
Chlorella vulgaris Attenuates Dermatophagoides Farinae-Induced Atopic Dermatitis-Like Symptoms in NC/Nga Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
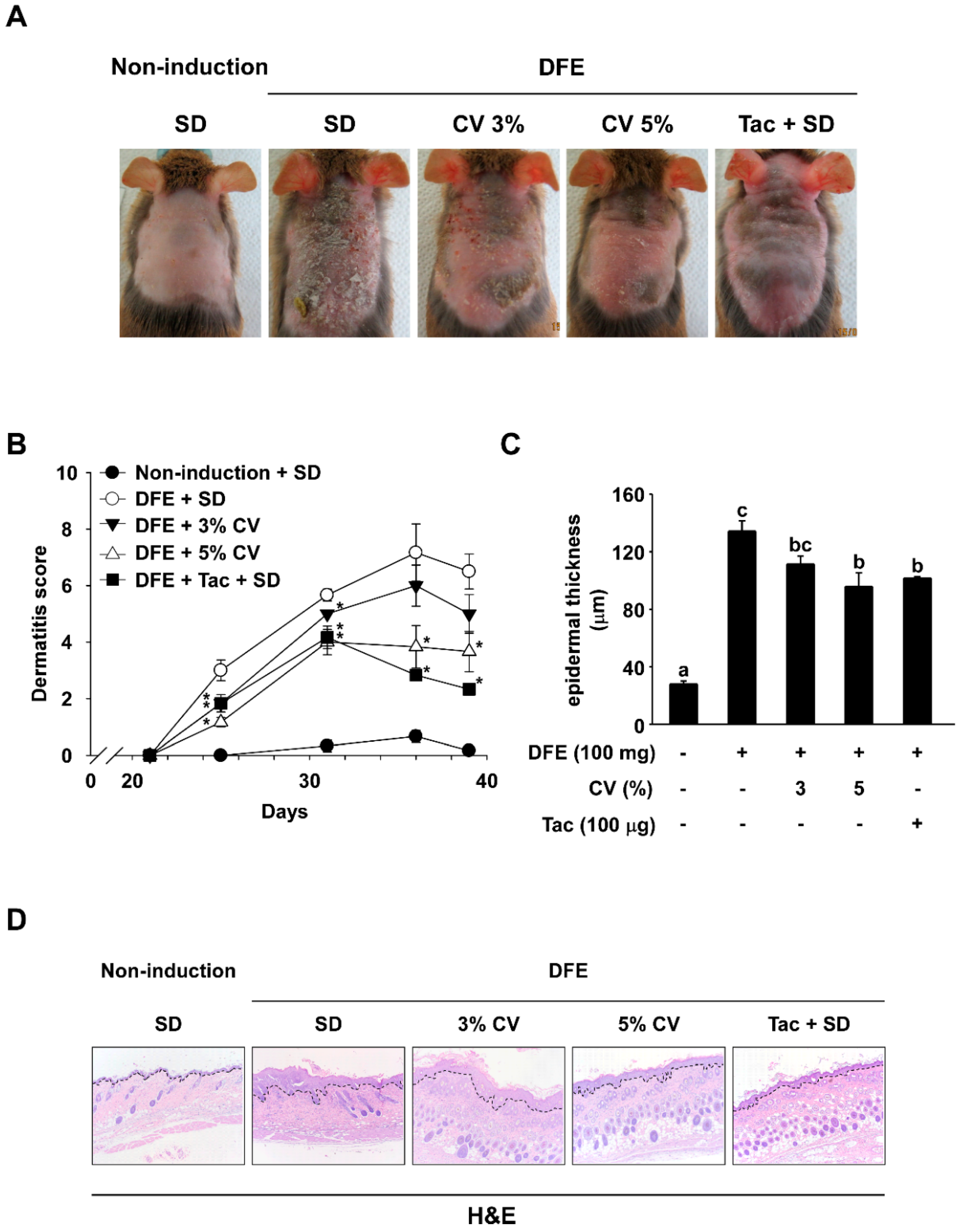
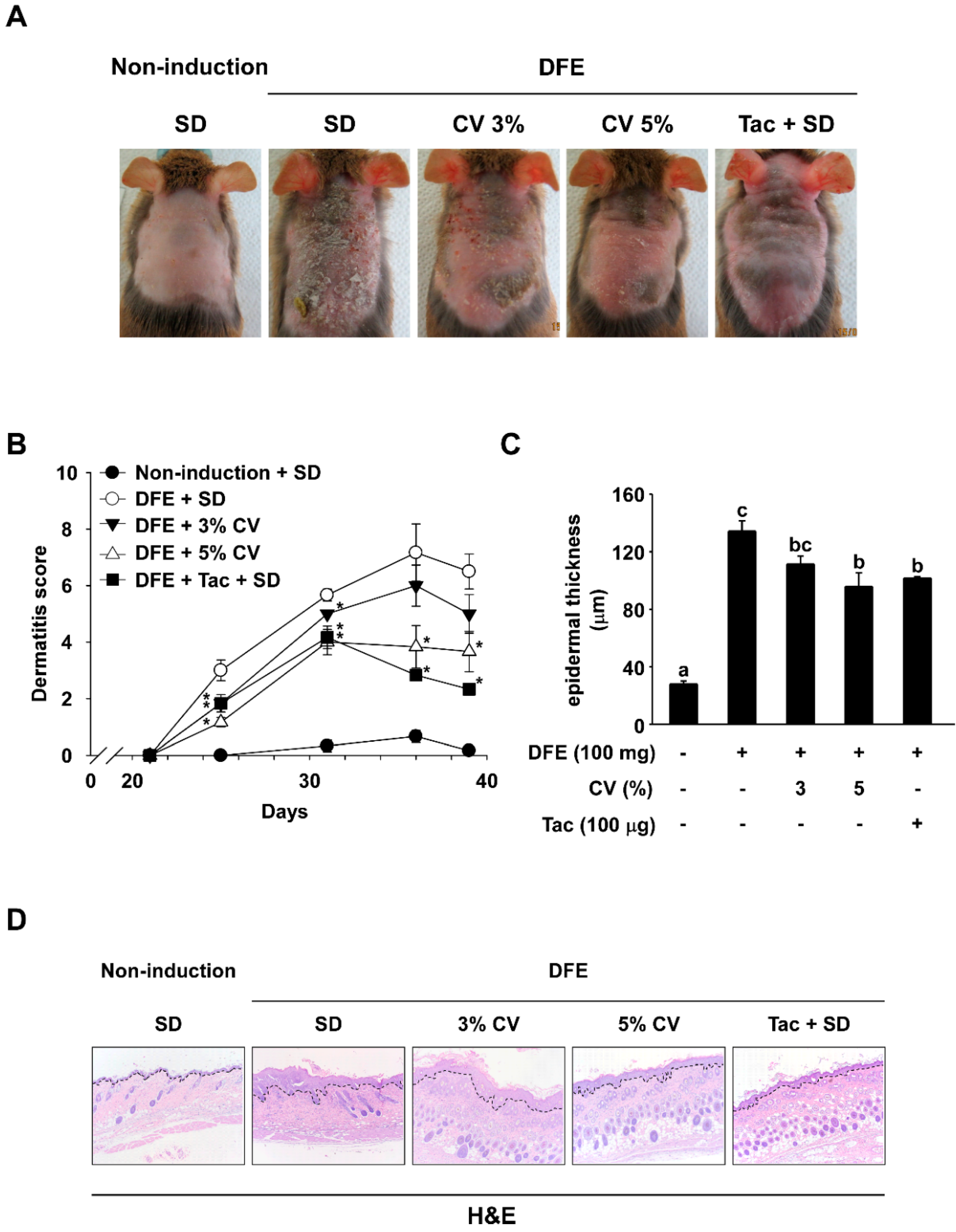
2.1. CV Supplementation Alleviates DFE-Induced AD-Like Skin Lesions in NC/Nga Mice
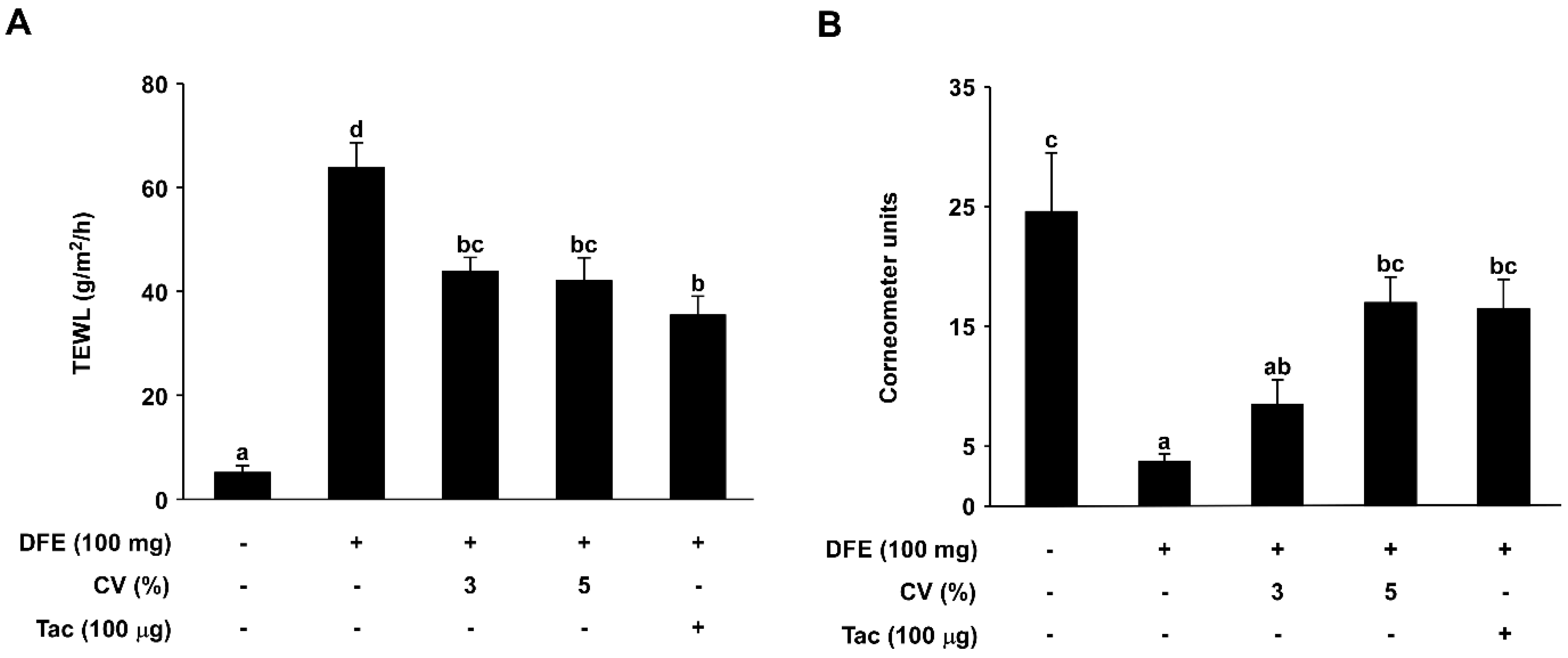
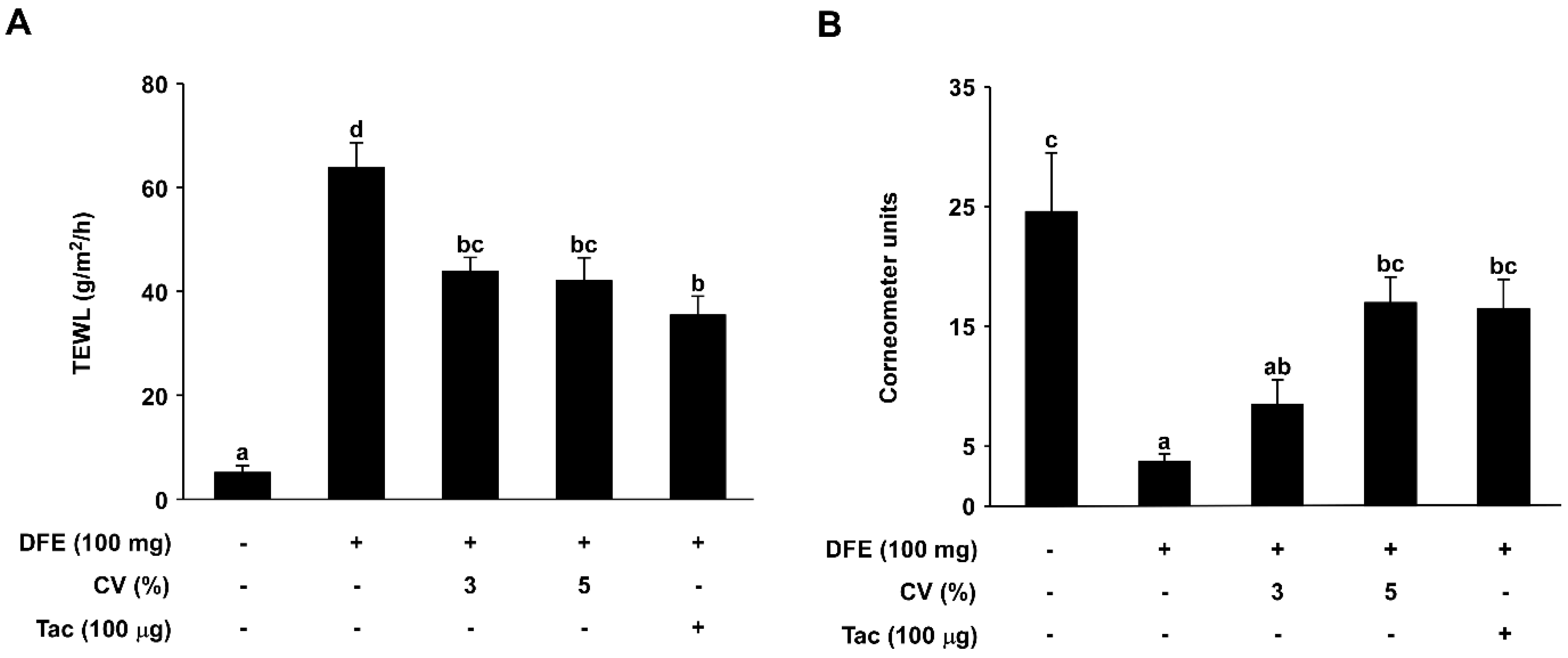
2.2. CV Supplementation Enhances Skin Hydration in DFE-Induced AD-Like Skin Lesions in NC/Nga Mice
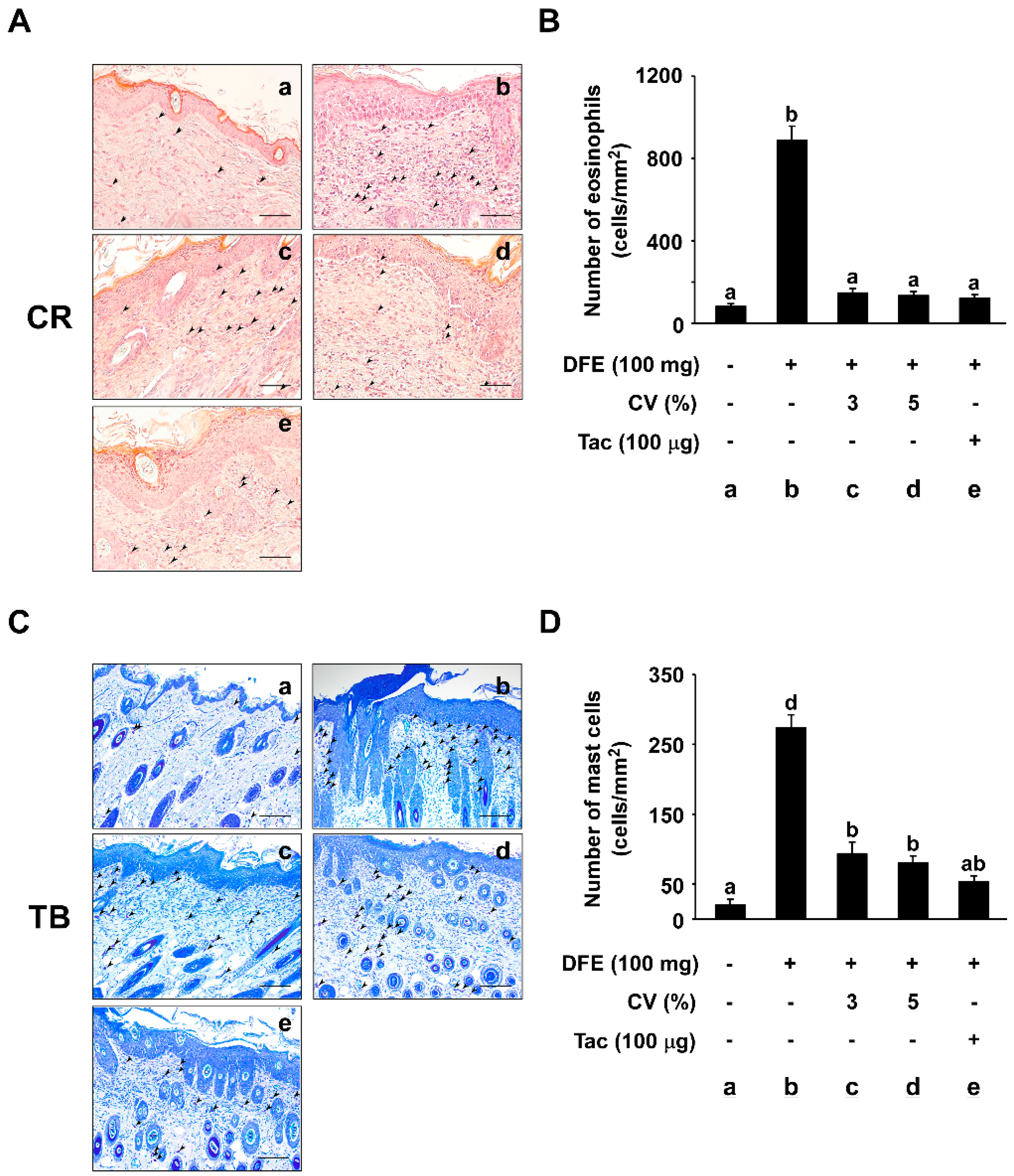
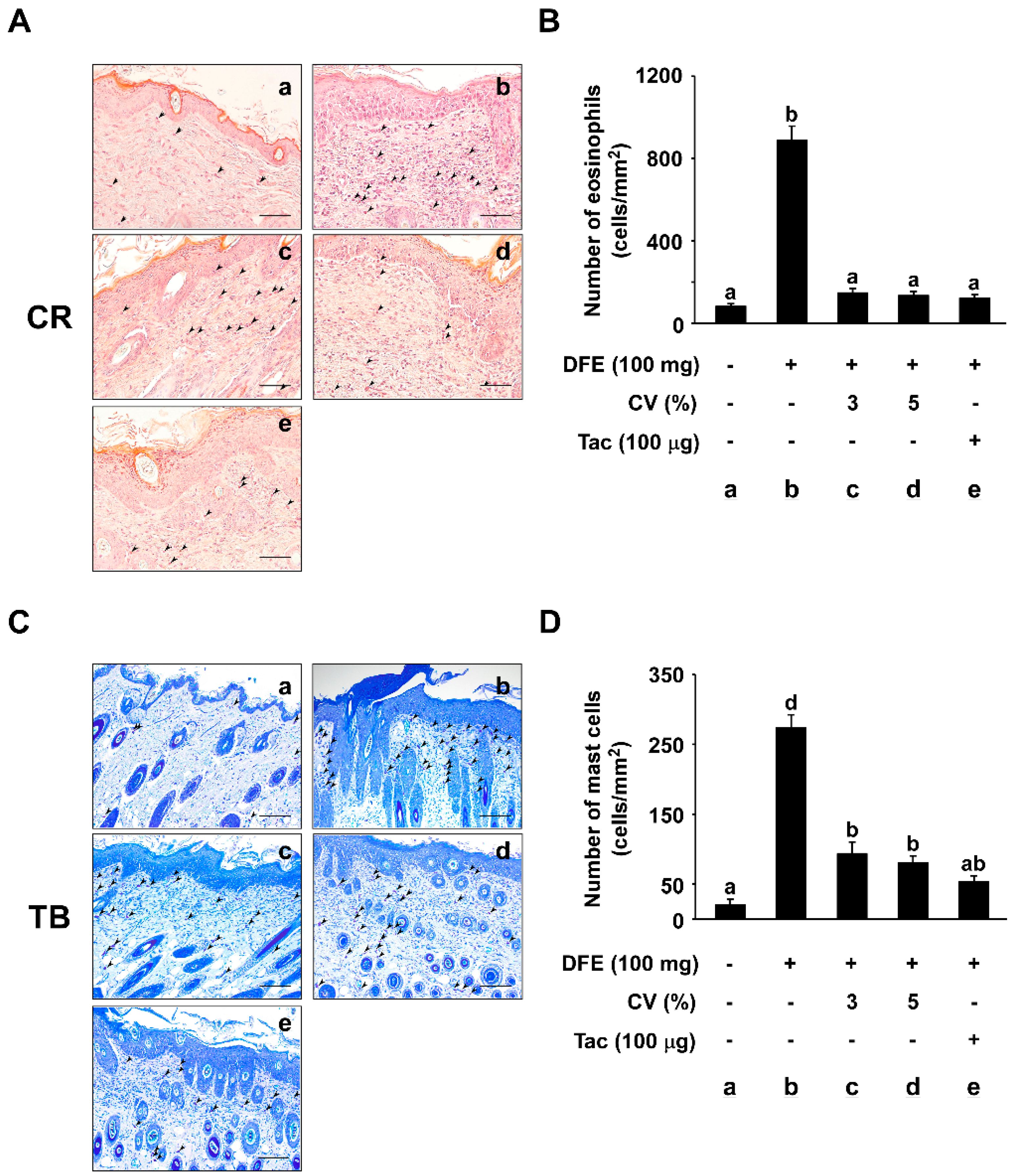
2.3. CV Supplementation Suppresses DFE-Induced Infiltration of Eosinophils and Mast Cells into Skin Lesions
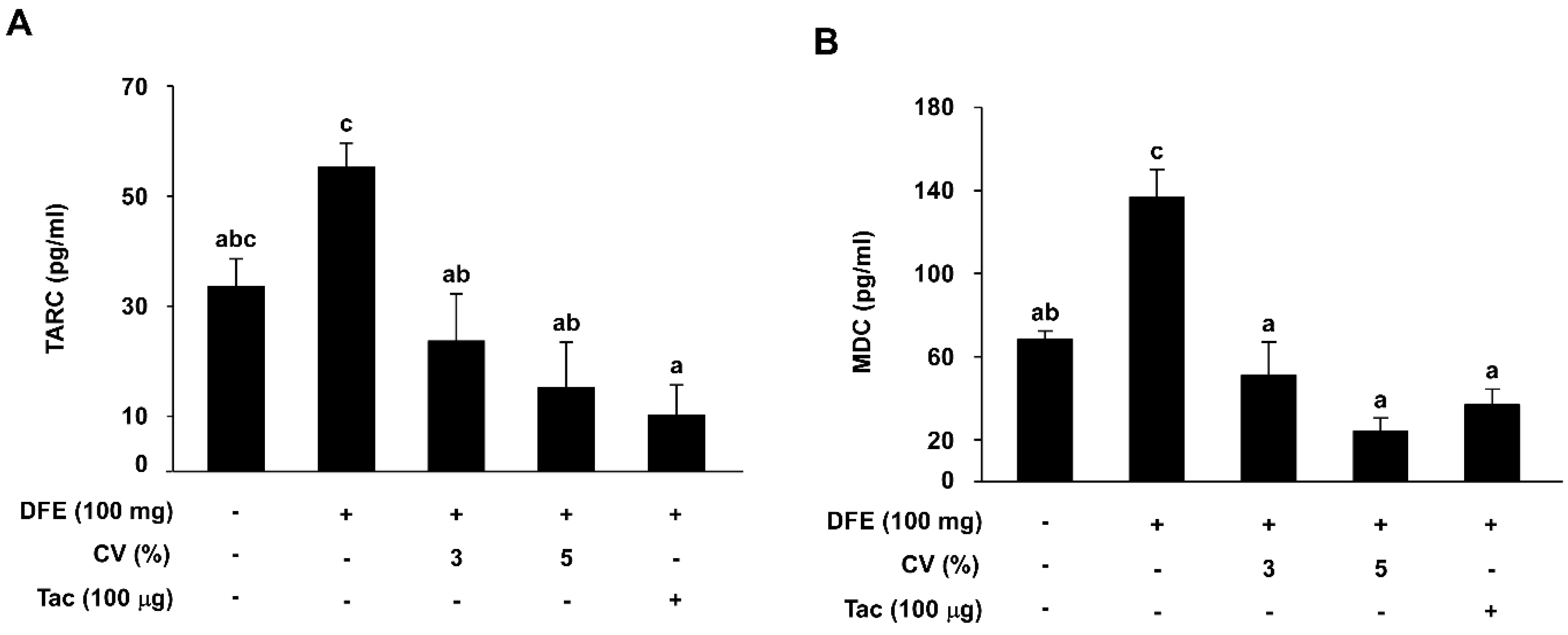
2.4. CV Supplementation Downregulates DFE-Induced Serum TARC and MDC Levels in NC/Nga Mice
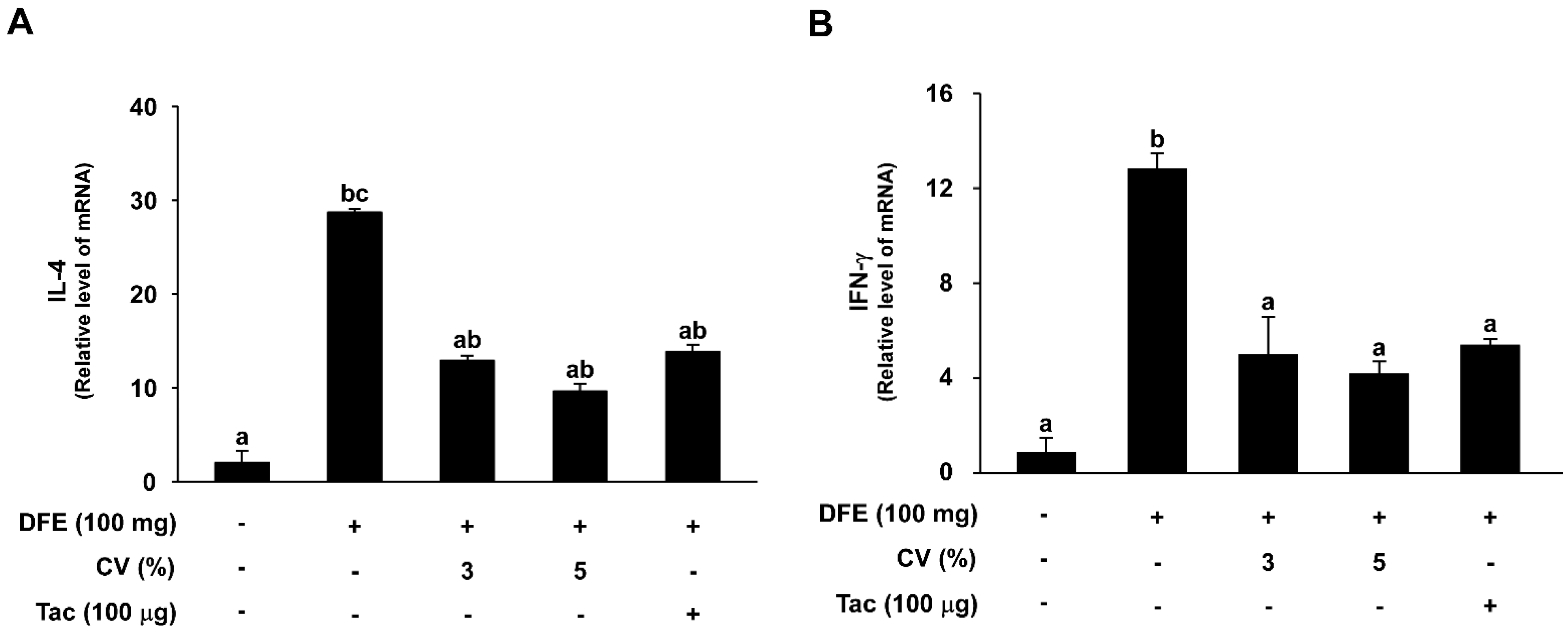
2.5. CV Supplementation Downregulates DFE-Induced Th2 and Th1 mRNA Expression Levels in NC/Nga Mice
3. Discussion
4. Experimental Section
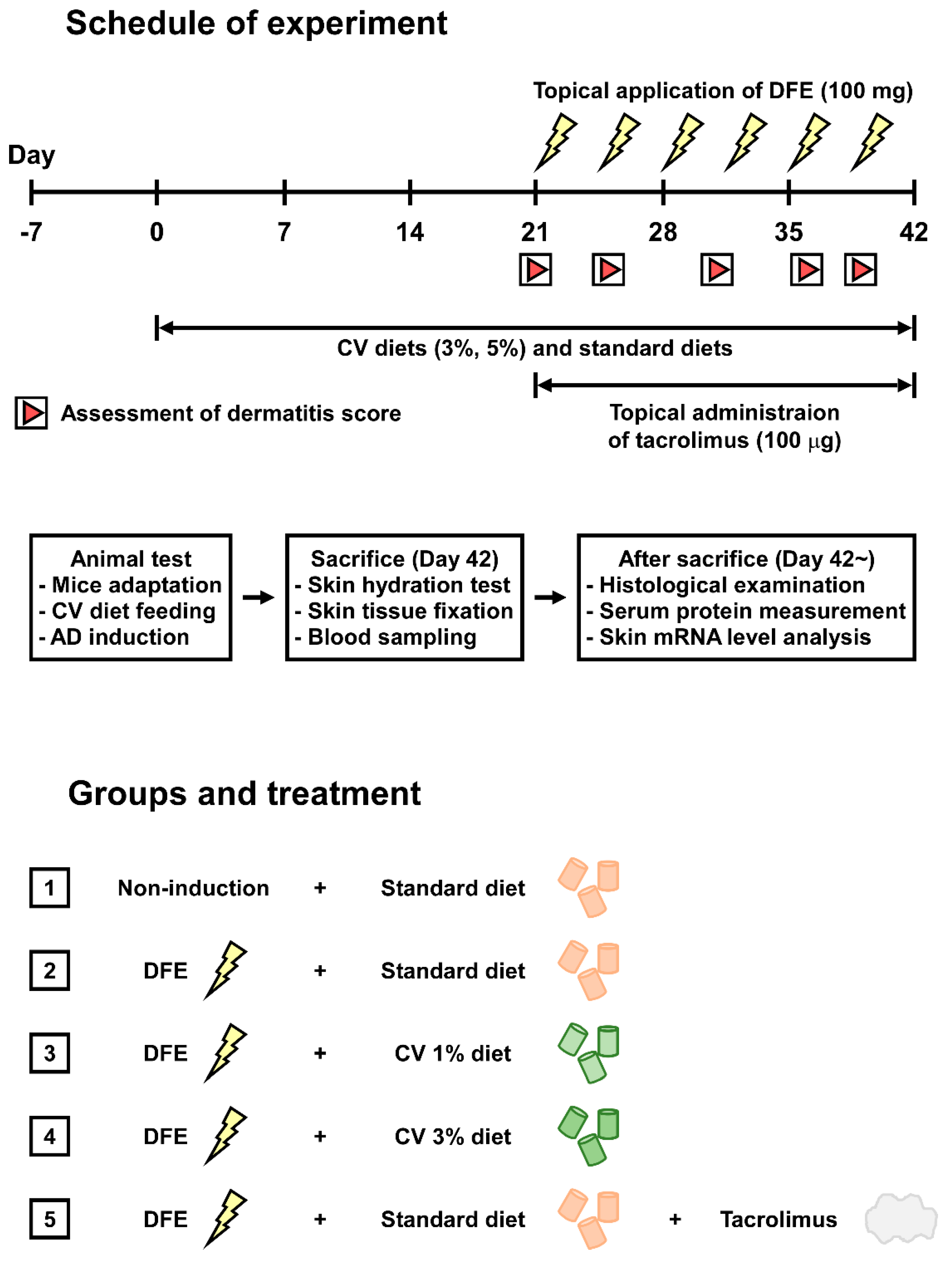
4.1. Chlorella Vulgaris (CV) Supplementation
4.2. Animals
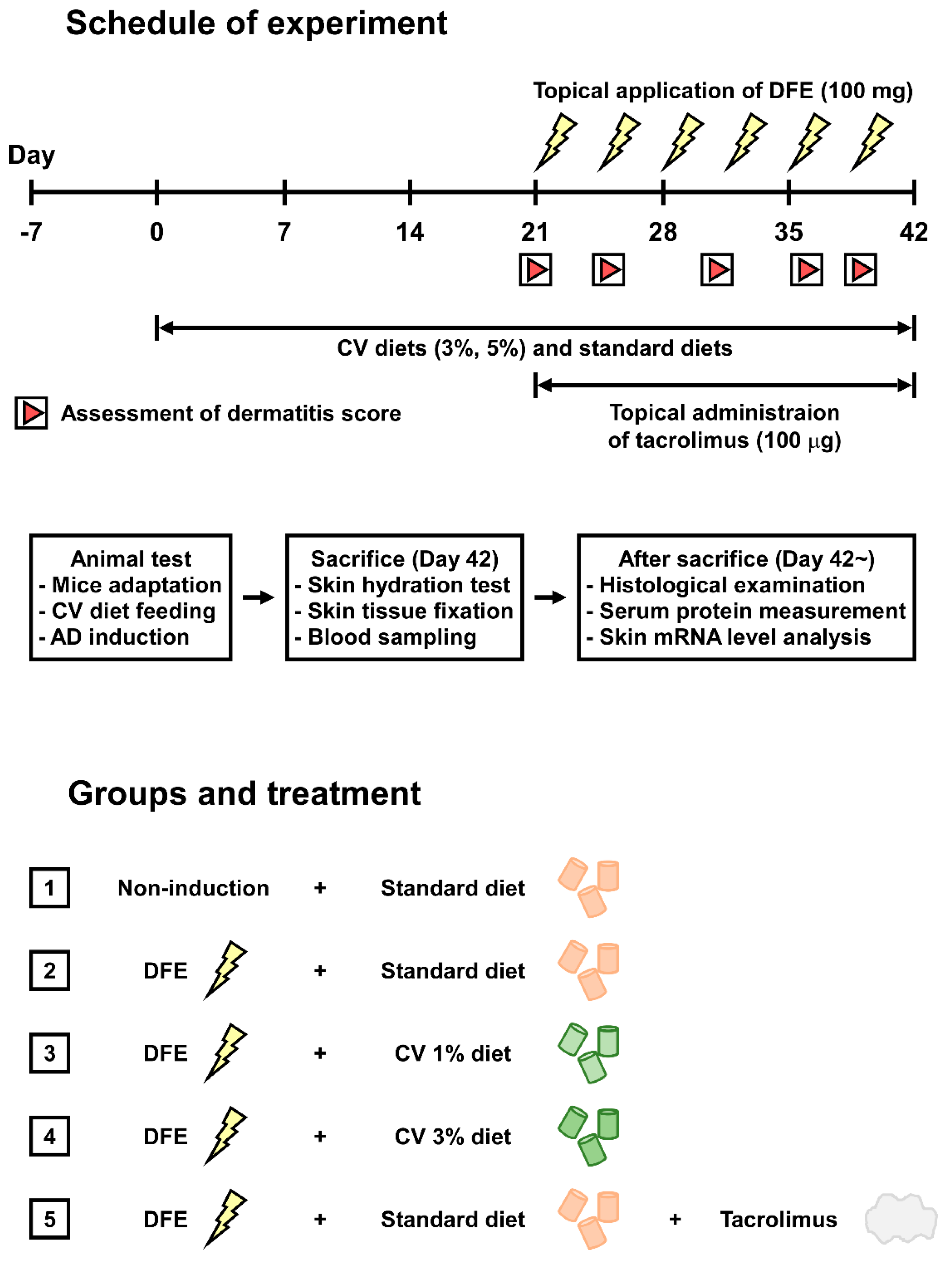
4.3. Induction of Atopic Dermatitis (AD)-Like Symptoms
4.4. Assessment of Skin Lesions and Dermatitis Scores
4.5. Histological Examination
4.6. Measurement of Serum TARC and MDC Levels
4.7. Measurement of Th2 and Th1 Cytokine mRNA Levels
4.8. Assessment of Transepidermal Water Loss (TEWL) and Corneometer Units
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Leung, D.Y. Atopic dermatitis: New insights and opportunities for therapeutic intervention. J. Allergy Clin. Immunol. 2000, 105, 860–876. [Google Scholar] [CrossRef] [PubMed]
- Flohr, C.; Mann, J. New insights into the epidemiology of childhood atopic dermatitis. Allergy 2014, 69, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Totri, C.R.; Diaz, L.; Eichenfield, L.F. 2014 update on atopic dermatitis in children. Curr. Opin. Pediatr. 2014, 26, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Spergel, J.M.; Paller, A.S. Atopic dermatitis and the atopic march. J. Allergy Clin. Immunol. 2003, 112, S118–S127. [Google Scholar] [CrossRef] [PubMed]
- Von Kobyletzki, L.B.; Bornehag, C.G.; Hasselgren, M.; Larsson, M.; Lindstrom, C.B.; Svensson, A. Eczema in early childhood is strongly associated with the development of asthma and rhinitis in a prospective cohort. BMC Dermatol. 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Bieber, T. Atopic dermatitis. N. Engl. J. Med. 2008, 358, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Takehara, K.; Sato, S. Both Th2 and Th1 chemokines (TARC/CCL17, MDC/CCL22, and Mig/CXCL9) are elevated in sera from patients with atopic dermatitis. J. Dermatol. Sci. 2004, 34, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Charbonnier, A.S.; Duez, C.; Jacquet, A.; Stewart, G.A.; Tonnel, A.B.; Pestel, J. Th2 polarization by Der p 1—Pulsed monocyte-derived dendritic cells is due to the allergic status of the donors. Blood 2001, 98, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Smits, H.H.; Ratajczak, C.; Nithiananthan, A.; Wierenga, E.A.; Stewart, G.A.; Jacquet, A.; Tonnel, A.B.; Pestel, J. Monocyte-derived dendritic cells exposed to Der p 1 allergen enhance the recruitment of Th2 cells: Major involvement of the chemokines TARC/CCL17 and MDC/CCL22. Eur. Cytokine Netw 2003, 14, 219–228. [Google Scholar] [PubMed]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Ong, P.Y.; Leung, D.Y. Immune dysregulation in atopic dermatitis. Curr. Allergy Asthma. Rep. 2006, 6, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.R.; Choi, J.; Kim, J.; Kim, H.; Kang, H.; Kim, E.H.; Chang, J.H.; Kim, Y.E.; Choi, Y.J.; Lee, K.W.; et al. 20-O-β-d-glucopyranosyl-20(S)-protopanaxadiol-fortified ginseng extract attenuates the development of atopic dermatitis-like symptoms in NC/Nga mice. J. Ethnopharmacol. 2014, 151, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, J.R.; Kang, H.; Choi, J.; Yang, H.; Lee, P.; Kim, J.; Lee, K.W. 7,8,4ʹ-Trihydroxyisoflavone attenuates DNCB-induced atopic dermatitis-like symptoms in NC/Nga mice. PLoS ONE 2014, 9, e104938. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Terao, H.; Rikihisa, W.; Urabe, K.; Kinukawa, N.; Nose, Y.; Koga, T. Clinical dose and adverse effects of topical steroids in daily management of atopic dermatitis. Br. J. Dermatol. 2003, 148, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Hengge, U.R.; Ruzicka, T.; Schwartz, R.A.; Cork, M.J. Adverse effects of topical glucocorticosteroids. J. Am. Acad. Dermatol. 2006, 54, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Micro-algae as sources of fine chemicals. Microbiol. Sci. 1986, 3, 372–375. [Google Scholar] [PubMed]
- Morimura, Y.; Tamiya, N. Preliminary experiments in the use of Chlorella as human food. Food Technol. 1954, 8, 179–182. [Google Scholar]
- Konishi, F.; Tanaka, K.; Himeno, K.; Taniguchi, K.; Nomoto, K. Antitumor effect induced by a hot water extract of Chlorella vulgaris (CE): Resistance to meth-A tumor growth mediated by CE-induced polymorphonuclear leukocytes. Cancer Immunol. Immunother. 1985, 19, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Chovančíková, M.; Šimek, V. Effects of hight-fat and Chlorella vulgaris feeding on changes in lipid metabolism in mice. Biol. Bratisl. 2001, 56, 661–666. [Google Scholar]
- Jong-Yuh, C.; Mei-Fen, S. Potential hypoglycemic effects of Chlorella in streptozotocin-induced diabetic mice. Life Sci. 2005, 77, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, W.; Kim, Y.H.; Lee, Y.W. Chlorella vulgaris extract ameliorates carbon tetrachloride-induced acute hepatic injury in mice. Exp. Toxicol. Pathol. 2013, 65, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Ito, K.; Ueno, S.; Kumamoto, S.; Ando, Y.; Yamada, A.; Nomoto, K.; Yasunobu, Y. Oral administration of hot water extracts of Chlorella vulgaris reduces IgE production against milk casein in mice. Int. J. Immunopharmacol. 1999, 21, 311–323. [Google Scholar] [CrossRef]
- Bae, M.J.; Shin, H.S.; Chai, O.H.; Han, J.G.; Shon, D.H. Inhibitory effect of unicellular green algae (Chlorella vulgaris) water extract on allergic immune response. J. Sci. Food Agric. 2013, 93, 3133–3136. [Google Scholar] [CrossRef] [PubMed]
- Spoehr, H.A. Chlorella as a source of food. Proc. Am. Philos. Soc. 1951, 95, 62–67. [Google Scholar]
- Kwak, J.H.; Baek, S.H.; Woo, Y.; Han, J.K.; Kim, B.G.; Kim, O.Y.; Lee, J.H. Beneficial immunostimulatory effect of short-term Chlorella supplementation: Enhancement of natural killer cell activity and early inflammatory response (randomized, double-blinded, placebo-controlled trial). Nutr. J. 2012, 11. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Werner, Y.; Lindberg, M. Transepidermal water loss in dry and clinically normal skin in patients with atopic dermatitis. Acta Derm. Venereol. 1985, 65, 102–105. [Google Scholar] [PubMed]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Lebman, D.A.; Coffman, R.L. Interleukin 4 causes isotype switching to IgE in T cell-stimulated clonal B cell cultures. J. Exp. Med. 1988, 168, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Jujo, K.; Renz, H.; Abe, J.; Gelfand, E.W.; Leung, D.Y. Decreased interferon gamma and increased interleukin-4 production in atopic dermatitis promotes IgE synthesis. J. Allergy Clin. Immunol. 1992, 90, 323–331. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.; Lee, C.H.; Kim, J.R.; Kwon, J.Y.; Seo, S.G.; Han, J.G.; Kim, B.G.; Kim, J.-E.; Lee, K.W. Chlorella vulgaris Attenuates Dermatophagoides Farinae-Induced Atopic Dermatitis-Like Symptoms in NC/Nga Mice. Int. J. Mol. Sci. 2015, 16, 21021-21034. https://doi.org/10.3390/ijms160921021
Kang H, Lee CH, Kim JR, Kwon JY, Seo SG, Han JG, Kim BG, Kim J-E, Lee KW. Chlorella vulgaris Attenuates Dermatophagoides Farinae-Induced Atopic Dermatitis-Like Symptoms in NC/Nga Mice. International Journal of Molecular Sciences. 2015; 16(9):21021-21034. https://doi.org/10.3390/ijms160921021
Chicago/Turabian StyleKang, Heerim, Chang Hyung Lee, Jong Rhan Kim, Jung Yeon Kwon, Sang Gwon Seo, Jae Gab Han, Byung Gon Kim, Jong-Eun Kim, and Ki Won Lee. 2015. "Chlorella vulgaris Attenuates Dermatophagoides Farinae-Induced Atopic Dermatitis-Like Symptoms in NC/Nga Mice" International Journal of Molecular Sciences 16, no. 9: 21021-21034. https://doi.org/10.3390/ijms160921021
APA StyleKang, H., Lee, C. H., Kim, J. R., Kwon, J. Y., Seo, S. G., Han, J. G., Kim, B. G., Kim, J.-E., & Lee, K. W. (2015). Chlorella vulgaris Attenuates Dermatophagoides Farinae-Induced Atopic Dermatitis-Like Symptoms in NC/Nga Mice. International Journal of Molecular Sciences, 16(9), 21021-21034. https://doi.org/10.3390/ijms160921021