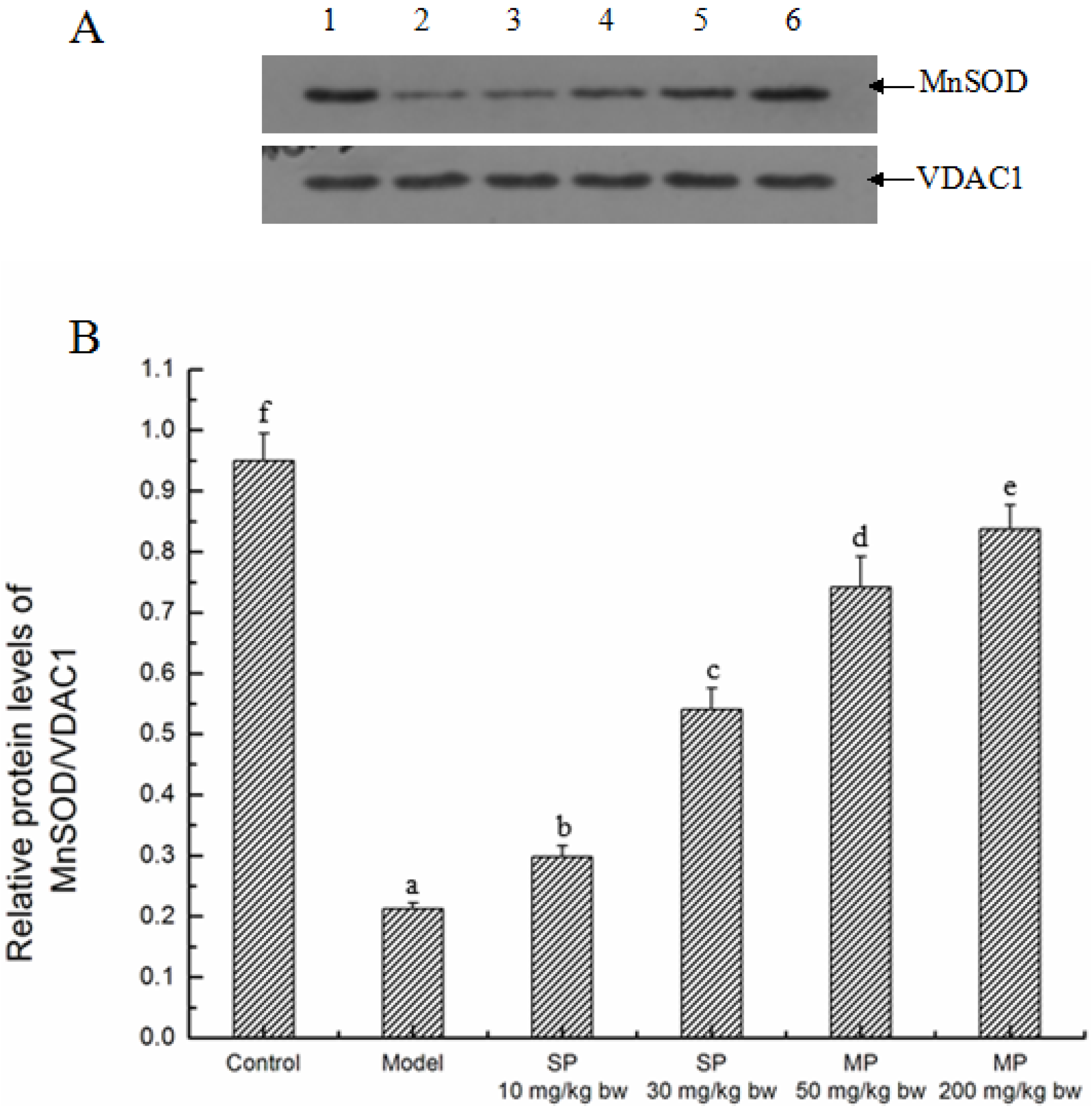
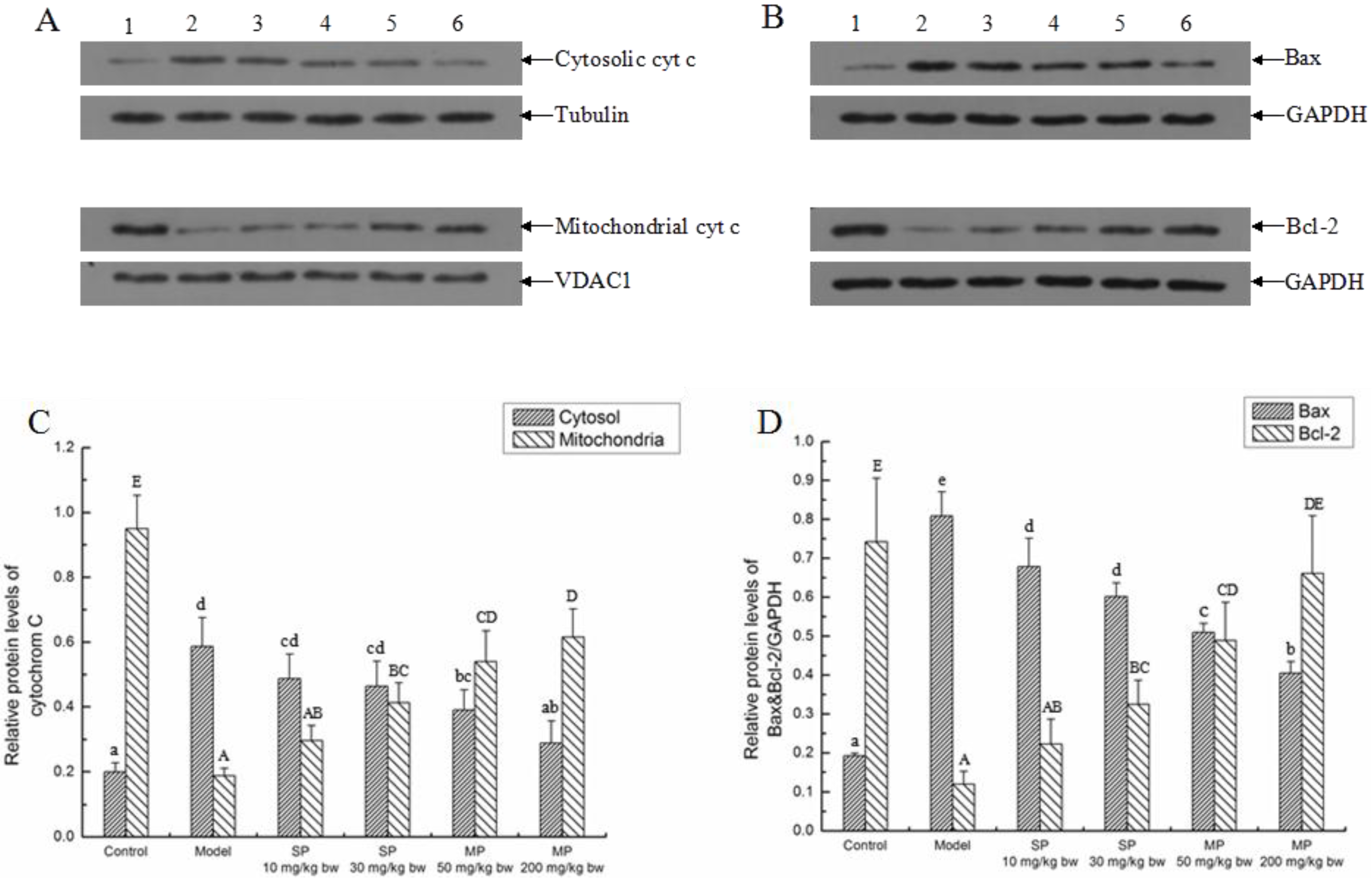
2.5.2. Effects of the Peptides on Intrinsic Pathway
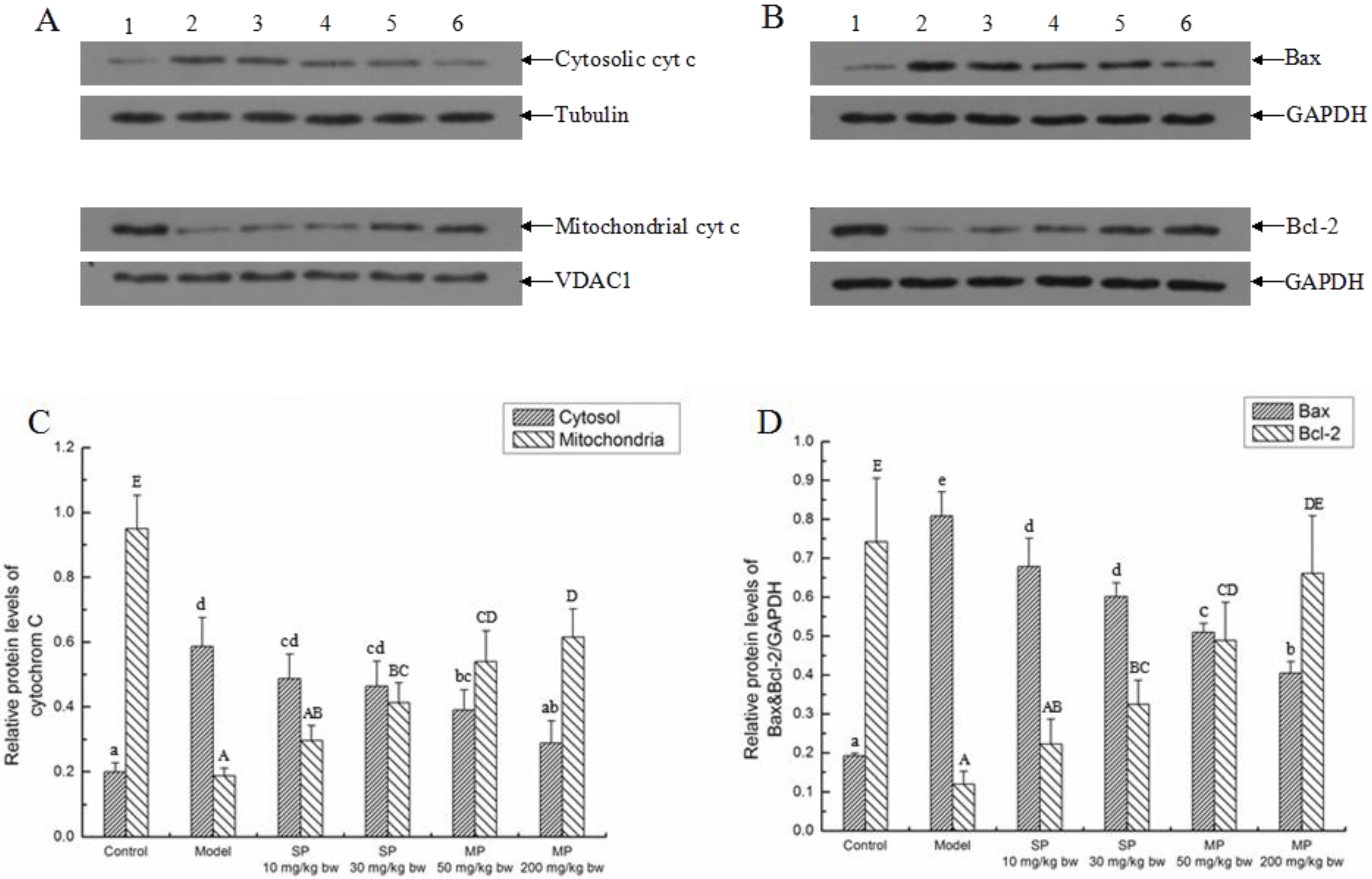
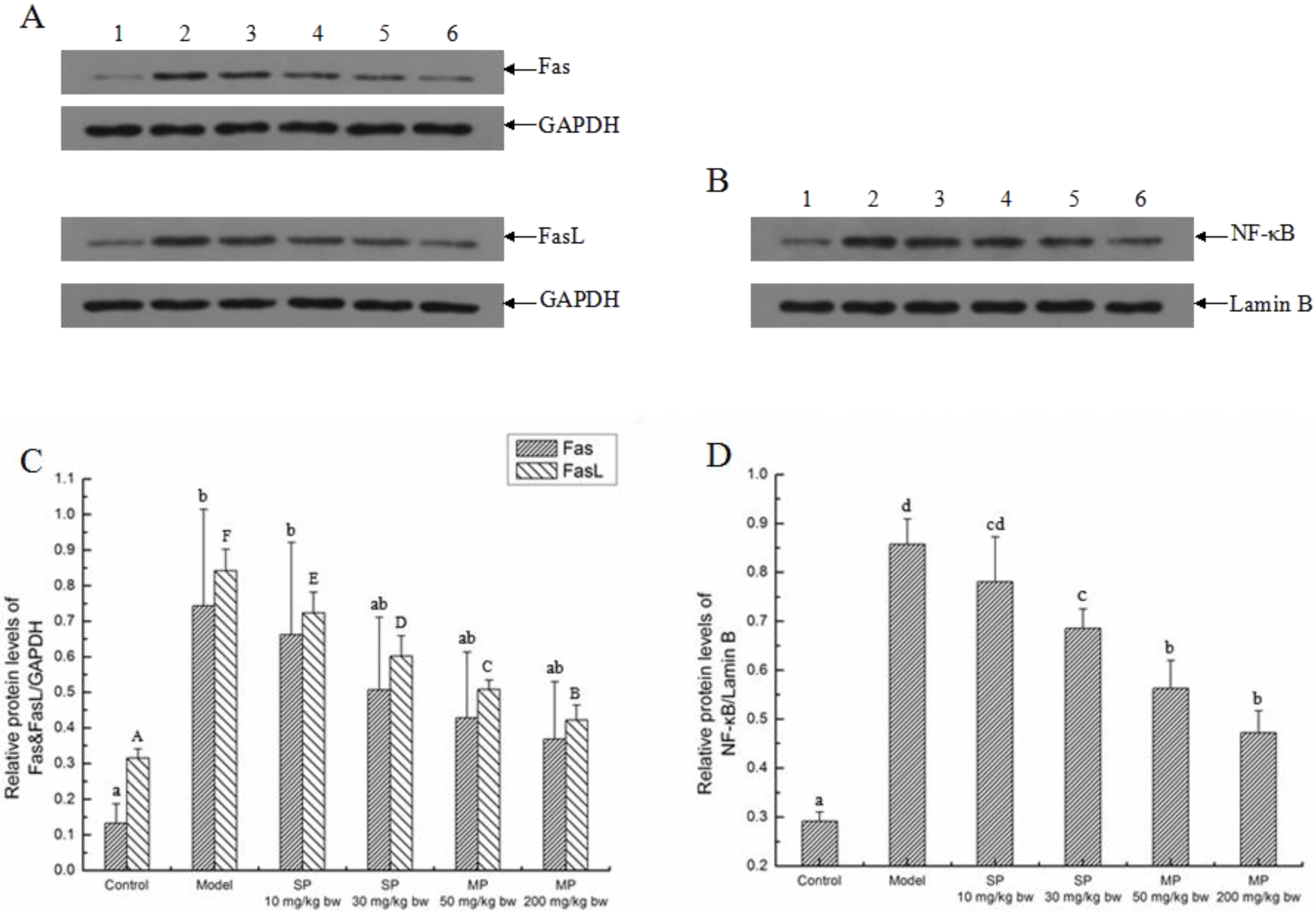
2.5.3. Effects of the Peptides on the Extrinsic Pathway
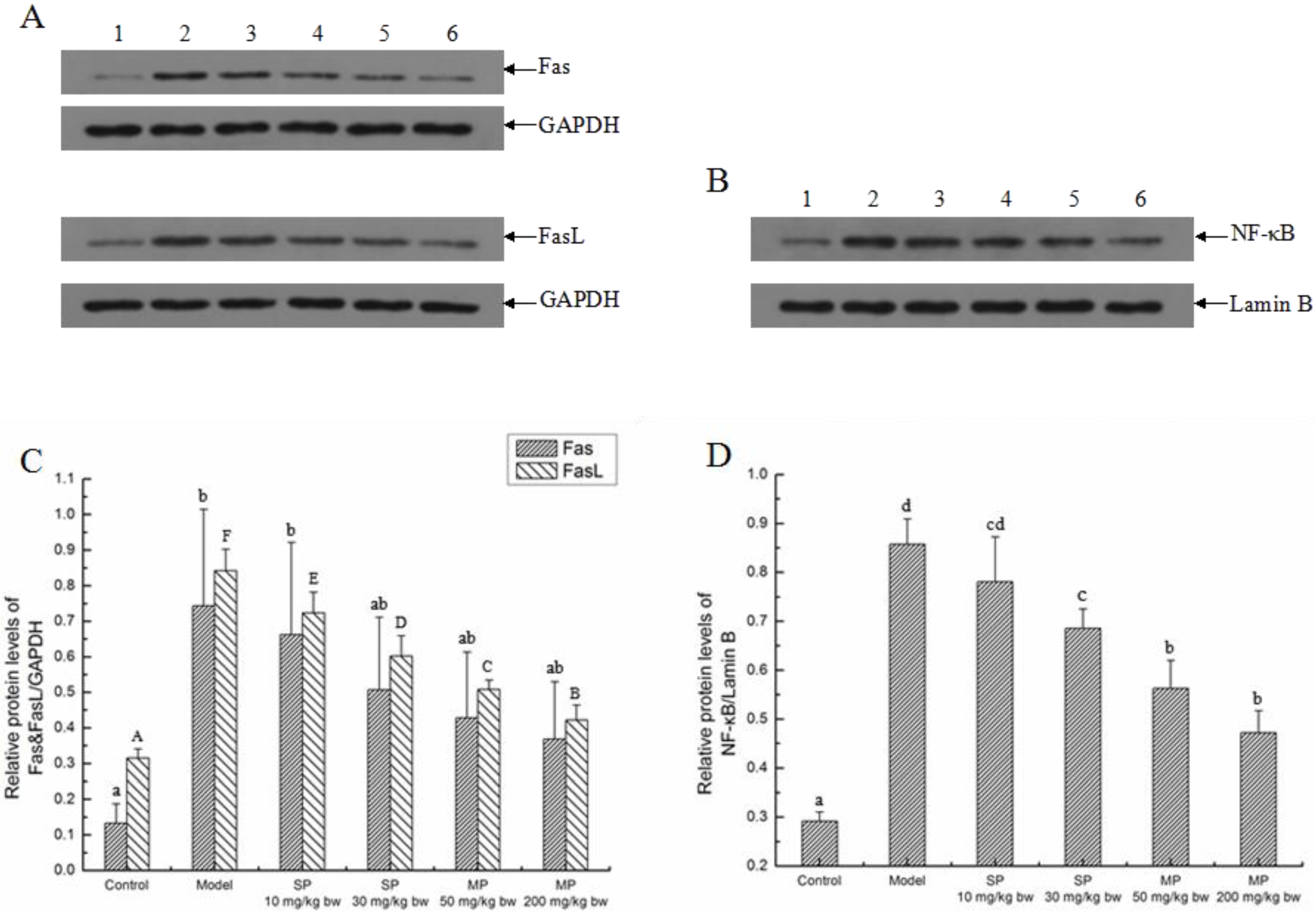
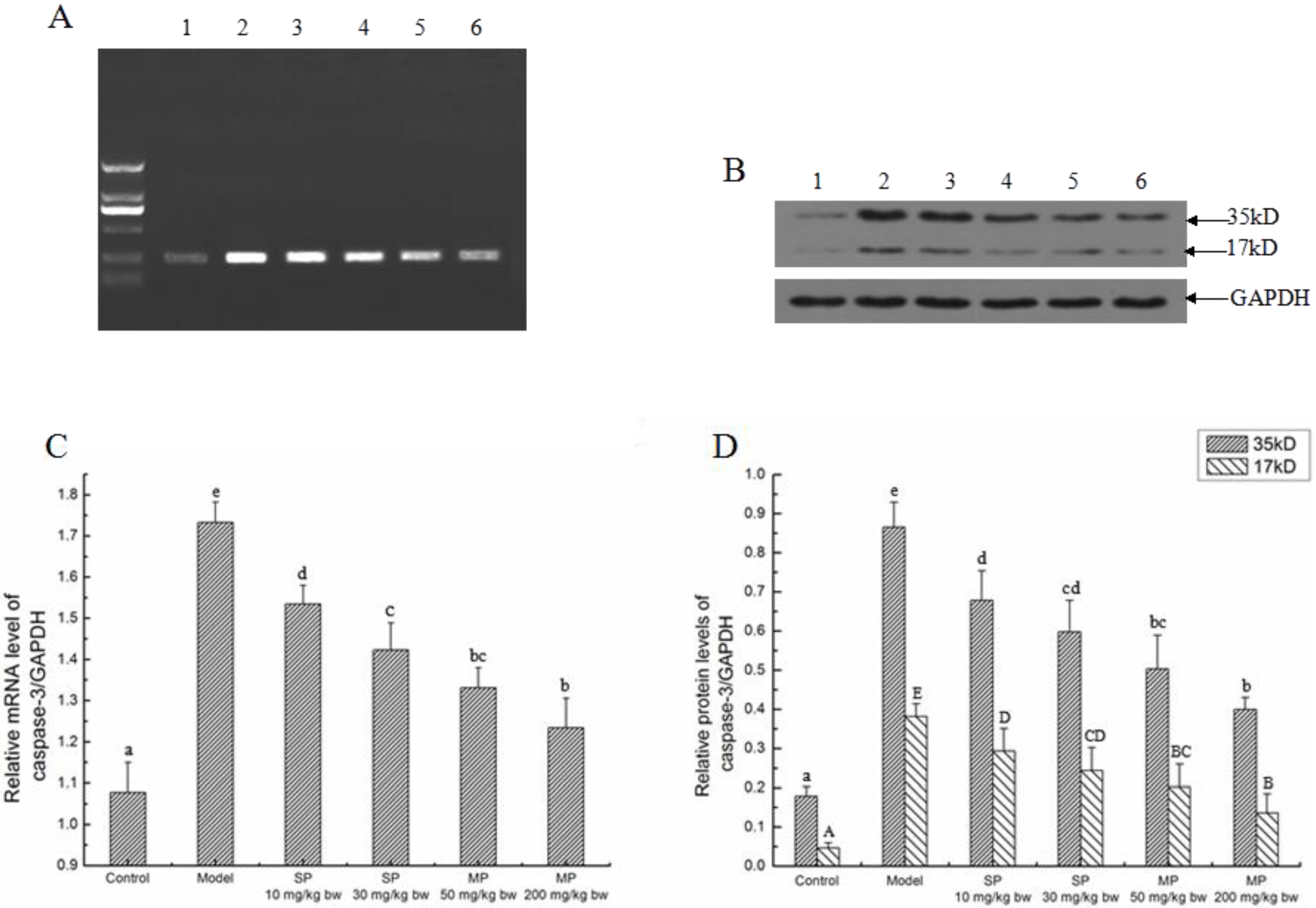
2.5.4. The Relative Expression Level of Caspase-3
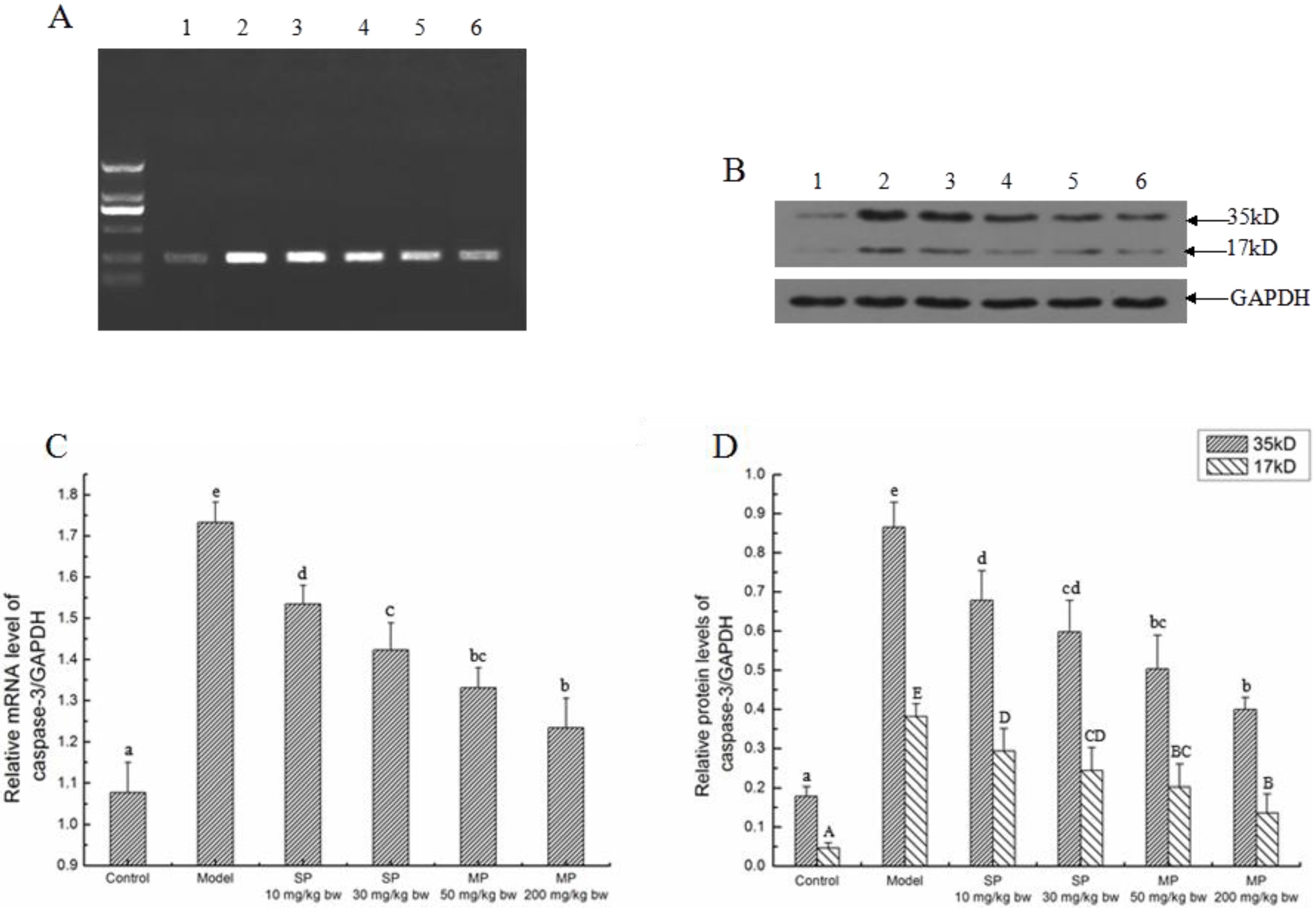
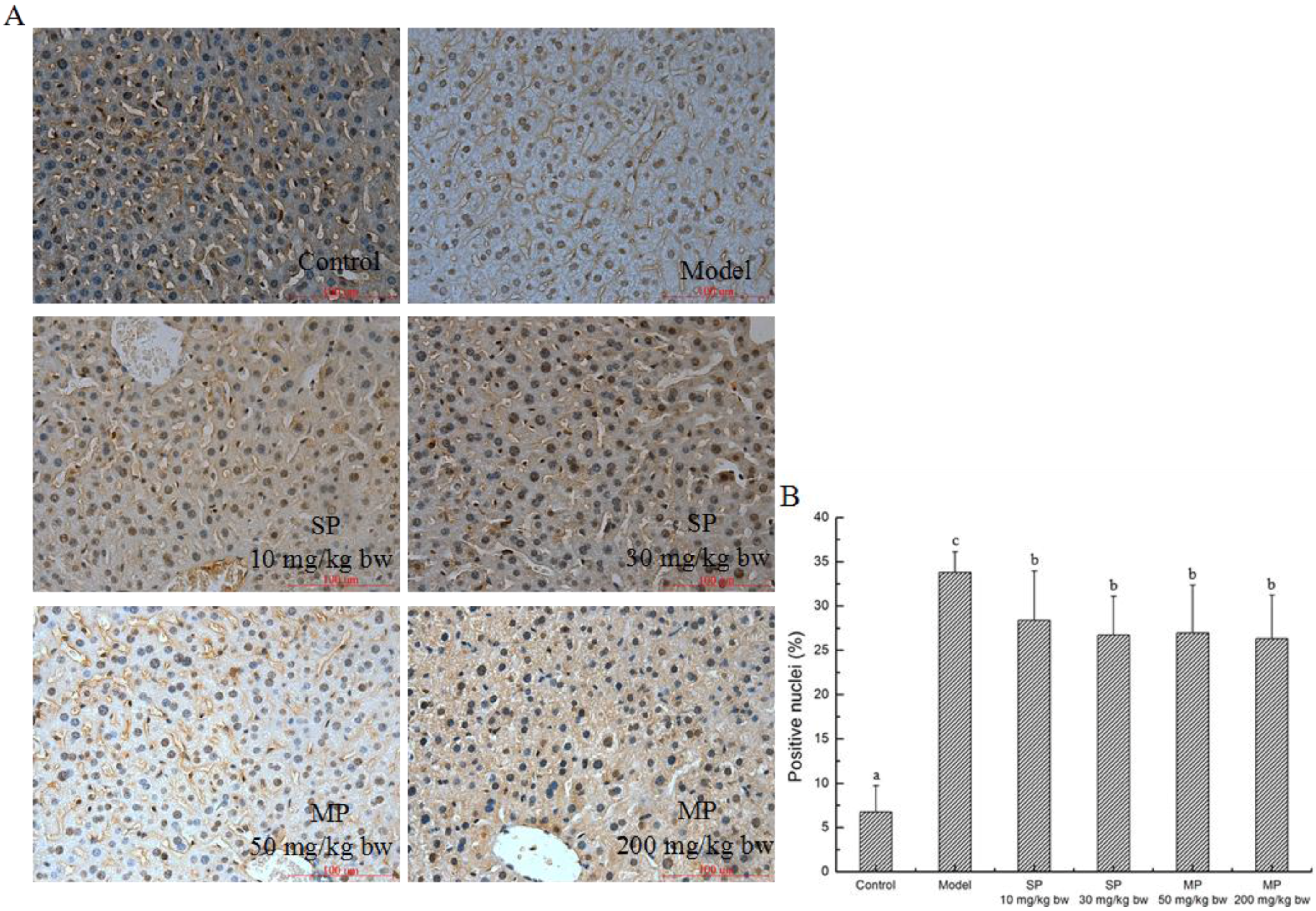
2.5.5. Effect of Peptides on Apoptosis
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of Concentrated Corn Protein and Corn Peptides
4.3. Animal and Experimental Design
4.4. Biochemical Parameters of Blood and Liver Analysis
4.5. Cytokines Analysis
4.6. Western Blot Analysis
4.7. Real-Time (RT) PCR Analysis
4.8. Transferase dUTP Nick End Labeling (TUNEL) Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Pari, L.; KarthiKesan, K. Protective role of caffeic acid against alcohol-induced biochemical changes in rats. Fundam. Clin. Pharmacol. 2007, 21, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Nanji, A.A. Apoptosis and alcoholic liver disease. Semin. Liver Dis. 1998, 18, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Yacoub, L.K.; Fogt, F.; Griniuviene, B.; Nanji, A.A. Apoptosis and Bcl-2 Protein Expression in Experimental Alcoholic Liver Disease in the Rat. Alcohol Clin. Exp. Res. 1995, 19, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Ziol, M.; Tepper, M.; Lohez, M.; Arcangeli, G.; Ganne, N.; Christidis, C.; Trinchet, J.C.; Beaugrand, M.; Guillet, J.G.; Guettier, C. Clinical and biological relevance of hepatocyte apoptosis in alcoholic hepatitis. J. Hepatol. 2001, 34, 254–260. [Google Scholar] [CrossRef]
- Sprick, M.R.; Walczak, H. The interplay between the Bcl-2 family and death receptor-mediated apoptosis. Biochim. Biophys. Acta 2004, 1644, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.F.; Xing, L.; Cui, P.F.; Wang, F.Z.; Xie, R.L.; Zhang, J.L.; Zhang, M.; He, Y.J.; Lyu, J.Y.; Qiao, J.B.; et al. Mitochondria apoptosis pathway synergistically activated by hierarchical targeted nanoparticles co-delivering siRNA and lonidamine. Biomaterials 2015, 61, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.W.; Zhang, J.; Yang, W.H.; Wang, B.; Wang, J.W. Glaucocalyxin A induces apoptosis in human leukemia HL-60 cells through mitochondria-mediated death pathway. Toxicol. In Vitro 2011, 25, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Raychaudhuri, S.; Skommer, J.; Henty, K.; Birch, N.; Brittain, T. Neuroglobin protects nerve cells from apoptosis by inhibiting the intrinsic pathway of cell death. Apoptosis 2010, 15, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, H.; Adachi, M.; Miura, S.; Gores, GJ.; Ishii, H. The mitochondrial permeability transition contributes to acute ethanol-induced apoptosis in rat hepatocytes. Hepatology 2001, 34, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Miñana, J.B.; Gómez-Cambronero, L.; Lloret, A.; Pallardó, F.V.; del Olmo, J.; Escudero, A.; Rodrigo, J.M.; Pellíin, A.; Viña, J.R.; Viña, J.; et al. Mitochondrial oxidative stress and CD95 ligand: A dual mechanism for hepatocyte apoptosis in chronic alcoholism. Hepatology 2002, 35, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.H.; Sun, J.; He, H.; Dong, H.W.; Li, J.T. Antihypertensive effect of corn peptides, produced by a continuous production in enzymatic membrane reactor, in spontaneously hypertensive rats. Food Chem. 2011, 128, 968–973. [Google Scholar] [CrossRef]
- Zhuang, H.; Tang, N.; Yuan, Y. Purification and identification of antioxidant peptides from corn gluten meal. J. Funct. Foods 2013, 5, 1810–1821. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Wang, Y.; Zhang, Y.; Liu, J. Isolation and characterisation of in vitro and cellular free radical scavenging peptides from corn peptide fractions. Molecules 2015, 20, 3221–3237. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.C.; Li, J.T.; He, H.; Huang, W.H.; Zhang, W.J. Ultrafiltration preparation of potent bioactive corn peptide as alcohol metabolism stimulator in vivo, and study on its mechanism of action. J. Food Biochem. 2013, 37, 161–167. [Google Scholar] [CrossRef]
- Kongo-Dia-Moukala, J.U.; Zhang, H.; Irakoze, P.C. In vitro binding capacity of bile acids by defatted corn protein hydrolysate. Int. J. Mol. Sci. 2011, 12, 1066–1080. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.J. Study on Preparation, Antialcoholism and Hepatic-Protection Mechanism of Corn Peptides. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, June 2006. [Google Scholar]
- Guo, H.; Sun, J.; He, H.; Yu, G.C.; Du, J. Antihepatotoxic effect of corn peptides against Bacillus Calmette-Guerin/lipopolysaccharide-induced liver injury in mice. Food Chem. Toxicol. 2009, 47, 2431–2435. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.C.; Lv, J.; He, H.; Huang, W.; Han, Y. Hepatoprotective effects of corn peptides against carbon tetrachloride-induced liver injury in mice. J. Food Biochem. 2012, 36, 458–464. [Google Scholar] [CrossRef]
- Lv, J.; Nie, Z.K.; Zhang, J.L.; Liu, F.Y.; Wang, Z.Z.; Ma, Z.L.; He, H. Corn peptides protect against thioacetamide-induced hepatic fibrosis in Rats. J. Med. Food 2013, 16, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, J.; Li, Y. Corn oligopeptides protect against early alcoholic liver injury in rats. Food Chem. Toxicol. 2012, 50, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Pan, X.C.; Zhang, S.X.; Wang, W.X.; Cai, M.Y.; Li, Y.R.; Yang, F.; Guo, H.W. Protective effect of corn peptides against alcoholic liver injury in men with chronic alcohol consumption: A randomized double-blind placebo-controlled study. Lipids Health Dis. 2014, 13, 192. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Cederbaum, I. Alcohol and oxidative liver injury. Hepatology 2006, 43, 63–74. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, C.; Colell, A.; Morales, A.; Kaplowitz, N.; Fernández-Checa, J.C. Role of oxidative stress generated from the mitochondrial electron transport chain and mitochondrial glutathione status in loss of mitochondrial function and activation of transcription factor nuclear factor-κB: Studies with isolated mitochondria and rat hepatocytes. Mol. Pharmacol. 1995, 48, 825–834. [Google Scholar] [PubMed]
- Neuman, M.G. Apoptosis in diseases of the liver. Crit. Rev. Clin. Lab. Sci. 2001, 38, 109–166. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ye, Y.; Kang, J.; Yao, X.; Zhang, Y.; Jiang, W.; Gao, M.; Dai, Y.; Xin, Y.; Wang, Q.; Yin, Z.; Luo, L. l-Theanine prevents alcoholic liver injury through enhancing the antioxidant capability of hepatocytes. Food Chem. Toxicol. 2012, 50, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Ge, N.; Liang, H.; Liu, Y.; Ma, A.G.; Han, L. Protective effect of Aplysin on hepatic injury in ethanol-treated rats. Food Chem. Toxicol. 2013, 62, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.L.; Zhang, W.J.; Yu, G.C.; He, H.; Zhang, Y. The primary structure identification of a corn peptide facilitating alcohol metabolism by HPLC-MS/MS. Peptides 2012, 37, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Lemon, P.W.; Nagle, F.J. Effects of exercise on protein and amino acid metabolism. Med. Sci. Sport Exerc. 1980, 13, 141–149. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Takada, M.; Nozaki, O.; Ito, M.; Furukawa, Y. Preparation of corn peptide from corn gluten meal and its administration effect on alcohol metabolism in stroke-prone spontaneously hypertensive rats. J. Nutr. Sci. Vitaminol. 1996, 42, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Dickman, M.B. Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Cederbaum, A.I. CYP2E1 and oxidative liver injury by alcohol. Free Radic. Biol. Med. 2008, 44, 723–738. [Google Scholar] [CrossRef] [PubMed]
- Je, J.Y.; Cha, J.Y.; Cho, Y.S.; Ahn, H.Y.; Lee, J.H.; Cho, Y.S.; Ahn, C.B. Hepatoprotective effect of peptic hydrolysate from salmon pectoral fin protein byproducts on ethanol-induced oxidative stress in Sprague-Dawley rats. Food Res. Int. 2013, 51, 648–653. [Google Scholar] [CrossRef]
- Louvet, A.; Mathurin, P. Alcoholic liver disease: Mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Mu, T.; Liu, X.; Zhang, M.; Chen, J. Purple sweet potato (Ipomoea batatas L.) anthocyanins: Preventive effect on acute and subacute alcoholic liver damage and dealcoholic effect. J. Agric. Food Chem. 2014, 62, 2364–2373. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.Y.; Ahn, H.Y.; Cho, Y.S.; Je, J.Y. Protective effect of cordycepin-enriched Cordyceps militaris on alcoholic hepatotoxicity in Sprague-Dawley rats. Food Chem. Toxicol. 2013, 60, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Danial, N.N.; Korsmeyer, S.J. Cell death: Critical control points. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef]
- Neuman, M.G.; Katz, G.G.; Malkiewicz, I.M.; Mathurin, P.; Tsukamoto, H.; Adachi, M.; Ishii, H.; Colell, A.; García-Ruiz, C.; Fernández-Checa, J.C.; et al. Alcoholic liver injury and apoptosis—Synopsis of the symposium held at ESBRA 2001: 8th Congress of the European Society for Biomedical Research on Alcoholism, Paris, September 16, 2001. Alcohol 2002, 28, 117–128. [Google Scholar] [CrossRef]
- Green, D.R.; Reed, J.C. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kim, C.N.; Yang, J.; Jemmerson, R.; Wang, X. Induction of apoptotic program in cell-free extracts: Requirement for dATP and cytochrome c. Cell 1996, 86, 147–157. [Google Scholar] [CrossRef]
- Korsmeyer, S.J. BCL-2 gene family and the regulation of programmed cell death. Cancer Res. 1999, 59, 1693–1700. [Google Scholar] [CrossRef]
- Yang, T.; Zhu, H.; Zhou, H.; Lin, Q.; Li, W.; Liu, J. Rice protein hydrolysate attenuates hydrogen peroxide induced apoptosis of myocardiocytes H9c2 through the Bcl-2/Bax pathway. Food Res. Int. 2012, 48, 736–741. [Google Scholar] [CrossRef]
- Maher, S.; Toomey, D.; Condron, C.; Bouchier-Hayes, D. Activation-induced cell death: The controversial role of Fas and Fas ligand in immune privilege and tumour counterattack. Immunol. Cell Biol. 2002, 80, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Chinnaiyan, A.M.; O’Rourke, K.; Tewari, M.; Dixit, V.M. FADD, a novel death domain-containing protein, interacts with the death domain of Fas and initiates apoptosis. Cell 1995, 81, 505–512. [Google Scholar] [CrossRef]
- Kucharczak, J.; Simmons, M.J.; Fan, Y.J.; Gélinas, C. To be, or not to be: NF-κB is the answer-role of Rel/NF-κB in the regulation of apoptosis. Oncogene 2003, 22, 8961–8982. [Google Scholar] [CrossRef] [PubMed]
- Diessenbacher, P.; Hupe, M.; Sprick, M.R.; Kerstan, A.; Geserick, P.; Haas, T.L.; Wachter, T.; Neumann, M.; Walczak, H.; Silke, J.; et al. NF-κB inhibition reveals differential mechanisms of TNF versus TRAIL-induced apoptosis upstream or at the level of caspase-8 activation independent of cIAP2. J. Investig. Dermatol. 2008, 22, 1134–1147. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Abdalla, M.; Li, Y.; Bai, Y.S. NF-κB mediates the induction of Fas receptor and Fas ligand by microcystin-LR in HepG2 cells. Mol. Cell. Biochem. 2011, 352, 209–219. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Hou, T.; Shi, W.; Liu, W.; He, H. Inhibition of Hepatocyte Apoptosis: An Important Mechanism of Corn Peptides Attenuating Liver Injury Induced by Ethanol. Int. J. Mol. Sci. 2015, 16, 22062-22080. https://doi.org/10.3390/ijms160922062
Ma Z, Hou T, Shi W, Liu W, He H. Inhibition of Hepatocyte Apoptosis: An Important Mechanism of Corn Peptides Attenuating Liver Injury Induced by Ethanol. International Journal of Molecular Sciences. 2015; 16(9):22062-22080. https://doi.org/10.3390/ijms160922062
Chicago/Turabian StyleMa, Zhili, Tao Hou, Wen Shi, Weiwei Liu, and Hui He. 2015. "Inhibition of Hepatocyte Apoptosis: An Important Mechanism of Corn Peptides Attenuating Liver Injury Induced by Ethanol" International Journal of Molecular Sciences 16, no. 9: 22062-22080. https://doi.org/10.3390/ijms160922062
APA StyleMa, Z., Hou, T., Shi, W., Liu, W., & He, H. (2015). Inhibition of Hepatocyte Apoptosis: An Important Mechanism of Corn Peptides Attenuating Liver Injury Induced by Ethanol. International Journal of Molecular Sciences, 16(9), 22062-22080. https://doi.org/10.3390/ijms160922062