
Gene Expression Signature in Endemic Osteoarthritis by Microarray Analysis


 and
and
Abstract
:1. Introduction
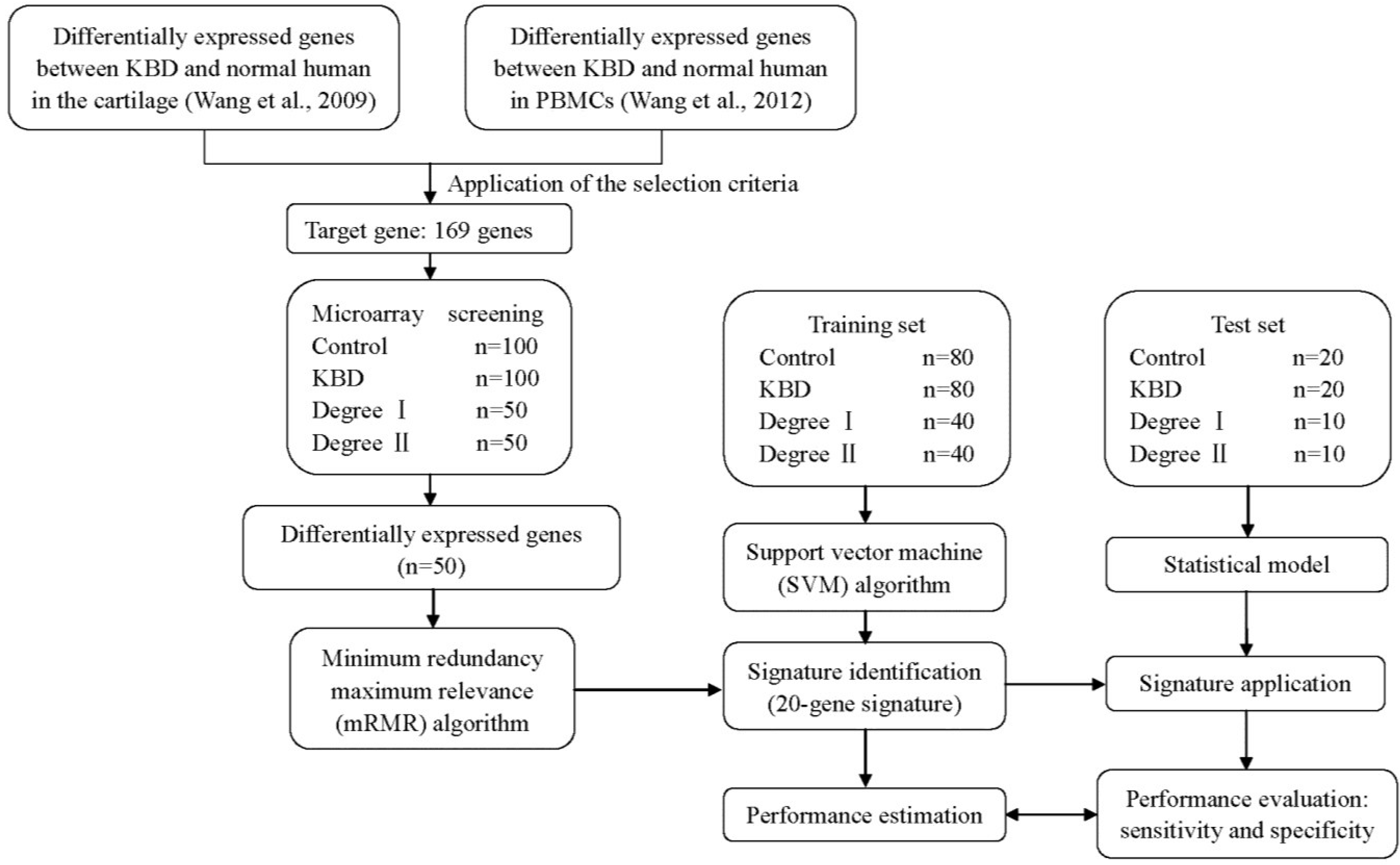
2. Results
2.1. Differentially Expressed Genes
2.2. Identification of a 20-Gene Signature
2.3. Statistical Analysis of the KBD Degree with the 20-Gene Signature

{kind=link}
{kind=link}
| Gene Name | Symbol | Public ID | Fold Change b |
|---|---|---|---|
| down-regulated genes | |||
| ATP-binding cassette, sub-family C, member 13, pseudogene | ABCC13 | NR_003087 | 0.42 ± 0.03 |
| ABI family, member 3 (NESH) binding protein | ABI3BP | NM_015429 | 0.33 ± 0.03 |
| Branched chain amino-acid transaminase 1, cytosolic | BCAT1 | NM_001178091 | 0.43 ± 0.04 |
| Calcium channel, voltage-dependent, gamma subunit 6 | CACNG6 | NM_145814 | 0.39 ± 0.02 |
| Chondroitin sulfate N-acetylgalactosaminyltransferase 1 | CSGALNACT1 | NM_001130518 | 0.21 ± 0.03 |
| Cathepsin C | CTSC | NM_001114173 | 0.39 ± 0.02 |
| Cytochrome b5 reductase 3 | CYB5R3 | NM_000398 | 0.48 ± 0.04 |
| Dystrophin, muscular dystrophy | DMD | NM_007868 | 0.37 ± 0.02 |
| Enhancer of rudimentary homolog (Drosophila) | ERH | NM_004450 | 0.38 ± 0.02 |
| F11 receptor | F11R | NM_016946 | 0.46 ± 0.02 |
| FK506 binding protein 9, 63 kDa | FKBP9 | NM_007270 | 0.49 ± 0.06 |
| Frizzled family receptor 1 | FZD1 | NM_003505 | 0.47 ± 0.03 |
| Growth differentiation factor 5 | GDF5 | NM_000557 | 0.44 ± 0.03 |
| Hemoglobin, alpha 2 | HBA2 | NM_000517 | 0.49 ± 0.03 |
| Zinc family member 5 | ZIC5 | NM_033132 | 0.38 ± 0.02 |
| up-regulated genes | |||
| Baculoviral IAP repeat containing 3 | BIRC3 | NM_001165 | 4.26 ± 0.35 |
| FGFR1 oncogene partner 2 | FGFR1OP2 | NM_001171887 | 2.39 ± 0.15 |
| Sialic acid binding lg-like lectin 8 | SIGLEC8 | NM_014442 | 2.50 ± 0.29 |
| Single-stranded DNA binding protein 1, mitochondrial | SSBP1 | NM_001256510 | 3.12 ± 0.25 |
| Tetratricopeptide repeat domain 25 | TTC25 | NM_031421 | 3.19 ± 0.25 |
| Diagnosed Group Membership | Predicted Group Membership | Total | |||
|---|---|---|---|---|---|
| KBD Degree I | KBD Degree II | ||||
| Original | Count | Degree I | 46 | 4 | 50 |
| Degree II | 5 | 45 | 50 | ||
| % | Degree I | 92.0 | 8.0 | 100.0 | |
| Degree II | 10.0 | 90.0 | 100.0 | ||
| Cross-validated a | Count | Degree I | 41 | 9 | 50 |
| Degree II | 9 | 41 | 50 | ||
| % | Degree I | 82.0 | 18.0 | 100.0 | |
| Degree II | 18.0 | 82.0 | 100.0 | ||
2.4. Quantitative RT-PCR Analyses
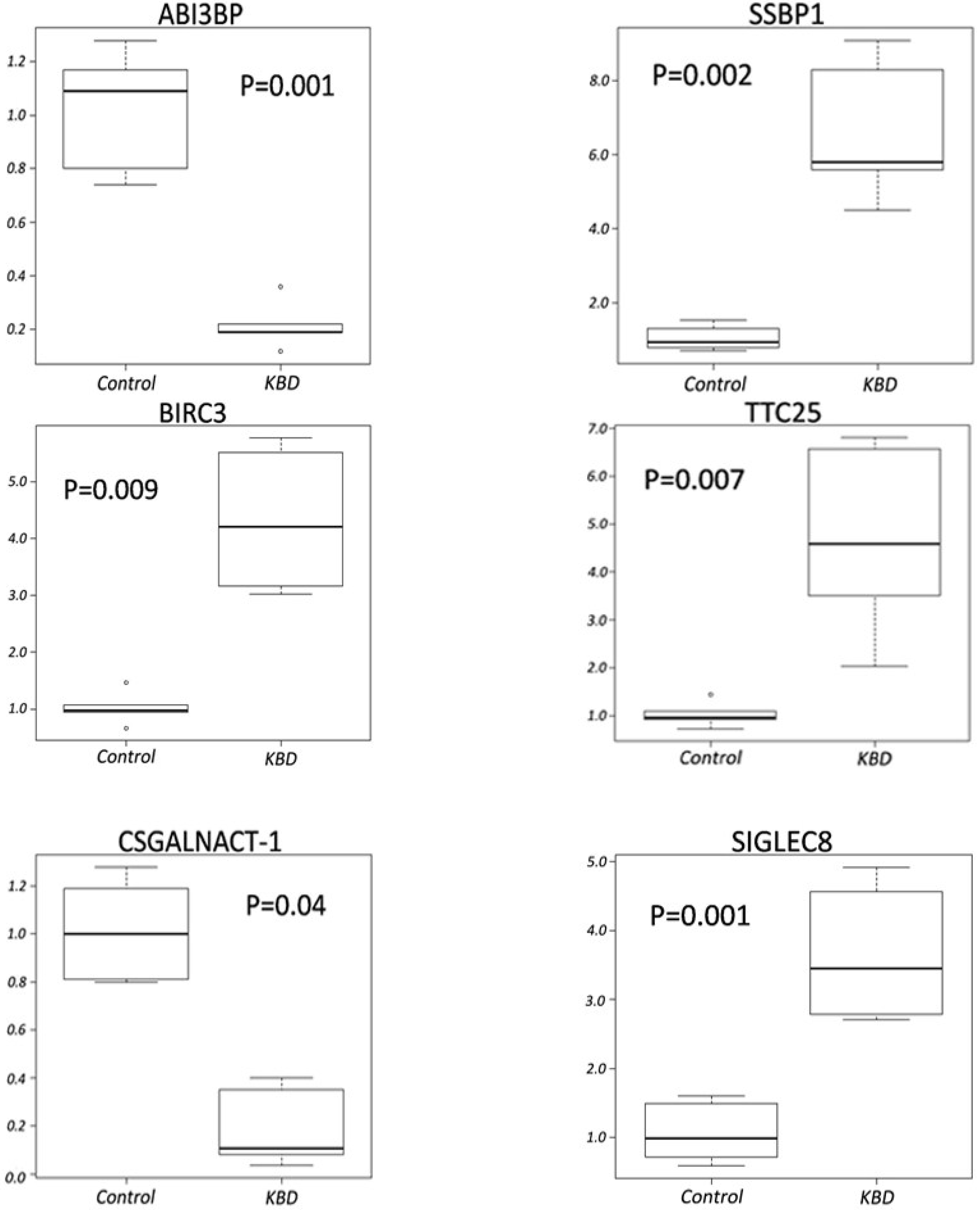
3. Discussion
4. Experimental Section
4.1. Ethics Statement
4.2. Patients and Study Design
| Variable | Microarray | Signature Identification | ||||
|---|---|---|---|---|---|---|
| KBD | Control | Training Set | Test Set | |||
| (N = 100) | (N = 100) | KBD | Control | KBD | Control | |
| (N = 80) | (N = 80) | (N = 20) | (N = 20) | |||
| Age, year | ||||||
| mean | 57.91 | 53.43 | 57.50 | 53.20 | 56.20 | 52.34 |
| range | 43–79 | 40–77 | 43–79 | 40–77 | 43–75 | 46–72 |
| Gender, N (%) | ||||||
| Male | 44 (44.0) | 44 (44.0) | 37 (46.2) | 35 (43.7) | 7 (35.0) | 9 (45.0) |
| Female | 56 (56.0) | 56 (56.0) | 43 (53.8) | 45 (56.3) | 13 (65.0) | 11 (55.0) |
| KBD Degree, N (%) | ||||||
| Degree I | 50 (50.0) | - | 38 (47.5) | - | 12 (60.0) | - |
| Degree II | 50 (50.0) | 42 (52.5) | 8 (40.0) | |||
| Clinical Sign, N (%) | ||||||
| EP a | 95 (95.0%) | 77 (96.2%) | 18 (90.0%) | |||
| Brachydactylia | 53 (53.0%) | 45 (56.2%) | 8 (40.0%) | |||
| Variable | KBD | Normal | ||||||
|---|---|---|---|---|---|---|---|---|
| n | Age (Years) | Male | Female | n | Age (Years) | Male | Female | |
| 1 | 1 | 69 | 1 | 0 | 1 | 65 | 1 | 0 |
| 2 | 1 | 62 | 1 | 0 | 1 | 56 | 1 | 0 |
| 3 | 1 | 57 | 1 | 0 | 1 | 56 | 1 | 0 |
| 4 | 1 | 58 | 0 | 1 | 1 | 55 | 0 | 1 |
| 5 | 1 | 52 | 0 | 1 | 1 | 48 | 0 | 1 |
| Total | 5 | 59.6 | 3 | 2 | 5 | 56.0 | 3 | 2 |
4.3. Selection of the Target Genes
4.4. Blood Collection and the PBMC Isolation
4.5. RNA Preparation
4.6. Microarray Hybridization
4.7. Gene Expression Analysis
4.8. Statistical Analysis
4.9. Quantitative Real-Time PCR Validation
| Symbol | 5' Primer Sequence | 3' Primer Sequence | Amplicon Size (bp) |
|---|---|---|---|
| ABI3BP | GAAGATCACTGCCAGTTTGTGGA | CCTGGCGAACTGCTCTGAAATA | 109 |
| BIRC3 | GACTCAGGTGTTGGGAATCTGGA | TGAGGGTAACTGGCTTGAACTTGAC | 127 |
| CSGALNACT1 | CAGCTCTTGCTGCTGCTGTG | AAGGATGATCTTGCAGGCAGAA | 139 |
| SIGLEC8 | CAGGTGTGACCACGACCAGTA | ACTGGCCCTCAAGGACTGAA | 141 |
| SSBP1 | CCTCATCAGATGTGCAGGAATGTT | TGACCCACTCGCCCAAGTAAG | 190 |
| TTC25 | AGATCGGCCGCTGCTACTTG | CCACCAGAACACTGGCATTCA | 126 |
| β-ACTIN | CGGAGTCAACGGATTTGGTCGTAT | AGCCTTCTCCATGGTGGTGAAGAC | 120 |
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Duan, C.; Guo, X.; Zhang, X.D.; Yu, H.J.; Yan, H.; Gao, Y.; Ma, W.J.; Gao, Z.Q.; Xu, P.; Lammi, M. Comparative analysis of gene expression profiles between primary knee osteoarthritis and an osteoarthritis endemic to Northwestern China, Kashin-Beck disease. Arthritis Rheumatol. 2010, 62, 771–780. [Google Scholar] [CrossRef]
- Fang, H.; Guo, X.; Farooq, U.; Xia, C.; Dong, R. Development and validation of a quality of life instrument for Kashin-Beck disease: An endemic osteoarthritis in China. Osteoarthr. Cartil. 2012, 20, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Zou, K.; Liu, G.; Wu, T.; Du, L. Selenium for preventing Kashin-Beck osteoarthropathy in children: A meta-analysis. Osteoarthr. Cartil. 2009, 17, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Chasseur, C.; Suetens, C.; Nolard, N.; Begaux, F.; Haubruge, E. Fungal contamination in barley and Kashin-Beck disease in Tibet. Lancet 1997, 350, 1074. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ma, W.J.; Zhang, F.; Ren, F.L.; Qu, C.J.; Lammi, M.J. Recent advances in the research of an endemic osteochondropathy in China: Kashin-Beck disease. Osteoarthr. Cartil. 2014, 22, 1774–1783. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Liew, C.C. Gene profiling identifies secreted protein transcripts from peripheral blood cells in coronary artery disease. J. Mol. Cell. Cardiol. 2003, 35, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.W.; Zhang, H.; Yager, T.D.; Nossova, N.; Dempsey, A.; Zheng, R.; Han, M.; Tang, H.; Chao, S.; Liew, C.C. Blood-based biomarkers for detecting mild osteoarthritis in the human knee. Osteoarthr. Cartil. 2005, 13, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Guo, X.; Wang, W.; Yan, H.; Li, C. Genome-wide gene expression analysis suggests an important role of hypoxia in the pathogenesis of endemic osteochondropathy Kashin-Beck disease. PLoS ONE 2011, 6, e22983. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Guo, X.; Wu, X.M.; Lammi, M.J. Genome-wide gene expression analysis suggests an important role of suppressed immunity in pathogenesis of Kashin-Beck disease. PLoS ONE 2012, 7, e28439. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Toyoda, H.; Tanahashi, T.; Tanaka, J.; Kumada, T.; Yoshioka, Y.; Kosaka, N.; Ochiya, T.; Taguchi, Y.H. Comprehensive miRNA expression analysis in peripheral blood can diagnose liver disease. PLoS ONE 2012, 7, e48366. [Google Scholar] [CrossRef] [PubMed]
- Ishimura, M.; Yamamoto, H.; Mizuno, Y.; Takada, H.; Goto, M.; Doi, T.; Hoshina, T.; Ohga, S.; Ohshima, K.; Hara, T. A non-invasive diagnosis of histiocytic necrotizing lymphadenitis by means of gene expression profile analysis of peripheral blood mononuclear cells. J. Clin. Immunol. 2013, 33, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Z.; Guo, X.; Duan, C.; Ma, W.J.; Zhang, Y.G.; Xu, P.; Gao, Z.Q.; Wang, Z.F.; Yan, H.; Zhang, Y.F.; et al. Comparative analysis of gene expression profiles between the normal human cartilage and the one with endemic osteoarthritis. Osteoarthr. Cartil. 2009, 17, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Tsuang, M.T.; Nossova, N.; Yager, T.; Tsuang, M.M.; Guo, S.C.; Shyu, K.G.; Glatt, S.J.; Liew, C.C. Assessing the validity of blood-based gene expression profiles for the classification of schizophrenia and bipolar disorder: A preliminary report. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2005, 133B, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.G.; Aronow, B.J.; Luyrink, L.K.; Moroldo, M.B.; Pavlidis, P.; Passo, M.H.; Grom, A.A.; Hirsch, R.; Giannini, E.H.; Colbert, R.A.; et al. Gene expression in juvenile arthritis and spondyloarthropathy: Pro-angiogenic ELR+ chemokine genes relate to course of arthritis. Rheumatology 2004, 43, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Niimoto, T.; Nakasa, T.; Ishikawa, M.; Okuhara, A.; Izumi, B.; Deie, M.; Suzuki, O.; Adachi, N.; Ochi, M. MicroRNA-146a expresses in interleukin-17 producing T cells in rheumatoid arthritis patients. BMC Musculoskelet. Disord. 2010, 11, 209. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.S.; Che, M.X. Calcium channel of osteoblast. Chin. J. Clin. Oncol. Rehabil. 2009, 9, 120–122. [Google Scholar]
- Guggino, S.E.; Wagner, J.A.; Snowman, A.M.; Hester, L.D.; Sacktor, B.; Snyder, S.H. Phenylalkylamine-sensitive calcium channels in osteoblast-like osteosarcoma cells. Characterization by ligand binding and single channel recordings. J. Biol. Chem. 1988, 263, 10155–10161. [Google Scholar] [PubMed]
- Wang, X.; Wang, S.; He, S.; Zhang, F.; Tan, W.; Lei, Y.; Yu, H.; Li, Z.; Ning, Y.; Xiang, Y.; et al. Comparing gene expression profiles of Kashin-Beck and Keshan diseases occurring within the same endemic areas of China. Sci. China Life Sci. 2013, 56, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Gyrd-Hansen, M.; Meier, P. IAPs: From caspase inhibitors to modulators of NF-κB, inflammation and cancer. Nat. Rev. Cancer 2010, 10, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.M.; Zammit, N.W.; Yam, A.O.; Slattery, R.; Walters, S.N.; Malle, E.; Grey, S.T. Baculoviral inhibitors of apoptosis repeat containing (BIRC) proteins fine-tune TNF-induced nuclear factor κB and c-Jun N-terminal kinase signalling in mouse pancreatic beta cells. Diabetologia 2013, 56, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Rothe, M.; Pan, M.G.; Henzel, W.J.; Ayres, T.M.; Goeddel, D.V. The TNFR2-TRAF signaling complex contains two novel proteins related to baculoviral-inhibitor of apoptosis proteins. Cell 1995, 83, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Hoang, B.; Thomas, J.T.; Vukicevic, S.; Luyten, F.P.; Ryba, N.J.; Kozak, C.A.; Reddi, A.H.; Moos, M., Jr. Cartilage-derived morphogenetic proteins. New members of the transforming growth factor-β superfamily predominantly expressed in long bones during human embryonic development. J. Biol. Chem. 1994, 269, 28227–28234. [Google Scholar] [PubMed]
- Mikic, B. Multiple effects of GDF-5 deficiency on skeletal tissues: Implications for therapeutic bioengineering. Ann. Biomed. Eng. 2004, 32, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Mabuchi, A.; Shi, D.; Kubo, T.; Takatori, Y.; Saito, S.; Fujioka, M.; Sudo, A.; Uchida, A.; Yamamoto, S.; et al. A functional polymorphism in the 5' UTR of GDF5 is associated with susceptibility to osteoarthritis. Nat. Genet. 2007, 39, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhao, Y.; Yu, M.; Tu, Q.; Wu, L.; Lai, J.; Guo, X. Association between GDF5 gene polymorphism and Kashin-Beck disease. J. Xi’an Jiaotong Univ. (Med. Sci.) 2010, 31, 501–504. [Google Scholar]
- Wolters, P.J.; Chapman, H.A. Importance of lysosomal cysteine proteases in lung disease. Respir. Res. 2000, 1, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Guo, F.; Serebriiskii, I.G.; Howard, A.; Zhang, Y.Z. A 1.55 Å resolution X-ray crystal structure of HEF2/ERH and insights into its transcriptional and cell-cycle interaction networks. Proteins 2007, 68, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Tiranti, V.; Rossi, E.; Ruiz-Carrillo, A.; Rossi, G.; Rocchi, M.; DiDonato, S.; Zuffardi, O.; Zeviani, M. Chromosomal localization of mitochondrial transcription factor A (TCF6), single-stranded DNA-binding protein (SSBP), and endonuclease G (ENDOG), three human housekeeping genes involved in mitochondrial biogenesis. Genomics 1995, 25, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Gong, Z.H.; Ghosal, G.; Chen, J.J. SOSS complexes participate in the maintenance of genomic stability. Mol. Cell 2009, 35, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.T.; Guo, X.; Ma, W.J.; Zhang, Y.G.; Xu, P.; Yao, J.F.; Bai, Y.D. Mitochondrial function is altered in articular chondrocytes of an endemic osteoarthritis, Kashin-Beck disease. Osteoarthr. Cartil. 2010, 18, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Gulberti, S.; Jacquinet, J.C.; Chabel, M.; Ramalanjaona, N.; Magdalou, J.; Netter, P.; Coughtrie, M.W.; Ouzzine, M.; Fournel-Gigleux, S. Chondroitin sulfate N-acetylgalactosaminyltransferase-1 (CSGalNAcT-1) involved in chondroitin sulfate initiation: Impact of sulfation on activity and specificity. Glycobiology 2012, 22, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Kimata, K.; Sato, T.; Gotoh, M.; Narimatsu, H.; Shinomiya, K.; Watanabe, H. Chondroitin sulfate N-acetylgalactosaminyltransferase-1 plays a critical role in chondroitin sulfate synthesis in cartilage. J. Biol. Chem. 2007, 282, 4152–4161. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kudo, T.; Ikehara, Y.; Ogawa, H.; Hirano, T.; Kiyohara, K.; Hagiwara, K.; Togayachi, A.; Ema, M.; Takahashi, S.; et al. Chondroitin sulfate N-acetylgalactosaminyltransferase 1 is necessary for normal endochondral ossification and aggrecan metabolism. J. Biol. Chem. 2011, 286, 5803–5812. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wu, C.; Ma, W.; Zhang, Y.; Hou, T.; Xu, H.; Wu, S.; Yao, X.; Guo, X. Abnormal expression of chondroitin sulphate N-acetylgalactosaminyltransferase 1 and Hapln-1 in cartilage with Kashin-Beck disease and primary osteoarthritis. Int. Orthop. 2013, 37, 2051–2059. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Tan, L.; Goldring, M.B.; Olsen, B.R.; Li, Y. Expression of frizzled genes in mouse costochondral chondrocytes. Matrix Biol. 2001, 20, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Lories, R.J.; Corr, M.; Lane, N.E. To Wnt or not to Wnt: The bone and joint health dilemma. Nat. Rev. Rheumatol. 2013, 9, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Qian, Y.; Wei, L.; Abraham, J.; Shi, X.; Castranova, V.; Harner, E.J.; Flynn, D.C.; Guo, L. Population-based molecular prognosis of breast cancer by transcriptional profiling. Clin. Cancer Res. 2007, 13, 2014–2022. [Google Scholar] [CrossRef] [PubMed]
- Prasad, N.B.; Somervell, H.; Tufano, R.P.; Dackiw, A.P.; Marohn, M.R.; Califano, J.A.; Wang, Y.; Westra, W.H.; Clark, D.P.; Umbricht, C.B.; et al. Identification of genes differentially expressed in benign versus versus malignant thyroid tumors. Clin. Cancer Res. 2008, 14, 3327–3337. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xu, Q.; Yang, L.; Ye, X.; Liu, F.; Wu, F.; Ni, S.; Tan, C.; Cai, G.; Meng, X.; et al. Identification and validation of a blood-based 18-gene expression signature in colorectal cancer. Clin. Cancer Res. 2013, 19, 3039–3049. [Google Scholar]
- Wen, Y.; Zhang, F.; Li, C.; He, S.; Tan, W.; Lei, Y.; Zhang, Q.; Yu, H.; Zheng, J.; Guo, X. Gene expression analysis suggests bone development-related genes GDF5 and DIO2 are involved in the development of Kashin-Beck disease in children rather than adults. PLoS ONE 2014, 9, e103618. [Google Scholar] [CrossRef] [PubMed]
- Hinsenkamp, M. Kashin-Beck disease. Int. Orthop. 2001, 25, 133. [Google Scholar] [CrossRef] [PubMed]
- Felson, D.T.; Lawrence, R.C.; Dieppe, P.A.; Hirsch, R.; Helmick, C.G.; Jordan, J.M.; Kington, R.S.; Lane, N.E.; Nevitt, M.C.; Zhang, Y.; et al. Osteoarthritis: New insights. Part 1: The disease and its risk factors. Ann. Intern. Med. 2000, 133, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, B.; Gronthos, S.; Kuliwaba, J.S.; Robey, P.G.; Findlay, D.M.; Fazzalari, N.L. Identification of differentially expressed genes between osteoarthritic and normal trabecular bone from the intertrochanteric region of the proximal femur using cDNA microarray analysis. Bone 2005, 36, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Peng, H. Minimum redundancy feature selection from microarray gene expression data. J. Bioinform. Comput. Biol. 2005, 3, 185–205. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Lin, C.J. LIBSVM: A library for support vector machines. ACM Trans. Intell. Syst. Technol. 2011, 2, 21–28. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Ning, Y.; Zhang, F.; Yu, F.; Tan, W.; Lei, Y.; Wu, C.; Zheng, J.; Wang, S.; Yu, H.; et al. Gene Expression Signature in Endemic Osteoarthritis by Microarray Analysis. Int. J. Mol. Sci. 2015, 16, 11465-11481. https://doi.org/10.3390/ijms160511465
Wang X, Ning Y, Zhang F, Yu F, Tan W, Lei Y, Wu C, Zheng J, Wang S, Yu H, et al. Gene Expression Signature in Endemic Osteoarthritis by Microarray Analysis. International Journal of Molecular Sciences. 2015; 16(5):11465-11481. https://doi.org/10.3390/ijms160511465
Chicago/Turabian StyleWang, Xi, Yujie Ning, Feng Zhang, Fangfang Yu, Wuhong Tan, Yanxia Lei, Cuiyan Wu, Jingjing Zheng, Sen Wang, Hanjie Yu, and et al. 2015. "Gene Expression Signature in Endemic Osteoarthritis by Microarray Analysis" International Journal of Molecular Sciences 16, no. 5: 11465-11481. https://doi.org/10.3390/ijms160511465
APA StyleWang, X., Ning, Y., Zhang, F., Yu, F., Tan, W., Lei, Y., Wu, C., Zheng, J., Wang, S., Yu, H., Li, Z., Lammi, M. J., & Guo, X. (2015). Gene Expression Signature in Endemic Osteoarthritis by Microarray Analysis. International Journal of Molecular Sciences, 16(5), 11465-11481. https://doi.org/10.3390/ijms160511465