
Comparative Transcriptome Analysis of Genes Involved in GA-GID1-DELLA Regulatory Module in Symbiotic and Asymbiotic Seed Germination of Anoectochilus roxburghii (Wall.) Lindl. (Orchidaceae)

Abstract
:
1. Introduction
2. Results
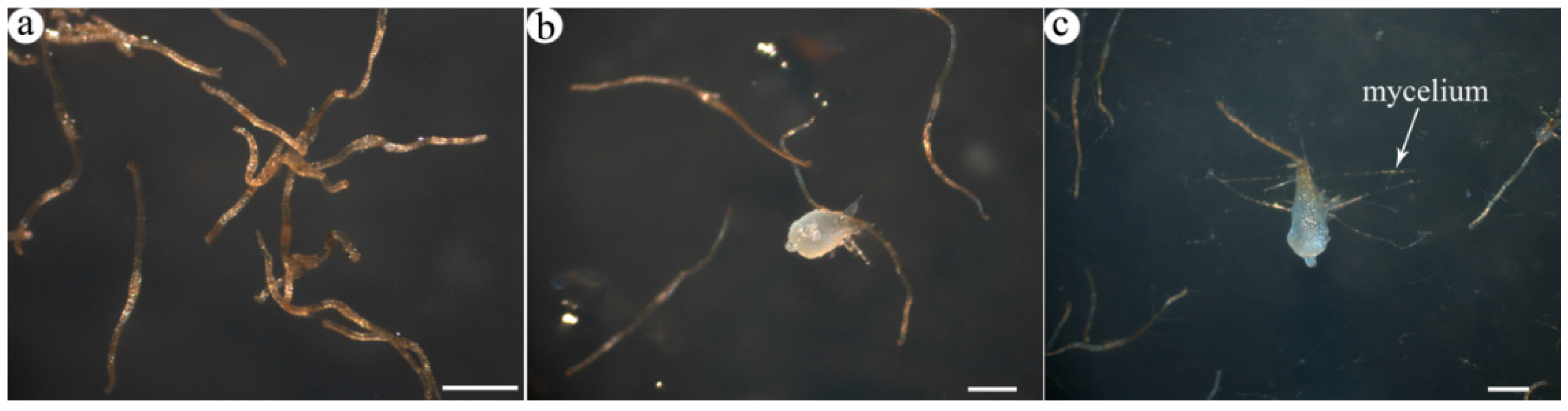
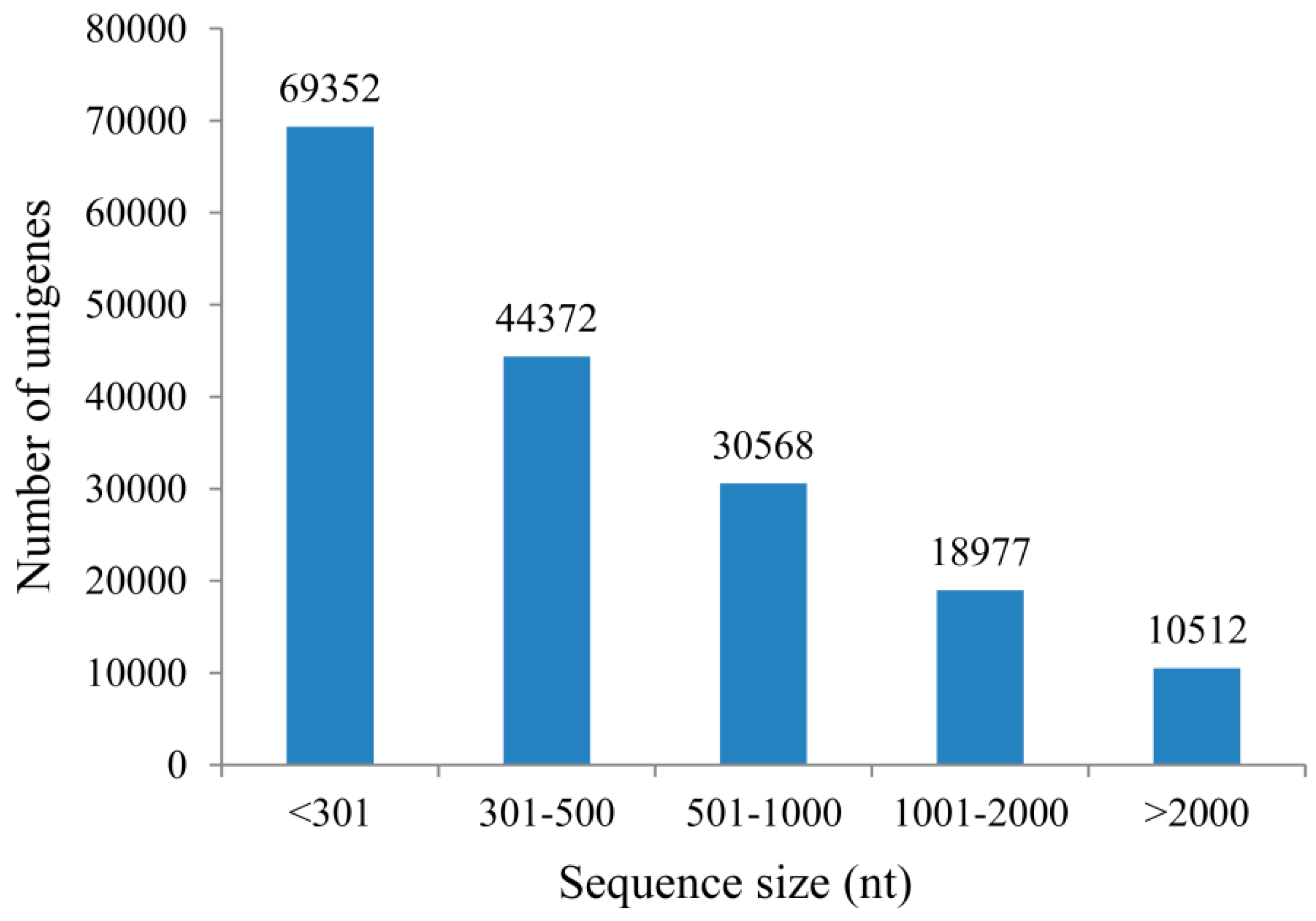
2.1. Transcriptome Profile of A. roxburghii Dry and Germinating Seeds
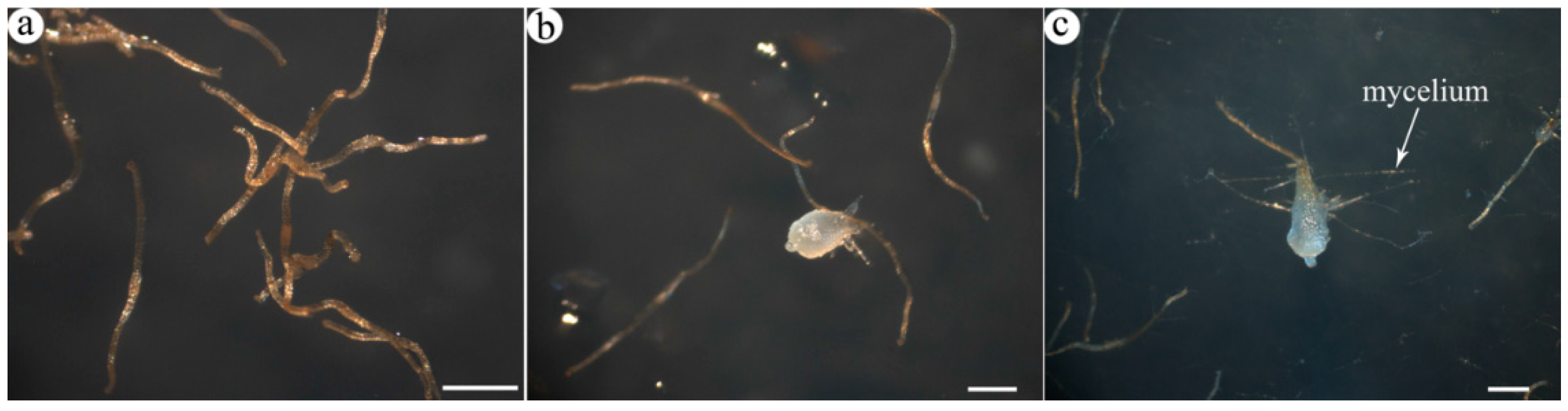

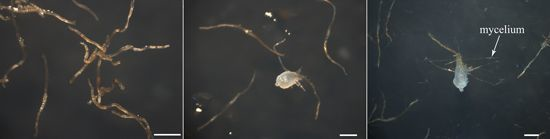{kind=link}
{kind=link}
{kind=link}
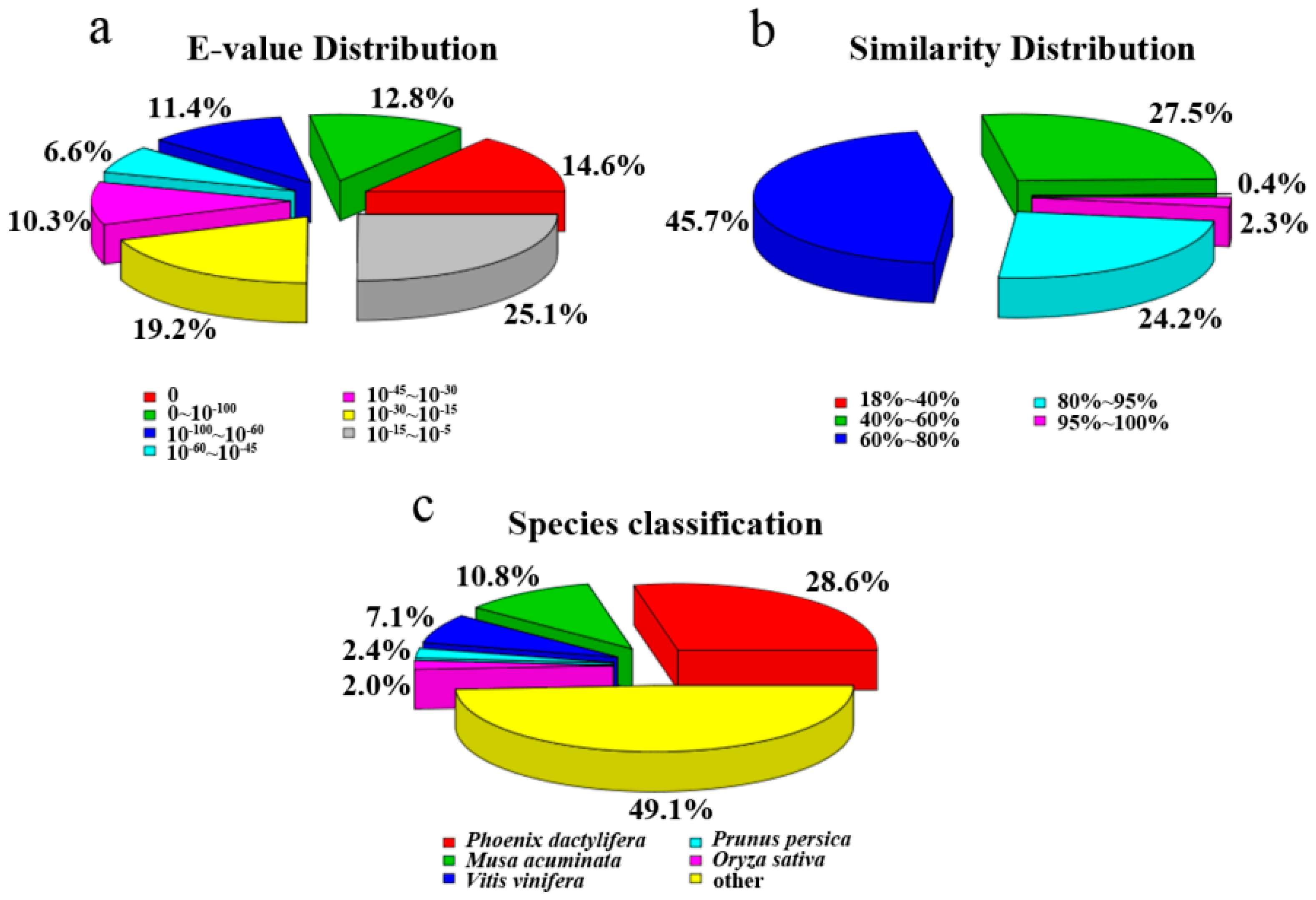
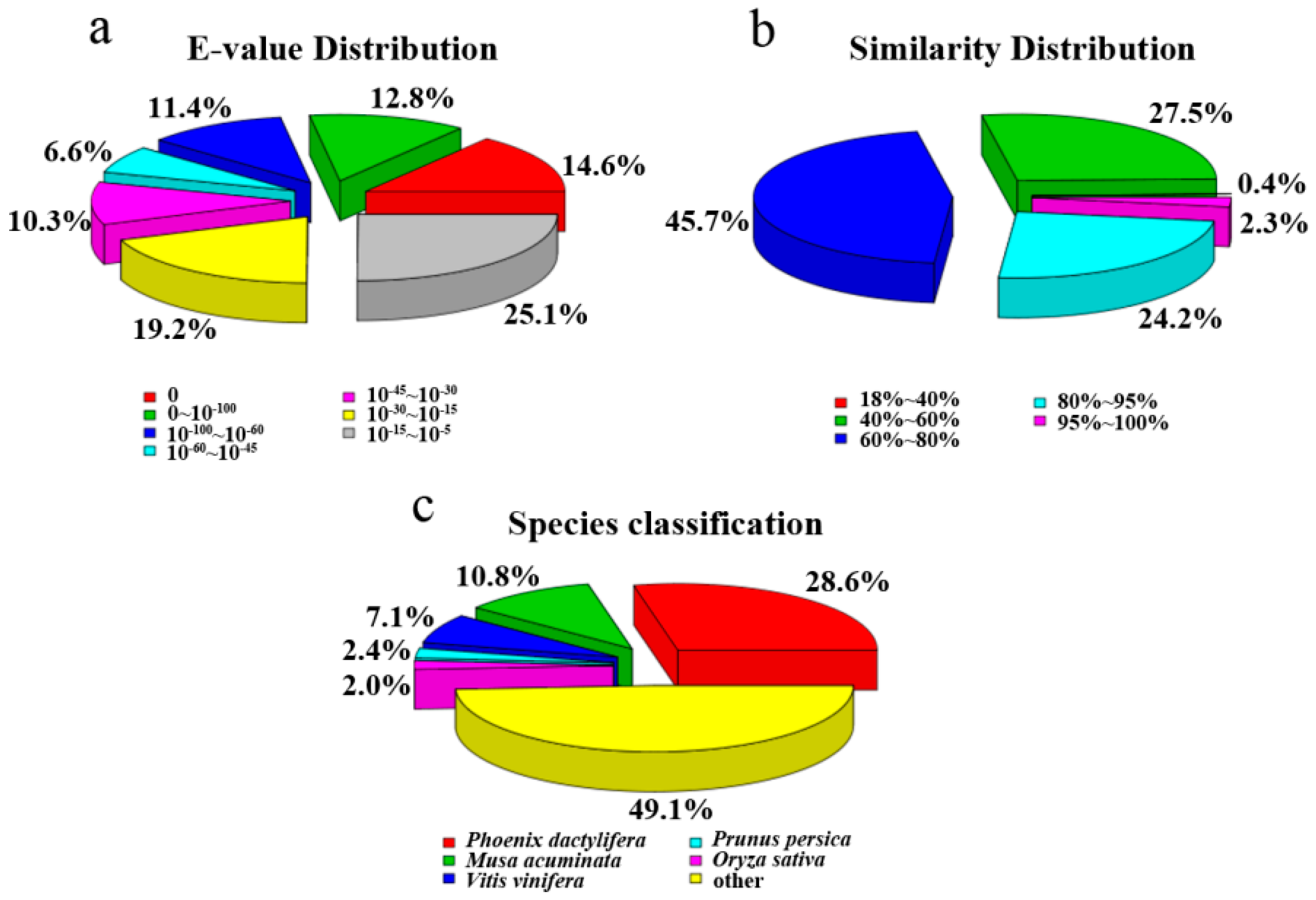{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Error (%) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|
| DS_1 1 | 56,491,050 | 48,108,916 | 0.03 | 94.13 | 86.82 | 51.13 |
| DS_2 1 | 53,963,022 | 45,099,394 | 0.03 | 94.22 | 87.19 | 51.27 |
| AGS_1 2 | 54,662,424 | 41,747,128 | 0.02 | 95.59 | 89.26 | 47.85 |
| AGS_2 2 | 54,531,822 | 42,560,466 | 0.03 | 94.79 | 87.91 | 48.06 |
| SGS_1 3 | 53,273,636 | 43,590,926 | 0.03 | 93.96 | 86.59 | 48.74 |
| SGS_2 3 | 53,140,236 | 44,182,242 | 0.03 | 94.31 | 87.24 | 48.96 |
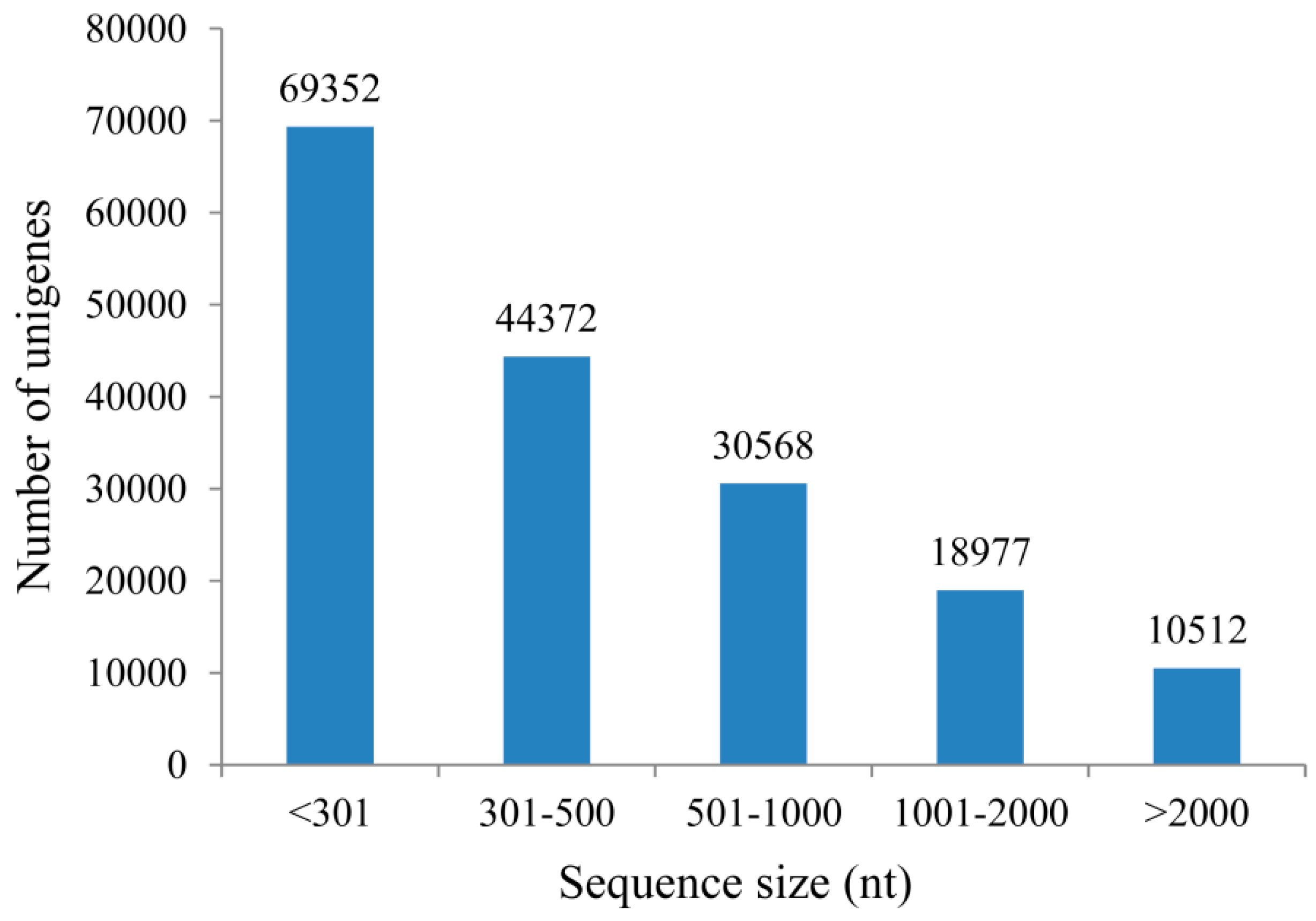
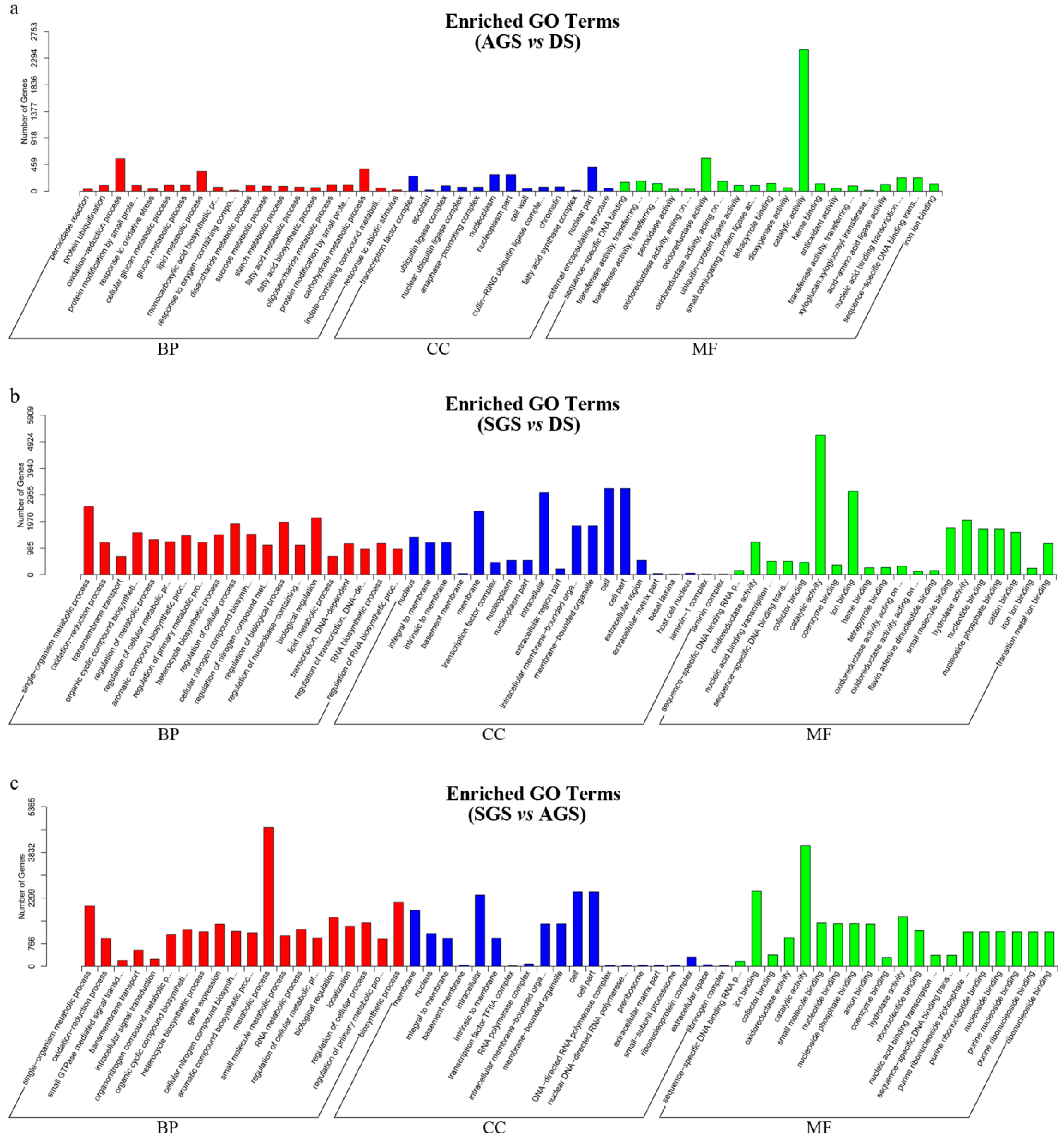
2.2. Unigene Functional Annotation
| Pathway | Gene Name | Number of Unigenes in Transcriptome |
|---|---|---|
| GA biosynthesis | ent-Copalyl diphosphate synthase (CPS) | 2 |
| ent-Kaurene synthase (KS) | 1 | |
| ent-Kaurene oxidase (KO) | 2 | |
| ent-Kaurenoic acid oxidase (KAO) | 1 | |
| GA20-oxidase (GA20ox) | 9 | |
| GA3 beta-hydroxylase (GA3ox) | 5 | |
| GA catabolism | GA2-oxidase (GA2ox) | 11 |
| GA signaling downstream | GA receptor GID1 | 5 |
| DELLA family protein | 11 | |
| F-box protein GID2 | 2 |
2.3. The Unigenes Encoding the Proteins Involved in GA-GID1-DELLA Regulatory Module
2.4. Differential Expression Genes at Seed Germination with or without Mycorrhiza Fungi
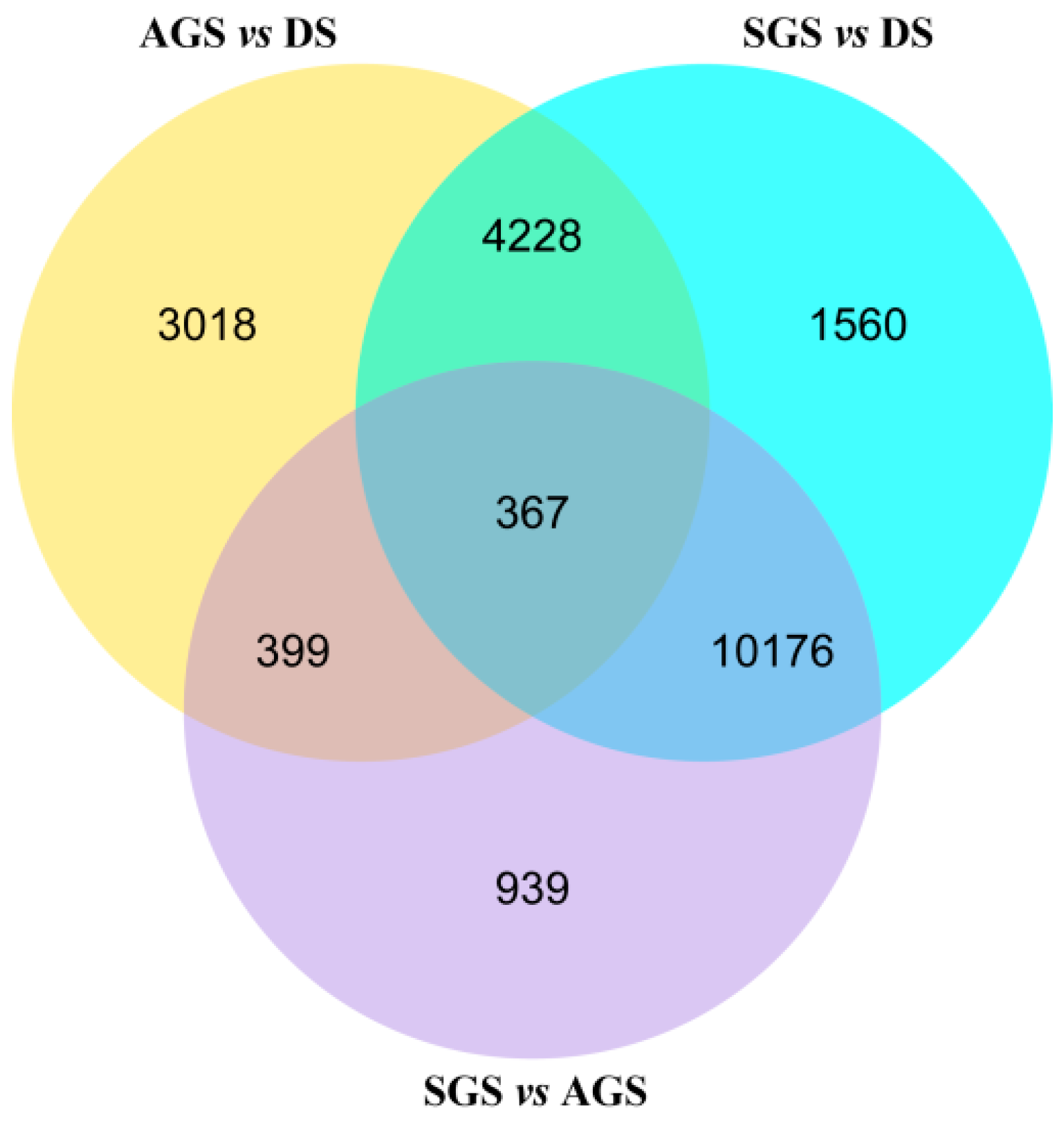
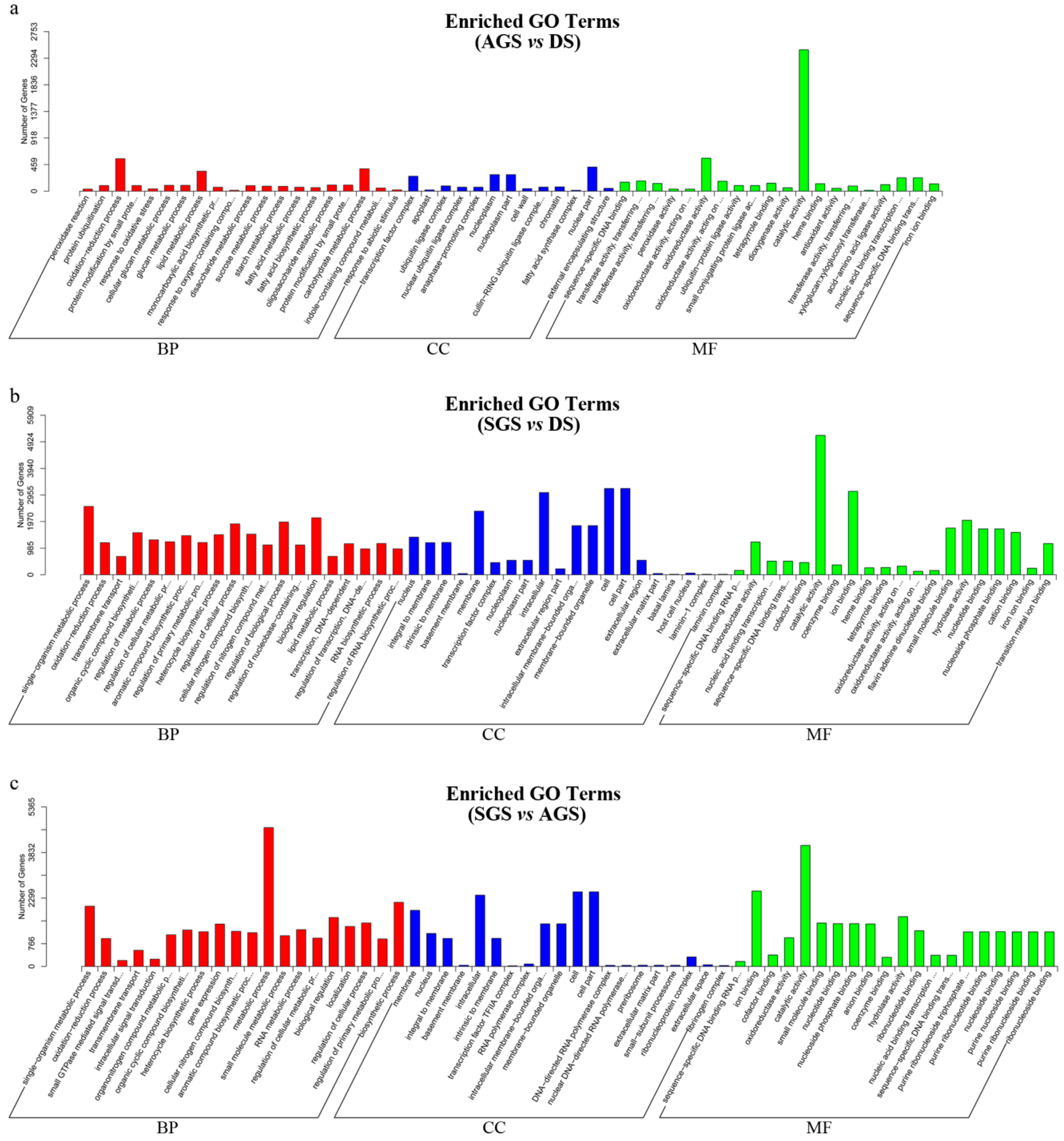
| AGS vs. DS | |||
| # | KEGG Pathway | Number of Unigenes | Percentage (%) |
| 1 | Phenylpropanoid biosynthesis | 89 | 2.91 |
| 2 | Starch and sucrose metabolism | 73 | 2.39 |
| 3 | Carbon metabolism | 71 | 2.32 |
| 4 | Plant hormone signal transduction | 68 | 2.22 |
| 5 | Biosynthesis of amino acids | 68 | 2.22 |
| 6 | Plant-pathogen interaction | 62 | 2.03 |
| 7 | Phenylalanine metabolism | 53 | 1.73 |
| 8 | Glycolysis/Gluconeogenesis | 39 | 1.28 |
| 9 | Protein processing in endoplasmic reticulum | 33 | 1.08 |
| 10 | Cyanoamino acid metabolism | 32 | 1.05 |
| 11 | Endocytosis | 32 | 1.05 |
| 12 | Glycerophospholipid metabolism | 30 | 0.98 |
| 13 | Glyoxylate and dicarboxylate metabolism | 29 | 0.95 |
| 14 | Photosynthesis | 27 | 0.88 |
| 15 | Amino sugar and nucleotide sugar metabolism | 27 | 0.88 |
| 16 | Purine metabolism | 27 | 0.88 |
| 17 | Flavonoid biosynthesis | 26 | 0.85 |
| 18 | Carbon fixation in photosynthetic organisms | 24 | 0.79 |
| 19 | Glutathione metabolism | 23 | 0.75 |
| 20 | Pyruvate metabolism | 23 | 0.75 |
| SGS vs. DS | |||
| # | KEGG Pathway | Number of Unigenes | Percentage (%) |
| 1 | Biosynthesis of amino acids | 135 | 1.70 |
| 2 | Carbon metabolism | 134 | 1.68 |
| 3 | Starch and sucrose metabolism | 112 | 1.41 |
| 4 | Ribosome | 107 | 1.34 |
| 5 | Phenylpropanoid biosynthesis | 105 | 1.32 |
| 6 | Spliceosome | 98 | 1.23 |
| 7 | Purine metabolism | 87 | 1.09 |
| 8 | Protein processing in endoplasmic reticulum | 87 | 1.09 |
| 9 | RNA transport | 83 | 1.04 |
| 10 | Amino sugar and nucleotide sugar metabolism | 71 | 0.89 |
| 11 | Plant-pathogen interaction | 71 | 0.89 |
| 12 | Glycolysis / Gluconeogenesis | 70 | 0.88 |
| 13 | Ubiquitin mediated proteolysis | 66 | 0.83 |
| 14 | Pyrimidine metabolism | 66 | 0.83 |
| 15 | Endocytosis | 66 | 0.83 |
| 16 | Oxidative phosphorylation | 63 | 0.79 |
| 17 | Phenylalanine metabolism | 62 | 0.78 |
| 18 | Cell cycle | 57 | 0.72 |
| 19 | Peroxisome | 56 | 0.70 |
| 20 | Pyruvate metabolism | 55 | 0.69 |
| SGS vs. AGS | |||
| # | KEGG Pathway | Number of Unigenes | Percentage (%) |
| 1 | Biosynthesis of amino acids | 111 | 1.69 |
| 2 | Ribosome | 103 | 1.56 |
| 3 | Carbon metabolism | 97 | 1.47 |
| 4 | Spliceosome | 96 | 1.46 |
| 5 | Protein processing in endoplasmic reticulum | 76 | 1.15 |
| 6 | RNA transport | 75 | 1.14 |
| 7 | Purine metabolism | 72 | 1.09 |
| 8 | Starch and sucrose metabolism | 70 | 1.06 |
| 9 | Oxidative phosphorylation | 64 | 0.97 |
| 10 | Endocytosis | 60 | 0.91 |
| 11 | Ubiquitin mediated proteolysis | 57 | 0.87 |
| 12 | Pyrimidine metabolism | 55 | 0.84 |
| 13 | Phenylpropanoid biosynthesis | 53 | 0.80 |
| 14 | Ribosome biogenesis in eukaryotes | 51 | 0.77 |
| 15 | Amino sugar and nucleotide sugar metabolism | 50 | 0.76 |
| 16 | Cell cycle | 50 | 0.76 |
| 17 | mRNA surveillance pathway | 46 | 0.70 |
| 18 | RNA degradation | 45 | 0.68 |
| 19 | Glycolysis/Gluconeogenesis | 45 | 0.68 |
| 20 | Peroxisome | 43 | 0.65 |
2.5. DEGs Involved in GA-GID1-DELLA Regulatory Module
| Pathway | Gene Name | Number | Unigene ID | log2fold 2 | padj 3 |
|---|---|---|---|---|---|
| GA biosynthesis | ent-Copalyl diphosphate synthase (CPS) | 0 | – | – | – |
| ent-Kaurene synthase (KS) | 0 | – | – | – | |
| ent-Kaurene oxidase (KO) | 0 | – | – | – | |
| ent-Kaurenoic acid oxidase (KAO) | 0 | – | – | – | |
| GA20-oxidase (GA20ox) | 2 | c99861_g1 | 2.17 | 1.97 × 10−3 | |
| c90765_g1 | 4.30 | 8.15 × 10−4 | |||
| GA3 beta-hydroxylase (GA3ox) | 0 | – | – | – | |
| GA catabolism | GA2-oxidase (GA2ox) | 2 | c99070_g1 | 2.98 | 3.03 × 10−7 |
| c106997_g1 | 4.98 | 5.18 × 10−5 | |||
| GA signaling | GA receptor GID1 | 0 | – | – | – |
| DELLA family protein SLR1 | 2 | c85242_g1 | 1.62 | 1.51 × 10−2 | |
| c93049_g1 | 1.83 | 3.58 × 10−3 | |||
| F-box protein GID2 | 0 | – | – | – |
3. Discussion
3.1. The Complexity of Orchid Seed Germination
3.2. The Role of Gibberellins in Seed Germination and Unigenes Related to the GA Signaling Pathway
3.3. Symbiotic Fungi Influence on the GA-GID1-DELLA Regulatory Module
4. Materials and Methods
4.1. Symbiotic and Asymbiotic Germination of Anoectochilus roxburghii Seeds
4.2. Total RNA Extraction
4.3. cDNA Library Construction and Sequencing
4.4. Data Filtering and de Novo Assembly
4.5. Functional Annotation and Metabolic Pathway Analysis of Unigenes
4.6. Analysis of Different Expression Genes
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| AM | arbuscular mycorrhiza |
| AGS | asymbiotic germinating seeds |
| CPS | ent-copalyl diphosphate synthase |
| DEGs | differentially expressed genes |
| DS | dry seeds |
| GA | gibberellin |
| GA20ox | gibberellin 20-oxidase |
| GA3ox | gibberellin 3-oxidase |
| GA2ox | gibberellin 2-oxidase |
| GGPP | trans-geranylgeranyl diphosphate |
| GID1 | GIBBERELLIN INSENSITIVE DWARF1 |
| GO | Gene Ontology |
| KAO | ent-kaurenoic acid oxidase |
| KEGG | the Kyoto Encyclopedia of Genes and Genomes dababase |
| KO | ent-kaurene oxidase |
| KOG | euKaryotic Ortholog Groups database |
| KS | ent-kaurene synthase |
| Nr | NCBI non-redundant protein sequences database |
| Nt | NCBI nucleotide database |
| ODDs | 2-oxoglutarate-dependent dioxygenases |
| Pfam | protein family database |
| RIN | RNA integrity numbers |
| SGS | symbiotic germinating seeds |
| TPSs | terpene synthases |
References
- Jiang, C.; Fu, X. GA action: Turning on de-DELLA repressing signaling. Curr. Opin. Plant Biol. 2007, 10, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.-P.; Gubler, F. Molecular mechanism of gibberellin signaling in plant. Annu. Rev. Plant Biol. 2004, 55, 197–223. [Google Scholar] [CrossRef] [PubMed]
- Voegele, A.; Linkies, A.; Müller, K.; Leubner-Metzger, G. Members of the gibberellin receptor gene family GID1 (GIBBERELLIN INSENSITIVE DWARF1) play distinct roles during Lepidium. sativum and Arabidopsis thaliana seed germination. J. Exp. Bot. 2011, 62, 5131–5147. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Genschik, P. Releasing the brakes of plant growth: How GAs shutdown DELLA proteins. J. Exp. Bot. 2009, 60, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.-P. The molecular mechanism and evolution of the GA–GID1–DELLA signaling module in plants. Curr. Biol. 2011, 21, R338–R345. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Deng, D.-X. Molecular basis and evolutionary pattern of GA–GID1–DELLA regulatory module. Mol. Genet. Genom. 2014, 289, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Delectis Florae Reipublicae Popularis Sinicae, Agendae Academiae Sinicae Edita. In Flora Reipublicae Popularis Sinicae; Tomus 27; Science Press: Beijing, China, 1999; p. 220.
- He, C.-N.; Wang, C.-L.; Guo, S.-X.; Yang, J.-S.; Xiao, P.-G. A novel flavonoid glucoside from Anoectochilus roxburghii (Wall.) Lindl. J. Integr. Plant Biol. 2006, 48, 359–363. [Google Scholar] [CrossRef]
- Gutiérrez, R.M.P. Orchids: A review of uses in traditional medicine, its phytochemistry and pharmacology. J. Med. Plant Res. 2010, 4, 592–638. [Google Scholar]
- Fujian Institute of Traditional Chinese Medicine. Record of Fujian Materia Medica, 2nd ed.; Fujian Science and Technology Press: Fuzhou, China, 1982. [Google Scholar]
- Stewart, S.L.; Kane, M.E. Asymbiotic seed germination and in vitro seedling development of Habenaria. macroceratitis (Orchidaceae), a rare Florida terrestrial orchid. Plant Cell Tissue Organ 2006, 86, 147–158. [Google Scholar] [CrossRef]
- Arditti, J. Factors affecting the germination of orchid seeds. Bot. Rev. 1967, 33, 1–97. [Google Scholar] [CrossRef]
- Johnson, T.R.; Stewart, S.L.; Dutra, D.; Kane, M.E.; Richardson, L. Asymbiotic and symbiotic seed germination of Eulophia. alta (Orchidaceae)—Preliminary evidence for the symbiotic culture advantage. Plant Cell Tissue Organ 2007, 90, 313–323. [Google Scholar] [CrossRef]
- Zhao, M.-M.; Zhang, G.; Zhang, D.-W.; Hsiao, Y.-Y.; Guo, S.-X. ESTs analysis reveals putative genes involved in symbiotic seed germination in Dendrobium officinale. PLoS ONE 2013, 8, e72705. [Google Scholar] [CrossRef] [PubMed]
- Perotto, S.; Rodda, M.; Benetti, A.; Sillo, F.; Ercole, E.; Rodda, M.; Girlanda, M.; Murat, C.; Balestrini, R. Gene expression in mycorrhizal orchid protocorms suggests a friendly plant–fungus relationship. Planta 2014, 239, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Tyler, L.; Thomas, S.G.; Hu, J.; Dill, A.; Alonso, J.M.; Ecker, J.R.; Sun, T.-P. DELLA proteins and gibberellin-regulated seed germination and floral development in Arabidopsis. Plant Physiol. 2004, 135, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Nambara, E.; Choi, G.; Yamaguchi, S. Interaction of light and hormone signals in germinating seeds. Plant Mol. Biol. 2009, 69, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Hu, J.; Mitchum, M.G.; Barnaby, N.; Ayele, B.T.; Ogawa, M.; Nam, E.; Lai, W.-C.; Hanada, A.; Alonso, J.M.; Ecker, J.R.; et al. Potential sites of bioactive gibberellin production during reproductive growth in Arabidopsis. Plant Cell 2008, 20, 320–336. [Google Scholar] [CrossRef] [PubMed]
- Murase, K.; Hirano, Y.; Sun, T.P.; Hakoshima, T. Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature 2008, 456, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Floss, D.S.; Levy, J.G.; Lévesque-Tremblay, V.; Pumplin, N.; Harrison, M.J. DELLA proteins regulate arbuscule formation in arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2013, 110, E5025–E5034. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, H.N. Terrestrial Orchids: From Seed to Mycotrophic Plant; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Zettler, L.W.; Hofer, C.J. Propagation of the little club-spur orchid (Platanthera. clavellata) by symbiotic seed germination and its ecological implications. Environ. Exp. Bot. 1998, 39, 189–195. [Google Scholar] [CrossRef]
- Weiss, D.; Ori, N. Mechanisms of cross talk between gibberellin and other hormones. Plant Physiol. 2007, 144, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.-P.; Kamiya, Y. The Arabidopsis GA1 locus encodes the cyclase ent-kaurene synthetase A of gibberellin biosynthesis. Plant Cell 1994, 6, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Cheng, H.; Wu, W.; Soo, H.M.; Peng, J. Gibberellin mobilizes distinct DELLA-dependent transcriptomes to regulate seed germination and floral development in Arabidopsis. Plant Physiol. 2006, 142, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Richards, D.E.; Fleck, B.; Xie, D.; Burton, N.; Harberd, N.P. The Arabidopsis mutant sleepy1gar2−1 protein promotes plant growth by increasing the affinity of the SCFSLY1 E3 ubiquitin ligase for DELLA protein substrates. Plant Cell 2004, 16, 1406–1418. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Ueguchi-Tanaka, M.; Sato, Y.; Ashikari, M.; Matsuoka, M. The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in Nuclei. Plant Cell 2002, 14, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Cao, D.-N.; Cheng, H.; Wen, Z.-L.; Peng, J.-R. Identification of the conserved serine/threonine residues important for gibberellin-sensitivity of Arabidopsis RGL2 protein. Plant J. 2005, 44, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Harberd, N.P.; Belfield, E.; Yasumura, Y. The angiosperm gibberellin-GID1-DELLA growth regulatory mechanism: how an “inhibitor of an inhibitor” enables flexible response to fluctuating environments. Plant Cell 2009, 21, 1328–1339. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, F.M.; Bizzell, C.M.; Lee, D.J.; Zeevaart, J.A.; Amasino, R.M. Overexpression of a novel class of gibberellin 2-oxidases decreases gibberellin levels and creates dwarf plants. Plant Cell 2003, 15, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.G.; Phillips, A.L.; Hedden, P. Molecular cloning and functional expression of gibberellin 2-oxidases, multifunctional enzymes involved in gibberellin deactivation. Proc. Natl. Acad. Sci. USA 1999, 96, 4638–4703. [Google Scholar] [CrossRef]
- Toh, S.; Imamura, A.; Watanabe, A.; Nakabayashi, K.; Okamoto, M.; Jikumaru, J.; Hanada, A.; Aso, Y.; Ishiyama, K.; Tamura, N.; et al. High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds. Plant Physiol. 2008, 146, 1368–1385. [Google Scholar] [CrossRef] [PubMed]
- Fleet, C.M.; Sun, T.-P. A DELLAcate balance: The role of gibberellin in plant morphogenesis. Curr. Opin. Plant Biol. 2005, 8, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Gutjahr, C. Phytohormone signaling in arbuscular mycorhiza development. Curr. Opin. Plant Biol. 2014, 20, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Foo, E.; Ross, J.J.; Jones, W.T.; Reid, J.B. Plant hormones in arbuscular mycorrhizal symbioses: An emerging role for gibberellins. Ann. Bot. 2013, 111, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Gismondi, A.; Rolfo, M.F.; Leonardi, D.; Rickards, O.; Canini, A. Identification of ancient Olea europaea L. and Cornus. mas L. by DNA barcoding. Cr. Biol. 2012, 335, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Gismondi, A.; Fanali, F.; Martínez Labarga, J.M.; Grilli Caiola, M.; Canini, A. Crocus sativus L. genomics and different DNA barcode applications. Plant Syst. Evol. 2013, 299, 1859–1863. [Google Scholar] [CrossRef]
- Gismondi, A.; di Marco, G.; Delorenzo, M.; Canini, A. Upgrade of Castanea. sativa (Mill.) genetic resources by sequencing of barcode markers. J. Genet. 2015, 94, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- NCBI. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 25 November 2015).
- UniProt. Available online: http://www.ebi.ac.uk/uniprot/ (accessed on 25 November 2015).
- KOG/COG. Available online: http://www.ncbi.nlm.nih.gov/COG/ (accessed on 25 November 2015).
- Pfam. Available online: http://pfam.sanger.ac.uk/ (accessed on 25 November 2015).
- Gene Ontology Consortium. Available online: http://geneontology.org/ (accessed on 25 November 2015).
- KEGG. Available online: http://www.genome.jp/kegg/ (accessed on 25 November 2015).
- Li, B.; Dewey, C. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
- Rasmussen, H.N.; Dixon, K.W.; Jersáková, J.; Těšitelová, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. 2015. [Google Scholar] [CrossRef] [PubMed]
- Potokina, E.; Sreenivasulu, N.; Altschmied, L.; Michalek, W.; Graner, A. Differential gene expression during seed germination in barley (Hordeum vulgare L.). Funct. Integr. Genom. 2002, 2, 28–39. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.-S.; Chen, J.; Li, S.-C.; Zeng, X.; Meng, Z.-X.; Guo, S.-X. Comparative Transcriptome Analysis of Genes Involved in GA-GID1-DELLA Regulatory Module in Symbiotic and Asymbiotic Seed Germination of Anoectochilus roxburghii (Wall.) Lindl. (Orchidaceae). Int. J. Mol. Sci. 2015, 16, 30190-30203. https://doi.org/10.3390/ijms161226224
Liu S-S, Chen J, Li S-C, Zeng X, Meng Z-X, Guo S-X. Comparative Transcriptome Analysis of Genes Involved in GA-GID1-DELLA Regulatory Module in Symbiotic and Asymbiotic Seed Germination of Anoectochilus roxburghii (Wall.) Lindl. (Orchidaceae). International Journal of Molecular Sciences. 2015; 16(12):30190-30203. https://doi.org/10.3390/ijms161226224
Chicago/Turabian StyleLiu, Si-Si, Juan Chen, Shu-Chao Li, Xu Zeng, Zhi-Xia Meng, and Shun-Xing Guo. 2015. "Comparative Transcriptome Analysis of Genes Involved in GA-GID1-DELLA Regulatory Module in Symbiotic and Asymbiotic Seed Germination of Anoectochilus roxburghii (Wall.) Lindl. (Orchidaceae)" International Journal of Molecular Sciences 16, no. 12: 30190-30203. https://doi.org/10.3390/ijms161226224
APA StyleLiu, S.-S., Chen, J., Li, S.-C., Zeng, X., Meng, Z.-X., & Guo, S.-X. (2015). Comparative Transcriptome Analysis of Genes Involved in GA-GID1-DELLA Regulatory Module in Symbiotic and Asymbiotic Seed Germination of Anoectochilus roxburghii (Wall.) Lindl. (Orchidaceae). International Journal of Molecular Sciences, 16(12), 30190-30203. https://doi.org/10.3390/ijms161226224