
A Natural Triterpene Derivative from Euphorbia kansui Inhibits Cell Proliferation and Induces Apoptosis against Rat Intestinal Epithelioid Cell Line in Vitro

 , and
, and
Abstract
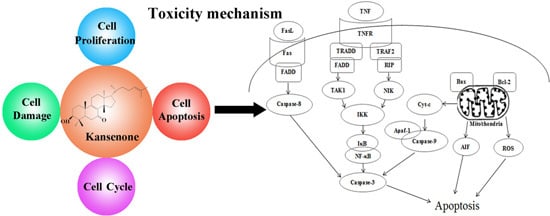
:
1. Introduction
2. Results and Discussion
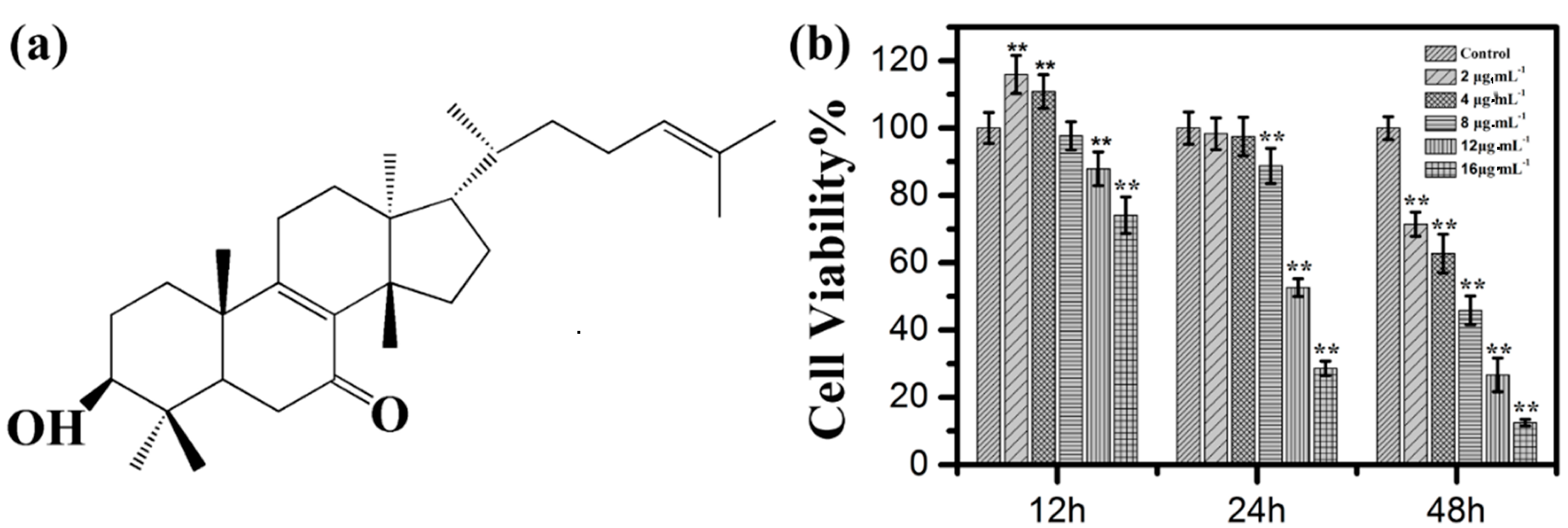
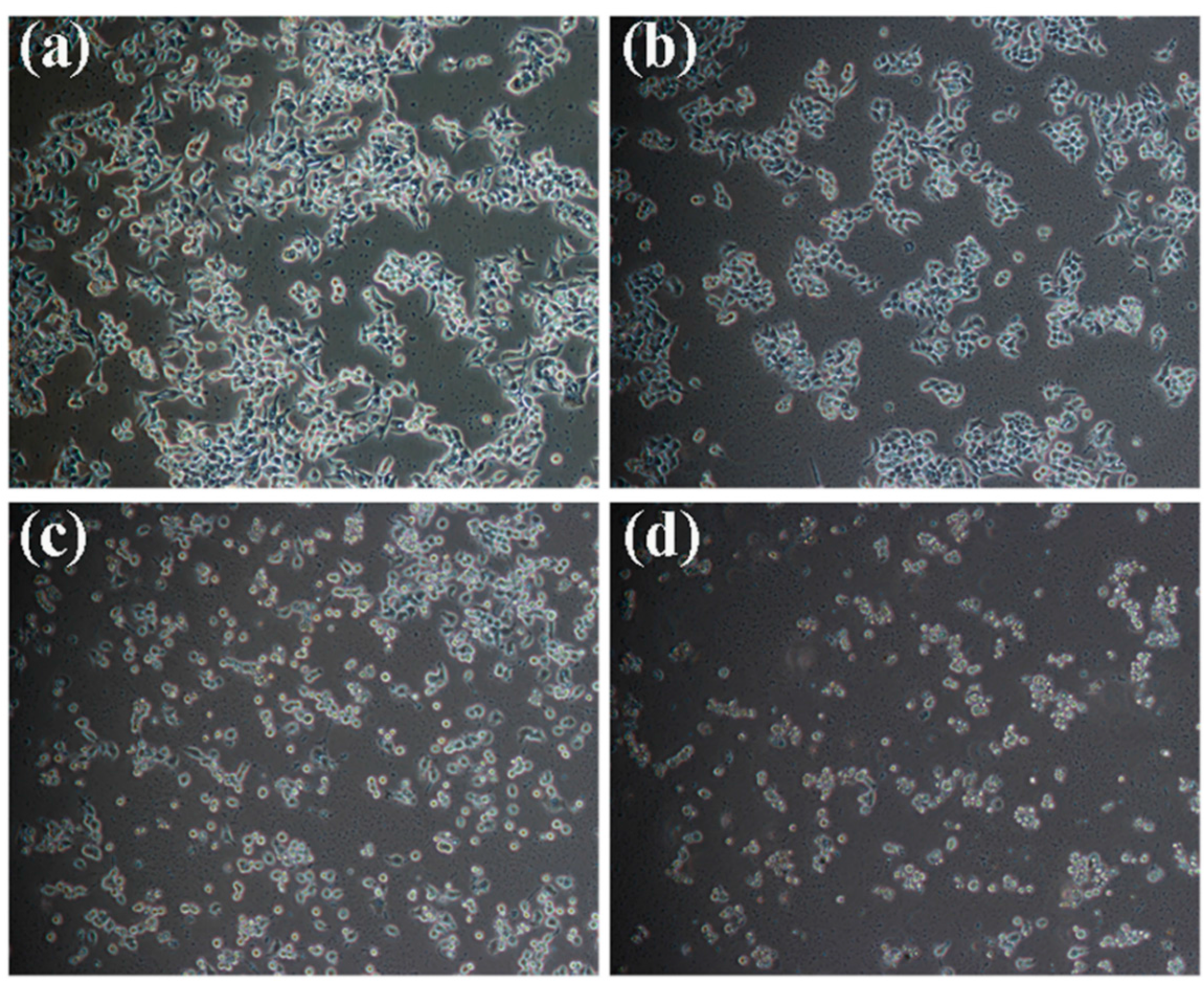
2.1. Effects of Kansenone on Cell Proliferation and Cell Morphology
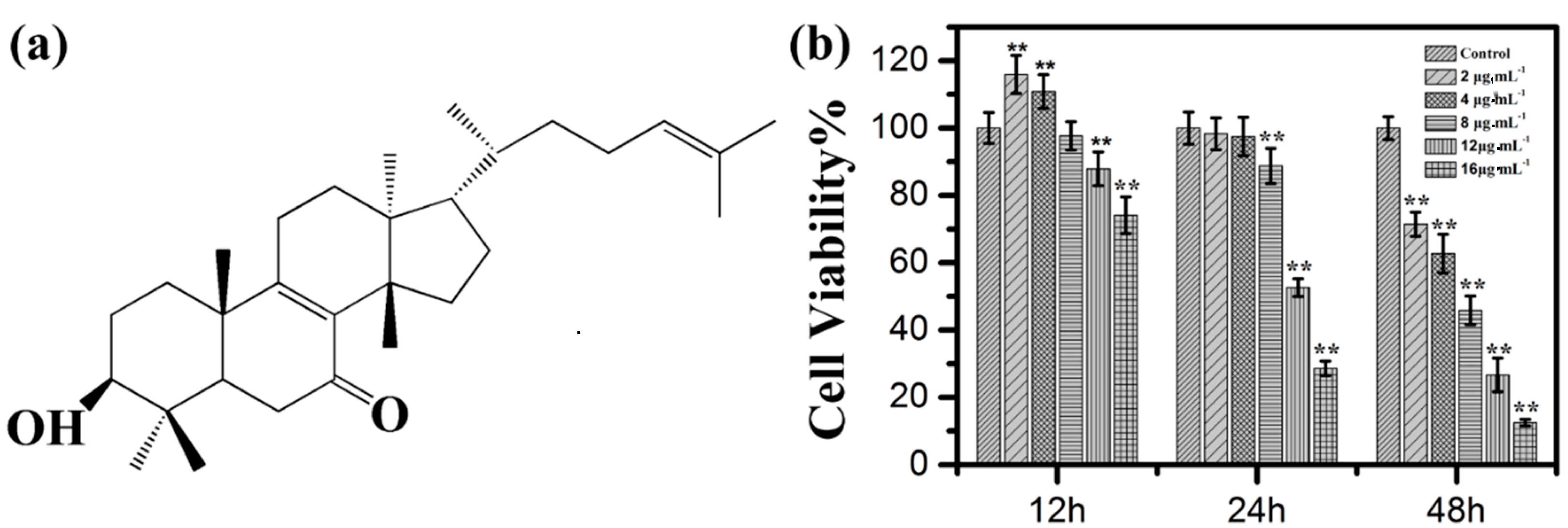
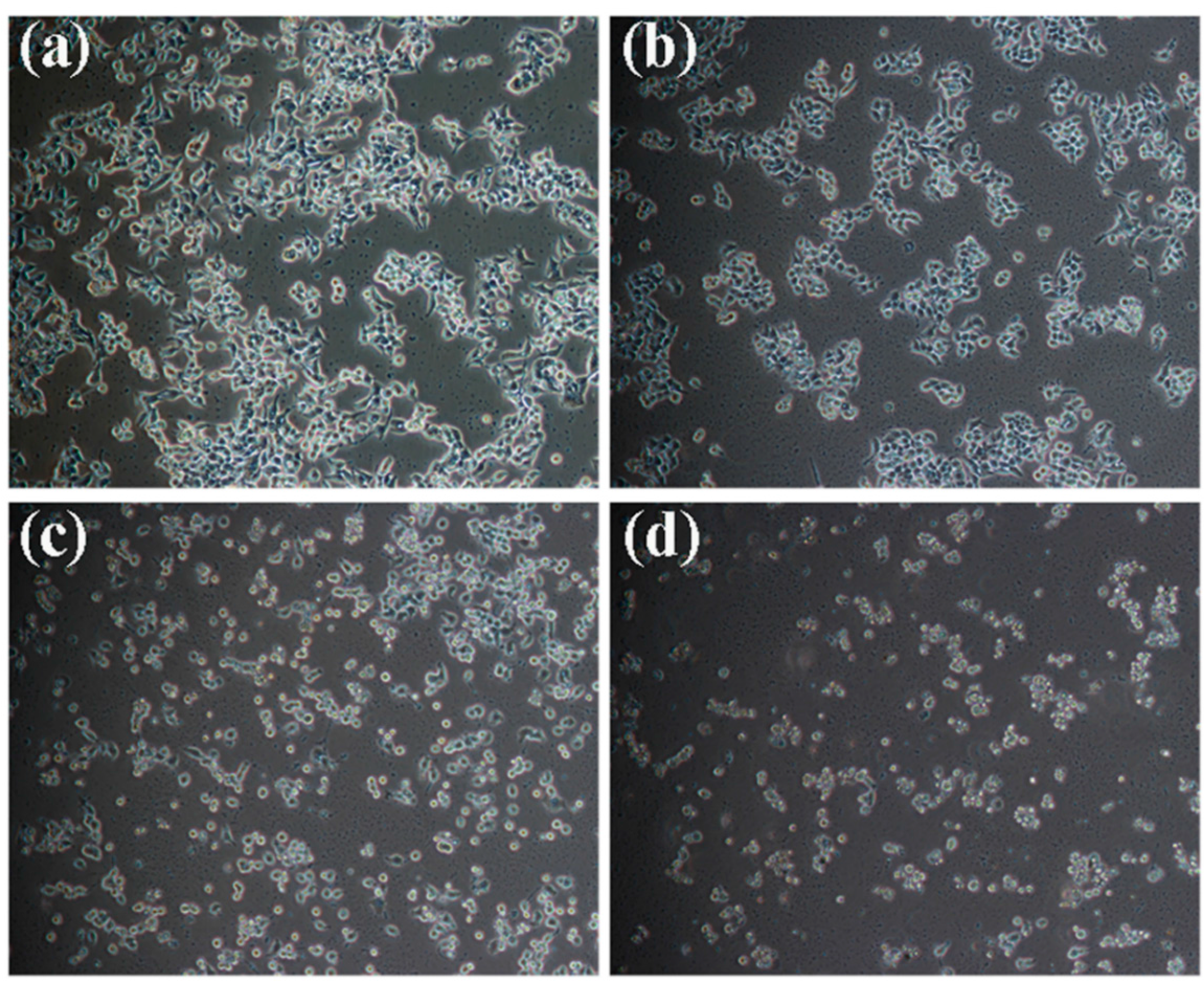
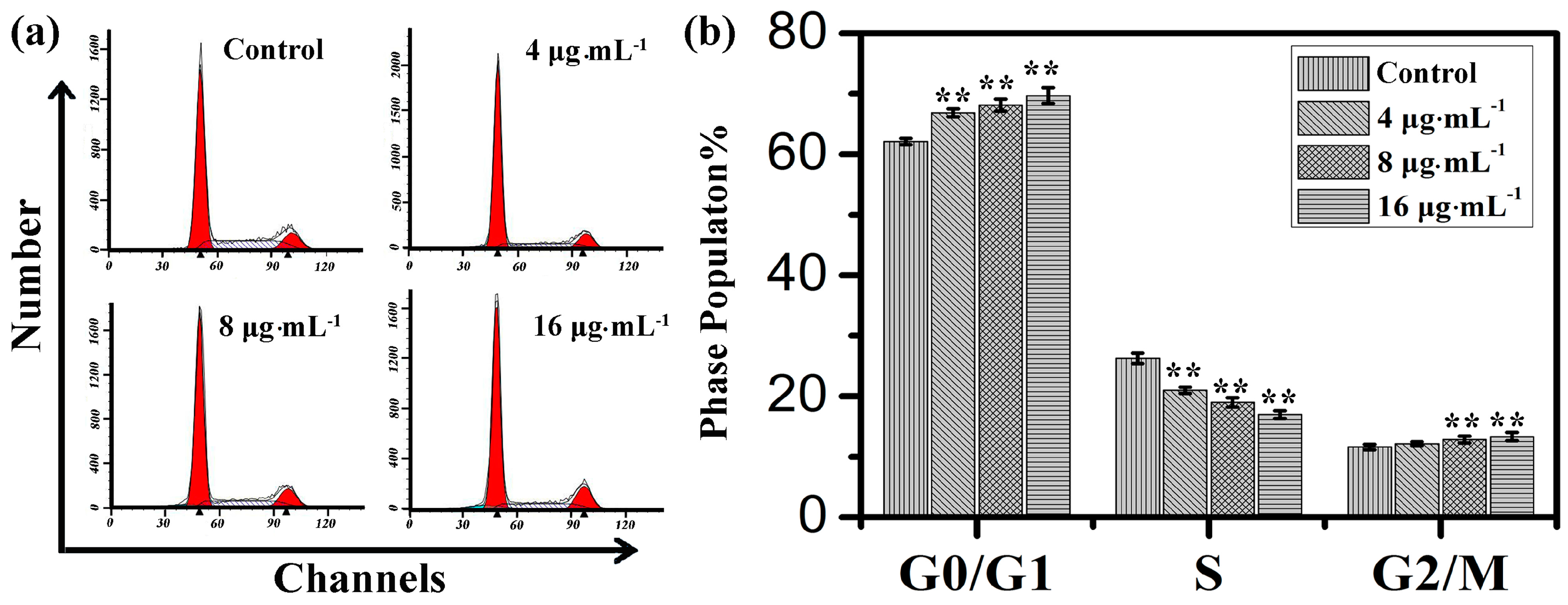
2.2. Effects of Kansenone on Cell Cycle
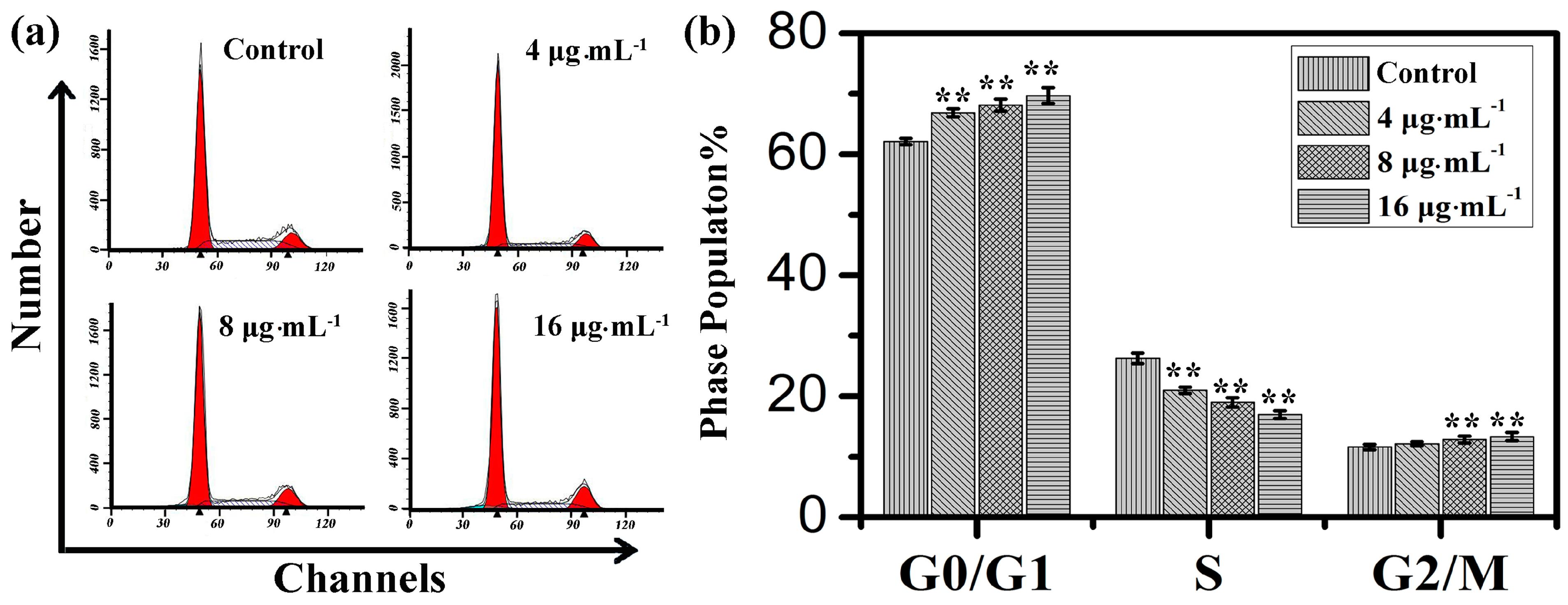
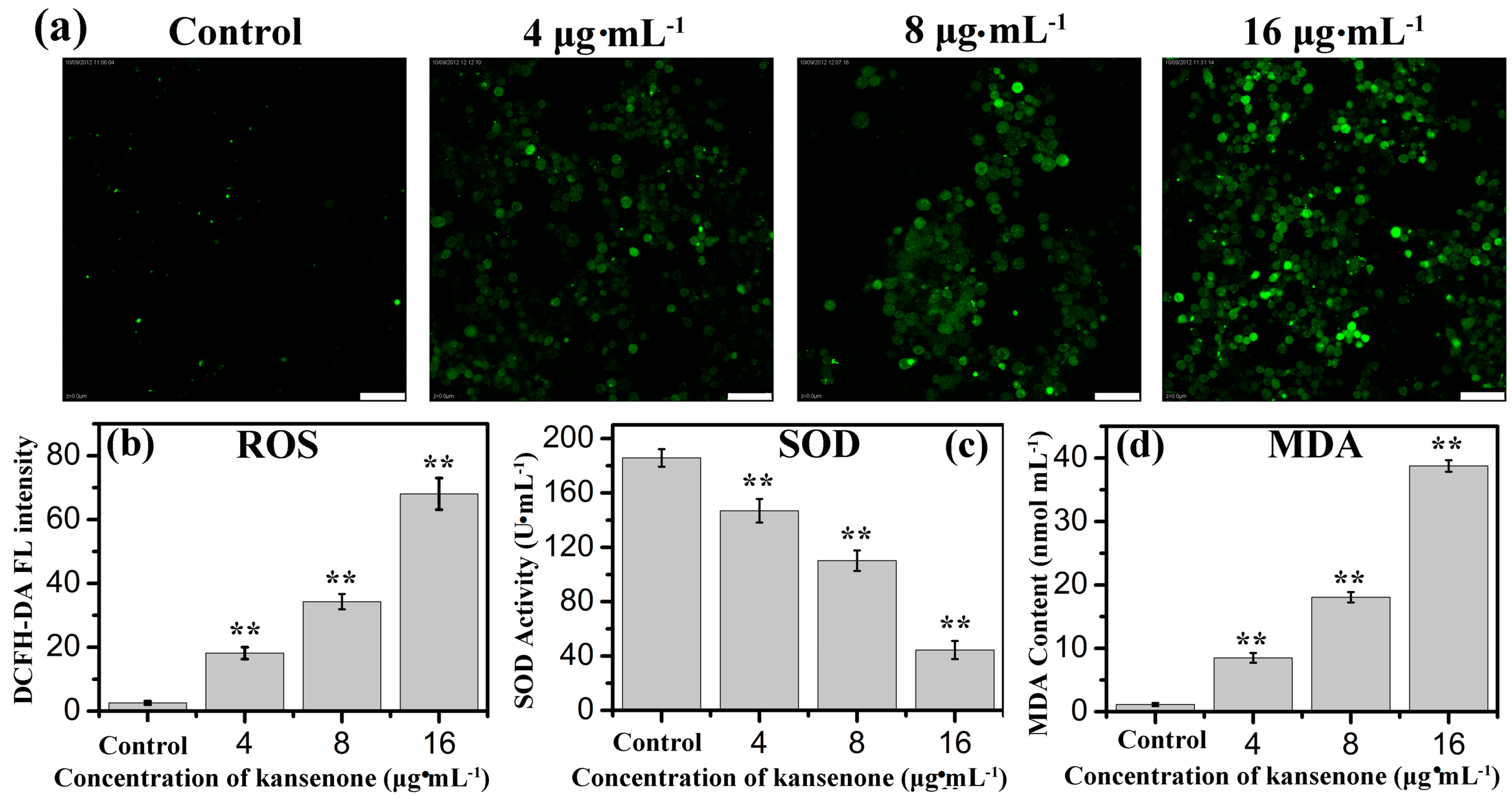
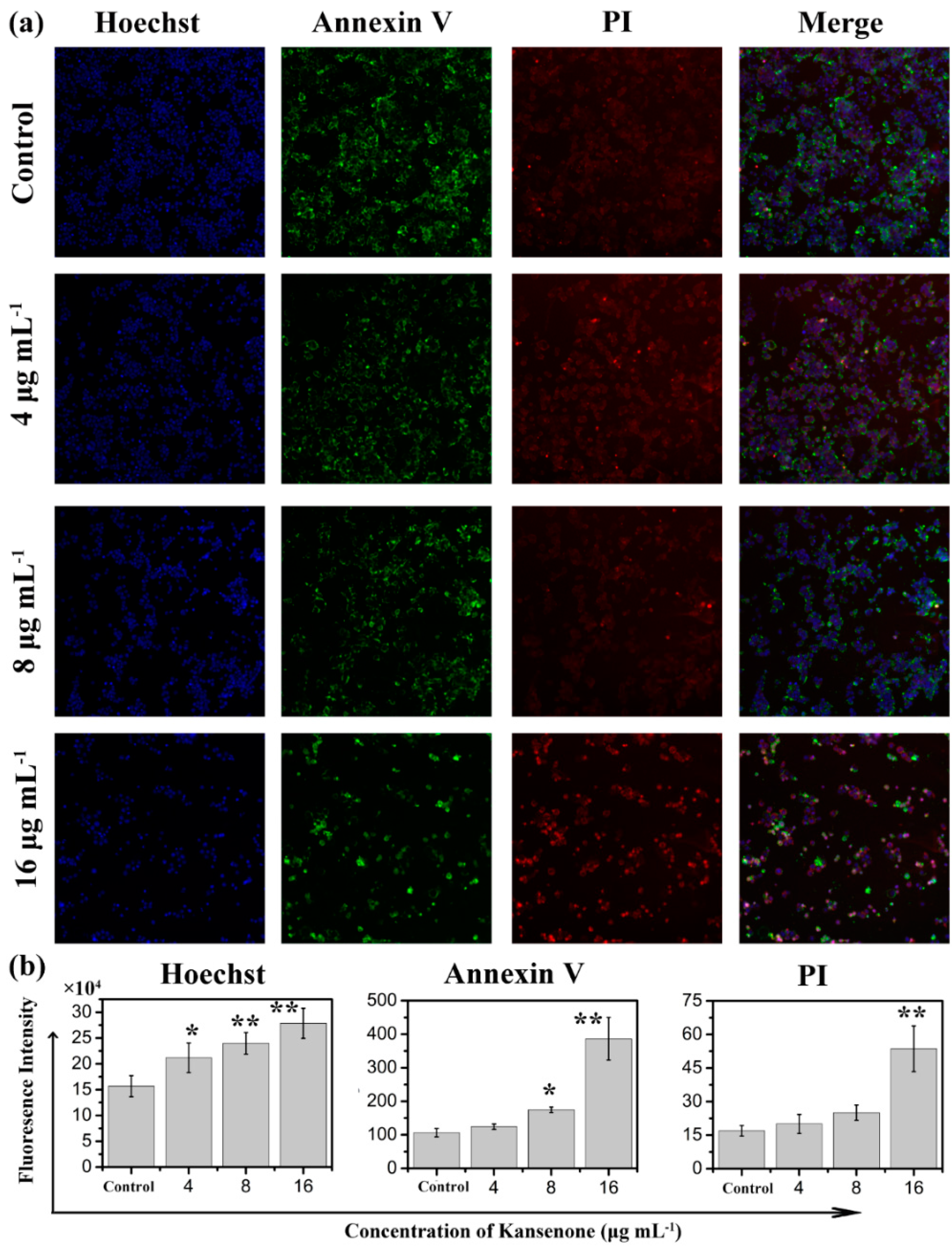
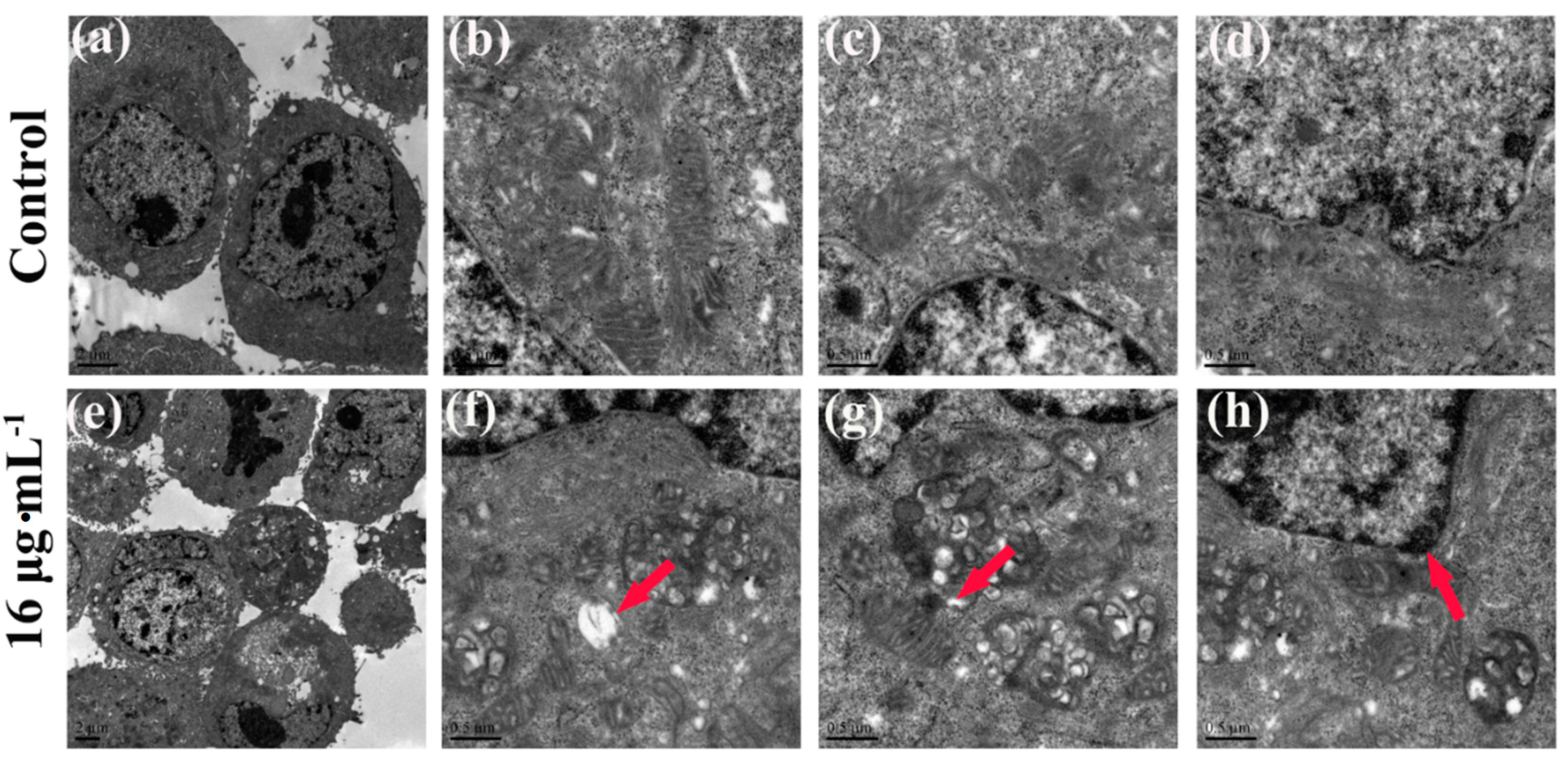
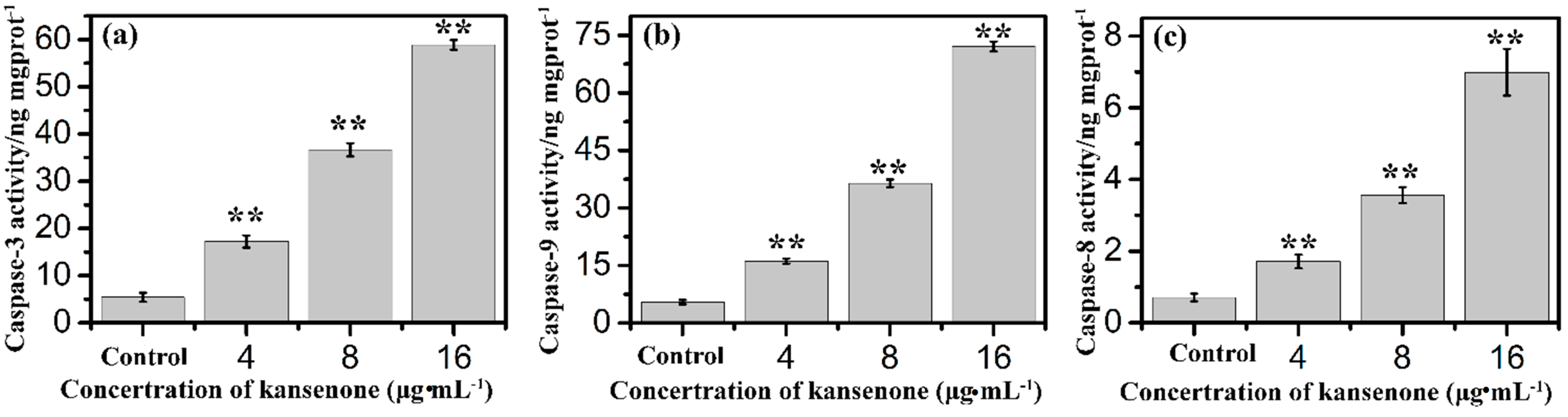
2.3. Effects of Kansenone on Cell Damage
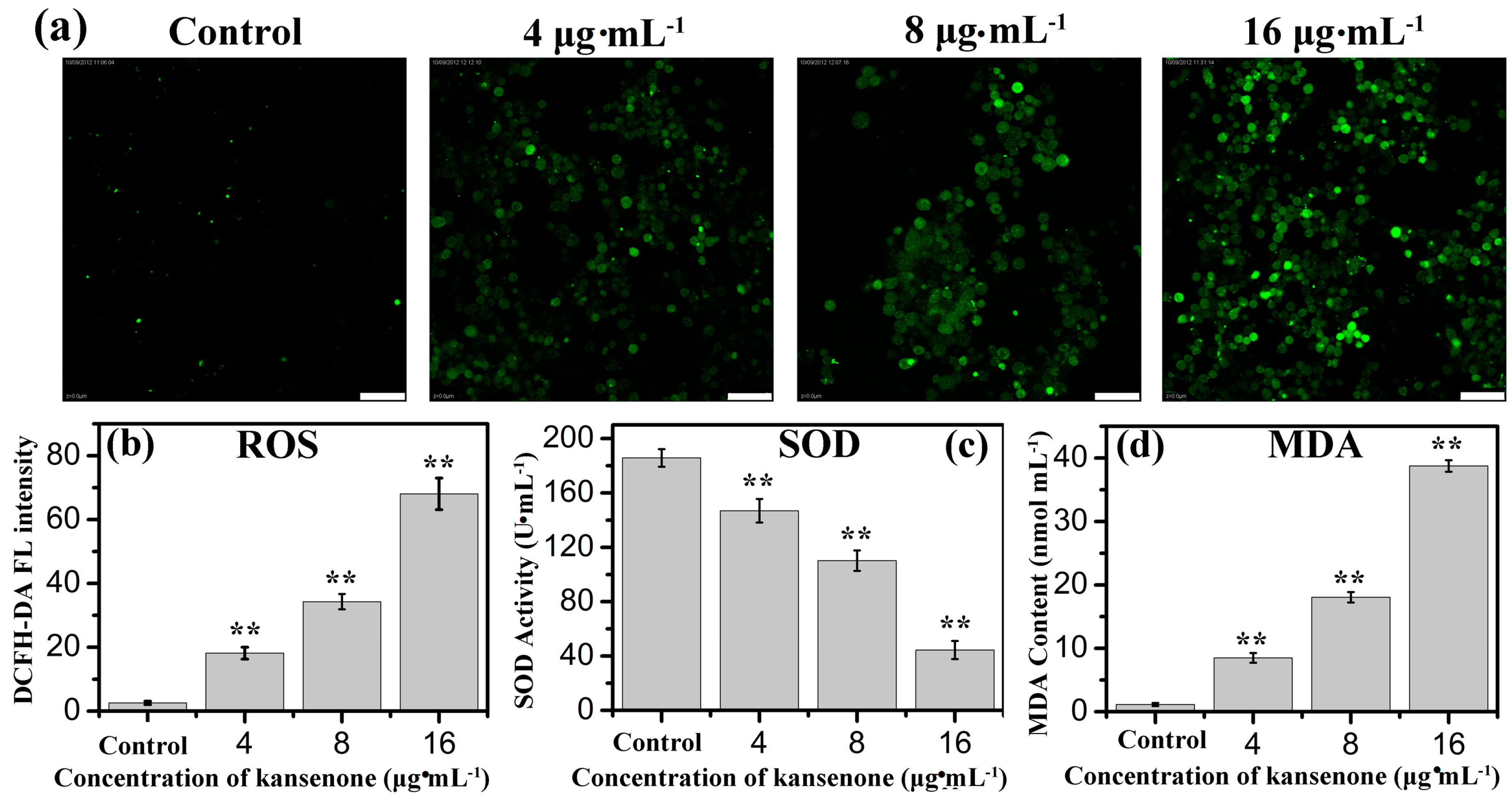
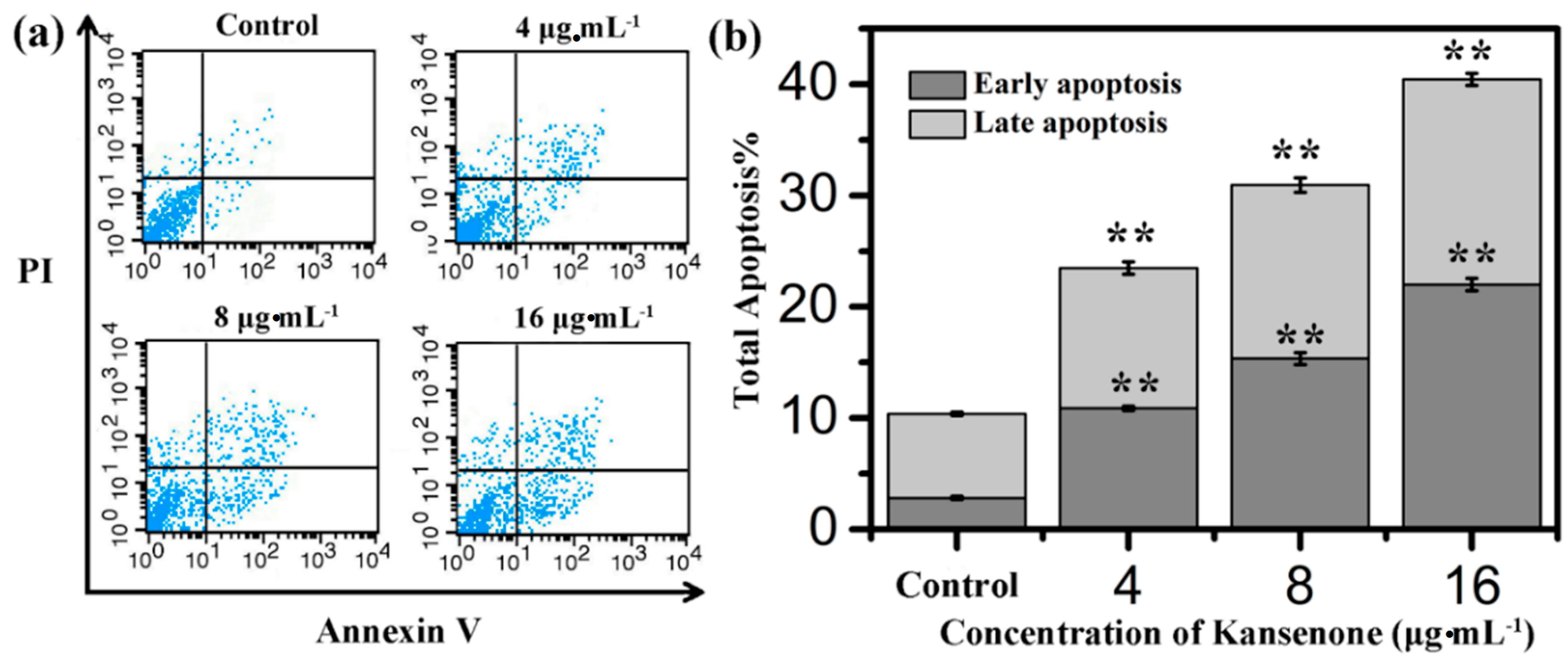
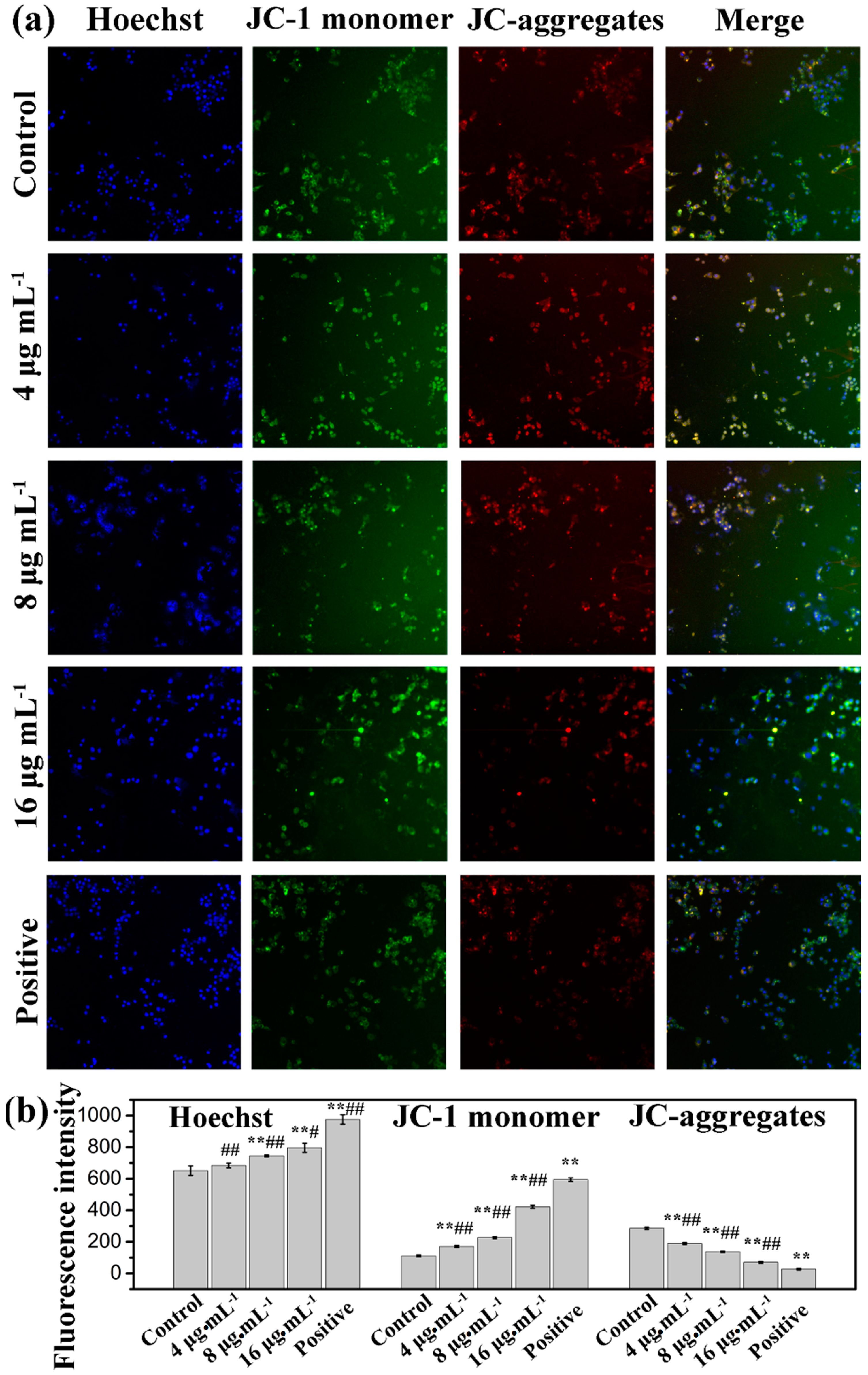
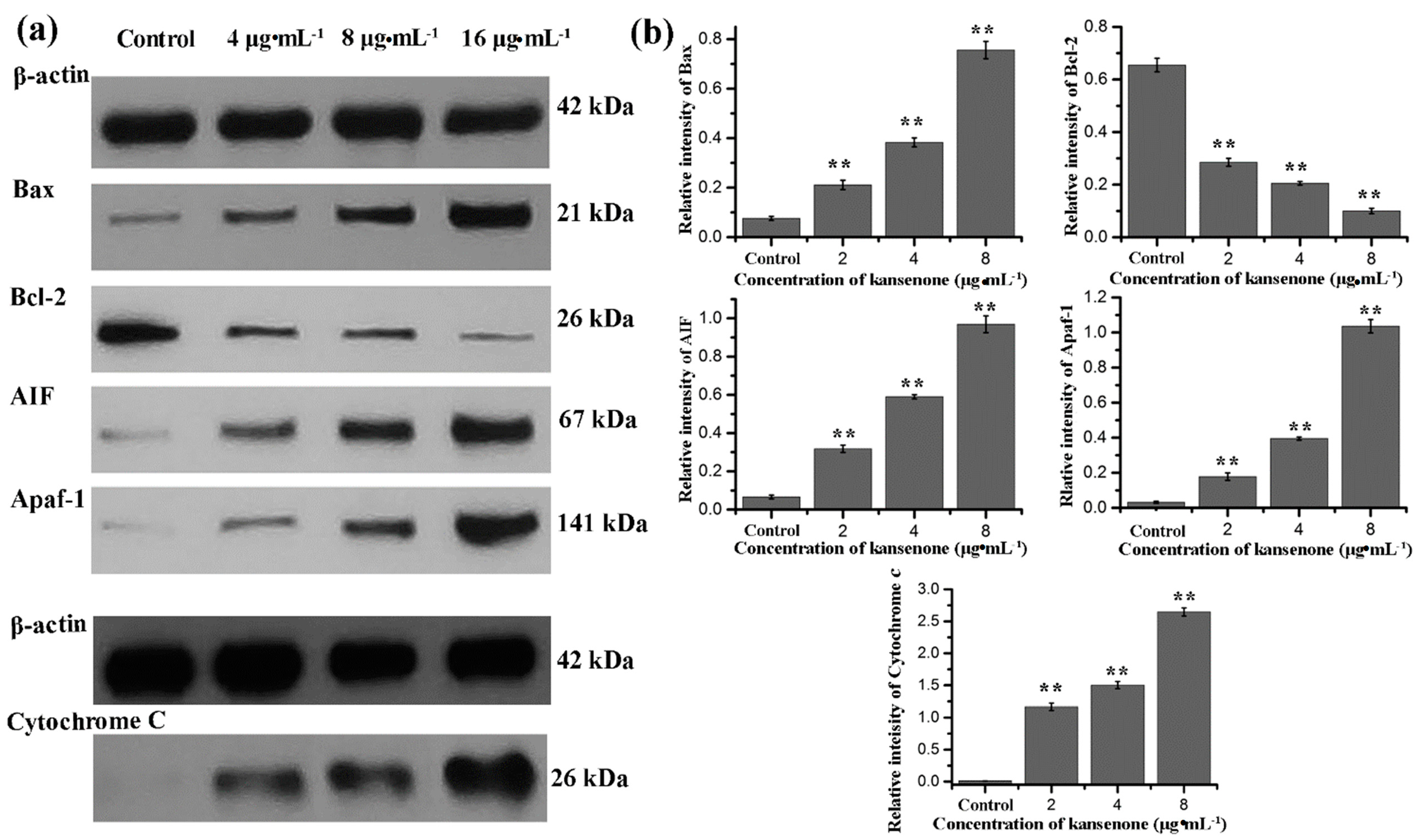
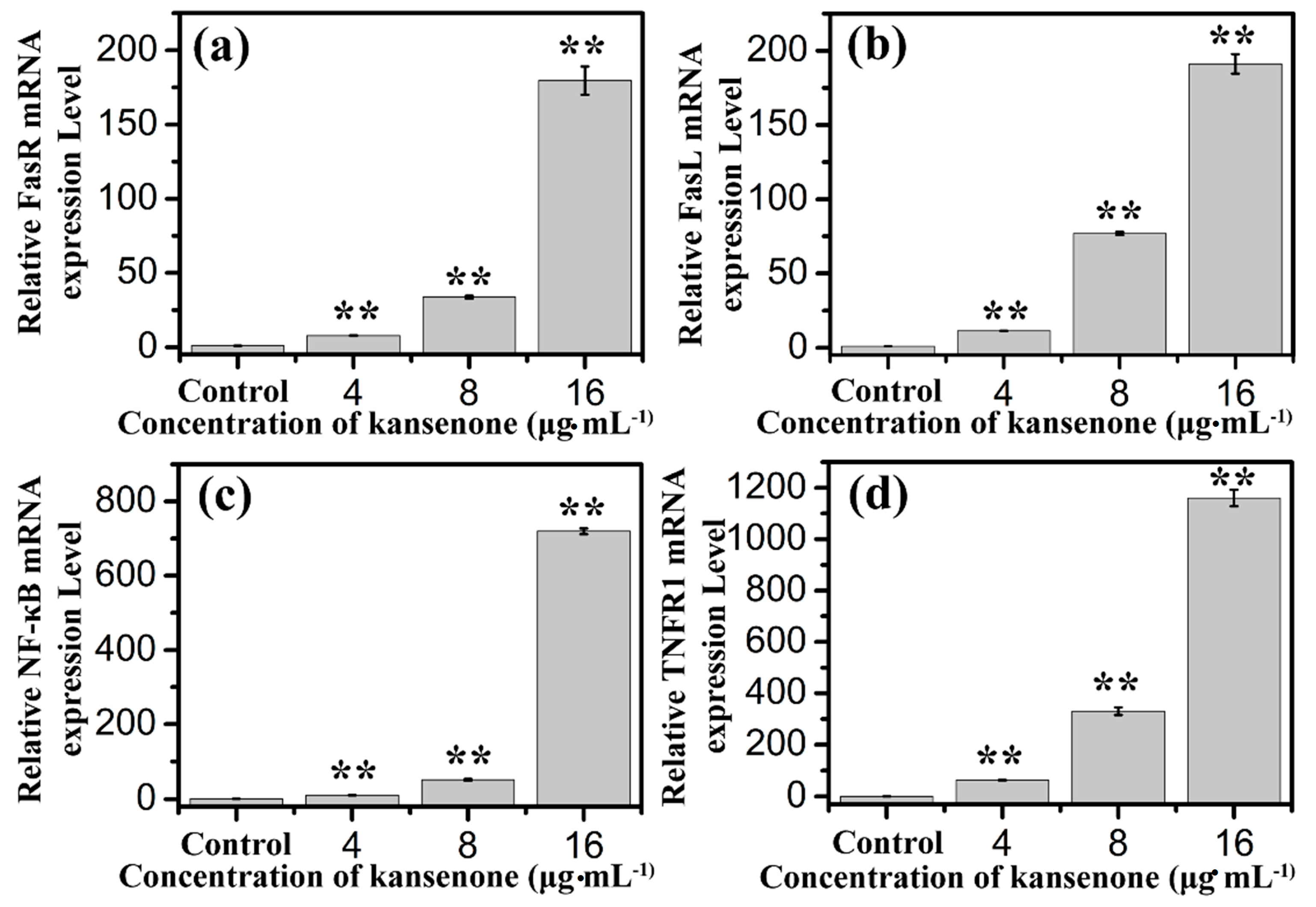
2.4. Effects of Kansenone on Cell Apoptosis
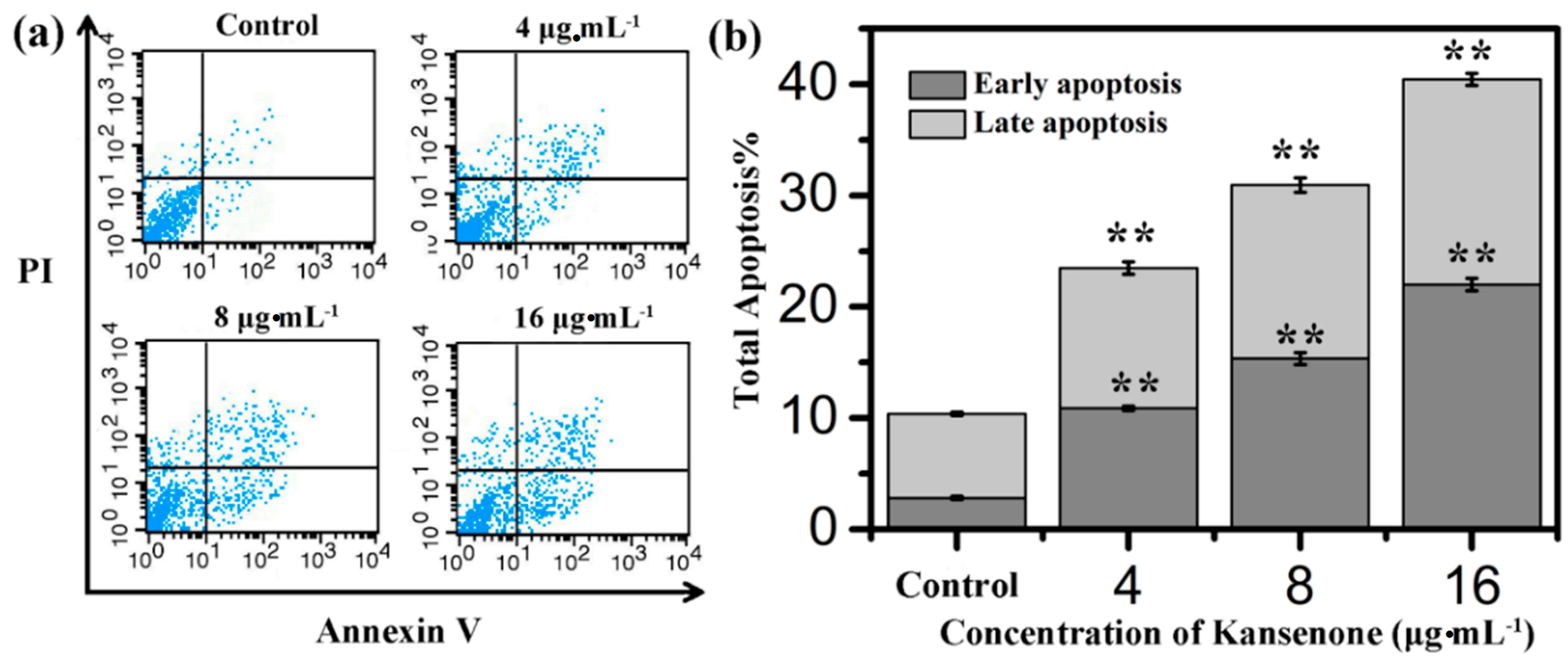

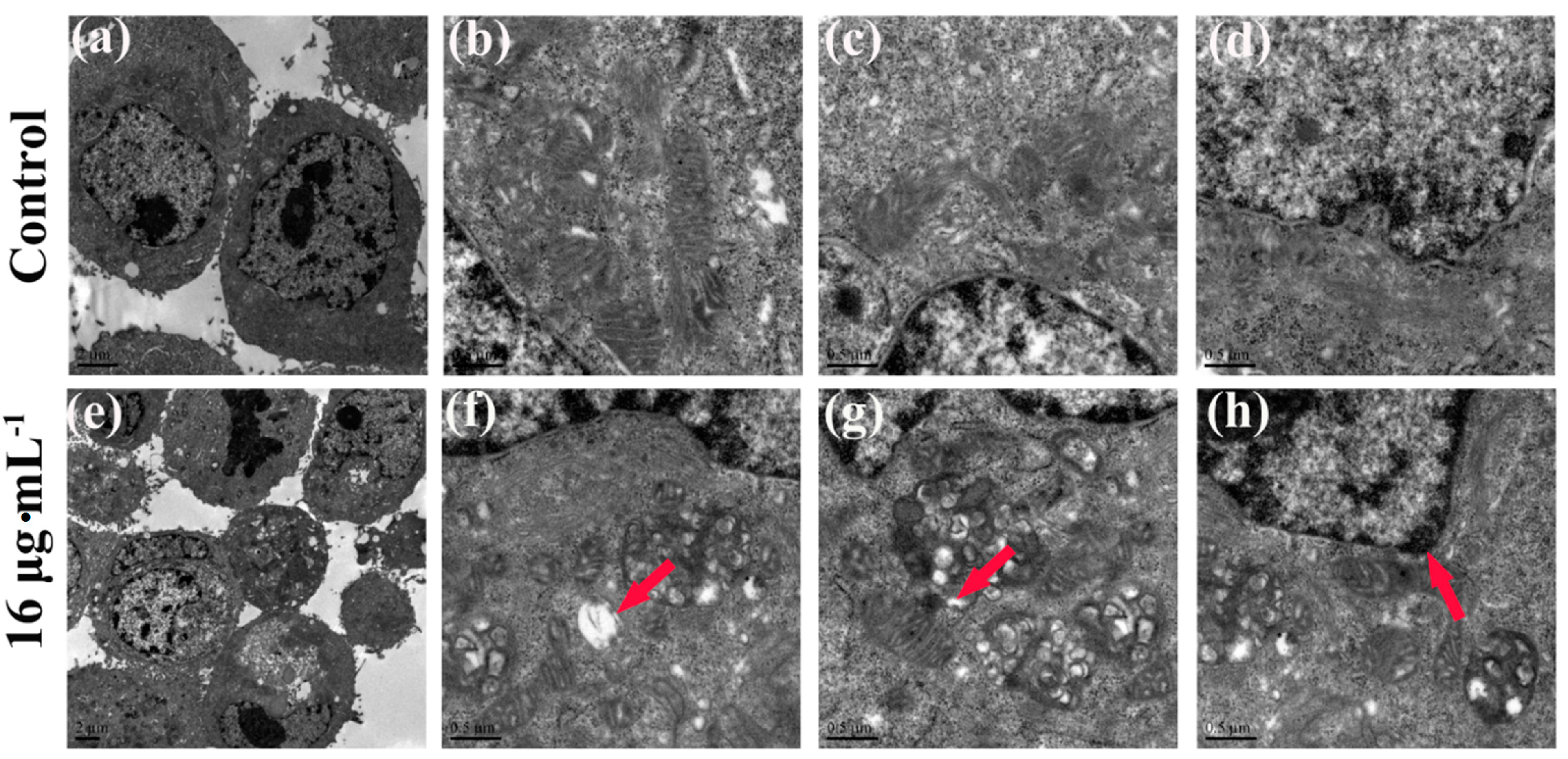
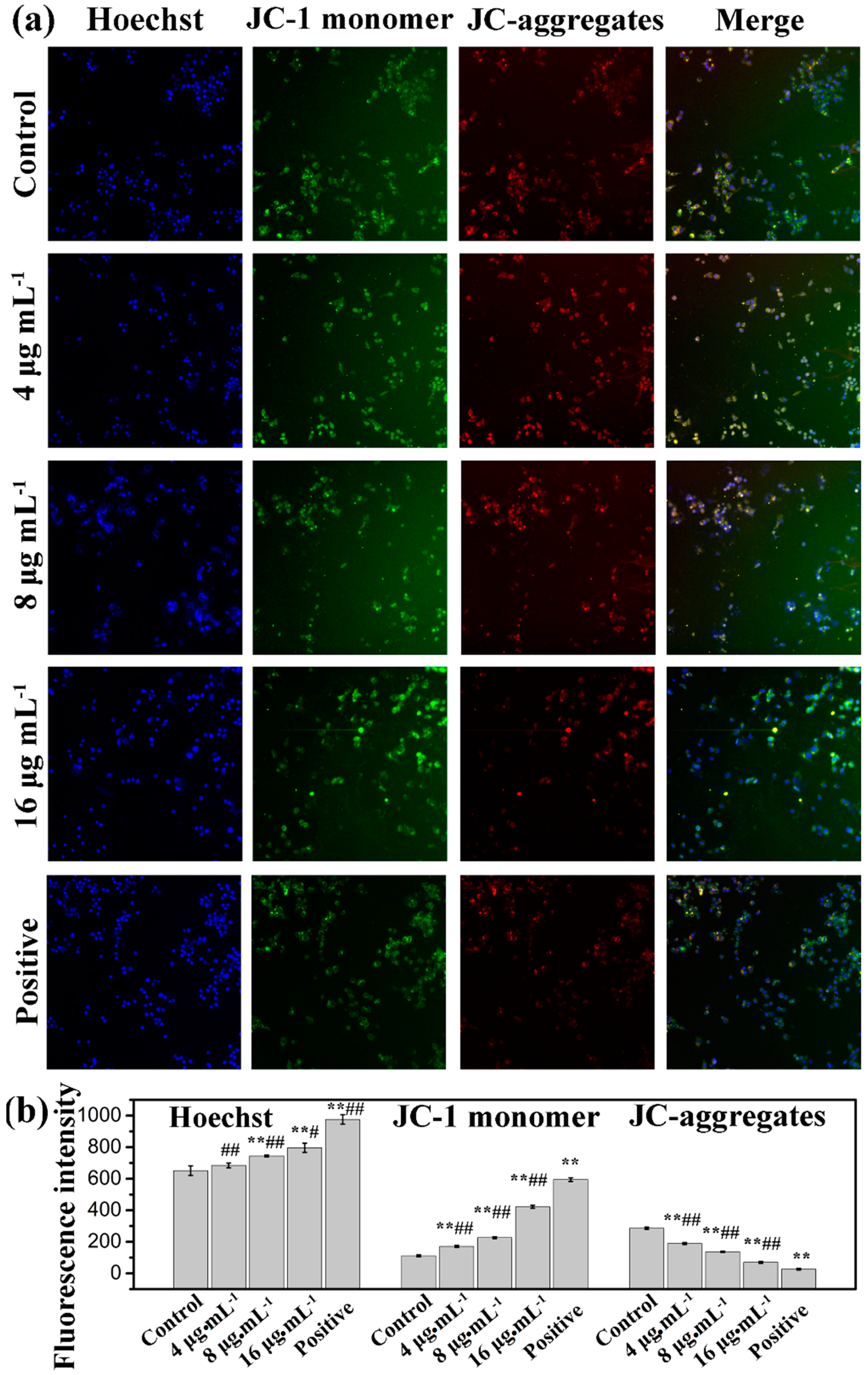
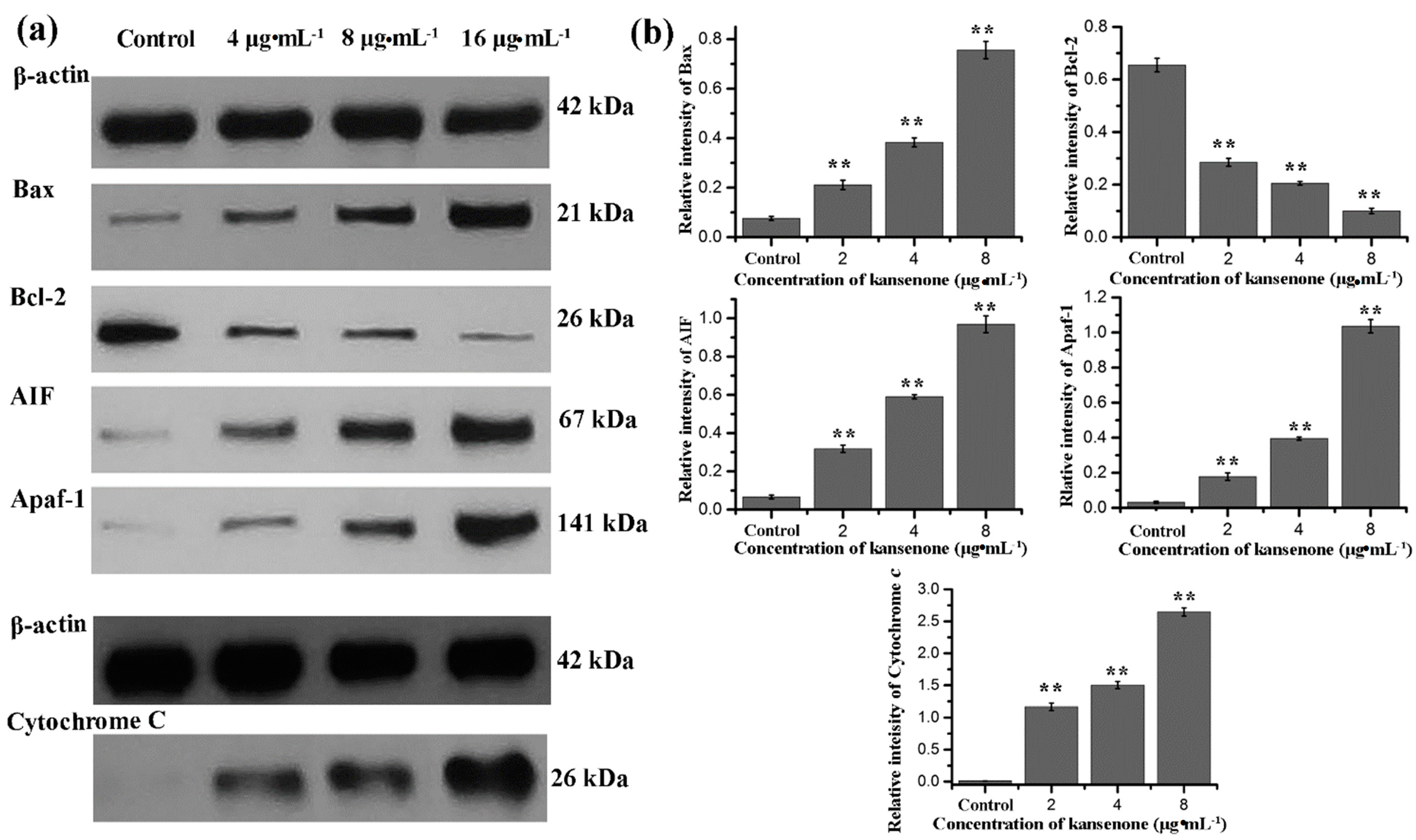

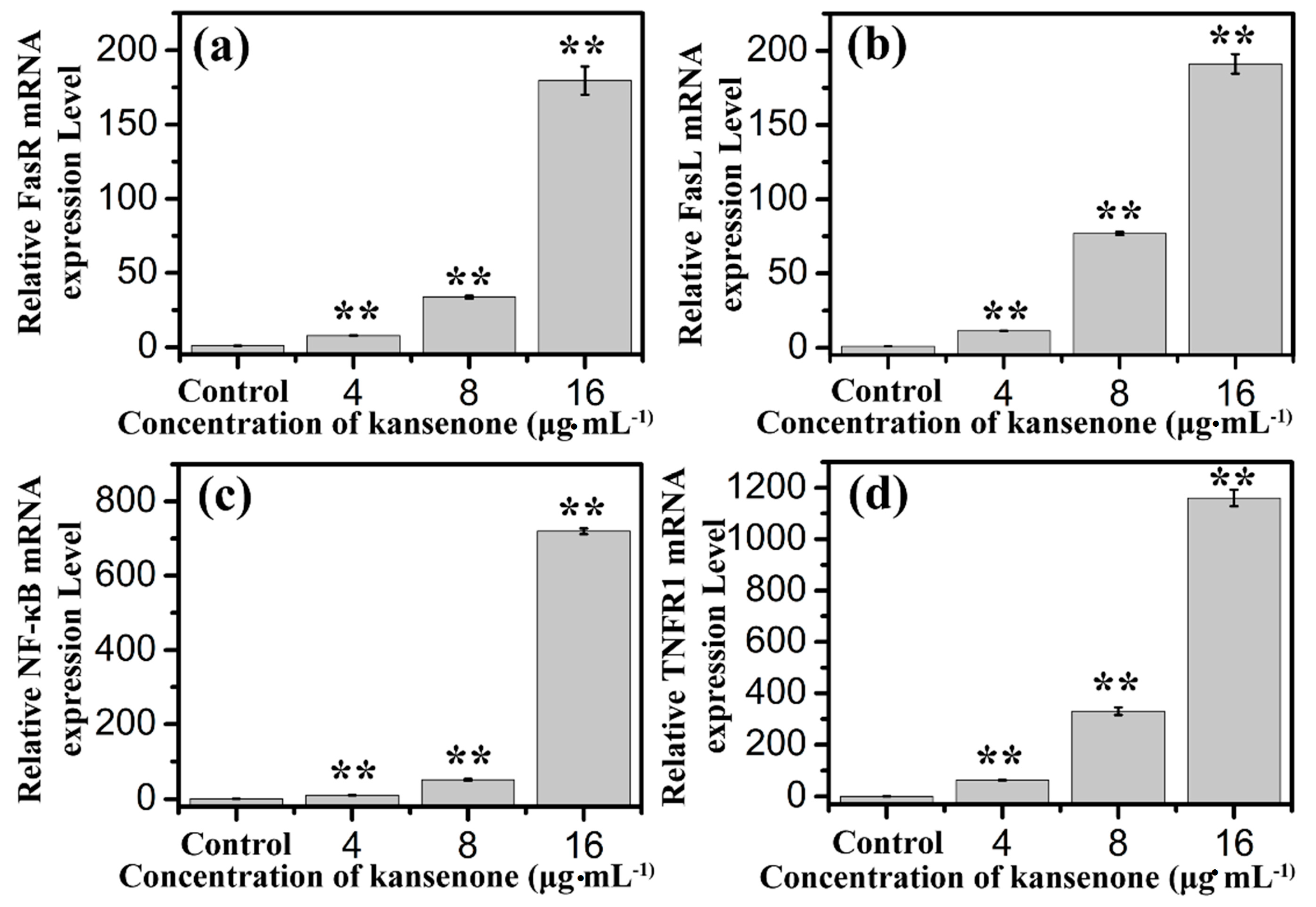
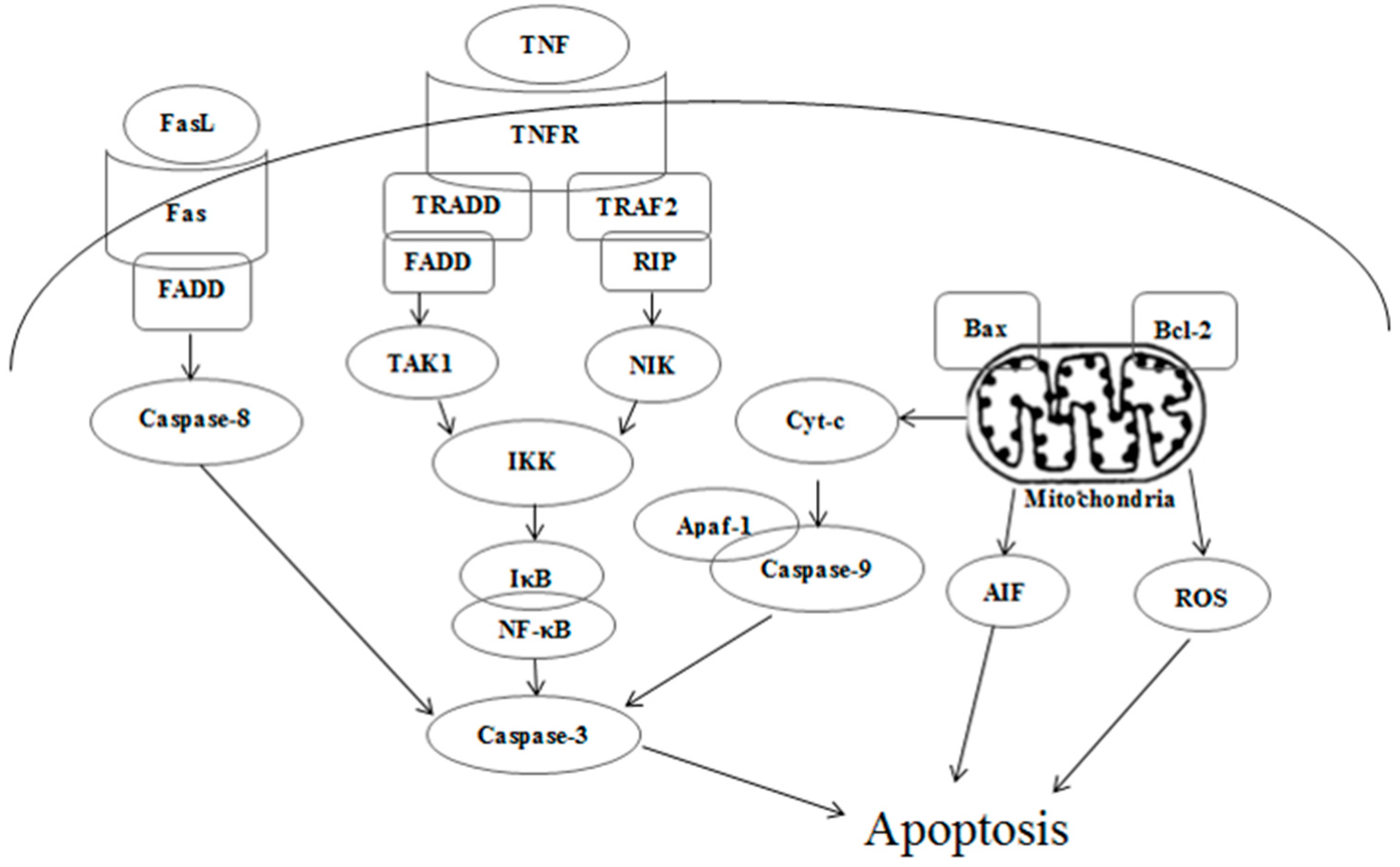
3. Experimental Section
3.1. Chemicals and Regents
3.2. Sample Preparation
3.3. Cell Culture and Cytotoxicity Assays
3.4. Cell Morphology Analysis
3.5. Measurement of Intracellular Reactive Oxygen Species
3.6. Measurement of SOD Activity, MDA Contents, Caspase-3, Caspase-8, and Caspase-9
3.7. Cell Cycle Analysis
3.8. Cell Apoptosis Analysis
3.9. Measurement of the Mitochondrial Membrane Potential (ΔΨm)
3.10. RNA Isolation and Real-Time PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA | Sense (5ʹ→3ʹ) | Antisense (5ʹ→3ʹ) |
|---|---|---|
| Rat-GAPDH | TCAAGAAGGTGGTGAAGCAG | AGGTGGAAGAATGGGAGTTG |
| Rat-Fas | AAGATGCAGCTGAGCAGAAA | GGATTAAAGCTTGACACGCA |
| Rat-FasL | CACAAGGTCCAACAGGTCAG | TTCTCTTTGCCTCTGCATTG |
| Rat-TNFR1 | CCCGTCTTCGGTCCTAGTAA | GTTGAGGGATCCGTAGAGGA |
| Rat-NF-κB | GTGTTCACAGACCTGGCATC | TTCAGGGTACTCCATCAGCA |
3.11. Western Blot Analysis
3.12. Statistical Analysis
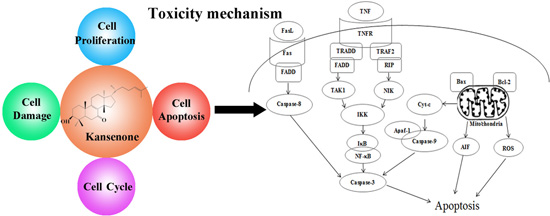
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yan, X.J.; Zhang, L.; Guo, J.M.; Cao, Y.D.; Shang, E.X.; Tang, Y.P.; Ding, A.W.; Duan, J.A. Processing of kansui roots stir-baked with vinegar reduces kansui-induced hepatocyte cytotoxicity by decreasing the contents of toxic terpenoids and regulating the cell apoptosis pathway. Molecules 2014, 19, 7237–7254. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Wang, J.S.; Wei, D.D.; Wang, X.B.; Luo, J.; Yang, M.H.; Kong, L.Y. Bioactivity-guided isolation of antiproliferative diterpenoids from Euphorbia kansui. Phytother. Res. 2012, 26, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Zhong, J.M.; Ye, S.Q.; Ni, Z.Y.; Miao, X.Q.; Mo, Y.K.; Li, Z.L. Screening of Epstein-Barr virus early antigen expression inducers from Chinese medicinal herbs and plants. Biomed. Environ. Sci. 1994, 7, 50–55. [Google Scholar] [PubMed]
- Zheng, W.F.; Cui, Z.; Zhu, Q. Cytotoxicity and antiviral activity of the compounds from Euphorbia kansui. Planta Med. 1998, 64, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Nunomura, S.; Kitanaka, S.; Ra, C. 3-O-(2,3-dimethylbutanoyl)-13-O-decan-oylingenolfrom Euphorbia kansui suppresses IgE-mediated mast cell activation. Biol. Pharm. Bull. 2006, 29, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Khiev, P.; Kim, J.W.; Sung, S.J.; Song, H.H.; Choung, D.H.; Chin, Y.W.; Lee, H.; Oh, S.R. Ingenane-type Diterpenes with a Modulatory Effect on IFN-γ Production from the Roots of Euphorbia kansui. Arch. Pharm. Res. 2012, 35, 1553–1558. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhou, L.Y.; He, H.P.; Leng, Y.; Yang, Z.; Hao, X.J. Inhibition of 11b-HSD1 by tetracyclic triterpenoids from Euphorbia kansui. Molecules 2012, 17, 1826–11838. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.W.; Ding, J.J.; Wu, F.H.; Chen, L.; Yang, Y.X.; Song, F.Y. 1HNMR-based metabonomics study of the urinary biochemical changes in kansui treated rat. J. Ethnopharmacol. 2012, 141, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.W.; Ding, J.J.; Yang, Y.X.; Wu, F.H.; Song, F.Y. Systems biochemical responses of rats to kansui and vinegar-processed kansui exposure by integrated metabonomics. J. Ethnopharmacol. 2014, 153, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.S.; Lee, S.W.; Park, M.H.; Kim, M.S.; Hudson, B.I.; Park, S.J.; Lee, W.S. Kansuinine A and Kansuinine B from Euphorbia kansui L. inhibit IL-6-induced Stat3 activation. Planta Med. 2010, 76, 1544–1549. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Wang, N.L.; Yao, X.S.; Miyata, S.; Kitanaka, S. Euphane and tirucallane triterpenes from the roots of Euphorbia kansui and their in vitro effects on the cell division of Xenopus. J. Nat. Prod. 2003, 66, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; He, H.P.; Fang, X. Kansuinone, a novel euphane-type triterpene from Euphorbia kansui. Tetrahedron Lett. 2010, 51, 6286–6289. [Google Scholar] [CrossRef]
- Pan, Q.; Ip, F.C.; Ip, N.Y.; Zhu, H.X.; Min, Z.D. Activity of macrocyclic jatrophane diterpenes from Euphorbia kansui in a TrkA fibroblast survival assay. J. Nat. Prod. 2004, 67, 1548–1551. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.S.; Lin, Y.M.; Haruna, M.; Pan, D.J.; Shingu, T; Chen, T.P.; Hsu, H.Y.; Nakano, T.; Lee, K.H. Antitumor agent, 119 kansuiphorins A and B, two novel antileukemic diterpene ester from Euphorbia kansui. J. Nat. Prod. 1991, 54, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Dang, Q.L.; Choi, Y.H.; Choi, G.J.; Jang, K.S.; Park, M.S.; Park, N.J.; Lim, C.H.; Kim, H.; Ngoc, L.H.; Kim, J.C. Pesticidal activity of ingenane diterpenes isolated from Euphorbia kansui against Nilaparvata lugens and Tetranychus urticae. J. Asia Pac. Entomol. 2010, 13, 51–54. [Google Scholar] [CrossRef]
- Zhang, L.; Gao, L.; Li, Z.J.; Yan, X.J.; Yang, Y.J.; Tang, Y.P.; Cao, Y.D.; Ding, A.W. Bio-Guided isolation of the cytotoxic terpenoids from the roots of Euphorbia kansui against human normal cell lines L-O2 and GES-1. Int. J. Mol. Sci. 2012, 13, 11247–11259. [Google Scholar] [CrossRef] [PubMed]
- Matthews, G.M.; Newbold, A.; Johnstone, R.W. Intrinsic and extrinsic apoptotic pathway signaling as determinants of histone deacetylase inhibitor antitumor activity. Adv. Cancer Res. 2012, 116, 165–197. [Google Scholar] [PubMed]
- Zhai, D.; Jin, C.; Huang, Z.; Satterthwait, A.C.; Reed, J.C. Differential regulation of Bax and Bak by anti-apoptotic Bcl-2 family proteins Bcl-B and Mcl-1. J. Biol. Chem. 2008, 283, 9580–9586. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Dixit, V.M.D. Apoptosis control by death and decoy receptors. Curr. Opin. Cell Biol. 1999, 11, 255–260. [Google Scholar] [CrossRef]
- Consuelo, G.; Faustin, M. Lipid rafts and raft-mediated supramolecular entities in the regulation of CD95 death receptor apoptotic signaling. Apoptosis 2015, 20, 584–606. [Google Scholar]
- Cullen, S.P.; Martin, S.J. Fas and TRAIL death receptors as initiators of inflammation: Implications for cancer. Semin. Cell Dev. Biol. 2015, 39, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Reyes, F.J.; Centelles, J.J.; Lupiáñez, J.A.; Cascante, M. (2α,3β)-2,3-Dihydroxyolean-12-en-28-oic acid, a new natural triterpene from Olea europea, induces caspase dependent apoptosis selectively in colon adenocarcinoma cells. FEBS Lett. 2006, 580, 6302–6310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Yang, Z.H.; Li, R.J.; Geng, H.; Dong, C. Investigation of fine chalk dust particles’ chemical compositions and toxicities on alveolar macrophages in vitro. Chemosphere 2015, 120, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Elledge, S.J. Cell Cycle Checkpoints: Preventing an Identity Crisis. Science 1996, 274, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Mlynarczyk, C.; Fåhraeus, R. Endoplasmic reticulum stress sensitizes cells to DNA damage-induced apoptosis through p53-dependent suppression of p21CDKN1A. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Ma, B.Y.; Wang, Y.G.; Xiao, R.J.; Kong, Y.P.; Zhou, X.M.; Xia, D.J. Targeting gene-virus-mediated manganese superoxide dismutase effectively suppresses tumor growth in hepatocellular carcinoma in vitro and in vivo. Cancer Biother. Radiopharm. 2014, 29, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qin, C.X.; Shu, X.Q.; Chen, R.; Song, H.H.; Li, Q.; Xu, H. 2-Effects of copper on induction of thiol-compounds and antioxidant enzymes by the fruiting body of Oudemansiella radicata. Ecotox. Environ. Saf. 2015, 111, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.K.; Ghosh, T.; Roy, S.; Chattopadhyay, S.; Das, D. Zinc sulfide nanoparticles selectively induce cytotoxic and genotoxic effects on leukemic cells: Involvement of reactive oxygen species and tumor necrosis factor α. J. Appl. Toxicol. 2014, 34, 1130–1144. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- García, A.; Morales, P.; Arranz, N.; Delgado, M.E.; Rafter, J.; Haza, A.I. Antiapoptotic effects of dietary antioxidants towards N-nitrosopiperidine and N-nitrosodibutylamine-induced apoptosis in HL-60 and HepG2 cells. J. Appl. Toxicol. 2009, 29, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Shin, S.Y.; Lee, M.S.; Koh, D.; Lee, Y.H.; Lim, Y.A. A new synthetic 2-hydroxy-2,4,6-trimethoxy-5′,6′-naphthochalcone induces G2/M cell cycle arrest and apoptosis by disrupting the microtubular network of human colon cancer cells. Cancer Lett. 2014, 354, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.M.; Xu, L.N.; Yin, L.H.; Qi, Y.; Li, H.; Xu, Y.W.; Han, X.; Peng, J.Y.; Wan, X.Y. Cytotoxicity of dioscin in human gastric carcinoma cells through death receptor and mitochondrial pathways. J. Appl. Toxicol. 2011, 33, 712–722. [Google Scholar] [CrossRef] [PubMed]
- You, R.X.; Liu, J.Y.; Li, S.J.; Wang, L.; Wang, K.P.; Zhang, Y. Alkali-soluble polysaccharide, isolated from lentinus edodes, induces apoptosis and G2/M cell cycle arrest in H22 cells through microtubule depolymerization. Phytother. Res. 2014, 28, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.J.; Zhang, N.; Wang, X.L.; Cai, C.; Cun, J.J.; Li, Y.M.; Lv, S.G.; Yang, Q.F. Nitidine chloride induces apoptosis, cell cycle arrest, and synergistic cytotoxicity with doxorubicin in breast cancer cells. Tumor Biol. 2014, 35, 10201–10212. [Google Scholar] [CrossRef] [PubMed]
- Hennet, T.; Bertoni, G.; Richter, C.; Peterhans, E. Expression of BCL-2 protein enhances the survival of mouse fibrosarcoid cells in tumor necrosis factor-mediated cytotoxicity. Cancer Res. 1993, 53, 1456–1460. [Google Scholar] [PubMed]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Calandria, C.; Irurzun, A.; Barco, A.; Carrasco, L. Individual expression of poliovirus 2Apro and 3Cpro induces activation of caspase-3 and PARP cleavage in hela cells. Virus Res. 2004, 104, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Yap, E.; Tan, W.L.; Ng, I.; Ng, Y.K. Combinatorial-approached neuroprotection using pan-caspase inhibitor and poly (ADP-ribose) polymerase (PARP) inhibitor following experimental stroke in rats; is there additional benefit? Brain Res. 2008, 1195, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Vucic, D.; Dixit, V.M.D.; Wertz, I.E. Ubiquitylation in apoptosis: A post-translational modification at the edge of life and death. Nat. Rev. Mol. Cell Biol. 2011, 12, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Lichter, P.; Walczak, H.; Weitz, S.; Behrmann, I.; Krammer, P.H. The human APO-1 (APT) antigen maps to 10q23, a region that is syntenic with mouse chromosome 19. Genomics 1992, 14, 179–180. [Google Scholar] [CrossRef]
- Inazawa, J.; Itoh, N.; Abe, T.; Nagata, S. Assignment of the human Fas antigen gene (Fas) to 10q24.1. Genomics 1992, 14, 821–822. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A.T. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Dash, S.K.; Tripathy, S; Das, B.; Mahapatra, S.K.; Pramanik, P.; Roy, S. Cobalt oxide nanoparticles induced oxidative stress linked to activation of TNF-α/caspase-8/p38-MAPK signaling in human leukemia cells. J. Appl. Toxicol. 2015, 35, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Dong, W.B.; Fu, H.; Li, Q.P.; Deng, C.L.; Lei, X.P.; Guo, L. Involvement of the nuclear factor-κB pathway in the adhesion of neutrophils to renal tubular cells after injury induced by neonatal postasphyxial serum. Mol. Cell. Biochem. 2014, 388, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, F.; Yang, Y.; Zhang, L.; Cao, Y.; Yao, W.; Tang, Y.; Ding, A. A Natural Triterpene Derivative from Euphorbia kansui Inhibits Cell Proliferation and Induces Apoptosis against Rat Intestinal Epithelioid Cell Line in Vitro. Int. J. Mol. Sci. 2015, 16, 18956-18975. https://doi.org/10.3390/ijms160818956
Cheng F, Yang Y, Zhang L, Cao Y, Yao W, Tang Y, Ding A. A Natural Triterpene Derivative from Euphorbia kansui Inhibits Cell Proliferation and Induces Apoptosis against Rat Intestinal Epithelioid Cell Line in Vitro. International Journal of Molecular Sciences. 2015; 16(8):18956-18975. https://doi.org/10.3390/ijms160818956
Chicago/Turabian StyleCheng, Fangfang, Yanjing Yang, Li Zhang, Yudan Cao, Weifeng Yao, Yuping Tang, and Anwei Ding. 2015. "A Natural Triterpene Derivative from Euphorbia kansui Inhibits Cell Proliferation and Induces Apoptosis against Rat Intestinal Epithelioid Cell Line in Vitro" International Journal of Molecular Sciences 16, no. 8: 18956-18975. https://doi.org/10.3390/ijms160818956
APA StyleCheng, F., Yang, Y., Zhang, L., Cao, Y., Yao, W., Tang, Y., & Ding, A. (2015). A Natural Triterpene Derivative from Euphorbia kansui Inhibits Cell Proliferation and Induces Apoptosis against Rat Intestinal Epithelioid Cell Line in Vitro. International Journal of Molecular Sciences, 16(8), 18956-18975. https://doi.org/10.3390/ijms160818956