
Transcriptional Responses of Glutathione Transferase Genes in Ruditapes philippinarum Exposed to Microcystin-LR


 and
and 
Abstract
:1. Introduction
2. Results and Discussion
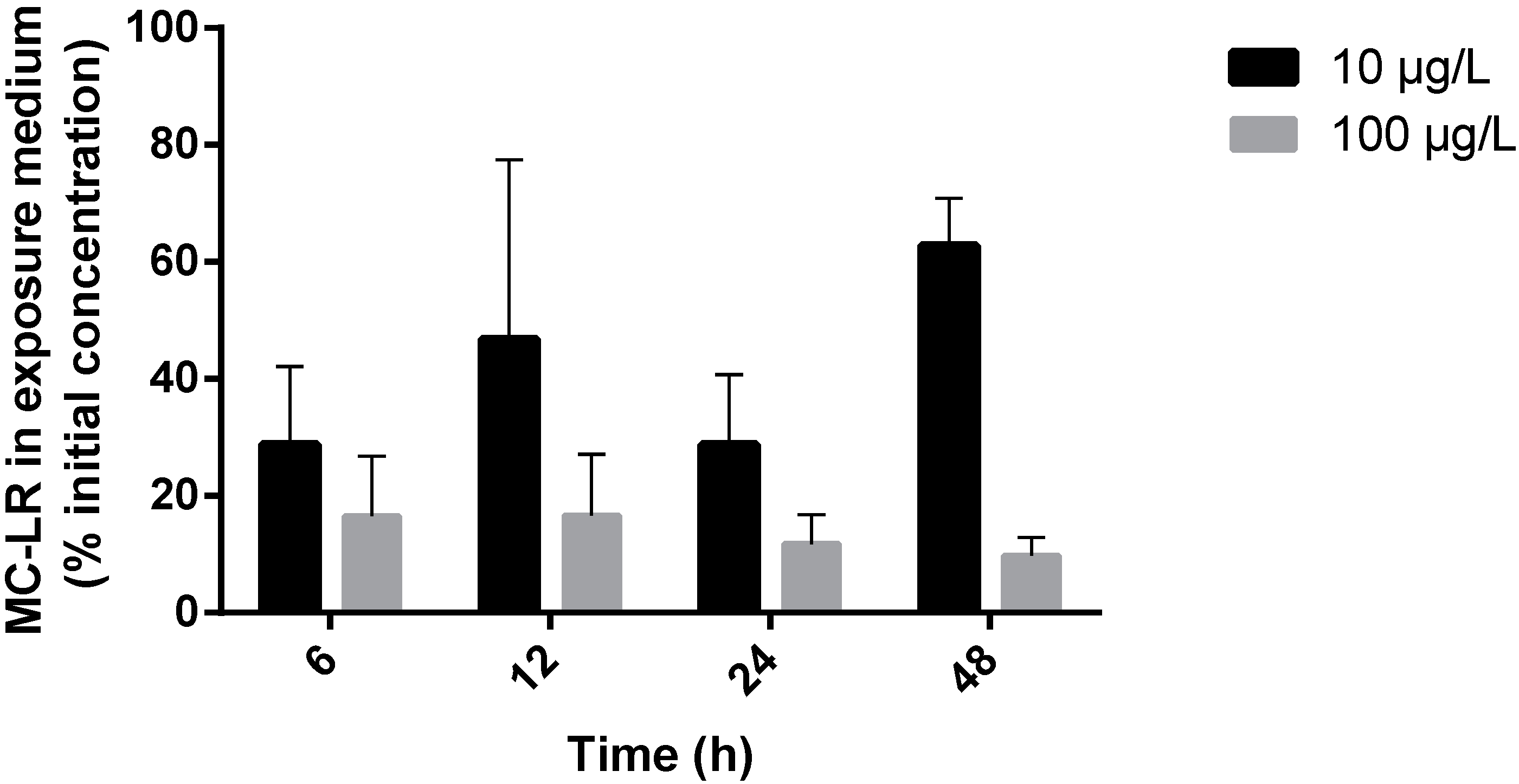
2.1. MC-LR Uptake by R. philippinarum
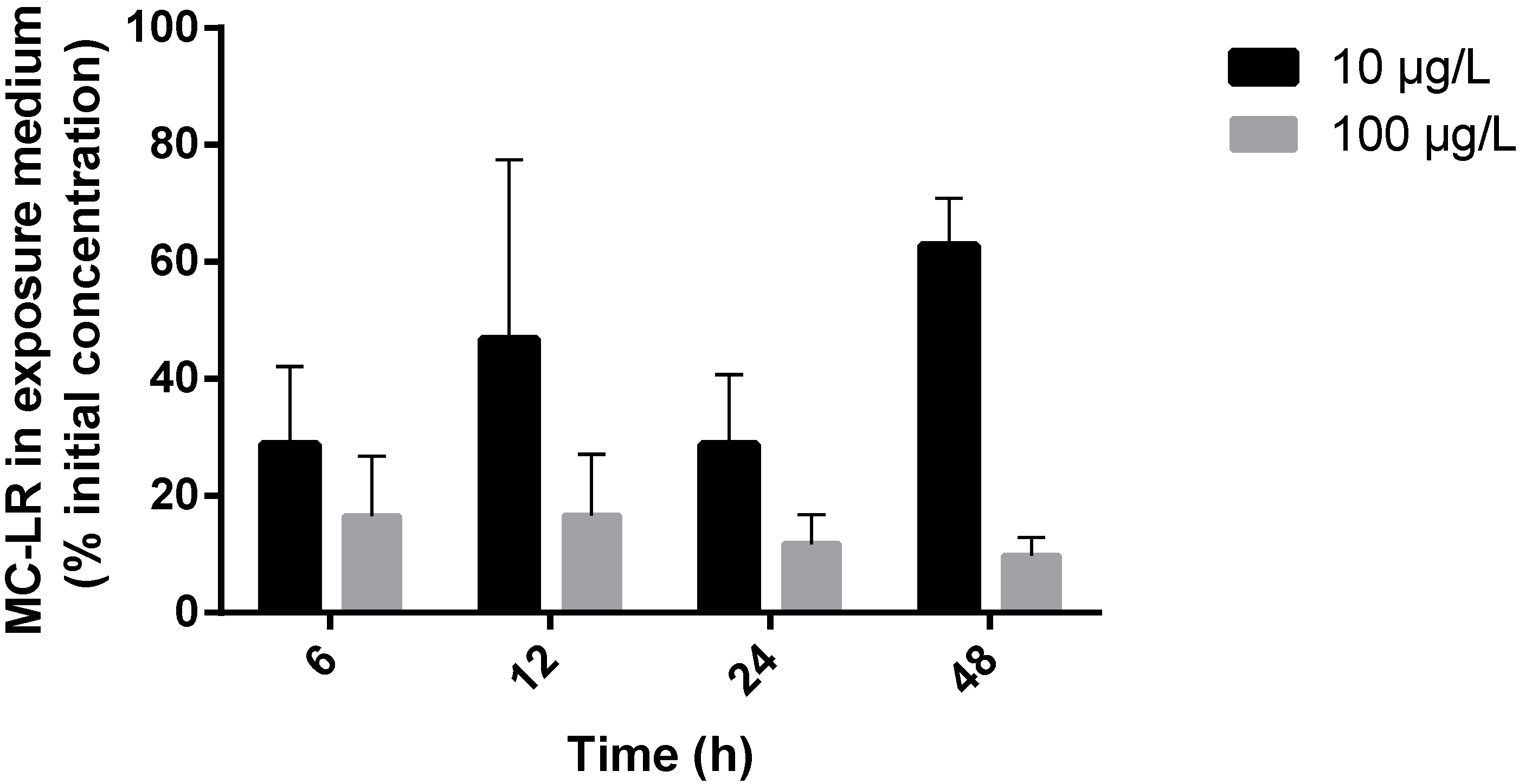
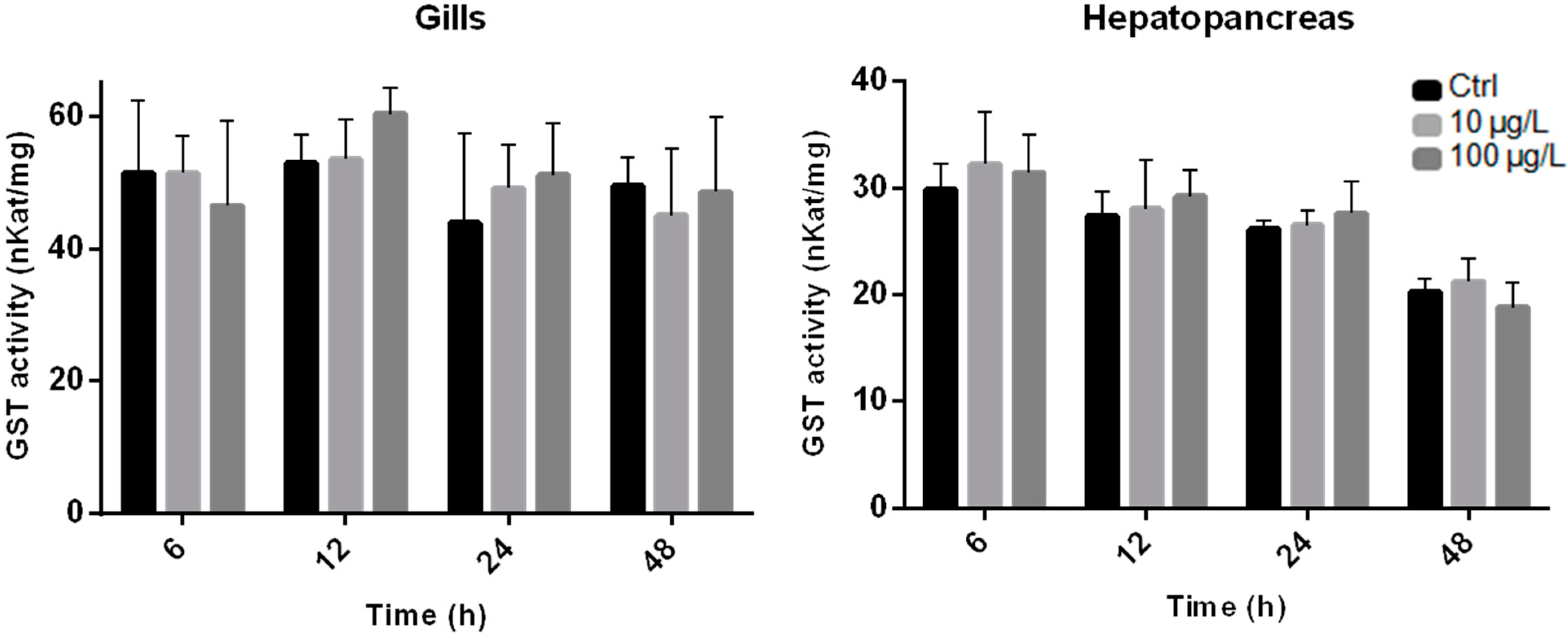
2.2. Enzyme Activity Measurements
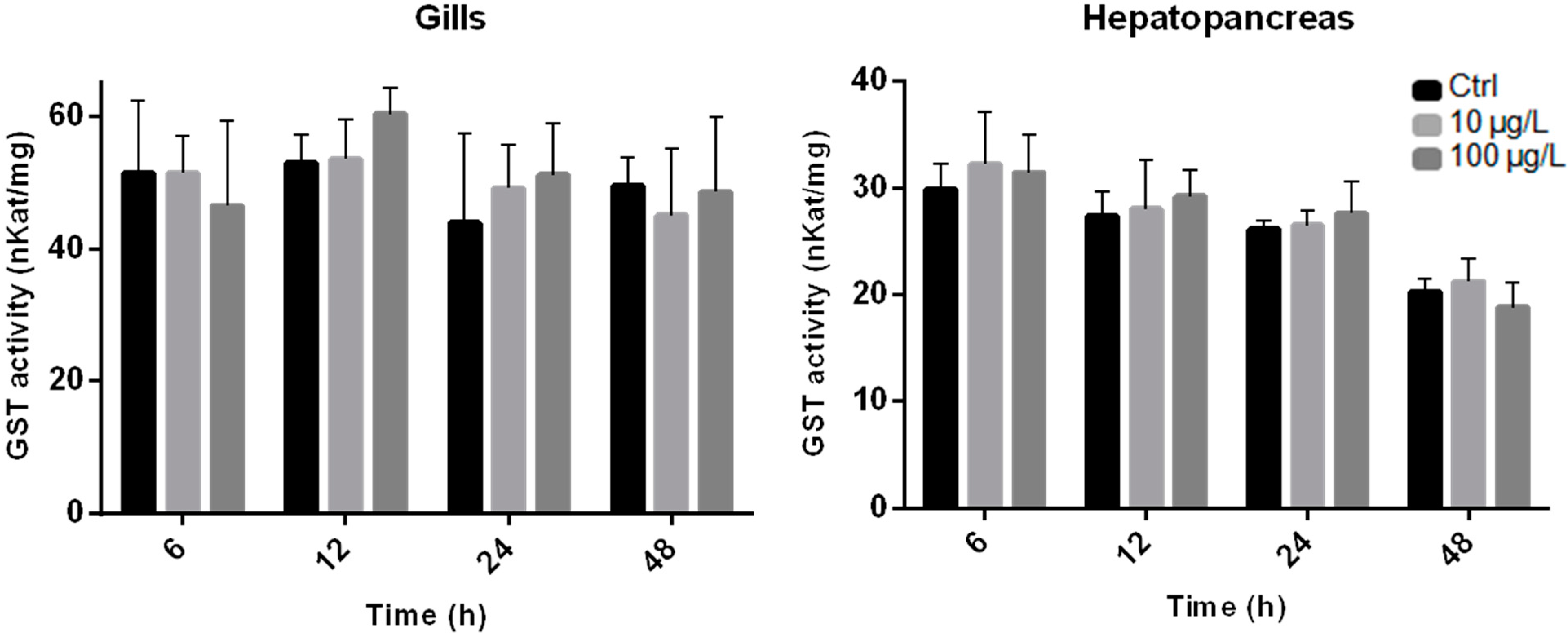
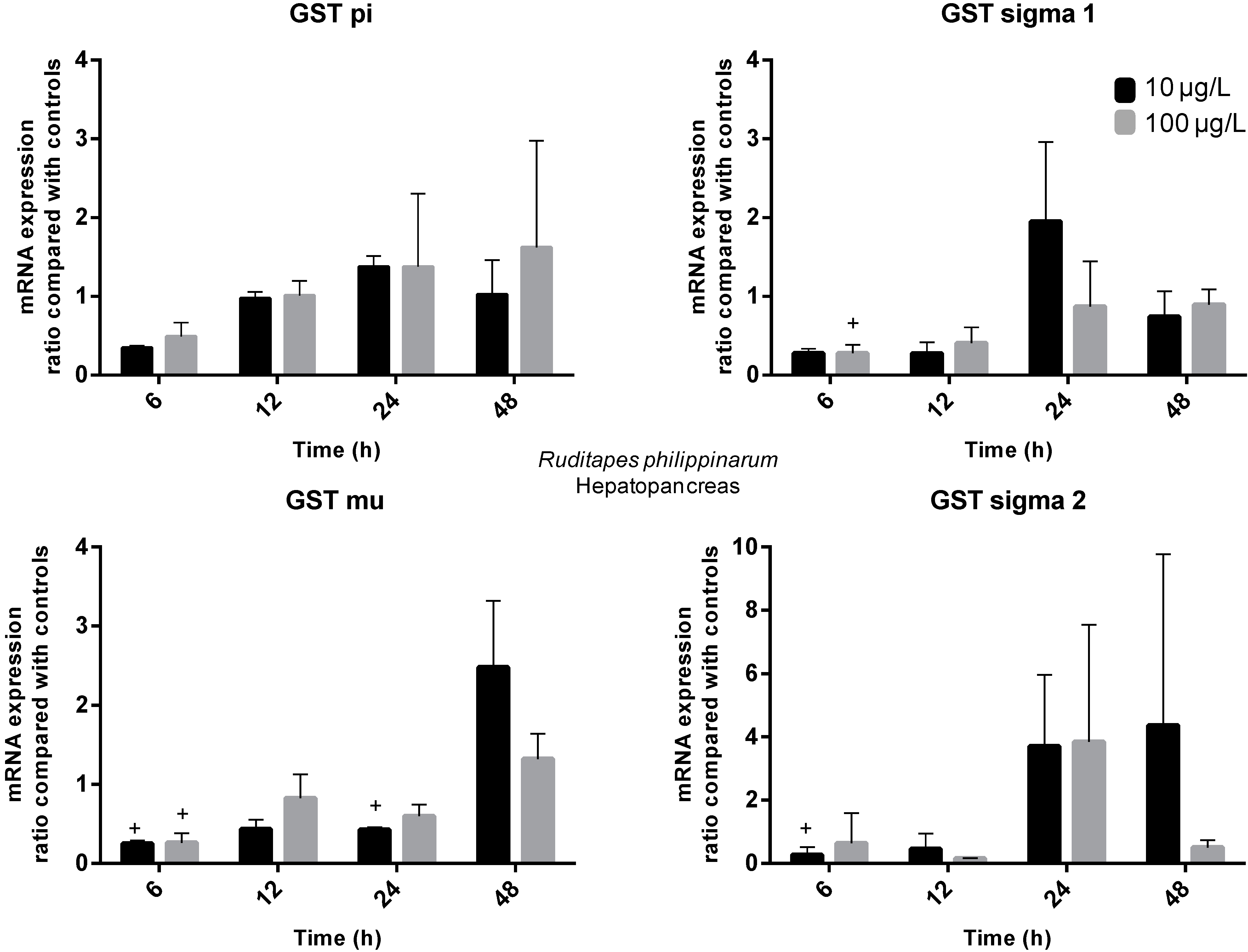
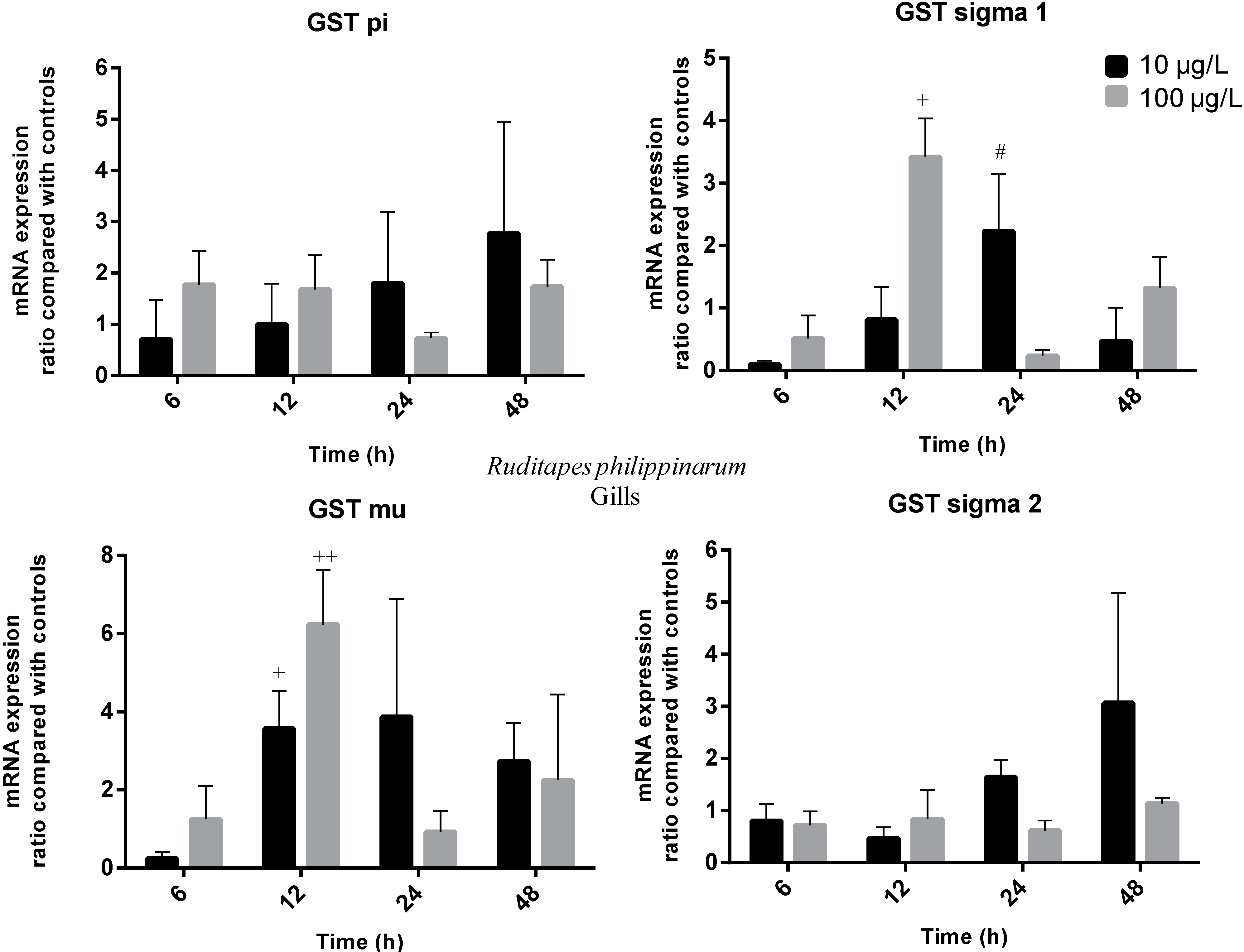
2.3. Gene Expression
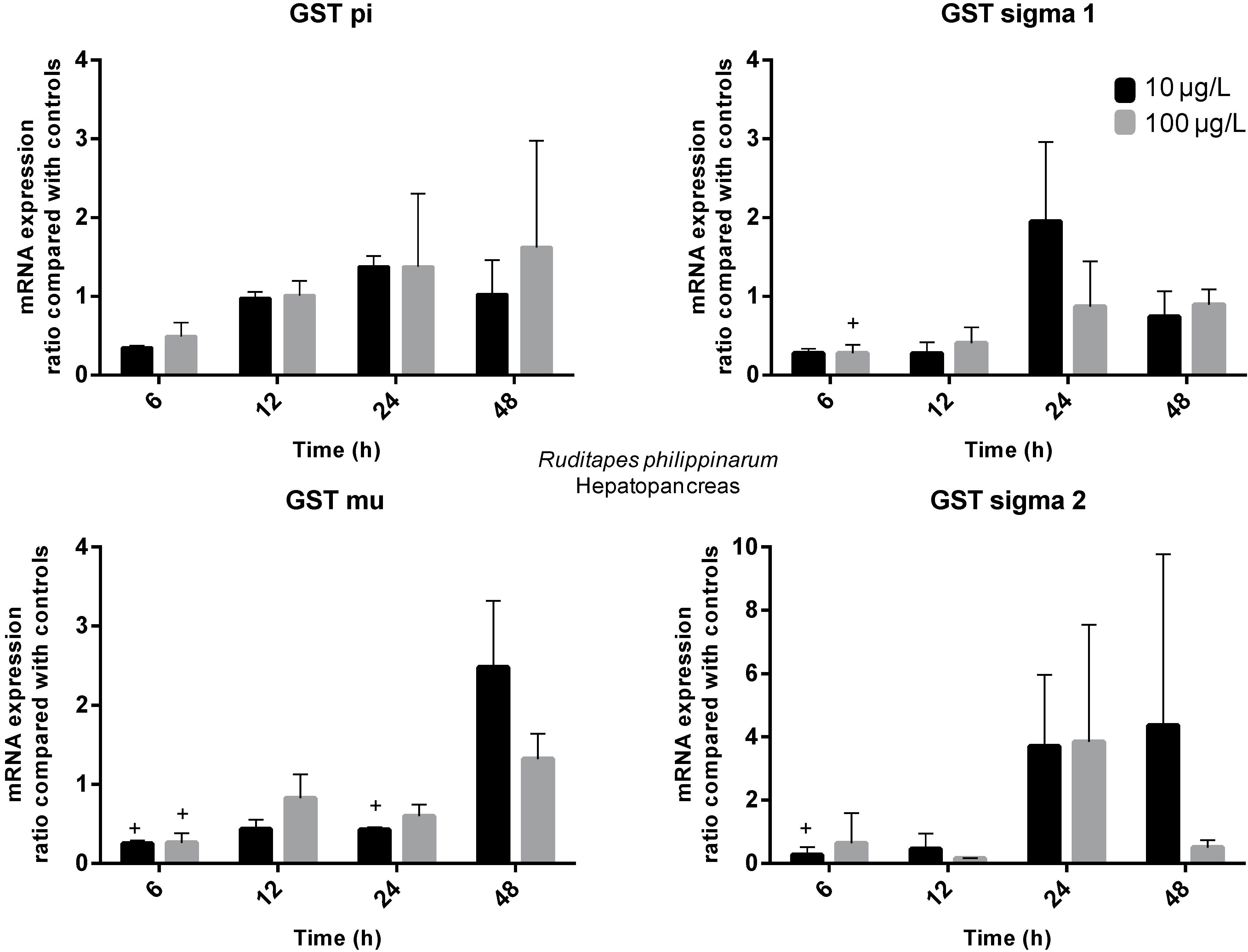
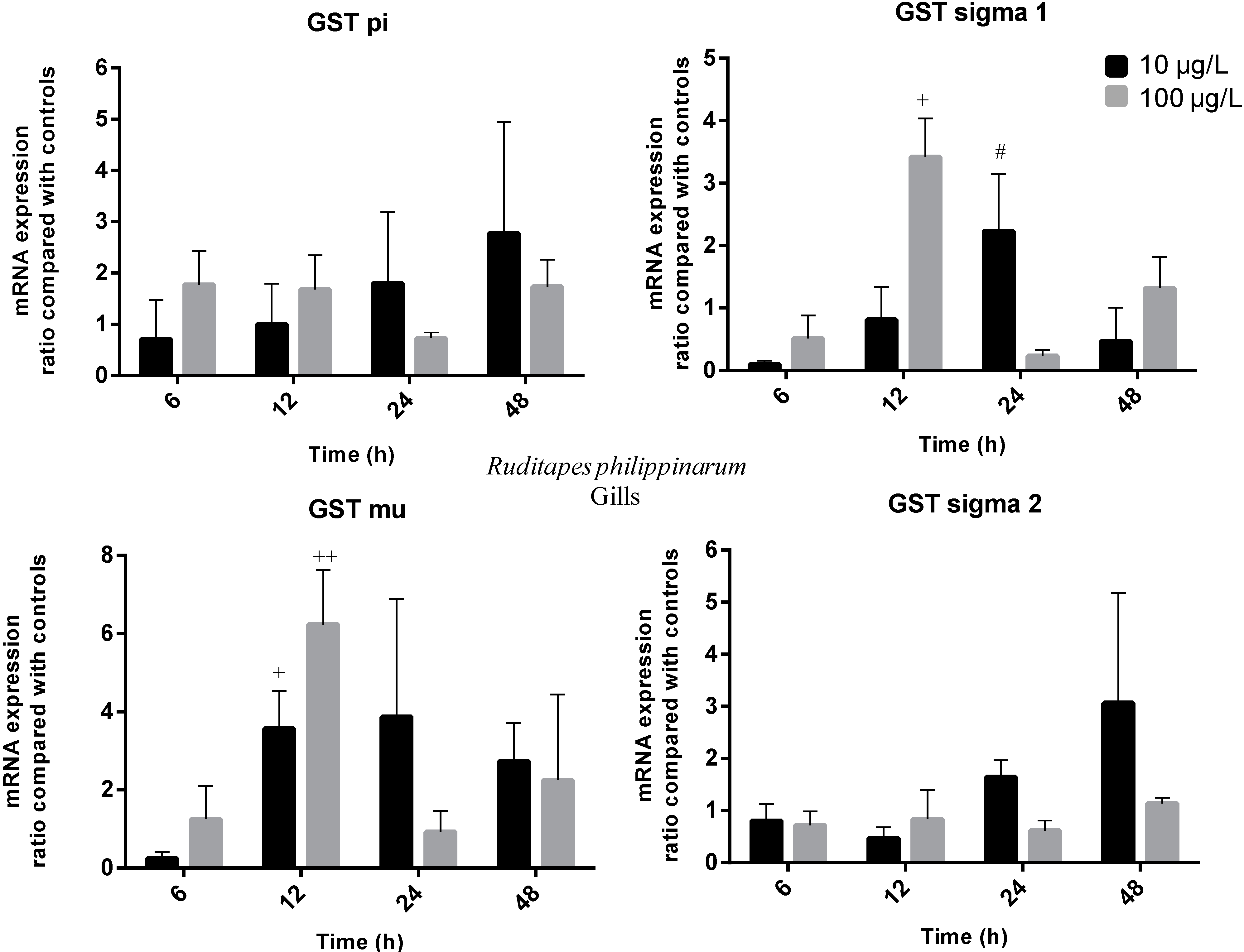
3. Experimental Section
3.1. Test Species and Cultures
3.2. MC-LR HPLC Analysis
3.2.1. Microcystin-LR Extraction for Analytical and Semi-Preparative HPLC
3.2.2. MC-LR Purification
3.2.3. MC-LR Quantification
3.3. Exposure Experiments
3.4. Toxin Analysis in Exposure Medium
3.5. Enzyme Activity Measurements
3.6. Gene Expression
3.6.1. RNA Extraction
3.6.2. cDNA Synthesis
3.6.3. Primers Design

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GST Gene | Primer Sequence (5'–3' Order) | Product Length (bp) | |
|---|---|---|---|
| Forward | Reverse | ||
| sigma1 | CAGAAGAATTTGGCAGAAGTAG | AAGACAGCAAGATCAGCGAG | 121 |
| sigma 2 | AAGGCTAAACTTACAGAGGAG | GTGTTTCTTGAGTTCAGGGT | 209 |
| mu | GACTTCCCAATGTACGAGCTT | ACACTTTCCTGAGCGAGATAC | 139 |
| pi | GCATTACCGACCCTCAAAGC | CCATTGACGGGCATTTTCTT | 101 |
| EF1-α | GCTCACAGAAGCTGTACCAGG | CTGGGCATAGAAGCTTGCAG | 136 |
3.6.4. Quantitative RT-PCR
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.M.; Pereira, M.J. Microcystin-producing blooms—A serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef]
- Vichi, S.; Lavorini, P.; Funari, E.; Scardala, S.; Testai, E. Contamination by Microcystis and microcystins of blue–green algae food supplements (BGAS) on the italian market and possible risk for the exposed population. Food Chem. Toxicol. 2012, 50, 4493–4499. [Google Scholar] [CrossRef] [PubMed]
- Ufelmann, H.; Krüger, T.; Luckas, B.; Schrenk, D. Human and rat hepatocyte toxicity and protein phosphatase 1 and 2A inhibitory activity of naturally occurring desmethyl-microcystins and nodularins. Toxicology 2012, 293, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, C.; Pflugmacher, S. Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicol. Appl. Pharmacol. 2005, 203, 201–218. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Maynes, J.T.; Luu, H.A.; Cherney, M.M.; Andersen, R.J.; Williams, D.; Holmes, C.F.; James, M.N. Crystal structures of protein phosphatase-1 bound to motuporin and dihydromicrocystin-LA: Elucidation of the mechanism of enzyme inhibition by cyanobacterial toxins. J. Mol. Biol. 2006, 356, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Vasconcelos, V. Molecular mechanisms of microcystin toxicity in animal cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.J.; Dietrich, D.R. Pathological and biochemical characterization of microcystin-induced hepatopancreas and kidney damage in Carp (Cyprinus carpio). Toxicol. Appl. Pharmacol. 2000, 164, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Dong, H.; Thompson, D.C.; Shertzer, H.G.; Nebert, D.W.; Vasiliou, V. Glutathione defense mechanism in liver injury: Insights from animal models. Food Chem. Toxicol. 2013, 60, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Y.; Song, L.; Liu, J. Responses of antioxidant systems in the hepatocytes of common carp (Cyprinus carpio L.) to the toxicity of microcystin-LR. Toxicon 2003, 42, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Pinho, G.L.L.; Moura da Rosa, C.; Maciel, F.E.; Bianchini, A.; Yunes, J.S.; Proença, L.A.O.; Monserrat, J.M. Antioxidant responses and oxidative stress after microcystin exposure in the hepatopancreas of an estuarine crab species. Ecotoxicol. Environ. Saf. 2005, 61, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Prieto, A.I.; Pichardo, S.; Jos, Á.; Moreno, I.; Cameán, A.M. Time-dependent oxidative stress responses after acute exposure to toxic cyanobacterial cells containing microcystins in tilapia fish (Oreochromis niloticus) under laboratory conditions. Aquat. Toxicol. 2007, 84, 337–345. [Google Scholar] [CrossRef]
- Žegura, B.; Zajc, I.; Lah, T.T.; Filipič, M. Patterns of microcystin-LR induced alteration of the expression of genes involved in response to DNA damage and apoptosis. Toxicon 2008, 51, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Cazenave, J.; Bistoni Mde, L.; Zwirnmann, E.; Wunderlin, D.A.; Wiegand, C. Attenuating effects of natural organic matter on microcystin toxicity in zebra fish (Danio rerio) embryos—Benefits and costs of microcystin detoxication. Environ. Toxicol. 2006, 21, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Amado, L.L.; Garcia, M.L.; Ramos, P.B.; Freitas, R.F.; Zafalon, B.; Ferreira, J.L.R.; Yunes, J.S.; Monserrat, J.M. A method to measure total antioxidant capacity against peroxyl radicals in aquatic organisms: Application to evaluate microcystins toxicity. Sci. Total Environ. 2009, 407, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Sherratt, P.J.; Hayes, J.D. Glutathione S-Transferases. In Enzyme Systems that Metabolize Drugs and Other Xenobiotics; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2002; pp. 319–352. [Google Scholar]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Pflugmacher, S.; Wiegand, C.; Oberemm, A.; Beattie, K.A.; Krause, E.; Codd, G.A.; Steinberg, C.E.W. Identification of an enzymatically formed glutathione conjugate of the cyanobacterial hepatotoxin microcystin-LR: The first step of detoxication. Biochim. Biophys. Acta. 1998, 1425, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Amorim, Á.; Vasconcelos, V.T. Dynamics of microcystins in the mussel Mytilus galloprovincialis. Toxicon 1999, 37, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, V.M. Uptake and depuration of the heptapeptide toxin microcystin-LR in Mytilus galloprovincialis. Aquat. Toxicol. 1995, 32, 227–237. [Google Scholar] [CrossRef]
- Martins, J.C.; Vasconcelos, V.M. Microcystin dynamics in aquatic organisms. J. Toxicol. Environ. Health B 2008, 12, 65–82. [Google Scholar] [CrossRef]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Amado, L.L.; Monserrat, J.M. Oxidative stress generation by microcystins in aquatic animals: Why and how. Environ. Int. 2010, 36, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.; Ramos Pinto, L.; Sampaio, M.; Costa, A.; Silva, M.; Rodrigues, A.M.; Quintino, V.; Figueira, E. Effects of depuration on the element concentration in bivalves: Comparison between sympatric Ruditapes decussatus and Ruditapes philippinarum. Estuar. Coast. Shelf Sci. 2012, 110, 43–53. [Google Scholar] [CrossRef]
- Lee, K.-W.; Raisuddin, S.; Rhee, J.-S.; Hwang, D.-S.; Yu, I.T.; Lee, Y.-M.; Park, H.G.; Lee, J.-S. Expression of glutathione S-transferase (GST) genes in the marine copepod Tigriopus japonicus exposed to trace metals. Aquat. Toxicol. 2008, 89, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Umasuthan, N.; Saranya Revathy, K.; Lee, Y.; Whang, I.; Choi, C.Y.; Lee, J. A novel molluscan sigma-like glutathione S-transferase from Manila clam, Ruditapes philippinarum: Cloning, characterization and transcriptional profiling. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.K.; Jones, G. Cyanobacterial Toxins. In Toxic Cyanobacteria in Water; Chorus, I., Bartram, J., Eds.; E&FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Figueira, E.; Freitas, R. Consumption of Ruditapes philippinarum and Ruditapes decussatus: Comparison of element accumulation and health risk. Environ. Sci. Pollut. Res. 2013, 20, 5682–5691. [Google Scholar] [CrossRef]
- Donaghy, L.; Lambert, C.; Choi, K.-S.; Soudant, P. Hemocytes of the carpet shell clam (Ruditapes decussatus) and the Manila clam (Ruditapes philippinarum): Current knowledge and future prospects. Aquaculture 2009, 297, 10–24. [Google Scholar] [CrossRef]
- Tanguy, A.; Bierne, N.; Saavedra, C.; Pina, B.; Bachère, E.; Kube, M.; Bazin, E.; Bonhomme, F.; Boudry, P.; Boulo, V.; et al. Increasing genomic information in bivalves through new EST collections in four species: Development of new genetic markers for environmental studies and genome evolution. Gene 2008, 408, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Contardo-Jara, V.; Pflugmacher, S.; Wiegand, C. Multi-xenobiotic-resistance a possible explanation for the insensitivity of bivalves towards cyanobacterial toxins. Toxicon 2008, 52, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef] [PubMed]
- Bourne, D.G.; Blakeley, R.L.; Riddles, P.; Jones, G.J. Biodegradation of the cyanobacterial toxin microcystin-LR in natural water and biologically active slow sand filters. Water Res. 2006, 40, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Mazur, H.; Plinski, M. Stability of cyanotoxins, microcystin-LR, microcystin-RR and nodularin in seawater and BG-11 medium of different salinity. Oceanologia 2001, 43, 329–339. [Google Scholar]
- Fernandes, S.; Welker, M.; Vasconcelos, V.M. Changes in the GST activity of the mussel Mytilus galloprovincialis during exposure and depuration of microcystins. J. Exp. Zool. A Ecol. Genet. Physiol. 2009, 311, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Hoarau, P.; Garello, G.; Gnassia-Barelli, M.; Roméo, M.; Girard, J.-P. Effect of three xenobiotic compounds on Glutathione S-Transferase in the clam Ruditapes decussatus. Aquat. Toxicol. 2004, 68, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, V.M.; Wiegand, C.; Pflugmacher, S. Dynamics of glutathione-S-transferases in Mytilus galloprovincialis exposed to toxic Microcystis aeruginosa cells, extracts and pure toxins. Toxicon 2007, 50, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Bebianno, M.J.; Lopes, B.; Guerra, L.; Hoarau, P.; Ferreira, A.M. Glutathione S-tranferases and cytochrome P450 activities in Mytilus galloprovincialis from the South coast of Portugal: Effect of abiotic factors. Environ. Int. 2007, 33, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Burmester, V.; Nimptsch, J.; Wiegand, C. Adaptation of freshwater mussels to cyanobacterial toxins: Response of the biotransformation and antioxidant enzymes. Ecotoxicol. Environ. Saf. 2012, 78, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Hoarau, P.; Damiens, G.; Roméo, M.; Gnassia-Barelli, M.; Bebianno, M.J. Cloning and expression of a GST-pi gene in Mytilus galloprovincialis. Attempt to use the GST-pi transcript as a biomarker of pollution. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Gowland, B.T.G.; McIntosh, A.D.; Davies, I.M.; Moffat, C.F. Glutathione S-transferase activity in mussels, Mytilus edulis, exposed to discharges from an aluminum smelter. Bull. Environ. Contam. Toxicol. 2002, 69, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Xie, P. The acute effects of microcystin LR on the transcription of nine glutathione S-transferase genes in common carp Cyprinus carpio L. Aquat. Toxicol. 2006, 80, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Xie, P.; Fu, J.; Li, G.; Xiong, Q.; Li, H. The effect of cyanobacterial crude extract on the transcription of GST mu, GST kappa and GST rho in different organs of goldfish (Carassius auratus). Aquat. Toxicol. 2008, 90, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xie, P.; Fu, J.; Hao, L.; Xiong, Q.; Li, H. Microcystin-induced variations in transcription of GSTs in an omnivorous freshwater fish, goldfish. Aquat. Toxicol. 2008, 88, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Qiu, L.; Wu, H.; Liu, X.; You, L.; Pei, D.; Chen, L.; Wang, Q.; Zhao, J. Expression profiles of seven glutathione S-transferase (GST) genes from Venerupis philippinarum exposed to heavy metals and benzo[α]pyrene. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Bathige, S.D.; Umasuthan, N.; Saranya Revathy, K.; Lee, Y.; Kim, S.; Cho, M.Y.; Park, M.A.; Whang, I.; Lee, J. A mu class glutathione S-transferase from Manila clam Ruditapes philippinarum(RpGSTmu): Cloning, mRNA expression, and conjugation assays. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2014, 162, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Li, L.; Iwamoto, N.; Nakajima-Takagi, Y.; Kaneko, H.; Nakayama, Y.; Eguchi, M.; Wada, Y.; Kumagai, Y.; Yamamoto, M. The antioxidant defense system Keap1-Nrf2 comprises a multiple sensing mechanism for responding to a wide range of chemical compounds. Mol. Cell. Biol. 2009, 29, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.K.; Wakabayashi, N.; Kensler, T.W. Chemoprevention through the Keap1-Nrf2 signaling pathway by phase 2 enzyme inducers. Mutat. Res. 2004, 555, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Toroser, D.; Yarian, C.S.; Orr, W.C.; Sohal, R.S. Mechanisms of γ-glutamylcysteine ligase regulation. Biochim. Biophys. Acta 2006, 1760, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.W.; Antoun, G.R.; Ali-Osman, F. The human glutathione S-transferase P1 protein is phosphorylated and its metabolic function enhanced by the Ser/Thr protein kinases, cAMP-dependent protein kinase and protein kinase C, in glioblastoma cells. Cancer Res. 2004, 64, 9131–9138. [Google Scholar] [CrossRef] [PubMed]
- Okamura, T.; Singh, S.; Buolamwini, J.; Haystead, T.; Friedman, H.; Bigner, D.; Ali-Osman, F. Tyrosine phosphorylation of the human glutathione S-transferase P1 by epidermal growth factor receptor. J. Biol. Chem. 2009, 284, 16979–16989. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Pan, L.; Liu, N.; Wang, L.; Miao, J. Cloning, characterization and tissue distribution of a pi-class glutathione S-transferase from clam (Venerupis philippinarum): Response to benzo[α]pyrene exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 152, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Ito, E.; Takai, A.; Kondo, F.; Masui, H.; Imanishi, S.; Harada, K. Comparison of protein phosphatase inhibitory activity and apparent toxicity of microcystins and related compounds. Toxicon 2002, 40, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [PubMed]
- Frasco, M.F.; Guilhermino, L. Effects of dimethoate and β-naphthoflavone on selected biomarkers of Poecilia reticulata. Fish. Physiol. Biochem. 2002, 26, 149–156. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Morga, B.; Arzul, I.; Faury, N.; Renault, T. Identification of genes from flat oyster Ostrea edulis as suitable housekeeping genes for quantitative real time PCR. Fish Shellfish. Immunol. 2010, 29, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.C.; Machado, J.; Martins, A.; Azevedo, J.; OlivaTeles, L.; Vasconcelos, V. Dynamics of protein phosphatase gene expression in Corbicula fluminea exposed to microcystin-LR and to toxic Microcystis aeruginosa cells. Int. J. Mol. Sci. 2011, 12, 9172–9188. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, B.; Carneiro, M.; Machado, J.; Azevedo, J.; Vasconcelos, V.; Martins, J.C. Transcriptional Responses of Glutathione Transferase Genes in Ruditapes philippinarum Exposed to Microcystin-LR. Int. J. Mol. Sci. 2015, 16, 8397-8414. https://doi.org/10.3390/ijms16048397
Reis B, Carneiro M, Machado J, Azevedo J, Vasconcelos V, Martins JC. Transcriptional Responses of Glutathione Transferase Genes in Ruditapes philippinarum Exposed to Microcystin-LR. International Journal of Molecular Sciences. 2015; 16(4):8397-8414. https://doi.org/10.3390/ijms16048397
Chicago/Turabian StyleReis, Bruno, Mariana Carneiro, João Machado, Joana Azevedo, Vitor Vasconcelos, and José Carlos Martins. 2015. "Transcriptional Responses of Glutathione Transferase Genes in Ruditapes philippinarum Exposed to Microcystin-LR" International Journal of Molecular Sciences 16, no. 4: 8397-8414. https://doi.org/10.3390/ijms16048397
APA StyleReis, B., Carneiro, M., Machado, J., Azevedo, J., Vasconcelos, V., & Martins, J. C. (2015). Transcriptional Responses of Glutathione Transferase Genes in Ruditapes philippinarum Exposed to Microcystin-LR. International Journal of Molecular Sciences, 16(4), 8397-8414. https://doi.org/10.3390/ijms16048397