
Aminoacyl-tRNA Synthetase Complexes in Evolution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
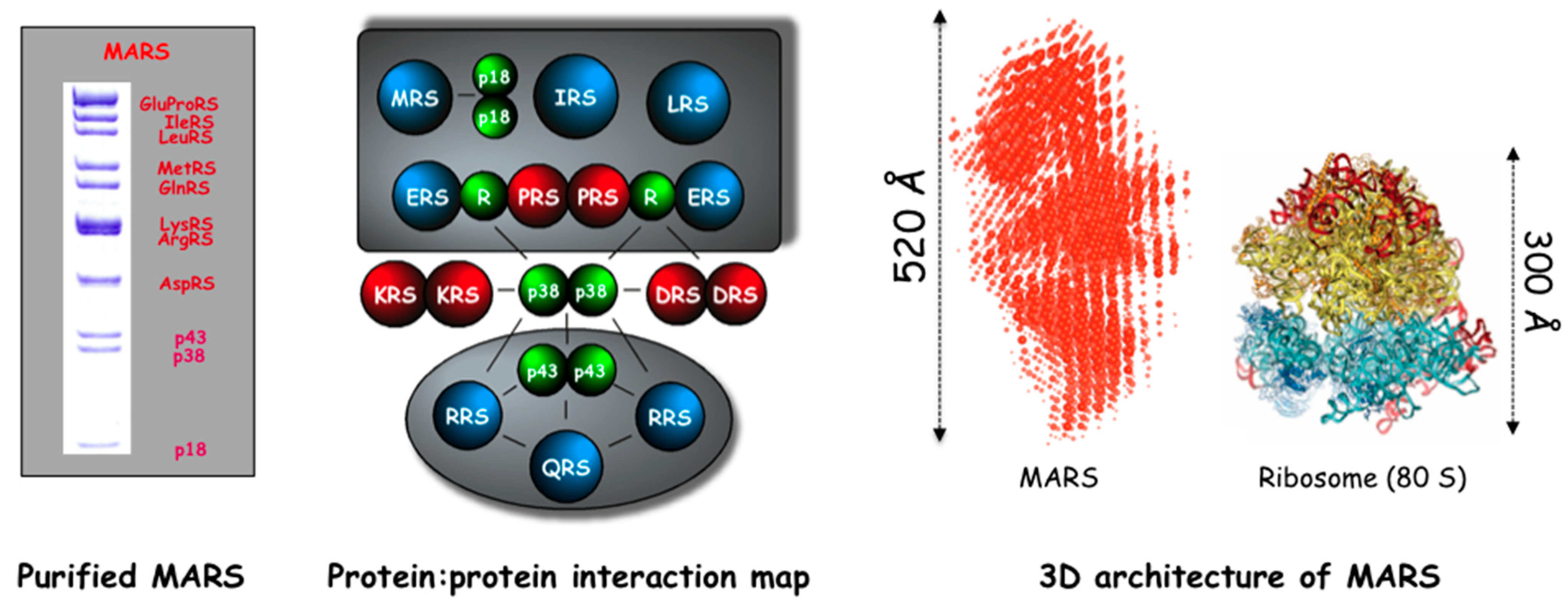
2. Multi-Aminoacyl-tRNA Synthetase Complexes (MARS)
2.1. Complexes in Archaea
2.2. Complexes in Unicellular Eukaryotes
2.2.1. The MARS in Saccharomyces cerevisiae

2.2.2. The MARS in Toxoplasma gondii
2.2.3. The MARS in Trypanosoma brucei
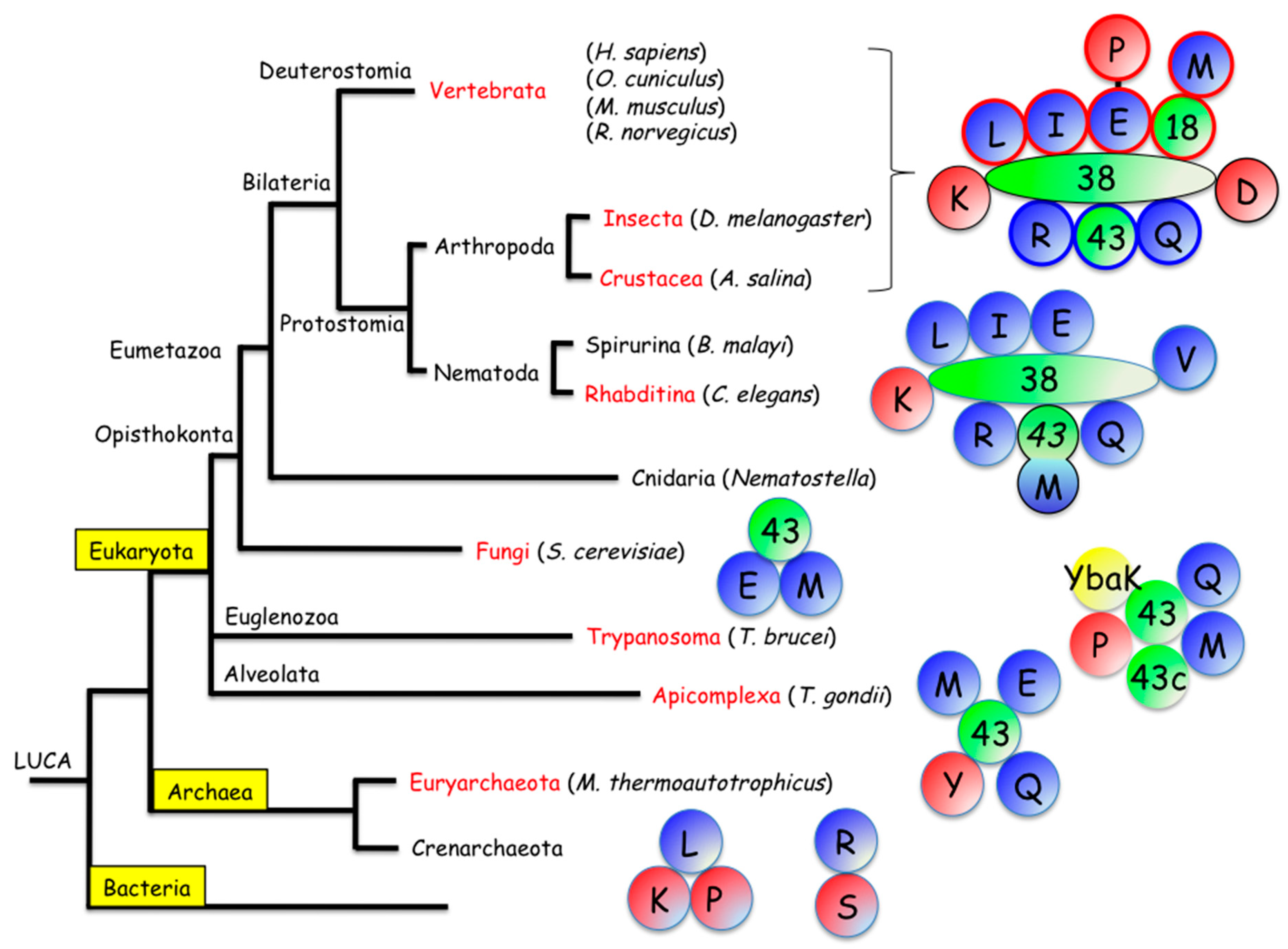
2.3. The MARS in Eumetazoa
2.3.1. The MARS in Deuterostomia
Composition of the MARS
Assembly of the MARS

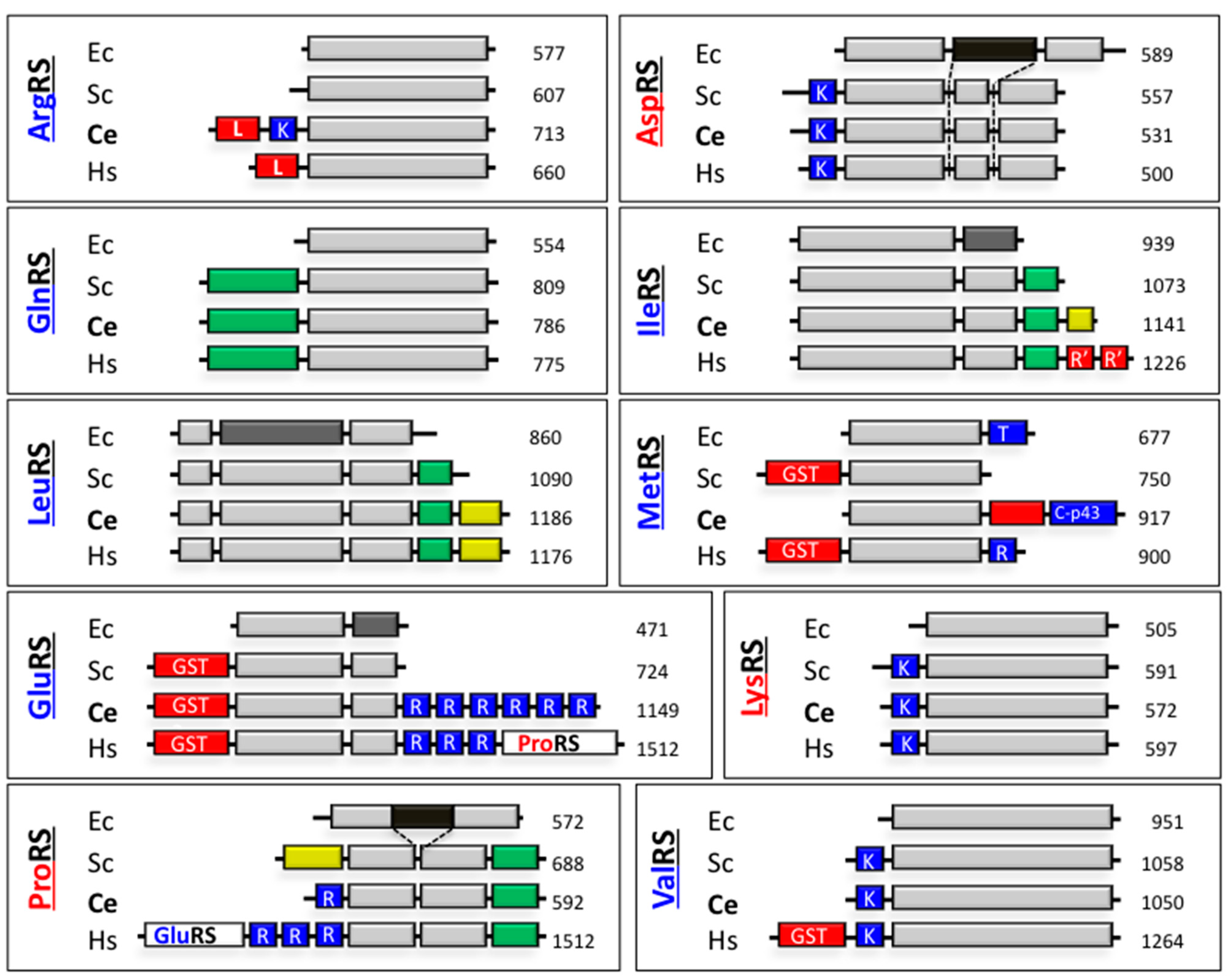
2.3.2. The MARS in Protostomia
Arthropoda
Nematoda
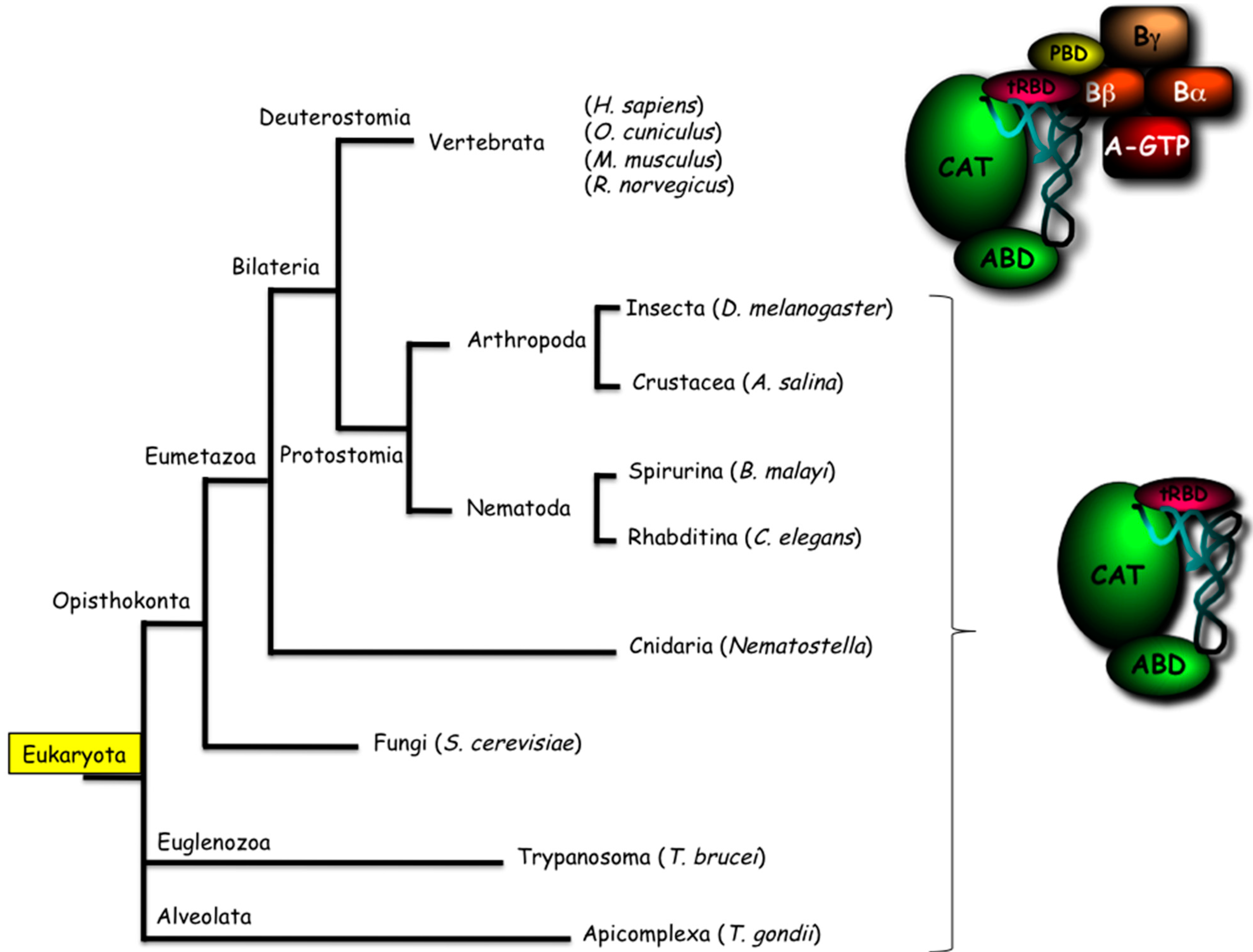
3. Integration of the MARS in Cellular Homeostasis
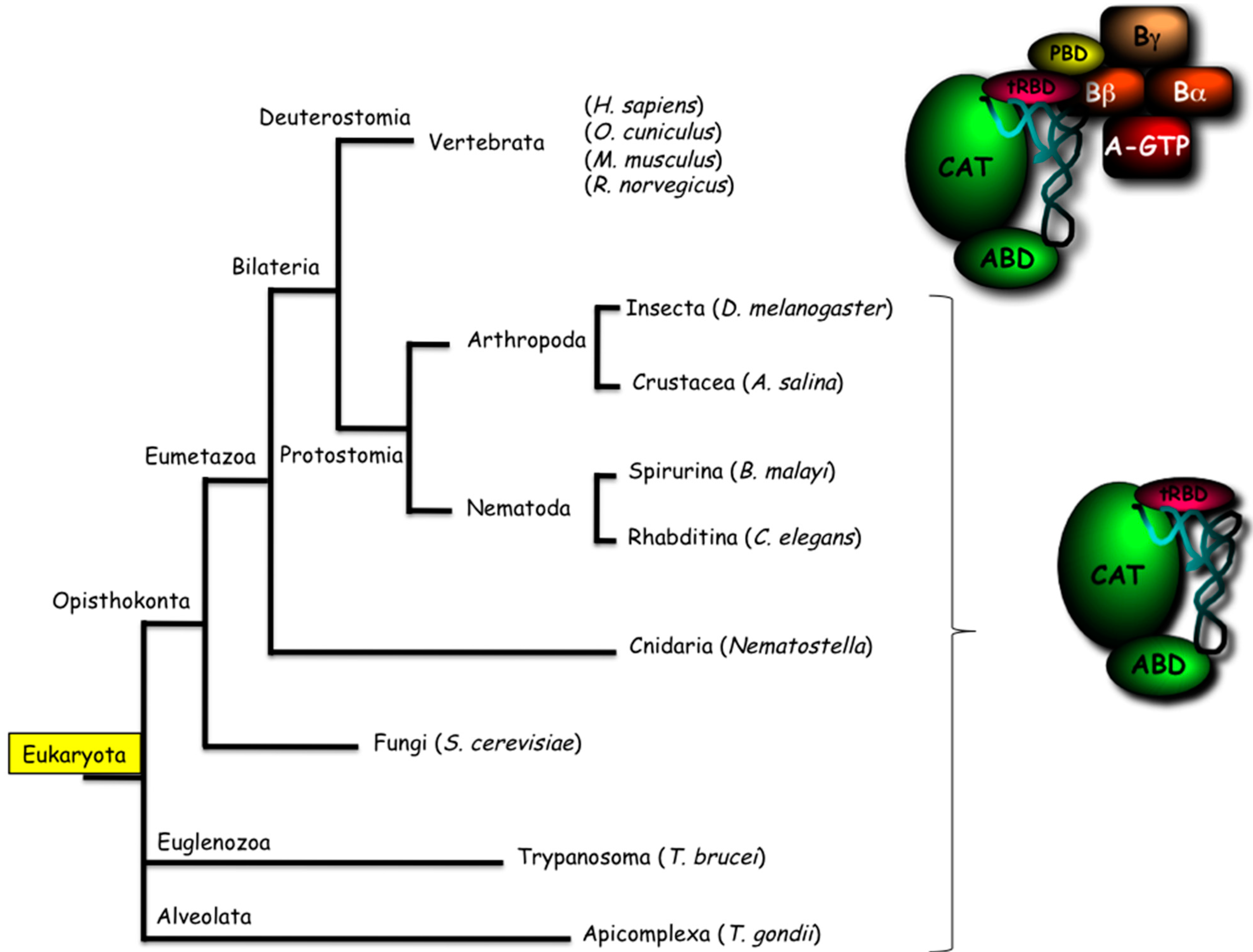
3.1. Integration of the MARS in Translation Apparatus
3.2. Role of the MARS in Regulation of Other Cellular Functions
4. Concluding Remarks
4.1. Origin of the MARS of the Eukaryotic-Type
4.2. Understanding the Balance between Translational and Non-Translational Functions
4.3. Perspectives
Acknowledgments
Conflicts of Interest
References
- Ibba, M.; Söll, D. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 2000, 69, 617–650. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, P.; Söll, D. Aminoacyl-tRNA synthetases: General features and recognition of transfer RNAs. Annu. Rev. Biochem. 1979, 48, 601–648. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Schimmel, P.; Yang, X.L. Functional expansion of human tRNA synthetases achieved by structural inventions. FEBS Lett. 2010, 584, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Yang, X.L.; Schimmel, P. New functions of aminoacyl-tRNA synthetases beyond translation. Nat. Rev. Mol. Cell Biol. 2011, 11, 668–674. [Google Scholar] [CrossRef]
- Hausmann, C.D.; Ibba, M. Aminoacyl-tRNA synthetase complexes: Molecular multitasking revealed. FEMS Microbiol. Rev. 2008, 32, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Schimmel, P. Essential nontranslational functions of tRNA synthetases. Nat. Chem. Biol. 2013, 9, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Motzik, A.; Nechushtan, H.; Foo, S.Y.; Razin, E. Non-canonical roles of lysyl-tRNA synthetase in health and disease. Trends Mol. Med. 2013, 19, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Fox, P.L. Aminoacyl-tRNA synthetases in medicine and disease. EMBO Mol. Med. 2013, 5, 332–343. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Musier Forsyth, K. Cys-tRNAPro editing by Haemophilus influenzae YbaK via a novel synthetase-YbaK-tRNA ternary complex. J. Biol. Chem. 2005, 280, 34465–34472. [Google Scholar] [CrossRef] [PubMed]
- Lipman, R.S.A.; Chen, J.; Evilia, C.; Vitseva, O.; Hou, Y.M. Association of an aminoacyl-tRNA synthetase with a putative metabolic protein in archaea. Biochemistry 2003, 42, 7487–7496. [Google Scholar] [CrossRef] [PubMed]
- Oza, J.P.; Sowers, K.R.; Perona, J.J. Linking energy production and protein synthesis in hydrogenotrophic methanogens. Biochemistry 2012, 51, 2378–2389. [Google Scholar] [CrossRef] [PubMed]
- Godinic, V.; Mocibob, M.; Rocak, S.; Ibba, M.; Weygand-Durasevic, I. Peroxin Pex21p interacts with the C-terminal noncatalytic domain of yeast seryl-tRNA synthetase and forms a specific ternary complex with tRNASer. FEBS J. 2007, 274, 2788–2799. [Google Scholar] [CrossRef] [PubMed]
- Laporte, D.; Huot, J.L.; Bader, G.; Enkler, L.; Senger, B.; Becker, H.D. Exploring the evolutionary diversity and assembly modes of multi-aminoacyl-tRNA synthetase complexes: Lessons from unicellular organisms. FEBS Lett. 2014, 588, 4268–4278. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.L. An aminoacyl-tRNA synthetase complex in Escherichia coli. J. Bacteriol. 1987, 169, 2718–2723. [Google Scholar] [PubMed]
- Harris, C.L.; Kolanko, C.J. Aminoacyl-tRNA synthetase complex in Saccharomyces cerevisiae. Biochem. J. 1995, 309, 321–324. [Google Scholar] [PubMed]
- Bandyopadhyay, A.K.; Deutscher, M.P. Complex of aminoacyl-tRNA synthetases. J. Mol. Biol. 1971, 60, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kaminska, M.; Havrylenko, S.; Decottignies, P.; Le Marechal, P.; Negrutskii, B.; Mirande, M. Dynamic organization of aminoacyl-tRNA synthetase complexes in the cytoplasm of human cells. J. Biol. Chem. 2009, 284, 13746–13754. [Google Scholar] [CrossRef] [PubMed]
- Barbarese, E.; Koppel, D.E.; Deutscher, M.P.; Smith, C.L.; Ainger, K.; Morgan, F.; Carson, J.H. Protein translation components are colocalized in granules in oligodendrocytes. J. Cell Sci. 1995, 108, 2781–2790. [Google Scholar] [PubMed]
- Negrutskii, B.S.; Stapulionis, R.; Deutscher, M.P. Supramolecular organization of the mammalian translation system. Proc. Natl. Acad. Sci. USA 1994, 91, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Praetorius-Ibba, M.; Hausmann, C.D.; Paras, M.; Rogers, T.E.; Ibba, M. Functional association between three archaeal aminoacyl-tRNA synthetases. J. Biol. Chem. 2007, 282, 3680–3687. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, C.D.; Ibba, M. Structural and functional mapping of the archaeal multi-aminoacyl-tRNA synthetase complex. FEBS Lett. 2008, 582, 2178–2182. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, C.D.; Praetorius-Ibba, M.; Ibba, M. An aminoacyl-tRNA synthetase:Elongation factor complex for substrate channeling in archaeal translation. Nucleic Acids Res. 2007, 35, 6094–6102. [Google Scholar] [CrossRef] [PubMed]
- Godinic-Mikulcic, V.; Jaric, J.; Hausmann, C.D.; Ibba, M.; Weygand-Durasevic, I. An archaeal tRNA-synthetase complex that enhances aminoacylation under extreme conditions. J. Biol. Chem. 2011, 286, 3396–3404. [Google Scholar] [CrossRef] [PubMed]
- Godinic-Mikulcic, V.; Jaric, J.; Greber, B.J.; Franke, V.; Hodnik, V.; Anderluh, G.; Ban, N.; Weygand-Durasevic, I. Archaeal aminoacyl-tRNA synthetases interact with the ribosome to recycle tRNAs. Nucleic Acids Res. 2014, 42, 5191–5201. [Google Scholar] [CrossRef] [PubMed]
- Raina, M.; Elgamal, S.; Santangelo, T.J.; Ibba, M. Association of a multi-synthetase complex with translating ribosomes in the archaeon Thermococcus kodakarensis. FEBS Lett. 2012, 586, 2232–2238. [Google Scholar] [CrossRef] [PubMed]
- Simos, G.; Segref, A.; Fasiolo, F.; Hellmuth, K.; Shevchenko, A.; Mann, M.; Hurt, E.C. The yeast protein Arc1p binds to tRNA and functions as a cofactor for the methionyl- and glutamyl-tRNA synthetases. EMBO J. 1996, 15, 5437–5448. [Google Scholar] [PubMed]
- Simos, G.; Sauer, A.; Fasiolo, F.; Hurt, E.C. A conserved domain within Arc1p delivers tRNA to aminoacyl-tRNA synthetases. Mol. Cell 1998, 1, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Golinelli-Cohen, M.P.; Mirande, M. Arc1p is required for cytoplasmic confinement of synthetases and tRNA. Mol. Cell. Biochem. 2007, 300, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Graindorge, J.S.; Senger, B.; Tritch, D.; Simos, G.; Fasiolo, F. Role of Arc1p in the modulation of yeast glutamyl-tRNA synthetase activity. Biochemistry 2005, 44, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Golinelli-Cohen, M.P.; Zakrzewska, A.; Mirande, M. Complementation of yeast Arc1p by the p43 component of the human multisynthetase complex does not require its association with yeast MetRS and GluRS. J. Mol. Biol. 2004, 340, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Galani, K.; Grosshans, H.; Deinert, K.; Hurt, E.C.; Simos, G. The intracellular location of two aminoacyl-tRNA synthetases depends on complex formation with Arc1p. EMBO J. 2001, 20, 6889–6898. [Google Scholar] [CrossRef] [PubMed]
- Frechin, M.; Enkler, L.; Tetaud, E.; Laporte, D.; Senger, B.; Blancard, C.; Hammann, P.; Bader, G.; Clauder-Münster, S.; Steinmetz, L.M.; et al. Expression of nuclear and mitochondrial genes encoding ATP synthase is synchronized by disassembly of a multisynthetase complex. Mol. Cell 2014, 56, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Frechin, M.; Senger, B.; Braye, M.; Kern, D.; Martin, R.P.; Becker, H.D. Yeast mitochondrial Gln-tRNAGln is generated by a GatFAB-mediated transamidation pathway involving Arc1p-controlled subcellular sorting of cytosolic GluRS. Genes Dev. 2009, 23, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Simader, H.; Hothorn, M.; Kohler, C.; Basquin, J.; Simos, G.; Suck, D. Structural basis of yeast aminoacyl-tRNA synthetase complex formation revealed by crystal structures of two binary sub-complexes. Nucleic Acids Res. 2006, 34, 3968–3979. [Google Scholar] [CrossRef] [PubMed]
- Karanasios, E.; Simader, H.; Panayotou, G.; Suck, D.; Simos, G. Molecular determinants of the yeast Arc1p-aminoacyl-tRNA synthetase complex assembly. J. Mol. Biol. 2007, 374, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Hoja, U.; Stolz, J.; Sauer, G.; Schweizer, E. Identification of the tRNA-binding protein Arc1p as a novel target of in vivo biotinylation in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 42445–42452. [Google Scholar] [CrossRef] [PubMed]
- Koehler, C.; Round, A.; Simader, H.; Suck, D.; Svergun, D. Quaternary structure of the yeast Arc1p-aminoacyl-tRNA synthetase complex in solution and its compaction upon binding of tRNAs. Nucleic Acids Res. 2013, 41, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Van Rooyen, J.M.; Murat, J.B.; Hammoudi, P.M.; Kieffer-Jaquinod, S.; Coute, Y.; Sharma, A.; Pelloux, H.; Belrhali, H.; Hakimi, M.A. Assembly of the novel five-component apicomplexan multi-aminoacyl-tRNA synthetase complex is driven by the hybrid scaffold protein Tg-p43. PLoS One 2014, 9, e89487. [Google Scholar] [CrossRef] [PubMed]
- Cestari, I.; Kalidas, S.; Monnerat, S.; Anupama, A.; Phillips, M.A.; Stuart, K. A multiple aminoacyl-tRNA synthetase complex that enhances tRNA-aminoacylation in African trypanosomes. Mol. Cell. Biol. 2013, 33, 4872–4888. [Google Scholar] [CrossRef] [PubMed]
- Vennegoor, C.; Bloemendal, H. Occurrence and particle character of aminoacyl-tRNA synthetases in the post-microsomal fraction from rat liver. Eur. J. Biochem. 1972, 26, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Som, K.; Hardesty, B. Isolation and partial characterization of an aminoacyl-tRNA synthetase complex from rabbit reticulocytes. Arch. Biochem. Biophys. 1975, 166, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Denney, R.M. Detection and partial purification of rapidly sedimenting forms of aminoacyl-transfer ribonucleic acid synthetases from human placenta. Arch. Biochem. Biophys. 1977, 183, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Ussery, M.A.; Tanaka, W.K.; Hardesty, B. Subcellular distribution of aminoacyl-tRNA synthetases in various eukaryotic cells. Eur. J. Biochem. 1977, 72, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Kellermann, O.; Brevet, A.; Tonetti, H.; Waller, J.P. Macromolecular complexes of aminoacyl-tRNA synthetases from eukaryotes. Extensive purification and characterization of the high-molecular-weight complex(es) of seven aminoacyl-tRNA synthetases from sheep liver. Eur. J. Biochem. 1979, 99, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.L.; Yang, D.C.H. Stoichiometry and composition of an aminoacyl-tRNA synthetase complex from rat liver. Proc. Natl. Acad. Sci. USA 1981, 78, 4059–4062. [Google Scholar] [CrossRef] [PubMed]
- Kellermann, O.; Tonetti, H.; Brevet, A.; Mirande, M.; Pailliez, J.P.; Waller, J.P. Macromolecular complexes from sheep and rabbit containing seven aminoacyl-tRNA synthetases I. Species specificity of the polypeptide composition. J. Biol. Chem. 1982, 257, 11041–11048. [Google Scholar] [PubMed]
- Godar, D.E.; Godar, D.E.; Garcia, V.; Jacobo, A.; Aebi, U.; Yang, D.C.H. Structural organization of the multienzyme complex of mammalian aminoacyl-tRNA synthetases. Biochemistry 1988, 27, 6921–6928. [Google Scholar] [CrossRef] [PubMed]
- Mirande, M.; Le Corre, D.; Waller, J.P. A complex from cultured Chinese hamster ovary cells containing nine aminoacyl-tRNA synthetases. Thermolabile leucyl-tRNA synthetase from the tsH1 mutant cell line is an integral component of this complex. Eur. J. Biochem. 1985, 147, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Norcum, M.T. Isolation and electron microscopic characterization of the high molecular mass aminoacyl-tRNA synthetase complex from murine erythroleukemia cells. J. Biol. Chem. 1989, 264, 15043–15051. [Google Scholar] [PubMed]
- Quevillon, S.; Mirande, M. The p18 component of the multisynthetase complex shares a protein motif with the b and g subunits of eukaryotic elongation factor 1. FEBS Lett. 1996, 395, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, S.; Robinson, J.C.; Berthonneau, E.; Siatecka, M.; Mirande, M. Macromolecular assemblage of aminoacyl-tRNA synthetases: Identification of protein-protein interactions and characterization of a core protein. J. Mol. Biol. 1999, 285, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, S.; Agou, F.; Robinson, J.C.; Mirande, M. The p43 component of the mammalian multi-synthetase complex is likely to be the precursor of the endothelial monocyte-activating polypeptide II cytokine. J. Biol. Chem. 1997, 272, 32573–32579. [Google Scholar] [CrossRef] [PubMed]
- Cérini, C.; Kerjan, P.; Astier, M.; Gratecos, D.; Mirande, M.; Semeriva, M. A component of the multisynthetase complex is a multifunctional aminoacyl-tRNA synthetase. EMBO J. 1991, 10, 4267–4277. [Google Scholar] [PubMed]
- Kerjan, P.; Triconnet, M.; Waller, J.P. Mammalian prolyl-tRNA synthetase corresponds to the 150 kDa subunit of the high-M(r) aminoacyl-tRNA synthetase complex. Biochimie 1992, 74, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.S.; Sullivan, J.C.; Jia, J.; Francis, J.; Finnerty, J.R.; Fox, P.L. Evolution of function of a fused metazoan tRNA synthetase. Mol. Biol. Evol. 2011, 28, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Berthonneau, E.; Mirande, M. A gene fusion event in the evolution of aminoacyl-tRNA synthetases. FEBS Lett. 2000, 470, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.S.; Fox, P.L. Origin and evolution of glutamyl-prolyl tRNA synthetase WHEP domains reveal evolutionary relationships within Holozoa. PLoS One 2014, 9, e98493. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.; Renault, L.; Perez, J.; Mirande, M. Small-angle X-ray solution scattering study of the multi-aminoacyl-tRNA synthetase complex reveals an elongated and multi-armed particle. J. Biol. Chem. 2013, 288, 23979–23989. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.G.; Wei, H.; Ling, C.; Xu, M.G.; Wang, E.D. Two forms of human cytoplasmic arginyl-tRNA synthetase produced from two translation initiations by a single mRNA. Biochemistry 2006, 45, 1338–1344. [Google Scholar] [CrossRef] [PubMed]
- Lazard, M.; Mirande, M. Cloning and analysis of a cDNA encoding mammalian arginyl-tRNA synthetase, a component of the multisynthetase complex with a hydrophobic N-terminal extension. Gene 1993, 132, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, S.V.; Deutscher, M.P. An important role for the multienzyme aminoacyl-tRNA synthetase complex in mammalian translation and cell growth. Mol. Cell 2008, 29, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Sivaram, P.; Deutscher, M.P. Existence of two forms of rat liver arginyl-tRNA synthetase suggests channeling of aminoacyl-tRNA for protein synthesis. Proc. Natl. Acad. Sci. USA 1990, 87, 3665–3669. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Potdar, A.A.; Arif, A.; Ray, P.S.; Mukhopadhyay, R.; Willard, B.; Xu, Y.; Yan, J.; Saidel, G.M.; Fox, P.L. Coding region polyadenylation generates a truncated tRNA synthetase that counters translation repression. Cell 2012, 149, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Tolkunova, E.; Park, H.; Xia, J.; King, M.P.; Davidson, E. The human lysyl-tRNA synthetase gene encodes both the cytoplasmic and mitochondrial enzymes by means of an unusual alternative splicing of the primary transcript. J. Biol. Chem. 2000, 275, 35063–35069. [Google Scholar] [CrossRef] [PubMed]
- Kaminska, M.; Shalak, V.; Francin, M.; Mirande, M. Viral hijacking of mitochondrial lysyl-tRNA synthetase. J. Virol. 2007, 81, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Wolfson, A.; Knight, R. Occurrence of the aminoacyl-tRNA synthetases in high-molecular weight complexes correlates with the size of substrate amino acids. FEBS Lett. 2005, 579, 3467–3472. [Google Scholar] [CrossRef] [PubMed]
- Eswarappa, S.M.; Fox, P.L. Citric acid cycle and the origin of MARS. Trends Biochem. Sci. 2013, 38, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.C.; Kim, S.; Lee, B.J. Solution structure and p43 binding of the p38 leucine zipper motif: Coiled-coil interactions mediate the association between p38 and p43. FEBS Lett. 2003, 542, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Kim, D.G.; Lee, A.E.; Kim, H.R.; Lee, J.Y.; Kwon, N.H.; Shin, Y.K.; Hwang, S.K.; Chang, S.H.; Cho, M.H.; et al. Cancer-associated splicing variant of tumor suppressor AIMP2/p38: Pathological implication in tumorigenesis. PLoS Genet. 2011, 7, e1001351. [Google Scholar] [CrossRef] [PubMed]
- Kaminska, M.; Havrylenko, S.; Decottignies, P.; Gillet, S.; Marechal, P.L.; Negrutskii, B.; Mirande, M. Dissection of the structural organization of the aminoacyl-tRNA synthetase complex. J. Biol. Chem. 2009, 284, 6053–6060. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kang, Y.S.; Lee, J.W.; Kim, H.J.; Ahn, Y.H.; Park, H.; Ko, Y.G.; Kim, S. p38 is essential for the assembly and stability of macromolecular tRNA synthetase complex: Implications for its physiological significance. Proc. Natl. Acad. Sci. USA 2002, 99, 7912–7916. [Google Scholar] [CrossRef] [PubMed]
- Corti, O.; Hampe, C.; Koutnikova, H.; Darios, F.; Jacquier, S.; Prigent, A.; Robinson, J.C.; Pradier, L.; Ruberg, M.; Mirande, M.; et al. The p38 subunit of the aminoacyl-tRNA synthetase complex is a Parkin substrate: Linking protein biosynthesis and neurodegeneration. Hum. Mol. Genet. 2003, 12, 1427–1437. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.S.; von Coelln, R.; Sriram, S.R.; Kim, S.W.; Chung, K.K.K.; Pletnikova, O.; Troncoso, J.; Johnson, B.; Saffary, R.; Goh, E.L.; et al. Accumulation of the authentic parkin substrate aminoacyl-tRNA synthetase cofactor, p38/JTV-1, leads to catecholaminergic cell death. J. Neurosci. 2005, 25, 7968–7978. [Google Scholar] [CrossRef] [PubMed]
- Sriram, S.R.; Li, X.J.; Ko, H.S.; Chung, K.K.K.; Wong, E.; Lim, K.L.; Dawson, V.L.; Dawson, T.M. Familial-associated mutations differentially disrupt the solubility, localization, binding and ubiquitination properties of parkin. Hum. Mol. Genet. 2005, 14, 2571–2586. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.J.; Park, M.C.; Choi, S.J.; Oh, Y.S.; Choi, E.C.; Cho, H.J.; Kim, M.H.; Kim, S.H.; Kim, D.W.; Kim, S.; et al. Determination of three-dimensional structure and residues of the novel tumor suppressor AIMP3/p18 required for the interaction with ATM. J. Biol. Chem. 2008, 283, 14032–14040. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.; Kwon, N.H.; Lee, J.Y.; Park, M.C.; Kang, E.; Kim, H.H.; Kang, T.J.; Kim, S. AIMP3/p18 controls translational initiation by mediating the delivery of charged initiator tRNA to initiation complex. J. Mol. Biol. 2012, 423, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Shalak, V.; Kaminska, M.; Mitnacht-Kraus, R.; Vandenabeele, P.; Clauss , M.; Mirande, M. The EMAPII cytokine is released from the mammalian multisynthetase complex after cleavage of its p43/proEMAPII component. J. Biol. Chem. 2001, 276, 23769–23776. [Google Scholar]
- Kao, J.; Ryan, J.; Brett, G.; Chen, J.; Shen, H.; Fan, Y.G.; Godman, G.; Familletti, P.C.; Wang, F.; Pan, Y.C.; et al. Endothelial monocyte-activating polypeptide II. A novel tumor-derived polypeptide that activates host-response mechanisms. J. Biol. Chem. 1992, 267, 20239–20247. [Google Scholar] [PubMed]
- Swairjo, M.A.; Morales, A.J.; Wang, C.C.; Ortiz, A.R.; Schimmel, P. Crystal structure of Trbp111: A structure-specific tRNA-binding protein. EMBO J. 2000, 19, 6287–6298. [Google Scholar] [CrossRef] [PubMed]
- Renault, L.; Kerjan, P.; Pasqualato, S.; Menetrey, J.; Robinson, J.C.; Kawaguchi, S.; Vassylyev, D.G.; Yokoyama, S.; Mirande, M.; Cherfils, J. Structure of the EMAPII domain of human aminoacyl-tRNA synthetase complex reveals evolutionary dimer mimicry. EMBO J. 2001, 20, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Shin, J.; Li, R.B.; Cheong, C.; Kim, K.; Kim, S. A novel anti-tumor cytokine contains an RNA binding motif present in aminoacyl-tRNA synthetases. J. Biol. Chem. 2000, 275, 27062–27068. [Google Scholar] [PubMed]
- Park, S.G.; Jung, K.H.; Lee, J.S.; Jo, Y.J.; Motegi, H.; Kim, S.; Shiba, K. Precursor of pro-apoptotic cytokine modulates aminoacylation activity of tRNA synthetase. J. Biol. Chem. 1999, 274, 16673–16676. [Google Scholar] [CrossRef] [PubMed]
- Guigou, L.; Shalak, V.; Mirande, M. The tRNA-interacting factor p43 associates with mammalian arginyl-tRNA synthetase but does not modify its tRNA aminoacylation properties. Biochemistry 2004, 43, 4592–4600. [Google Scholar] [CrossRef] [PubMed]
- Shalak, V.; Kaminska, M.; Mirande, M. Translation initiation from two in-frame AUGs generates mitochondrial and cytoplasmic forms of the p43 component of the multisynthetase complex. Biochemistry 2009, 48, 9959–9968. [Google Scholar] [CrossRef] [PubMed]
- Norcum, M.T.; Warrington, J.A. Structural analysis of the multienzyme aminoacyl-tRNA synthetase complex: A three-domain model based on reversible chemical crosslinking. Protein Sci. 1998, 7, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Rho, S.B.; Kim, M.J.; Lee, J.S.; Seol, W.G.; Motegi, H.; Kim, S.; Shiba, K. Genetic dissection of protein–protein interactions in multi-tRNA synthetase complex. Proc. Natl. Acad. Sci. USA 1999, 96, 4488–4493. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Yao, Y.N.; Zheng, Y.G.; Wei, H.; Wang, L.; Wu, X.F.; Wang, E.D. The C-terminal appended domain of human cytosolic leucyl-tRNA synthetase is indispensable in its interaction with arginyl-tRNA synthetase in the multi-tRNA synthetase complex. J. Biol. Chem. 2005, 280, 34755–34763. [Google Scholar] [CrossRef] [PubMed]
- Rho, S.B.; Lee, K.H.; Kim, J.W.; Shiba, K.; Jo, Y.J.; Kim, S. Interaction between human tRNA synthetases involves repeated sequence elements. Proc. Natl. Acad. Sci. USA 1996, 93, 10128–10133. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.C.; Kerjan, P.; Mirande, M. Macromolecular assemblage of aminoacyl-tRNA synthetases: Quantitative analysis of protein-protein interactions and mechanism of complex assembly. J. Mol. Biol. 2000, 304, 983–994. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Zu, L.D.; Yao, P.; Chen, X.; Wang, E.D. Two non-redundant fragments in the N-terminal peptide of human cytosolic methionyl-tRNA synthetase were indispensable for the multi-synthetase complex incorporation and enzyme activity. Biochim. Biophys. Acta 2009, 1794, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Mirande, M.; Kellermann, O.; Waller, J.P. Macromolecular complexes from sheep and rabbit containing seven aminoacyl-tRNA synthetases. II. Structural characterization of the polypeptide components and immunological identification of the methionyl-tRNA synthetase subunit. J. Biol. Chem. 1982, 257, 11049–11055. [Google Scholar] [PubMed]
- Kaminska, M.; Shalak, V.; Mirande, M. The appended C-domain of human methionyl-tRNA synthetase has a tRNA-sequestering function. Biochemistry 2001, 40, 14309–14316. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Park, S.G.; Kim, J.E.; Seol, W.; Ko, Y.G.; Kim, S. Catalytic peptide of human glutaminyl-tRNA synthetase is essential for its assembly to the aminoacyl-tRNA synthetase complex. J. Biol. Chem. 2000, 275, 21768–21772. [Google Scholar] [CrossRef] [PubMed]
- Kobbi, L.; Octobre, G.; Dias, J.; Comisso, M.; Mirande, M. Association of mitochondrial lysyl-tRNA synthetase with HIV-1 GagPol involves catalytic domain of the synthetase and transframe and integrase domains of Pol. J. Mol. Biol. 2011, 410, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Ofir-Birin, Y.; Fang, P.; Bennett, S.P.; Zhang, H.M.; Wang, J.; Rachmin, I.; Shapiro, R.; Song, J.; Dagan, A.; Pozo, J.; et al. Structural switch of lysyl-tRNA synthetase between translation and transcription. Mol. Cell 2013, 49, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Kim, Y.; Jin, K.S.; Kim, H.S.; Kim, J.H.; Wang, D.; Park, M.; Jo, C.H.; Kwon, N.H.; Kim, D.; et al. Structure of the ArgRS-GlnRS-AIMP1 complex and its implications for mammalian translation. Proc. Natl. Acad. Sci. USA 2014, 111, 15084–15089. [Google Scholar] [CrossRef] [PubMed]
- Lazard, M.; Kerjan, P.; Agou, F.; Mirande, M. The tRNA-dependent activation of arginine by arginyl-tRNA synthetase requires inter-domain communication. J. Mol. Biol. 2000, 302, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Norcum, M.T.; Boisset, N. Three-dimensional architecture of the eukaryotic multisynthetase complex determined from negatively stained and cryoelectron micrographs. FEBS Lett. 2002, 512, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Kerjan, P.; Cérini, C.; Semeriva, M.; Mirande, M. The multienzyme complex containing nine aminoacyl-tRNA synthetases is ubiquitous from Drosophila to mammals. Biochim. Biophys. Acta 1994, 1199, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Brandsma, M.; Kerjan, P.; Dijk, J.; Janssen, G.M.; Moller, W. Valyl-tRNA synthetase from Artemia. Purification and association with elongation factor 1. Eur. J. Biochem. 1995, 233, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Havrylenko, S.; Legouis, R.; Negrutskii, B.; Mirande, M. Caenorhabditis elegans evolves a new architecture for the multi-aminoacyl-tRNA synthetase complex. J. Biol. Chem. 2011, 286, 28476–28487. [Google Scholar] [CrossRef] [PubMed]
- Havrylenko, S.; Legouis, R.; Negrutskii, B.; Mirande, M. Methionyl-tRNA synthetase from Caenorhabditis elegans. A specific multi-domain organization for convergent functional evolution. Protein Sci. 2010, 19, 2475–2484. [Google Scholar] [CrossRef] [PubMed]
- Motorin, Y.A.; Wolfson, A.D.; Orlovsky, A.F.; Gladilin, K.L. Mammalian valyl-tRNA synthetase forms a complex with the first elongation factor. FEBS Lett. 1988, 238, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Bec, G.; Kerjan, P.; Zha, X.D.; Waller, J.P. Valyl-tRNA synthetase from rabbit liver. I. Purification as a heterotypic complex in association with elongation factor 1. J. Biol. Chem. 1989, 264, 21131–21137. [Google Scholar] [PubMed]
- Mansilla, F.; Friis, I.; Jadidi, M.; Nielsen, K.M.; Clark, B.F.C.; Knudsen, C.R. Mapping the human translation elongation factor eEF1H complex using the yeast two-hybrid system. Biochem. J. 2002, 365, 669–676. [Google Scholar] [PubMed]
- Bec, G.; Kerjan, P.; Waller, J.P. Reconstitution in vitro of the valyl-tRNA synthetase-elongation factor (EF) 1βγδ complex: Essential roles of the NH2-terminal extension of valyl-tRNA synthetase and of the EF-1δ subunit in complex formation. J. Biol. Chem. 1994, 269, 2086–2092. [Google Scholar] [PubMed]
- Reed, V.S.; Wastney, M.E.; Yang, D.C.H. Mechanisms of the transfer of aminoacyl-tRNA from aminoacyl-tRNA synthetase to the elongation factor 1α. J. Biol. Chem. 1994, 269, 32932–32936. [Google Scholar] [PubMed]
- David, A.; Netzer, N.; Strader, M.B.; Das, S.R.; Chen, C.Y.; Gibbs, J.; Pierre, P.; Bennink, J.R.; Yewdell, J.W. RNA binding targets aminoacyl-tRNA synthetases to translating ribosomes. J. Biol. Chem. 2011, 286, 20688–20700. [Google Scholar] [CrossRef] [PubMed]
- Stapulionis, R.; Kolli, S.; Deutscher, M.P. Efficient mammalian protein synthesis requires an intact F-Actin system. J. Biol. Chem. 1997, 272, 24980–24986. [Google Scholar] [CrossRef] [PubMed]
- Stapulionis, R.; Deutscher, M.P. A channeled tRNA cycle during mammalian protein synthesis. Proc. Natl. Acad. Sci. USA 1995, 92, 7158–7161. [Google Scholar] [CrossRef] [PubMed]
- Mirande, M. Processivity of translation in the eukaryote cell: Role of aminoacyl-tRNA synthetases. FEBS Lett. 2010, 584, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.S.; Arif, A.; Fox, P.L. Macromolecular complexes as depots for releasable regulatory proteins. Trends Biochem. Sci. 2007, 32, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Nechushtan, H.; Figov, N.; Razin, E. The function of lysyl-tRNA synthetase and Ap4A as signaling regulators of MITF activity in FceRI-activated mast cells. Immunity 2004, 20, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Yannay-Cohen, N.; Carmi-Levy, I.; Kay, G.; Yang, C.M.; Han, J.M.; Kemeny, D.M.; Kim, S.; Nechushtan, H.; Razin, E. LysRS serves as a key signaling molecule in the immune response by regulating gene expression. Mol. Cell 2009, 34, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, L.L.; Justesen, J.; Wolfson, A.D.; Frolova, L.Y. Diadenosine oligophosphates (ApnA), a novel class of signalling molecules? FEBS Lett. 1998, 427, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Sampath, P.; Mazumder, B.; Seshadri, V.; Gerber, C.A.; Chavatte, L.; Kinter, M.; Ting, S.M.; Dignam, J.D.; Kim, S.; Driscoll, D.M.; et al. Noncanonical function of glutamyl-prolyl-tRNA synthetase: Gene-specific silencing of translation. Cell 2004, 119, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Arif, A.; Jia, J.; Mukhopadhyay, R.; Willard, B.; Kinter, M.; Fox, P.L. Two-site phosphorylation of EPRS coordinates multimodal regulation of noncanonical translational control activity. Mol. Cell 2009, 35, 164–180. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.G.; Lee, J.Y.; Kwon, N.H.; Fang, P.; Zhang, Q.; Wang, J.; Young, N.L.; Guo, M.; Cho, H.Y.; Mushtaq, A.U.; et al. Chemical inhibition of prometastatic lysyl-tRNA synthetase-laminin receptor interaction. Nat. Chem. Biol. 2014, 10, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Park, S.G.; Kang, Y.S.; Ahn, Y.H.; Lee, S.H.; Kim, K.R.; Kim, K.W.; Koh, G.Y.; Ko, Y.G.; Kim, S. Dose-dependent biphasic activity of tRNA synthetase-associating factor, p43, in angiogenesis. J. Biol. Chem. 2002, 277, 45243–45248. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Jeong, S.J.; Park, M.C.; Kim, G.; Kwon, N.H.; Kim, H.K.; Ha, S.H.; Ryu, S.H.; Kim, S. Leucyl-tRNA synthetase is an intracellular leucine sensor for the mTORC1-signaling pathway. Cell 2012, 149, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Tsun, Z.Y.; Bar-Peled, L.; Chantranupong, L.; Zoncu, R.; Wang, T.; Kim, C.; Spooner, E.; Sabatini, D.M. The folliculin tumor suppressor is a GAP for the RagC/D GTPases that signal amino acid levels to mTORC1. Mol. Cell 2013, 52, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.G.; Kim, E.K.; Kim, T.; Park, H.; Park, H.S.; Choi, E.J.; Kim, S. Glutamine-dependent antiapoptotic interaction of human glutaminyl-tRNA synthetase with apoptosis signal-regulating kinase 1. J. Biol. Chem. 2001, 276, 6030–6036. [Google Scholar] [CrossRef] [PubMed]
- Meerschaert, K.; Remue, E.; de Ganck, A.; Staes, A.; Boucherie, C.; Gevaert, K.; Vandekerckhove, J.; Kleiman, L.; Gettemans, J. The tandem PDZ protein Syntenin interacts with the aminoacyl tRNA synthetase complex in a lysyl-tRNA synthetase-dependent manner. J. Proteome Res. 2008, 7, 4962–4973. [Google Scholar] [CrossRef] [PubMed]
- Javanbakht, H.; Halwani, R.; Cen, S.; Saadatmand, J.; Musier-Forsyth, K.; Gottlinger, H.; Kleiman, L. The interaction between HIV-1 Gag and human lysyl-tRNA synthetase during viral assembly. J. Biol. Chem. 2003, 278, 27644–27651. [Google Scholar] [CrossRef] [PubMed]
- Kunst, C.B.; Mezey, E.; Brownstein, M.J.; Patterson, D. Mutations in SOD1 associated with amyotrophic lateral sclerosis cause novel protein interactions. Nat. Genet. 1997, 15, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.S.; Gardiner, E.; Xu, Z.; Lau, C.F.; Wang, F.; Zhou, J.J.; Mendlein, J.D.; Nangle, L.A.; Chiang, K.P.; Yang, X.L.; et al. Human tRNA synthetase catalytic nulls with diverse functions. Science 2014, 345, 328–332. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havrylenko, S.; Mirande, M. Aminoacyl-tRNA Synthetase Complexes in Evolution. Int. J. Mol. Sci. 2015, 16, 6571-6594. https://doi.org/10.3390/ijms16036571
Havrylenko S, Mirande M. Aminoacyl-tRNA Synthetase Complexes in Evolution. International Journal of Molecular Sciences. 2015; 16(3):6571-6594. https://doi.org/10.3390/ijms16036571
Chicago/Turabian StyleHavrylenko, Svitlana, and Marc Mirande. 2015. "Aminoacyl-tRNA Synthetase Complexes in Evolution" International Journal of Molecular Sciences 16, no. 3: 6571-6594. https://doi.org/10.3390/ijms16036571
APA StyleHavrylenko, S., & Mirande, M. (2015). Aminoacyl-tRNA Synthetase Complexes in Evolution. International Journal of Molecular Sciences, 16(3), 6571-6594. https://doi.org/10.3390/ijms16036571