
Identification of Conserved and Novel MicroRNAs during Tail Regeneration in the Mexican Axolotl

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Identification of Conserved MicroRNAs in Regenerating Tail Tissue
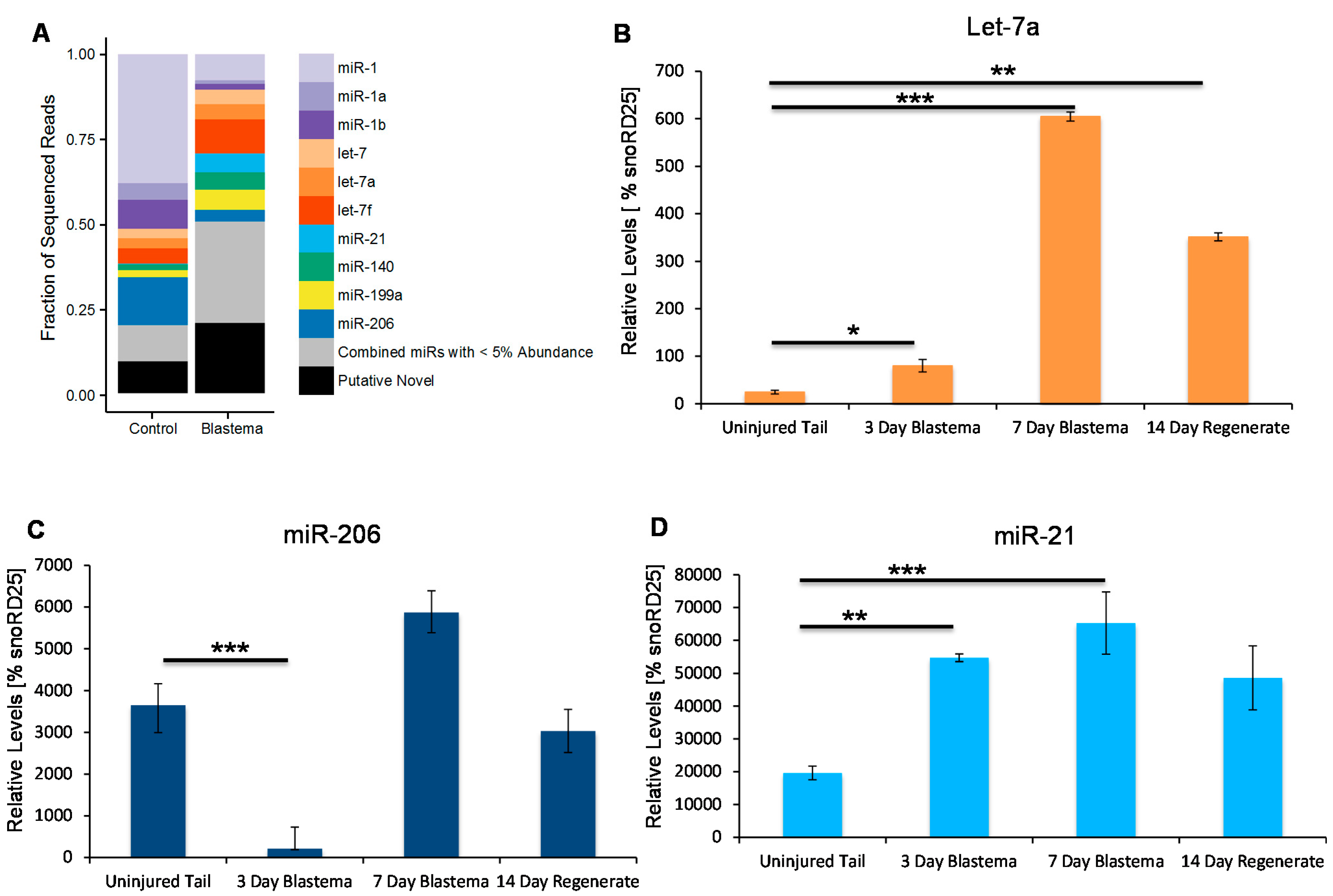
2.2. Identification of Putative Novel MicroRNAs Enriched in the Tail Blastema
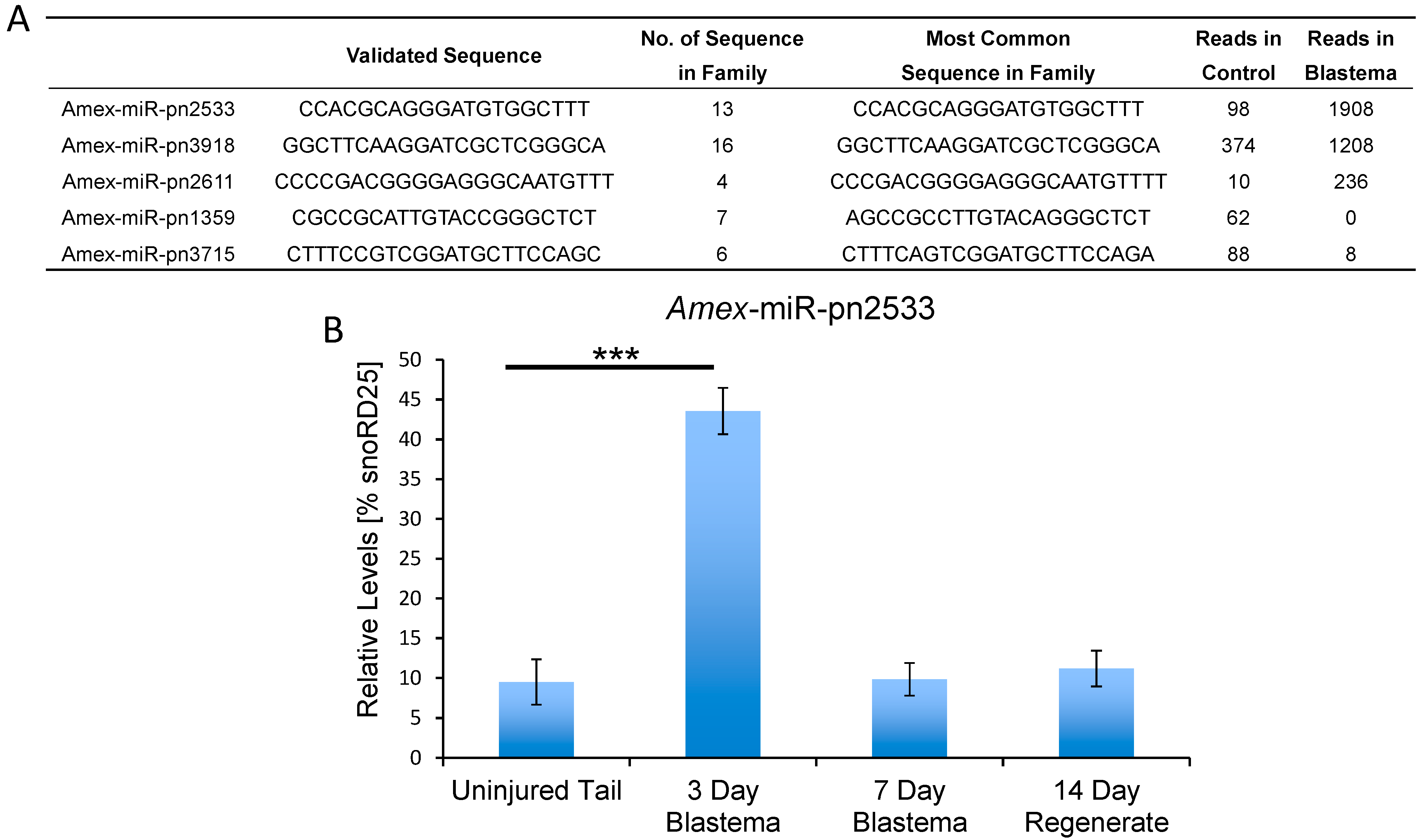

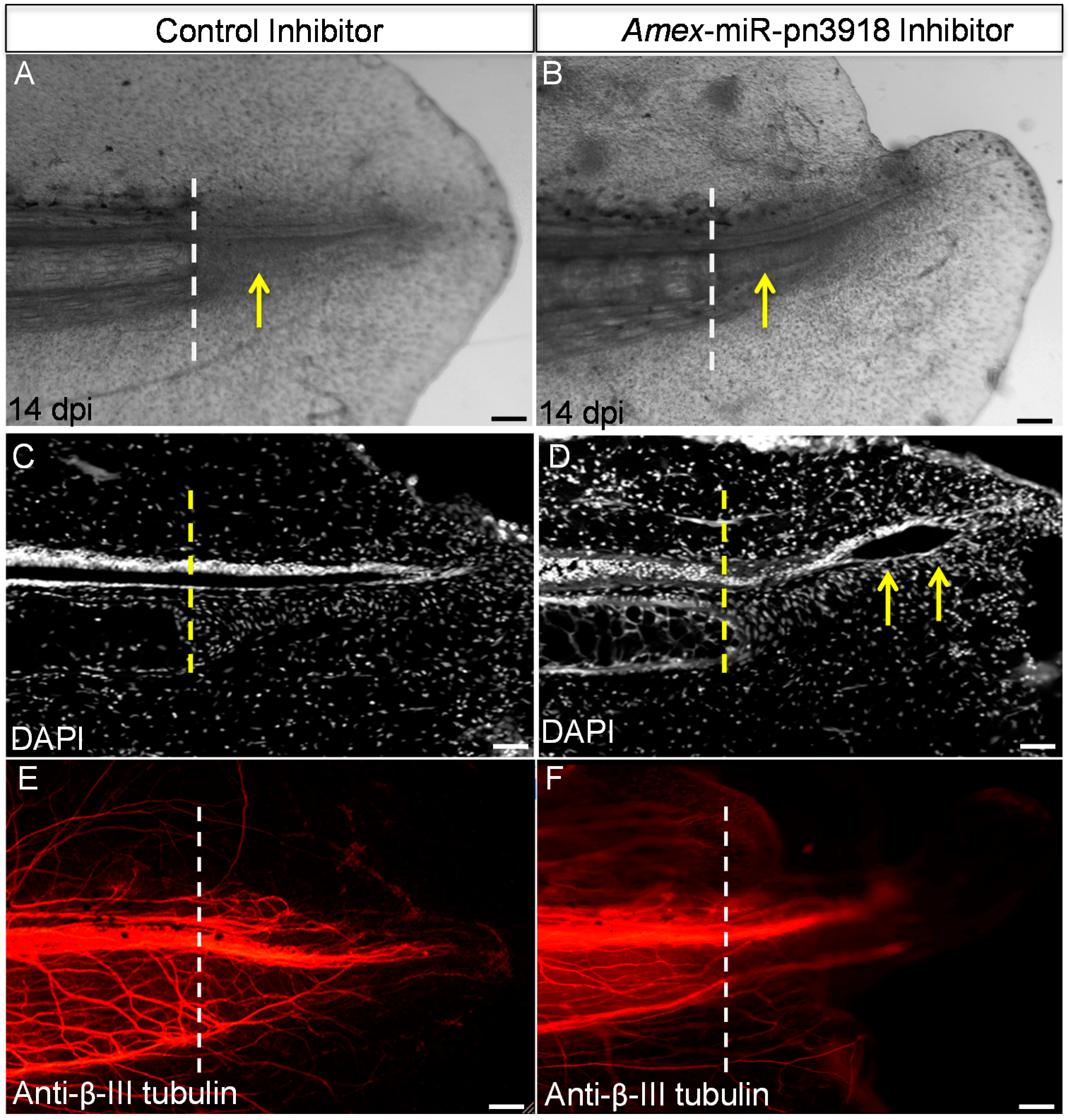
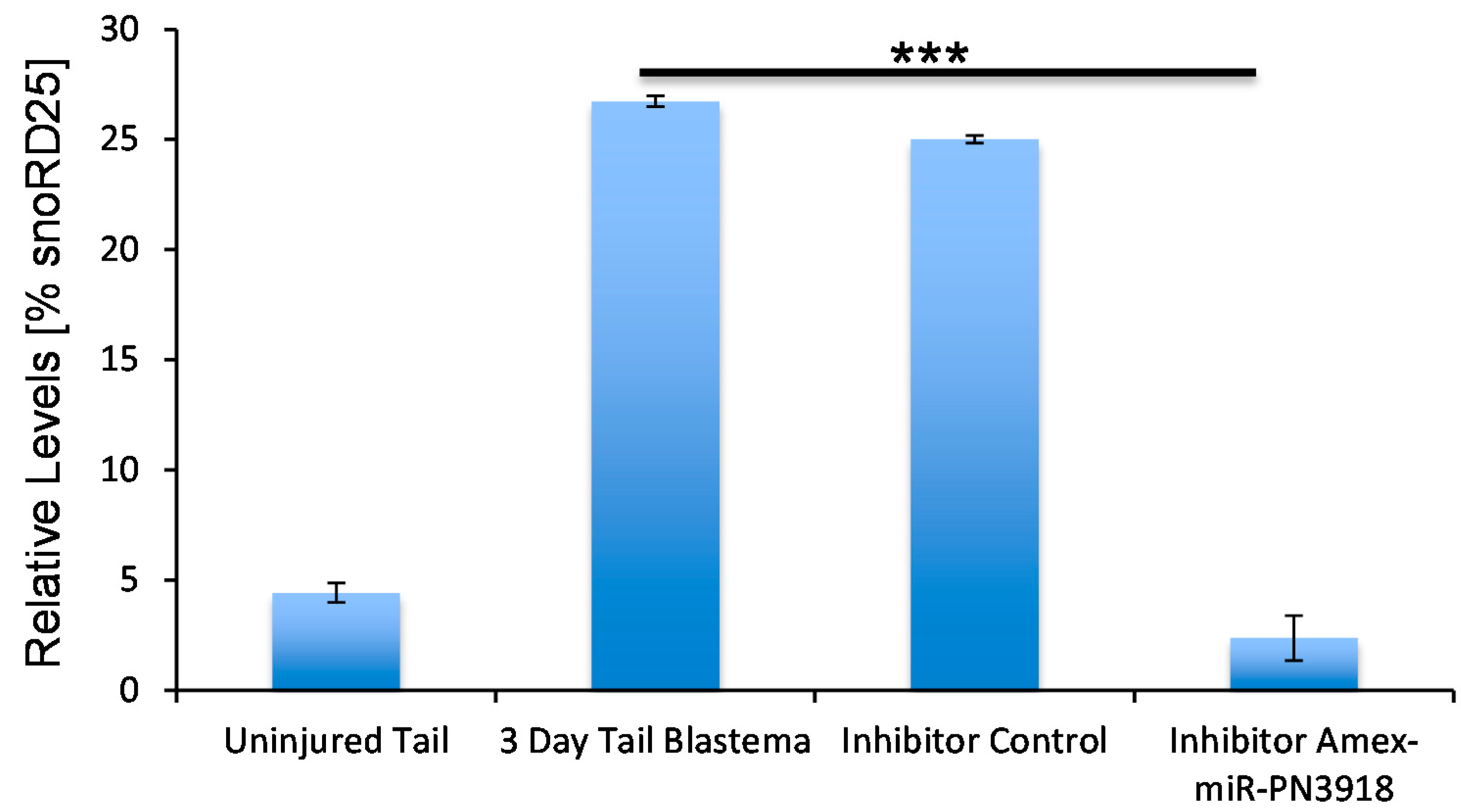
3. Discussion
4. Materials and Methods
4.1. Animal Handling
4.2. Deep Sequencing of Axolotl MicroRNAs
4.3. Quantitative Real-Time PCR
4.4. In Situ Hybridization
4.5. MicroRNA Inhibitor Injection
4.6. Whole-Mount Staining
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Agata, K.; Saito, Y.; Nakajima, E. Unifying principles of regeneration I: Epimorphosis versus morphallaxis. Dev. Growth Differ. 2007, 49, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Brockes, J.P. Amphibian limb regeneration: Rebuilding a complex structure. Science 1997, 276, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Brockes, J.P.; Kumar, A. Appendage regeneration in adult vertebrates and implications for regenerative medicine. Science 2005, 310, 1919–1923. [Google Scholar] [CrossRef] [PubMed]
- Brockes, J.P.; Kumar, A.; Velloso, C.P. Regeneration as an evolutionary variable. J. Anat. 2001, 199, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Call, M.K.; Grogg, M.W.; Tsonis, P.A. Eye on regeneration. Anat. Rec. B New Anat. 2005, 287, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, E.A. Spinal cord regeneration: A phenomenon unique to urodeles? Int. J. Dev. Biol. 1996, 40, 823–831. [Google Scholar] [PubMed]
- Diaz Quiroz, J.F.; Echeverri, K. Spinal cord regeneration: Where fish, frogs and salamanders lead the way, can we follow? Biochem. J. 2013, 451, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Frobisch, N.B.; Shubin, N.H. Salamander limb development: Integrating genes, morphology, and fossils. Dev. Dyn. 2011, 240, 1087–1099. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Bryant, S.V. Molecular mechanisms in the control of limb regeneration: The role of homeobox genes. Int. J. Dev. Biol. 1996, 40, 797–805. [Google Scholar] [PubMed]
- Gardiner, D.M.; Carlson, M.R.; Roy, S. Towards a functional analysis of limb regeneration. Semin. Cell Dev. Biol. 1999, 10, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Godwin, J.W.; Brockes, J.P. Regeneration, tissue injury and the immune response. J. Anat. 2006, 209, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Godwin, J.W.; Rosenthal, N. Scar-free wound healing and regeneration in amphibians: Immunological influences on regenerative success. Differentiation 2014, 87, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Kawasumi, A.; Sagawa, N.; Hayashi, S.; Yokoyama, H.; Tamura, K. Wound healing in mammals and amphibians: Toward limb regeneration in mammals. Curr. Top. Microbiol. Immunol. 2013, 367, 33–49. [Google Scholar] [PubMed]
- Maden, M. Regeneration: Every clot has a thrombin lining. Curr. Biol. 2003, 13, R517–R518. [Google Scholar] [CrossRef]
- Mescher, A.L. The cellular basis of limb regeneration in urodeles. Int. J. Dev. Biol. 1996, 40, 785–795. [Google Scholar] [PubMed]
- Murawala, P.; Tanaka, E.M.; Currie, J.D. Regeneration: The ultimate example of wound healing. Semin. Cell Dev. Biol. 2012, 23, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Nacu, E.; Tanaka, E.M. Limb regeneration: A new development? Annu. Rev. Cell Dev. Biol. 2011, 27, 409–440. [Google Scholar] [CrossRef] [PubMed]
- Nye, H.L.D.; Cameron, J.A.; Chernoff, E.A.G.; Stocum, D.L. Regeneration of the urodele limb: A review. Dev. Dyn. 2003, 226, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Odelberg, S.J. Cellular plasticity in vertebrate regeneration. Anat. Rec. B New Anat. 2005, 287, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Gatien, S. Regeneration in axolotls: A model to aim for! Exp. Gerontol. 2008, 43, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Seifert, A.W.; Maden, M. New insights into vertebrate skin regeneration. Int. Rev. Cell Mol. Biol. 2014, 310, 129–169. [Google Scholar] [PubMed]
- Tsonis, P.A. Regeneration of the lens in amphibians. Results Probl. Cell Differ. 2000, 31, 179–196. [Google Scholar] [PubMed]
- Zhu, W.; Pao, G.M.; Satoh, A.; Cummings, G.; Monaghan, J.R.; Harkins, T.T.; Bryant, S.V.; Voss, S.R.; Gardiner, D.M.; Hunter, T. Activation of germline-specific genes is required for limb regeneration in the Mexican axolotl. Dev. Biol. 2012, 370, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D. MicoRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Bartel, D.P.; Chen, C.Z. Micromanagers of gene expression: The potentially widespread influence of metazoan microRNAs. Nat. Rev. Genet. 2004, 5, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Bicker, S.; Schratt, G. MicroRNAs: Tiny regulators of synapse function in development and disease. J. Cell. Mol. Med. 2008, 12, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- De Felici, M.; Farini, D.; Dolci, S. In or out stemness: Comparing growth factor signalling in mouse embryonic stem cells and primordial germ cells. Curr. Stem Cell Res. Ther. 2009, 4, 87–97. [Google Scholar] [CrossRef] [PubMed]
- De Pietri Tonelli, D.; Fei, J.F.; Nomura, T.; Osumi, N.; Heisenberg, C.P.; Huttner, W.B. Single-cell detection of microRNAs in developing vertebrate embryos after acute administration of a dual-fluorescence reporter/sensor plasmid. Biotechniques 2006, 41, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Farh, K.K.H.; Grimson, A.; Jan, C.; Lewis, B.P.; Johnston, W.K.; Lim, L.P.; Burge, C.B.; Bartel, D.P. The widespread impact of mammalian microRNAs on mRNA repression and evolution. Science 2005, 310, 1817–1821. [Google Scholar] [CrossRef] [PubMed]
- Giraldez, A.J.; Cinalli, R.M.; Glasner, M.E.; Enright, A.J.; Thomson, J.M.; Baskerville, S.; Hammond, S.M.; Bartel, D.P.; Schier, A.F. MicroRNAs regulate brain morphogenesis in zebrafish. Science 2005, 308, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Hagen, J.; Lai, E. MicroRNA control of cell-cell signaling during development and disease. Cell Cycle 2008, 15, 2327–2332. [Google Scholar] [CrossRef]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsle, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.P.; Glasner, M.E.; Yekta, S.; Burge, C.B.; Bartel, D.P. Vertebrate microRNA genes. Science 2003, 299, 1540. [Google Scholar] [CrossRef] [PubMed]
- Alteri, A.; de Vito, F.; Nessina, G.; Pompili, M.; Calconi, A.; Visca, P.; Mottolese, M.; Presutti, C.; Grossi, M. Cyclin D1 is a major target of miR-206 in cell differentiation and transformation. Cell Cycle 2013, 12, 3781–3790. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Wang, Y.; Li, Y.; Cui, L.; Zhao, Y.; Zhao, B.; Li, K. miR-206, a key modulator of skeletal muscle development and disease. Int. J. Biol. Sci. 2015, 11, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Maki, N.; Trinh, A.; Trask, H.W.; Gui, J.; Tomlinson, C.R.; Tsonis, P.A. miRNAs in newt lens regeneration: Specific control of proliferation and evidence for miRNA networking. PLoS ONE 2010, 5, e12058. [Google Scholar] [CrossRef] [PubMed]
- Rybak, A.; Fuchs, H.; Smirnova, L.; Brandt, C.; Pohl, E.E.; Nitsch, R.; Wulczyn, F.G. A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment. Nat. Cell Biol. 2008, 10, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Tsonis, P.A.; Call, M.K.; Grogg, M.W.; Sartor, M.A.; Taylor, R.R.; Forge, A.; Fyffe, R.; Goldenberg, R.; Cowper-Sal·lari, R.; Tomlinson, C.R. MicroRNAs and regeneration: Let-7 members as potential regulators of dedifferentiation in lens and inner ear hair cell regeneration of the adult newt. Biochem. Biophys. Res. Commun. 2007, 362, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Hong, W. The emerging role of the hippo pathway in cell contact inhibition, organ size control, and cancer development in mammals. Cancer Cell 2008, 13, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Yin, V.P.; Lepilina, A.; Smith, A.; Poss, K.D. Regulation of zebrafish heart regeneration by miR-133. Dev. Biol. 2012, 365, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Yin, V.P.; Thomson, J.M.; Thummel, R.; Hyde, D.R.; Hammond, S.M.; Poss, K.D. Fgf-dependent depletion of microRNA-133 promotes appendage regeneration in zebrafish. Genes Dev. 2008, 22, 728–733. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, K.; Harding, R.L.; Hyde, D.R.; Patton, J.G. miR-203 regulates progenitor cell proliferation during adult zebrafish retina regeneration. Dev. Biol. 2014, 392, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, K.; Harding, R.L.; Bailey, T.; Patton, J.G.; Hyde, D.R. Dynamic miRNA expression patterns during retinal regeneration in zebrafish: Reduced dicer or miRNA expression suppresses proliferation of Muller glia-derived neuronal progenitor cells. Dev. Dyn. 2014, 243, 1591–1605. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R.; Fausett, B.V.; Goldman, D. Ascl1a regulates Muller glia dedifferentiation and retinal regeneration through a Lin-28-dependent, let-7 microRNA signalling pathway. Nat. Cell Biol. 2010, 12, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, E.M.; Weidinger, G. Micromanaging regeneration. Genes Dev. 2008, 22, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Yin, V.P.; Poss, K.D. New regulators of vertebrate appendage regeneration. Curr. Opin. Genet. Dev. 2008, 18, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.M.; Gibbs, K.M.; Davila, J.; Campbell, N.; Sung, S.; Todorova, T.I.; Otsuka, S.; Sabaawy, H.E.; Hart, R.P.; Schachner, M. MicroRNA miR-133b is essential for functional recovery after spinal cord injury in adult zebrafish. Eur. J. Neurosci. 2011, 33, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Makarev, E.; Spence, J.R.; Del Rio-Tsonis, K.; Tsonis, P.A. Identification of microRNAs and other small RNAs from the adult newt eye. Mol. Vis. 2006, 12, 1386–1391. [Google Scholar] [PubMed]
- Witman, N.; Heigwer, J.; Thaler, B.; Lui, W.O.; Morrison, J.I. miR-128 regulates non-myocyte hyperplasia, deposition of extracellular matrix and Islet1 expression during newt cardiac regeneration. Dev. Biol. 2013, 383, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Holman, E.C.; Campbell, L.J.; Hines, J.; Crews, C.M. Microarray analysis of microRNA expression during axolotl limb regeneration. PLoS ONE 2012, 7, e41804. [Google Scholar] [CrossRef] [PubMed]
- Diaz Quiroz, J.F.; Tsai, E.; Coyle, M.; Sehm, T.; Echeverri, K. Precise control of miR-125b levels is required to create a regeneration-permissive environment after spinal cord injury: A cross-species comparison between salamander and rat. Dis. Model. Mech. 2014, 7, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Sehm, T.; Sachse, C.; Frenzel, C.; Echeverri, K. miR-196 is an essential early-stage regulator of tail regeneration, upstream of key spinal cord patterning events. Dev. Biol. 2009, 334, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Caygill, E.E.; Johnston, L.A. Temporal regulation of metamorphic processes in Drosophila by the let-7 and miR-125 heterochronic microRNAs. Curr. Biol. 2008, 18, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Schulman, B.R.M.; Esquela-Kerscher, A.; Slack, F.J. Reciprocal expression of lin-41 and the microRNAs let-7 and mir-125 during mouse embryogenesis. Dev. Dyn. 2005, 234, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Wulczyn, F.G.; Smirnova, L.; Rybak, A.; Brandt, C.; Kwidzinski, E.; Ninnemann, O.; Strehle, M.; Seiler, A.; Schumacher, A.; Nitsch, R. Post-transcriptional regulation of the let-7 microRNA during neural cell specification. FASEB J. 2007, 21, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, E.A.G.; Stocum, D.L.; Nye, H.L.D.; Cameron, J.A. Urodele spinal cord regeneration and related processes. Dev. Dyn. 2003, 226, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Boettger, T.; Wüst, S.; Nolte, H.; Braun, T. The miR-206/133b cluster is dispensable for development, survival and regeneration of skeletal muscle. Skelet. Muscle 2014, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Crescenzi, M. Levels matter: miR-206 and cyclin D1. Cell Cycle 2013, 12, 3715. [Google Scholar] [CrossRef] [PubMed]
- Georgantas, R.W.; Streicher, K.; Greenberg, S.A.; Greenlees, L.M.; Zhu, W.; Brohawn, P.Z.; Higgs, B.W.; Czapiga, M.; Morehouse, C.A.; Amato, A.; et al. Inhibition of myogenic microRNAs 1, 133, and 206 by inflammatory cytokines links inflammation and muscle degeneration in adult inflammatory myopathies. Arthritis Rheumatol. 2014, 66, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Nachtigall, P.G.; Dias, M.C.; Pinhal, D. Evolution and genomic organization of muscle microRNAs in fish genomes. BMC Evol. Biol. 2014, 14, 196. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.K.; Kumar, R.M.; Farkhondeh, M.; Baskerville, S.; Lodish, H.F. Myogenic factors that regulate expression of muscle-specific microRNAs. Proc. Natl. Acad. Sci. USA 2006, 103, 8721–8726. [Google Scholar] [CrossRef] [PubMed]
- Winbanks, C.E.; Beyer, C.; Hagg, A.; Qian, H.; Sepulveda, P.V.; Gregorevic, P. miR-206 represses hypertrophy of myogenic cells but not muscle fibers via inhibition of HDAC4. PLoS ONE 2013, 8, e73589. [Google Scholar] [CrossRef] [PubMed]
- Holtzer, S.W. The inductive activity of the spinal cord in urodele amphibian regeneration. J. Morphol. 1956, 99, 1–33. [Google Scholar] [CrossRef]
- Erickson, J.R.; Echeverri, K. In vivo modulation and quantification of microRNAs during axolotl tail regeneration. Methods Mol. Biol. 2015, 1290, 159–167. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gearhart, M.D.; Erickson, J.R.; Walsh, A.; Echeverri, K. Identification of Conserved and Novel MicroRNAs during Tail Regeneration in the Mexican Axolotl. Int. J. Mol. Sci. 2015, 16, 22046-22061. https://doi.org/10.3390/ijms160922046
Gearhart MD, Erickson JR, Walsh A, Echeverri K. Identification of Conserved and Novel MicroRNAs during Tail Regeneration in the Mexican Axolotl. International Journal of Molecular Sciences. 2015; 16(9):22046-22061. https://doi.org/10.3390/ijms160922046
Chicago/Turabian StyleGearhart, Micah D., Jami R. Erickson, Andrew Walsh, and Karen Echeverri. 2015. "Identification of Conserved and Novel MicroRNAs during Tail Regeneration in the Mexican Axolotl" International Journal of Molecular Sciences 16, no. 9: 22046-22061. https://doi.org/10.3390/ijms160922046
APA StyleGearhart, M. D., Erickson, J. R., Walsh, A., & Echeverri, K. (2015). Identification of Conserved and Novel MicroRNAs during Tail Regeneration in the Mexican Axolotl. International Journal of Molecular Sciences, 16(9), 22046-22061. https://doi.org/10.3390/ijms160922046