
MRS of Brain Metabolite Levels Demonstrates the Ability of Scavenging of Excess Brain Glutamate to Protect against Nerve Agent Induced Seizures

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
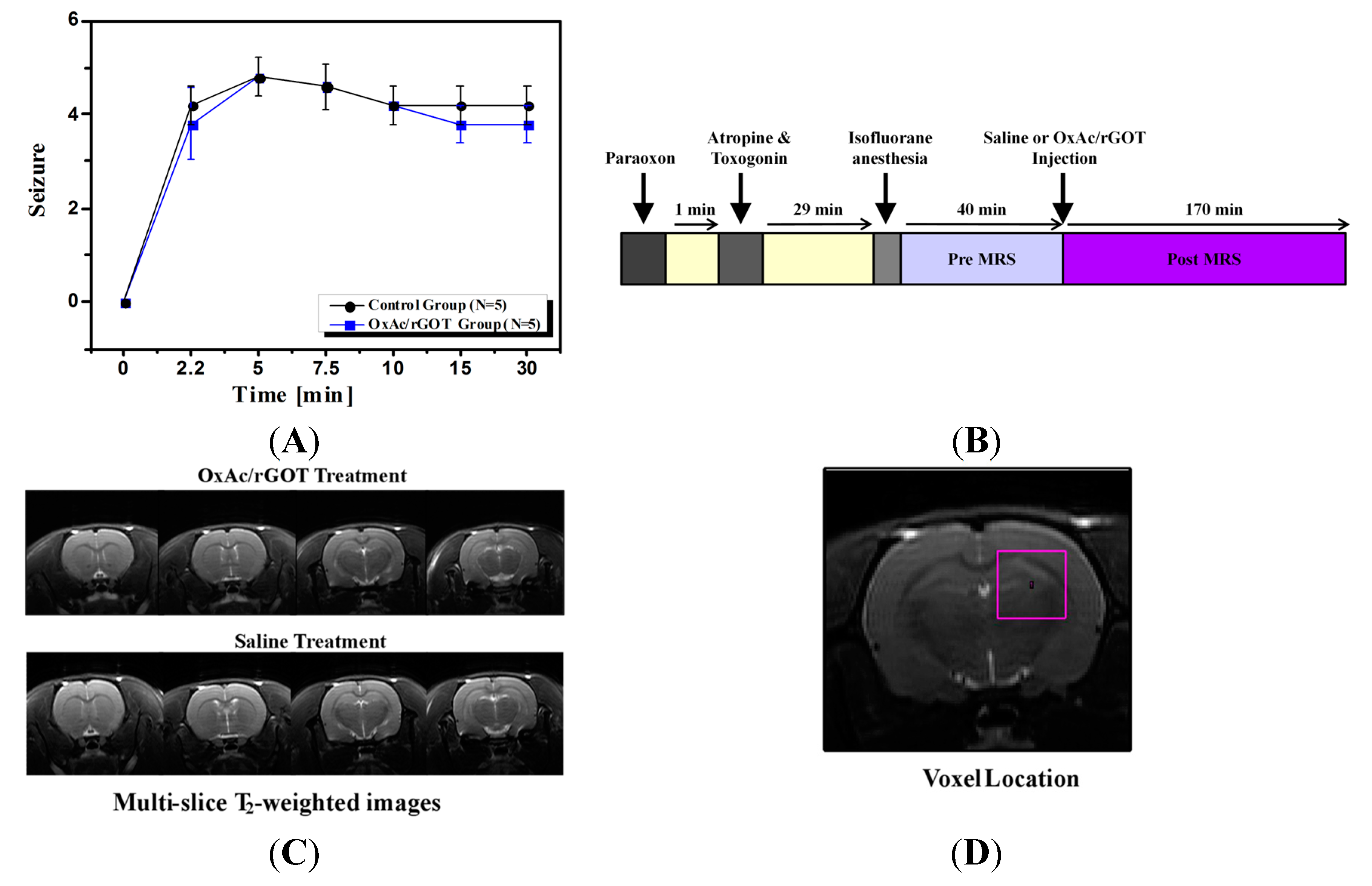
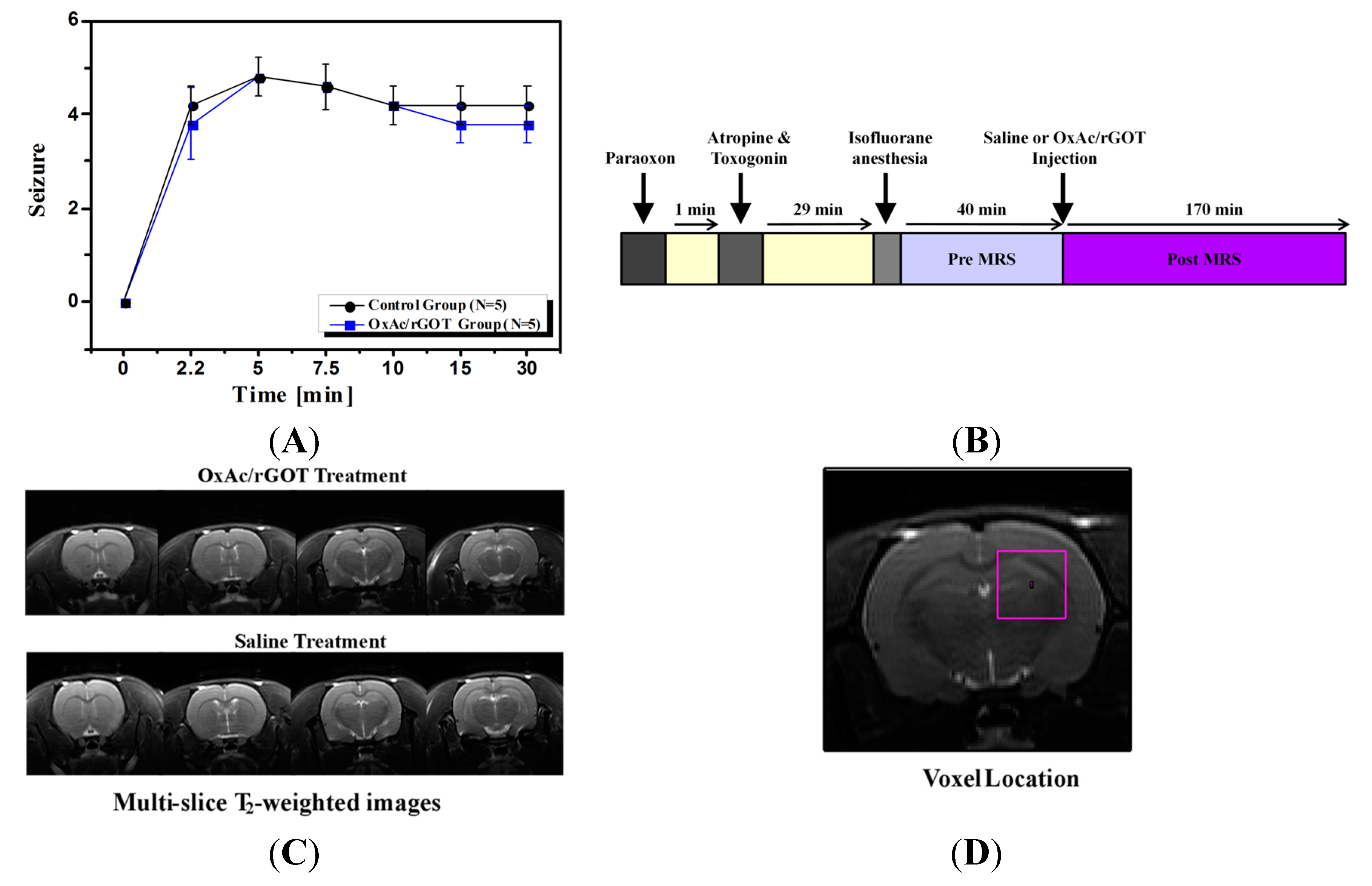
2.1. Clinical Signs
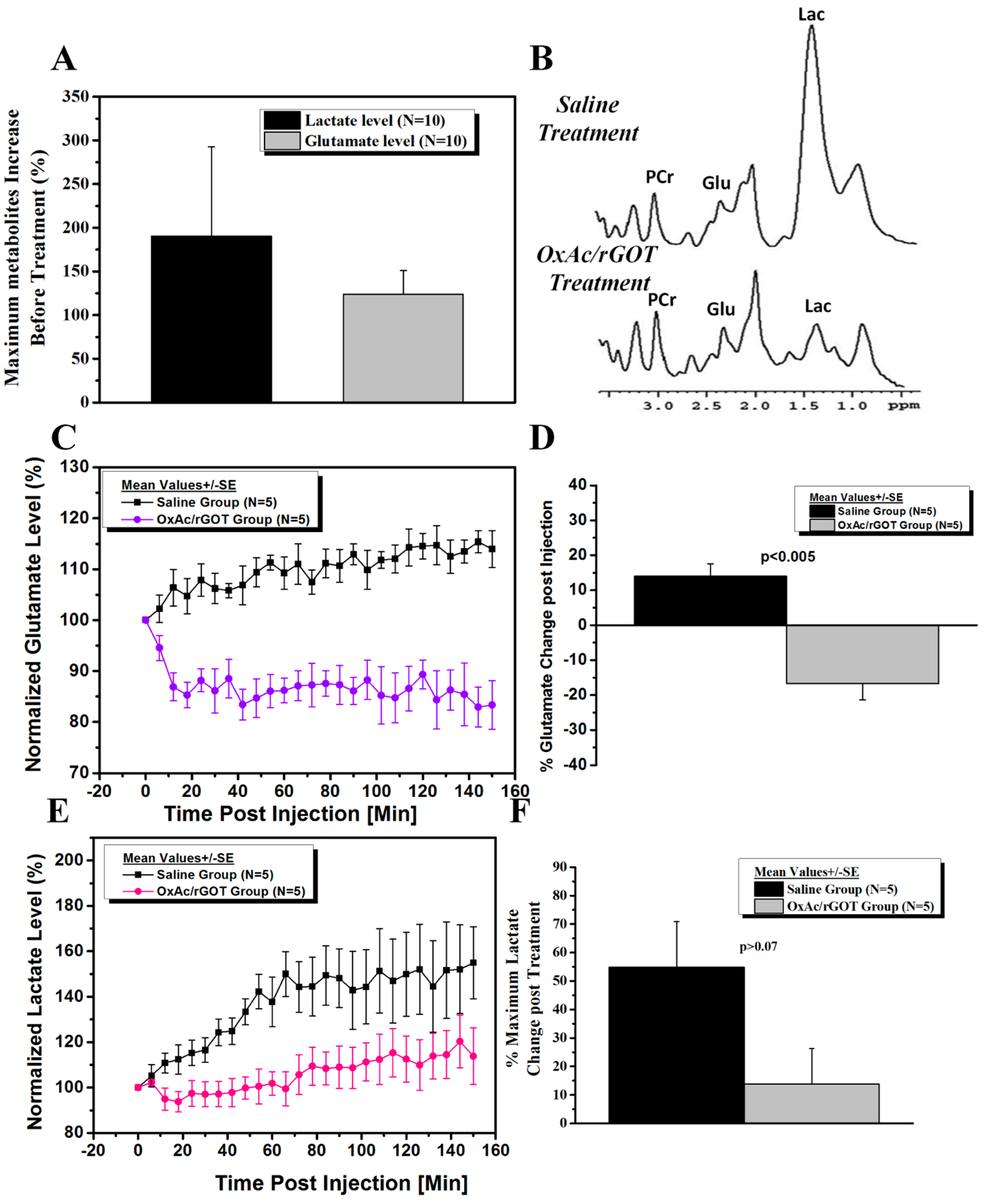
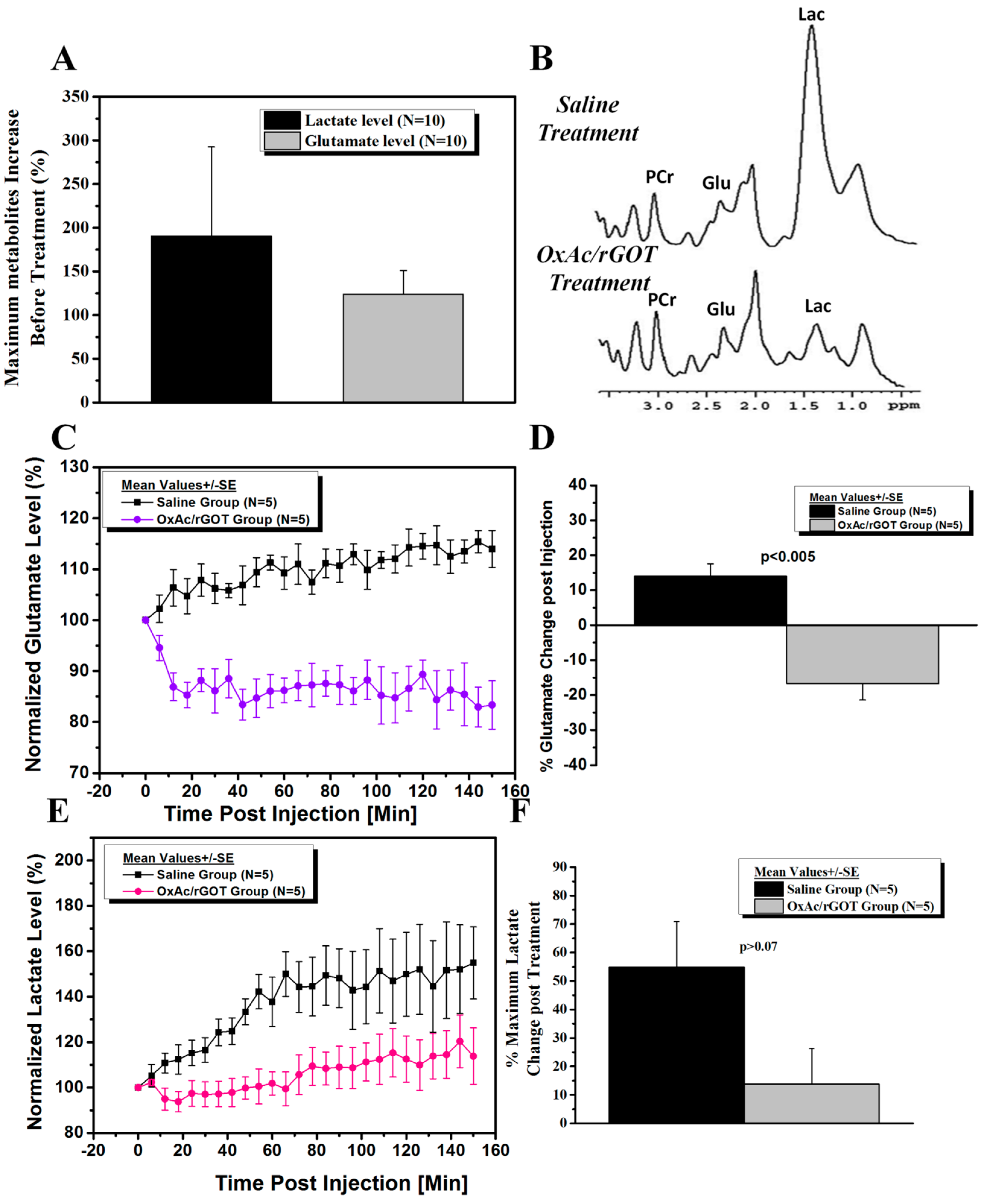
2.2. Effect of Blood Glutamate Scavenging (BGS) Treatment on Brain Glutamate and Lactate Levels
3. Experimental Section
3.1. Materials
3.2. Animals
3.3. Study Design
3.4. Clinical Signs
3.5. Magnetic Resonance Imaging (MRI)
3.6. Magnetic Resonance Spectrocopy (MRS)
3.7. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lallement, G.; Delamanche, I.S.; Pernot-Marino, I.; Baubichon, D.; Denoyer, M.; Carpentier, P.; Blanchet, G. Neuroprotective activity of glutamate receptor antagonists against soman-induced hippocampal damage: Quantification with an ω3 site ligand. Brain Res. 1993, 618, 227–237. [Google Scholar]
- Granacher, R.P., Jr. Traumatic Brain Injury: Methods for Clinical and Forensic Neuropsychiatric Assessment, 2nd ed.; Taylor & Francis Group (CRC Press): Boca Raton, FL, USA, 2008; pp. 18–31. [Google Scholar]
- Lallement, G.; Denoyer, M.; Collet, A.; Pernot-Marino, I.; Baubichon, D.; Monmaur, P.; Blanchet, G. Changes in hippocampal acetylcholine and glutamate extracellular levels during soman-induced seizures: Influence of septal cholinoceptive cells. Neurosci. Lett. 1992, 139, 104–107. [Google Scholar]
- Tattersall, J. Seizure activity post-organophosphate exposure. Front. Biosci. 2009, 14, 3688–3711. [Google Scholar]
- Sparenborg, S.; Brennecke, L.H.; Jaax, N.K.; Braitman, D.J. Dizocilpine (MK-801) arrests status epilepticus and prevents brain damage induced by soman. Neuropharmacology 1992, 31, 357–368. [Google Scholar]
- Cohen-Kashi-Malina, K.; Cooper, I.; Teichberg, V.I. Mechanisms of glutamate efflux at the blood-brain barrier: Involvement of glial cells. J. Cereb. Blood Flow Metab. 2012, 32, 177–189. [Google Scholar]
- Teichberg, V.I. GOT to rid the body of excess glutamate. J. Cereb. Blood Flow Metab. 2011, 31, 1376–1377. [Google Scholar]
- Cederberg-Helms, H.C.; Uhd-Nielsen, C.; Brodin, B. Glutamate efflux at the blood-brain barrier: Cellular mechanisms and potential clinical relevance. Arch. Med. Res. 2015, in press. [Google Scholar]
- Teichberg, V.I.; Cohen-Kashi-Malina, K.; Cooper, I.; Zlotnik, A. Homeostasis of glutamate in brain fluids: An accelerated brain-to-blood efflux of excess glutamate is produced by blood glutamate scavenging and offers protection from neuropathologies. Neuroscience 2009, 158, 301–308. [Google Scholar]
- Zlotnik, A.; Gruenbaum, S.E.; Artru, A.A.; Rozet, I.; Dubilet, M.; Tkachov, S.; Brotfain, E.; Klin, Y.; Shapira, Y.; Teichberg, V.I. The neuroprotective effects of oxaloacetate in closed head injury in rats is mediated by its blood glutamate scavenging activity. J. Neurosurg. Anesth. 2009, 21, 235–241. [Google Scholar]
- Campos, F.; Sobrino, T.; Ramos-Cabrer, P.; Argibay, B.; Agulla, J.; Pérez-Mato, M.; Rodrigues-Gonzalez, R.; Brea, D.; Castillo, J. Neuroprotection by glutamate oxaloacetate transaminase in ischemic stroke: an experimental study. J. Cereb. Blood Flow Metab. 2011, 31, 1378–1386. [Google Scholar]
- Ruban, A.; Berkutzki, T.; Cooper, I.; Mohar, B.; Teichberg, V.I. Blood glutamate scavengers prolong the survival of rats and mice with brain-implanted gliomas. Investig. New Drugs 2012, 30, 2226–2235. [Google Scholar]
- Ruban, A.; Mohar, B.; Jona, G.; Teichberg, V.I. Blood glutamate scavenging as a novel neuroprotective treatment for paraoxon intoxication. J. Cereb. Blood Flow Metab. 2013, 34, 221–227. [Google Scholar]
- Rink, C.; Gnyawali, S.; Peterson, L.; Khanna, S. Oxygen-inducible glutamate oxaloacetate transaminase as protective switch transforming neurotoxic glutamate to metabolic fuel during acute ischemic stroke. Antioxid. Redox Signal. 2011, 14, 1777–1785. [Google Scholar]
- Fauvelle, F.; Carpentier, P.; Dorandeu, F.; Foquin, A.; Testylier, G. Prediction of neuroprotective treatment efficiency using a HRMAS NMR-based statistical model of refractory status epilepticus on mouse: A metabolomic approach supported by histology. J. Proteome Res. 2012, 11, 3782–3795. [Google Scholar]
- Sun, P.Z.; Cheung, J.S.; Wang, E.; Lo, E.H. Association between pH-weighted endogenous amide proton chemical exchange saturation transfer MRI and tissue lactic acidosis during acute ischemic stroke. J. Cereb. Blood Flow Metab. 2011, 31, 1743–1750. [Google Scholar]
- Shrot, S.; Anaby, D.; Krivoy, A.; Makarovsky, I.; Rosman, Y.; Bloch-Shilderman, E.; Lazar, S.; Bar-Shir, A.; Cohen, Y. Early in vivo MR spectroscopy findings in organophosphate-induced brain damage-potential biomarkers for short-term survival. Magn. Reson. Med. 2012, 68, 1390–1398. [Google Scholar]
- Zhang, W.; Wang, P.J.; Li, M.H.; Gao, X.L.; Gu, G.J.; Shao, Z.H. 1H-MRS can monitor metabolites changes of lateral intraventrcular BDNF infusion into a mouse model of Alzheimer’s disease in vivo. Neuroscience 2013, 245, 40–49. [Google Scholar]
- Deshpande, L.S.; Carter, D.S.; Blair, R.E.; DeLorenzo, R.J. Development of a prolonged calcium plateau in hippocampal neurons in rats surviving status epilepticus induced by the organophosphate diisopropylfluorophosphate. Toxicol. Sci. 2010, 116, 623–631. [Google Scholar]
- Valadka, A.B.; Andrews, B.T. Neurotrauma: Evidence-Based Answers to Common Questions; Thieme: New York, NY, USA, 2005; pp. 3–4. [Google Scholar]
- Siesjö, B.K.; Siesjö, P. Mechanisms of secondary brain injury. Eur. J. Anesth. 1996, 13, 247–268. [Google Scholar]
- Sun, P.Z.; Benner, T.; Copen, W.A.; Sorensen, A.G. Early experience of translating pH-weighted MRI to image human subjects at 3 Tesla. Stroke J. Cereb. Circ. 2010, 41, S147–S151. [Google Scholar]
- Lallement, G.; Baubichon, D.; Clarençon, D.; Galonnier, M.; Peoc’h, M.; Carpentier, P. Review of the value of gacyclidine (GK-11) as adjuvant medication to conventional treatments of organophosphate poisoning: primate experiments mimicking various scenarios of military or terrorist attack by soman. Neurotoxicology 1999, 20, 675–684. [Google Scholar]
- Solberg, Y.; Belkin, M. The role of excitotoxicity in organophosphorous nerve agents central poisoning. Trends Pharmacol. Sci. 1997, 18, 183–185. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruban, A.; Biton, I.E.; Markovich, A.; Mirelman, D. MRS of Brain Metabolite Levels Demonstrates the Ability of Scavenging of Excess Brain Glutamate to Protect against Nerve Agent Induced Seizures. Int. J. Mol. Sci. 2015, 16, 3226-3236. https://doi.org/10.3390/ijms16023226
Ruban A, Biton IE, Markovich A, Mirelman D. MRS of Brain Metabolite Levels Demonstrates the Ability of Scavenging of Excess Brain Glutamate to Protect against Nerve Agent Induced Seizures. International Journal of Molecular Sciences. 2015; 16(2):3226-3236. https://doi.org/10.3390/ijms16023226
Chicago/Turabian StyleRuban, Angela, Inbal E. Biton, Arik Markovich, and David Mirelman. 2015. "MRS of Brain Metabolite Levels Demonstrates the Ability of Scavenging of Excess Brain Glutamate to Protect against Nerve Agent Induced Seizures" International Journal of Molecular Sciences 16, no. 2: 3226-3236. https://doi.org/10.3390/ijms16023226
APA StyleRuban, A., Biton, I. E., Markovich, A., & Mirelman, D. (2015). MRS of Brain Metabolite Levels Demonstrates the Ability of Scavenging of Excess Brain Glutamate to Protect against Nerve Agent Induced Seizures. International Journal of Molecular Sciences, 16(2), 3226-3236. https://doi.org/10.3390/ijms16023226