
Allostatic Load and Preterm Birth


Abstract
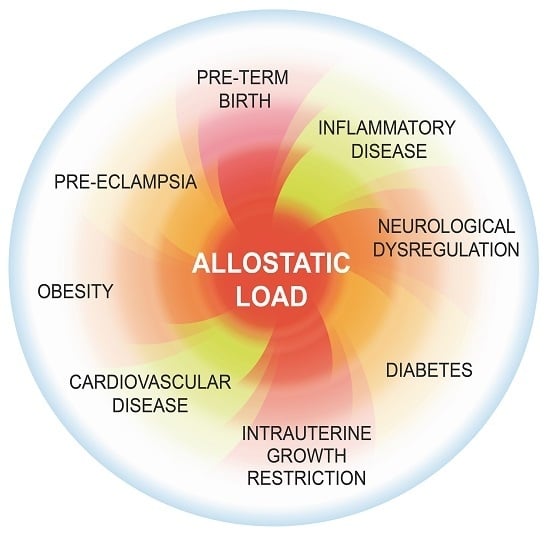
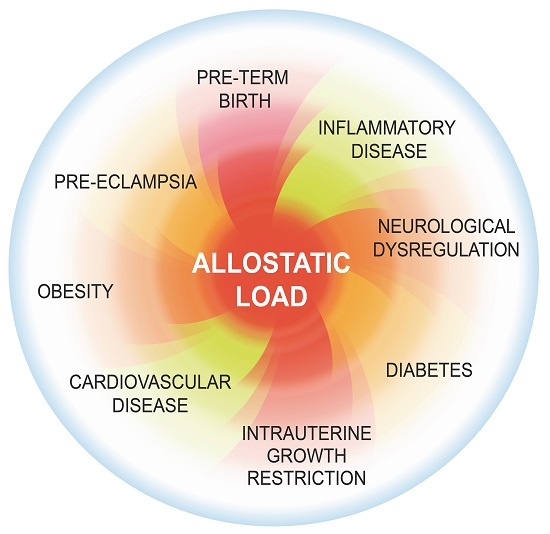
:
1. Introduction
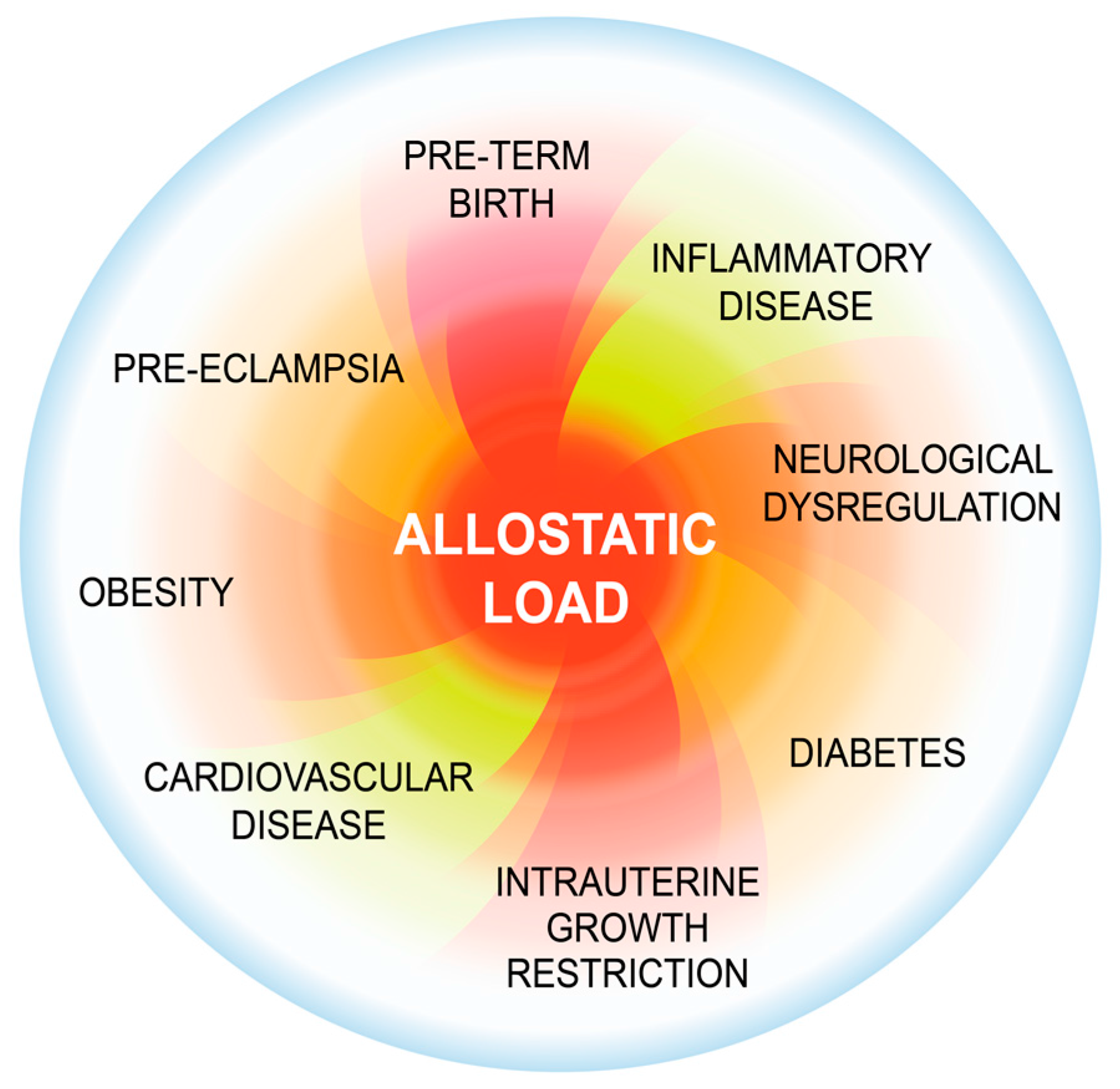
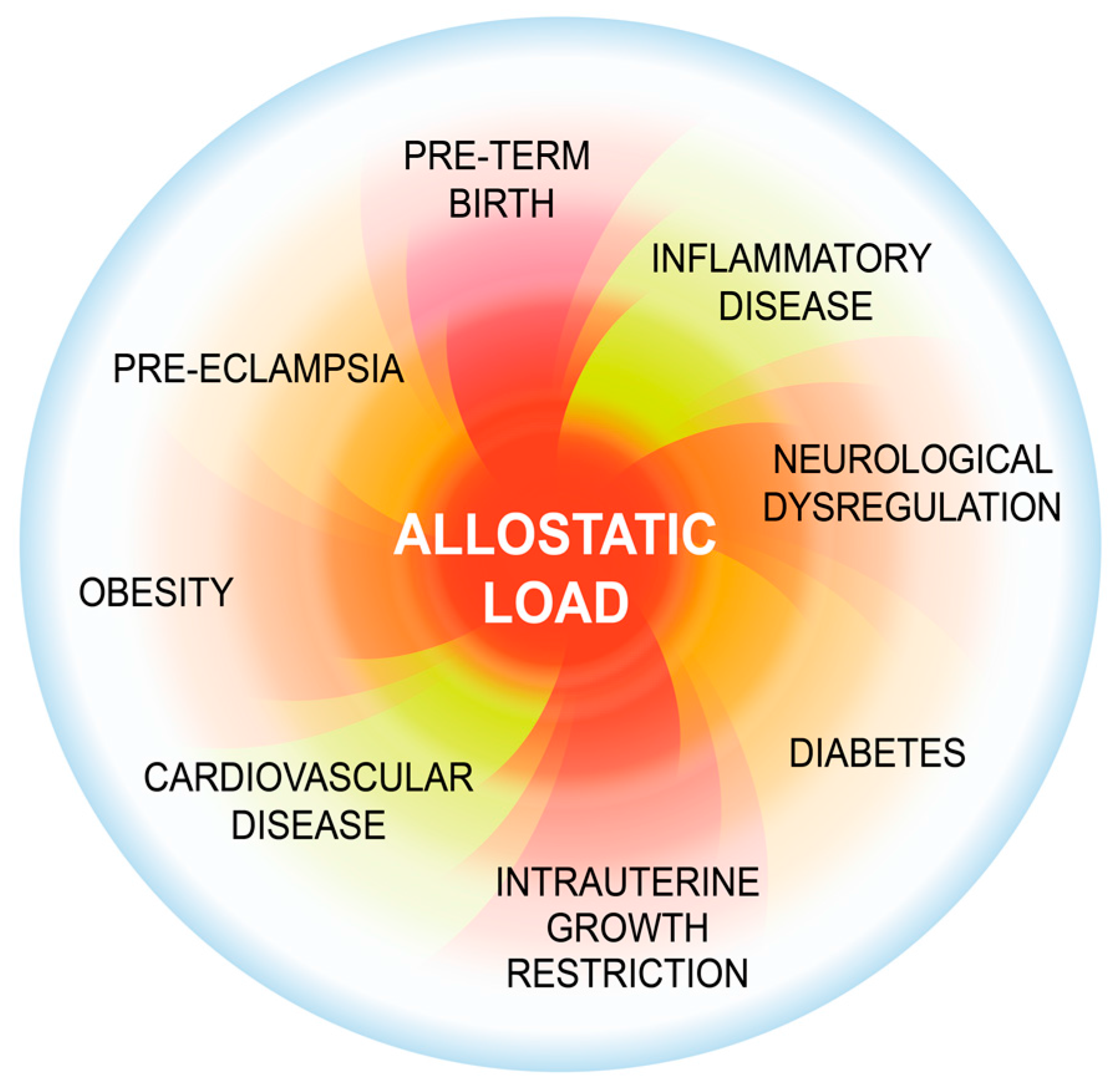
2. Background
2.1. Scope of the Issue
2.2. Our Working Construct
3. Evidence to Support Construct
3.1. Previous Attempts to Link Allostatic Load with Pregnancy and Birth Outcomes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Data Source | Population | Data Collection | Biomarkers | Allostatic Load Scoring | Outcome |
|---|---|---|---|---|---|---|
| Wallace et al., 2013 [26] | Bogalusa Heart Study | African-American women | Preconception |
| Contribution of each biomarker value to AL index weighted by loadings on the first principal component. This linear AL index is split into quartiles for analysis. Higher quartiles represent greater AL. | No evidence of a relationship between maternal preconception allostatic load and preterm birth or low birth weight infants. |
| Wallace et al., 2013[18] | Bogalusa Heart Study | African-American women | Preconception |
| Score of 1 or 0 based on whether biomarker within high risk percentile or below based on data sample’s distribution, respectively. Score is summed for each biomarker to obtain AL score ranging from 0 to 9, which larger score indicated higher AL. | No evidence of a relationship between maternal preconception allostatic load and preterm birth or low birth weight infants. |
| Wallace and Harville, 2013 [28] | Tulane-Lakeside Hospital Department of Obstetrics and Gynecology | White or African-American | Pregnant: 26–28 weeks gestation |
| Uses z-score for each biomarker based on the data sample’s distribution. AL score for each subject is the sum of z-scores. Higher scores presents higher AL. | Gestational age decreased significantly with increasing allostatic load. |
| Morrison et al., 2013 [27] | NHANES 1999–2006 | Civilian noninstitutionalized US population | Pregnant and non-pregnant |
| Score of 1 or 0 based on whether biomarker within high-risk percentile or below based on data sample’s distribution, respectively. Score is summed for each biomarker to obtain AL score ranging from 0 to 10, which larger score indicated higher AL. | AL may reflect proximal factors in pregnancy more strongly than they represent exposure to chronic stress over a woman’s lifetime. |
| Hux et al., 2014 [19] | NHANES 1999–2006 | Civilian noninstitutionalized US population | History of low birth weight infants and those who were preterm |
| Score of 1 or 0 based on whether biomarker is within high risk percentile or below based on data sample’s distribution, respectively. Score is summed for each biomarker to obtain AL score ranging from 0 to 9, which larger score indicated higher AL. | Women with history of SGA or PTB had higher AL than did those with normal birth weight outcomes. |
3.2. Chronic life Stress in Women
3.3. Animal Models
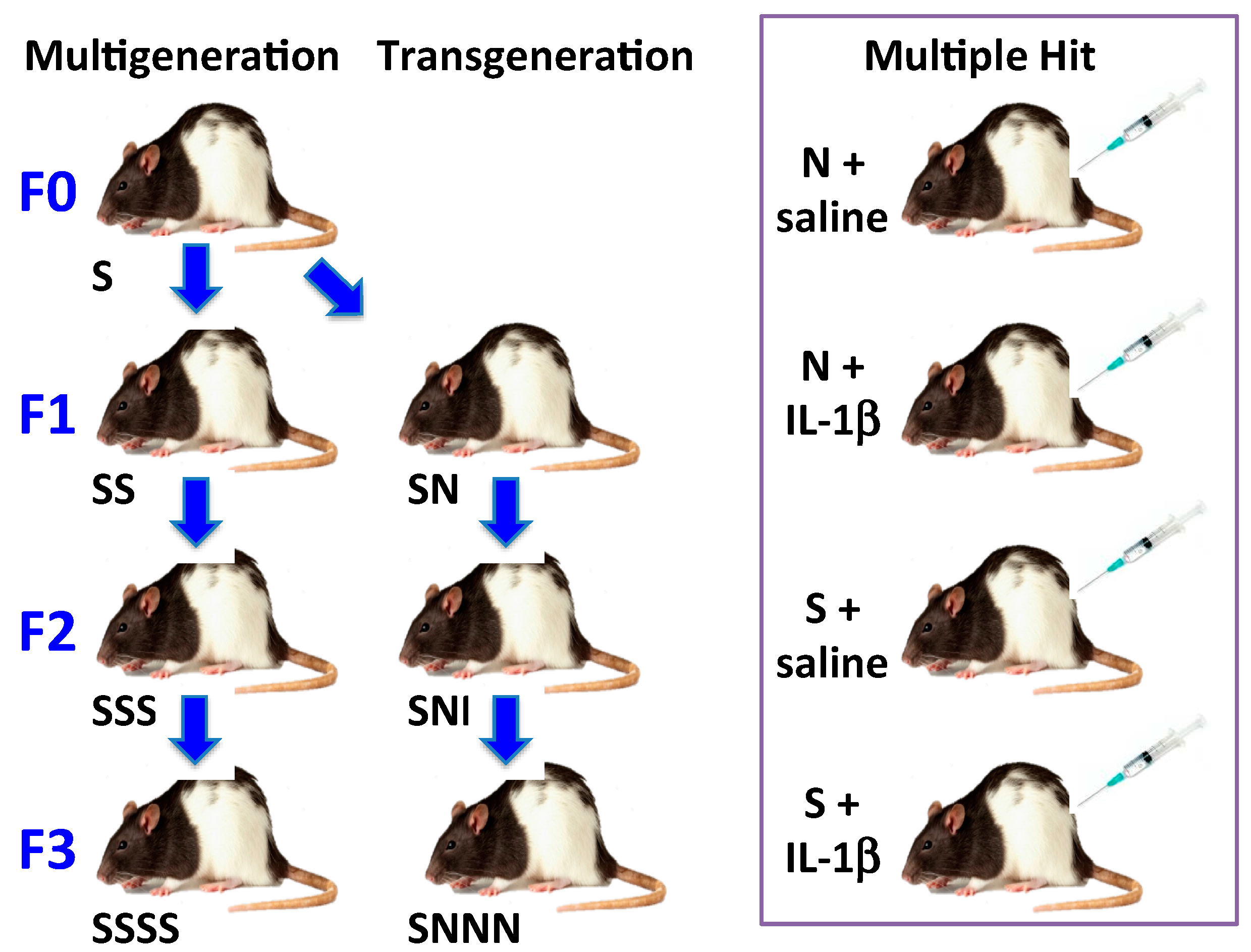
3.3.1. Stress by Generations
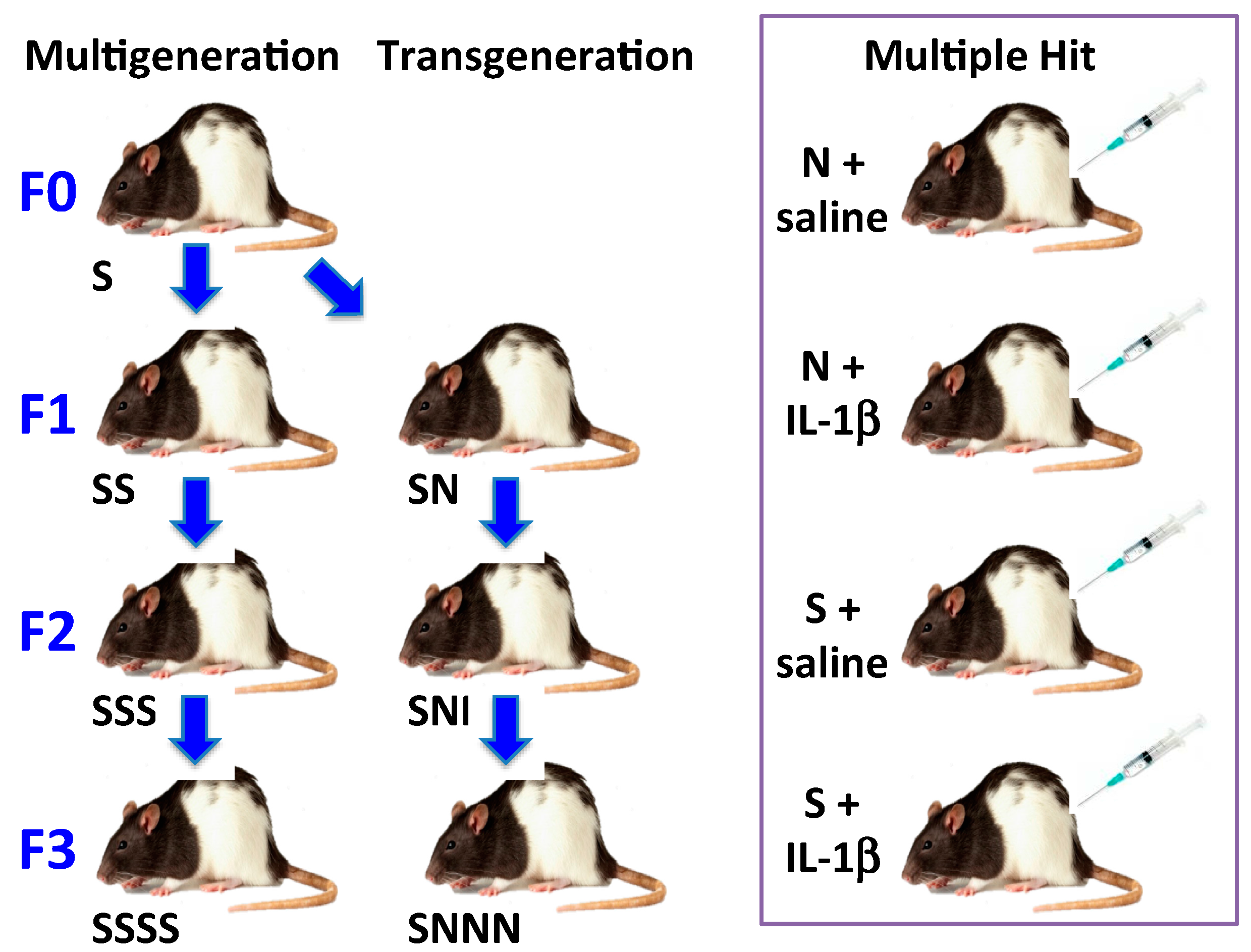
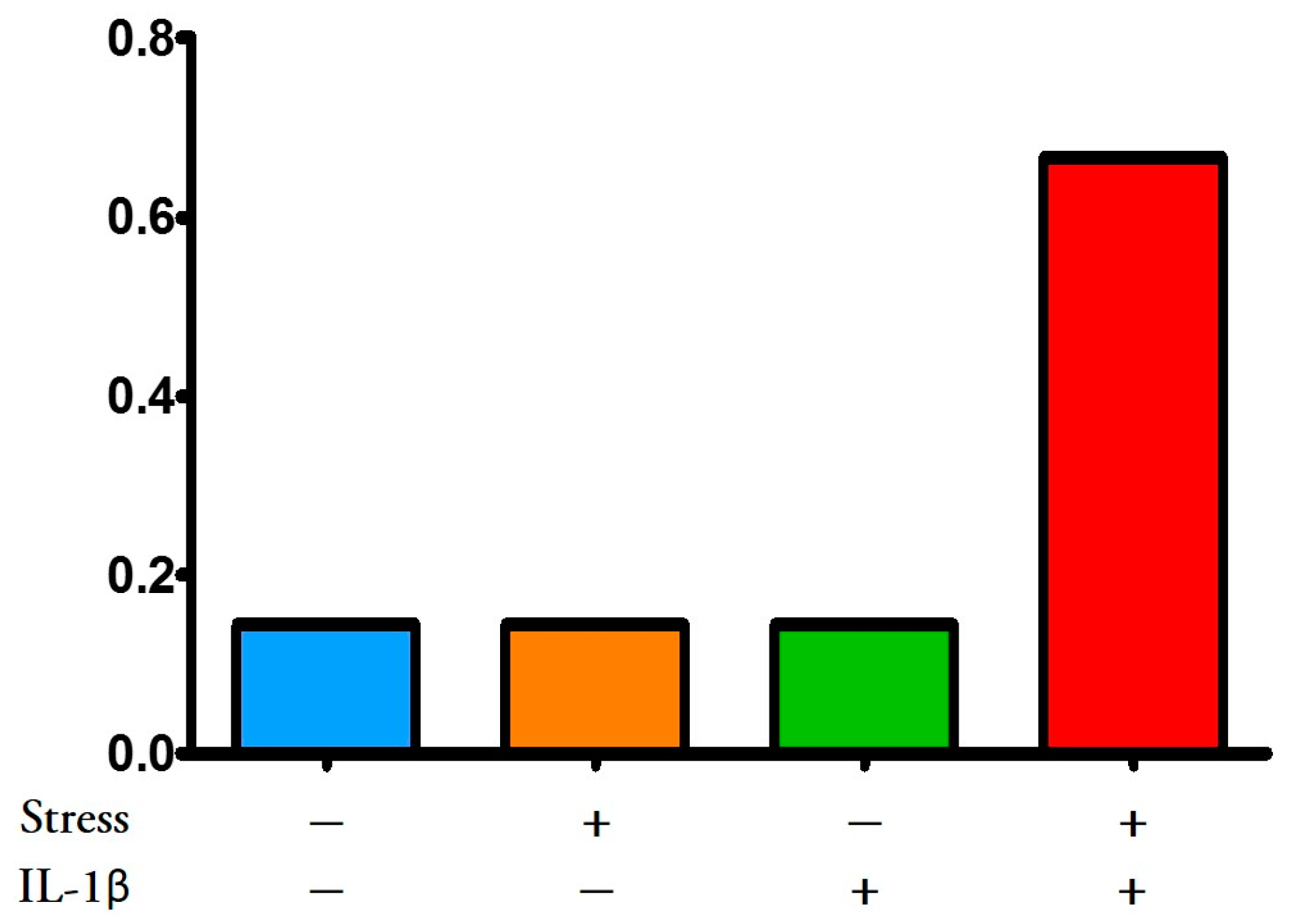
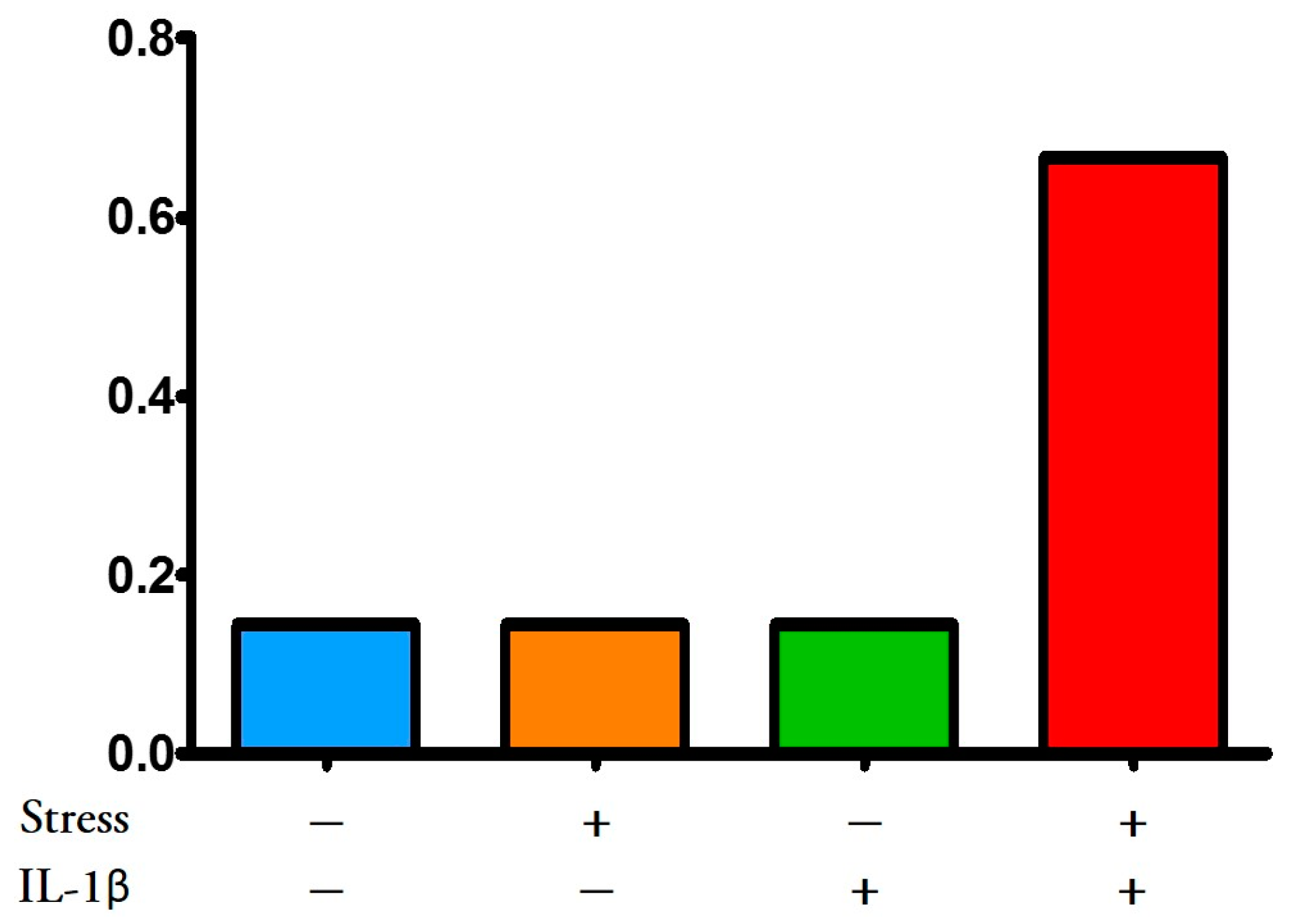
3.3.2. Multiple hit Model
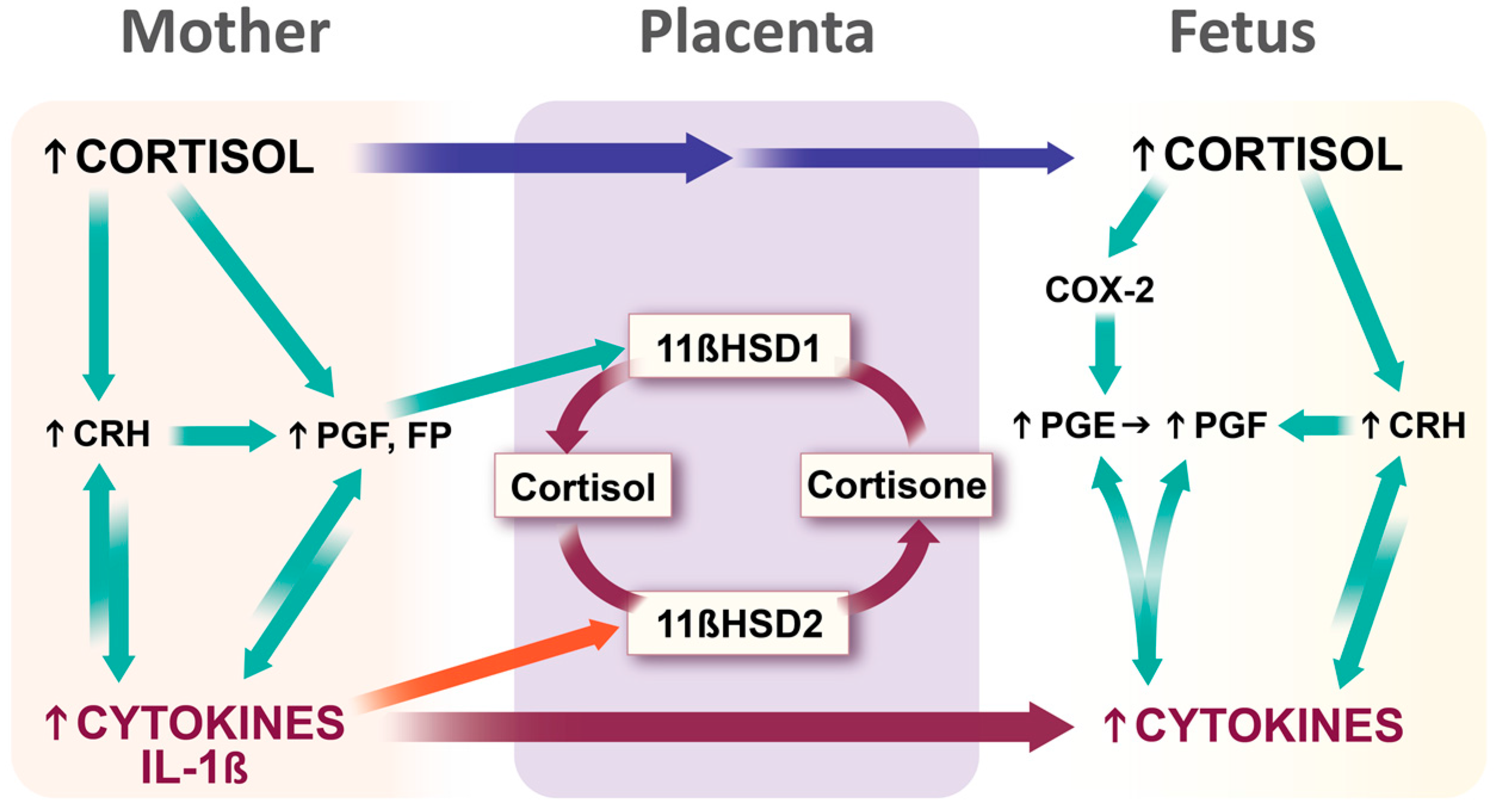
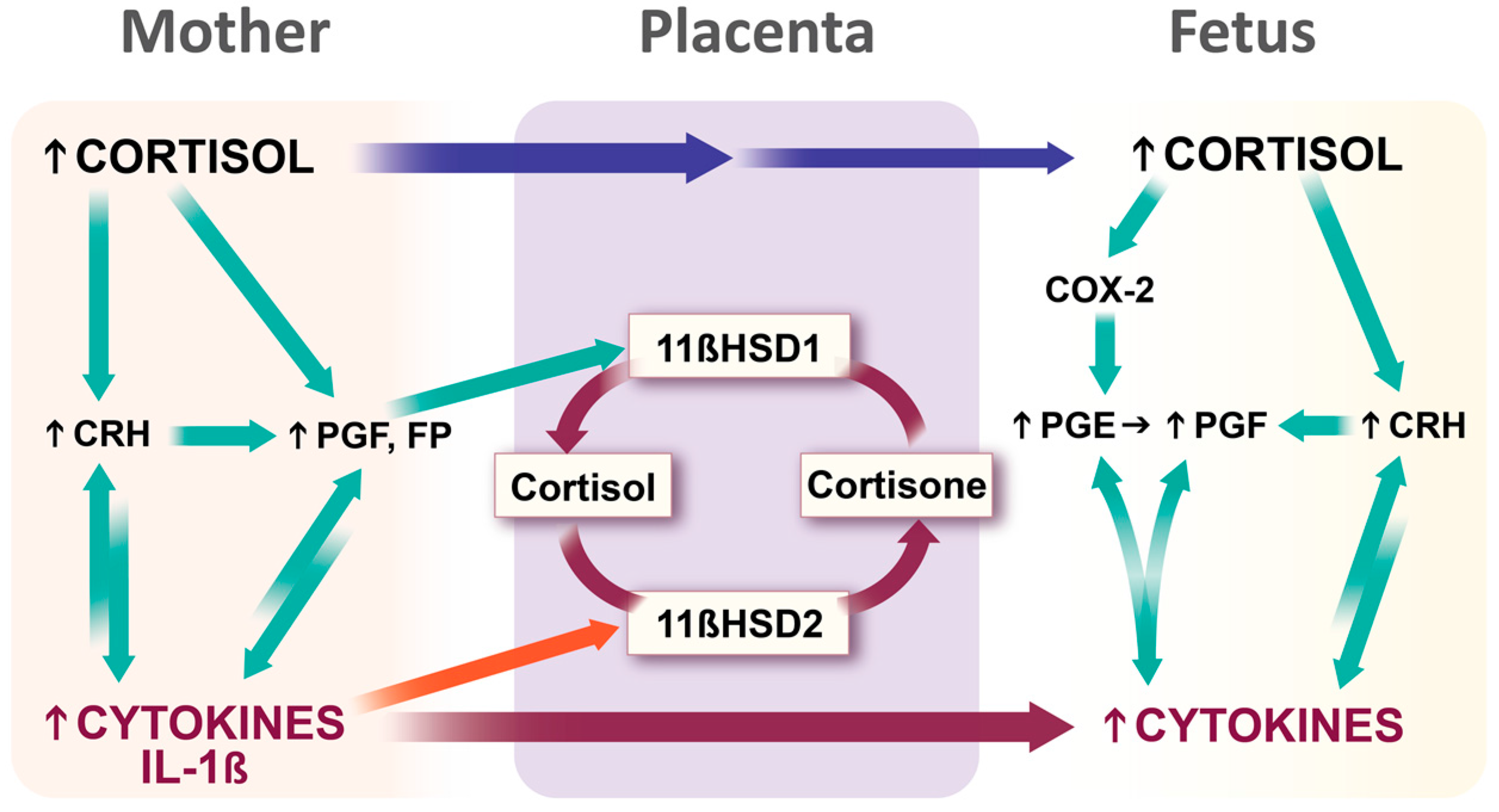
3.3.3. Stress Hormones and Inflammatory Mediators Intersect at the Placenta: Potential Transgenerational Transmission Mechanism
3.3.4. Epigenetic Associations
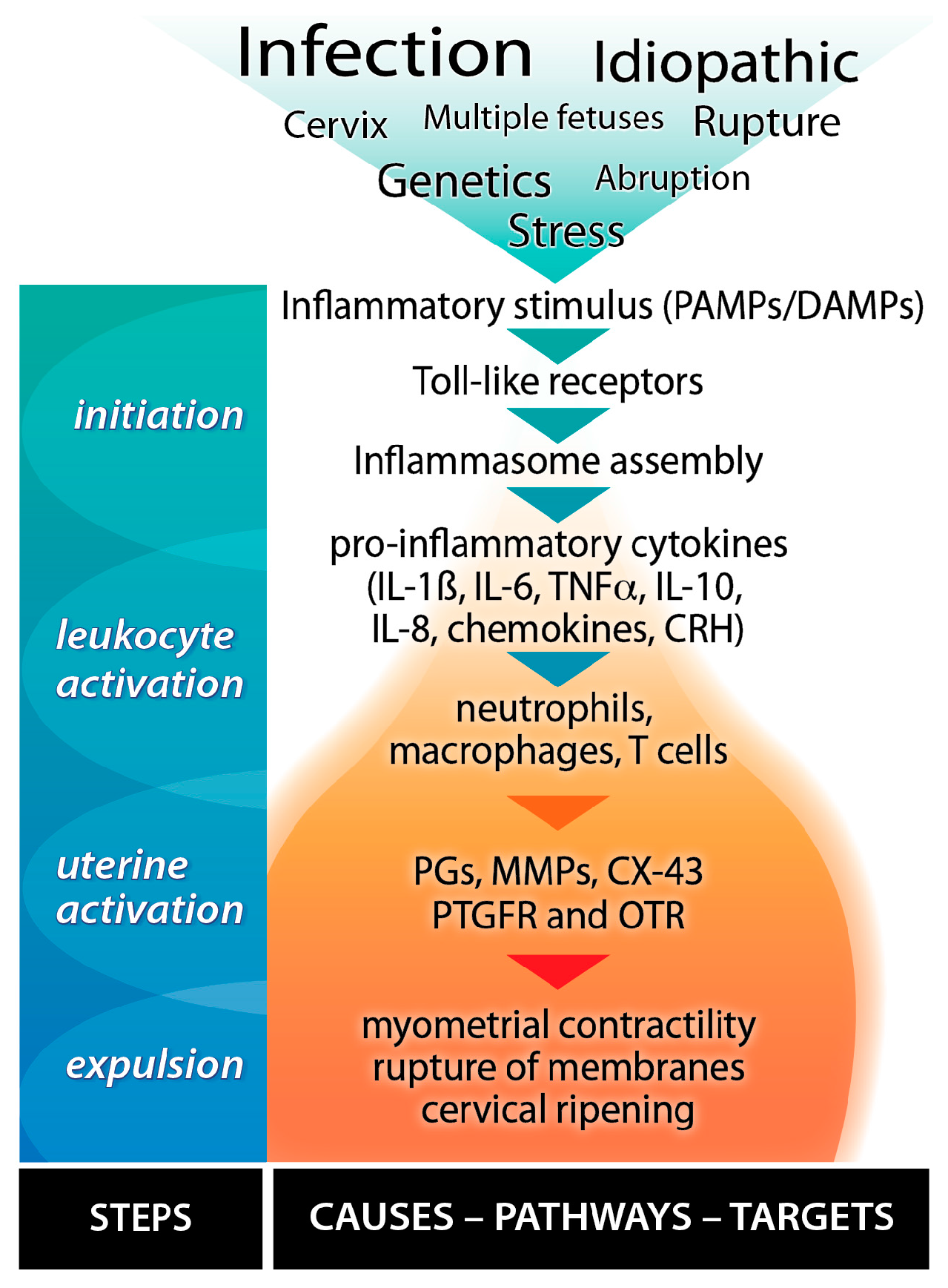
3.3.5. Stress, Inflammatory Mediators and Preterm Birth

4. Allostasis and Allostatic Load
4.1. Definitions
4.2. Types of Stressors
4.3. Our Conceptual Framework

4.4. Experimental Models of Allostatic Load and PTB
4.5. Using Allostatic Load to Predict PTB Risk
5. Developing an Allostatic Load Model for Predicting PTB Risk
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sterling, P.; Ever, J. Allostasis: A New Paradigm to Explain Arousal Pathology; Fisher, S.J.R., Ed.; John Wiley & Sons: New York, NY, USA, 1988. [Google Scholar]
- McEwen, B.S. Protective and damaging effects of stress mediators. N. Engl. J. Med. 1998, 338, 171–179. [Google Scholar] [PubMed]
- Christiaens, I.; Zaragoza, D.B.; Guilbert, L.; Robertson, S.A.; Mitchell, B.F.; Olson, D.M. Inflammatory processes in preterm and term parturition. J. Reprod. Immunol. 2008, 79, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, K.; Ritari, N.; Tommiska, V.; Salokorpi, T.; Lehtonen, L.; Tammela, O.; Pääkkönen, L.; Olsen, P.; Korkman, M.; Fellman, V. Neurodevelopmental outcome at 5 years of age of a national cohort of extremely low birth weight infants who were born in 1996–1997. Pediatrics 2005, 116, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.S.; Lydon, J.; Goulet, L.; Kahn, S.; Dahhou, M.; Platt, R.W.; Sharma, S.; Meaney, M.J.; Séguin, L. Maternal stress/distress, hormonal pathways and spontaneous preterm birth. Paediatr. Perinat. Epidemiol. 2013, 27, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Hobel, C.J. Stress and preterm birth. Clin. Obstet. Gynecol. 2004, 47, 856–880. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Tao, F.; Hao, J.; Sun, Y.; Jiang, X. Prenatal life events stress: Implications for preterm birth and infant birthweight. Am. J. Obstet. Gynecol. 2010, 203, 34.e1–34.e8. [Google Scholar] [CrossRef] [PubMed]
- Emanuel, I.; Leisenring, W.; Williams, M.A.; Kimpo, C.; Estee, S.; O’Brien, W.; Hale, C.B. The Washington State Intergenerational Study of Birth Outcomes: Methodology and some comparisons of maternal birthweight and infant birthweight and gestation in four ethnic groups. Paediatr. Perinat. Epidemiol. 1999, 13, 352–369. [Google Scholar] [CrossRef] [PubMed]
- Rich-Edwards, J.W.; Grizzard, T.A. Psychosocial stress and neuroendocrine mechanisms in preterm delivery. Am. J. Obstet. Gynecol. 2005, 192, S30–S35. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.E.; Anway, M.D.; Stanfield, J.; Skinner, M.K. Transgenerational epigenetic effects of the endocrine disruptor vinclozolin on pregnancies and female adult onset disease. Reproduction 2008, 135, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K.; Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Haque, M.; Nilsson, E.E. Ancestral dichlorodiphenyltrichloroethane (DDT) exposure promotes epigenetic transgenerational inheritance of obesity. BMC Med. 2013, 11, 228. [Google Scholar] [CrossRef] [PubMed]
- Veenendaal, M.V.; Painter, R.C.; de Rooij, S.R.; Bossuyt, P.M.; van der Post, J.A.; Gluckman, P.D.; Hanson, M.A.; Roseboom, T.J. Transgenerational effects of prenatal exposure to the 1944–45 Dutch famine. BJOG 2013, 120, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.E.; Riegle, G.D. Progesterone and luteinizing hormone secretion following stress-induced interruption of constant estrus in aged rats. J. Gerontol. 1985, 40, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Arck, P.C. Stress and pregnancy loss: Role of immune mediators, hormones and neurotransmitters. Am. J. Reprod. Immunol. 2001, 46, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Read, S.; Grundy, E. Allostatic load—A challenge to measure multisystem physiological regulation working paper. Available online: http://eprints.ncrm.ac.uk/2879/1/NCRM_workingpaper_0412.pdf (accessed on 24 August 2015).
- McEwen, B.S.; Seeman, T. Protective and damaging effects of mediators of stress. Elaborating and testing the concepts of allostasis and allostatic load. Ann. N. Y. Acad. Sci. 1999, 896, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, C.L.; Higham, J.P.; Heistermann, M.; Coe, C.L.; Prendergast, B.J.; Maestripieri, D. Immune function and HPA axis activity in free-ranging rhesus macaques. Physiol. Behav. 2011, 104, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.; Harville, E.; Theall, K.; Webber, L.; Chen, W.; Berenson, G. Neighborhood poverty, allostatic load, and birth outcomes in African American and white women: Findings from the Bogalusa Heart Study. Health Place 2013, 24, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Hux, V.J.; Catov, J.M.; Roberts, J.M. Allostatic load in women with a history of low birth weight infants: The national health and nutrition examination survey. J. Women’s Health 2014, 23, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Hux, V.J.; Roberts, J.M. A potential role for allostatic load in preeclampsia. Matern. Child Health J. 2014, 19, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Felitti, V.J.; Anda, R.F.; Nordenberg, D.; Williamson, D.F.; Spitz, A.M.; Edwards, V.; Koss, M.P.; Marks, J.S. Relationship of childhood abuse and household dysfunction to many of the leading causes of death in adults. The Adverse Childhood Experiences (ACE) Study. Am. J. Prev. Med. 1998, 14, 245–258. [Google Scholar] [CrossRef]
- Christiaens, I.; Hegadoren, K.; Olson, D.M. Adverse childhood experiences are associated with spontaneous preterm birth:A case-control study. BMC Med. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Robinson, A.M.; Zucchi, F.C.; Robbins, J.C.; Babenko, O.; Kovalchuk, O.; Kovalchuk, I.; Olson, D.M.; Metz, G.A.S. Ancestral exposure to stress epigenetically programs preterm birth risk and adverse maternal and newborn outcomes. BMC Med. 2014, 12, 121. [Google Scholar] [CrossRef] [PubMed]
- Juster, R.P.; McEwen, B.S.; Lupien, S.J. Allostatic load biomarkers of chronic stress and impact on health and cognition. Neurosci. Biobehav. Rev. 2010, 35, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.; Harville, E.; Theall, K.; Webber, L.; Chen, W.; Berenson, G. Preconception biomarkers of allostatic load and racial disparities in adverse birth outcomes: The Bogalusa Heart Study. Paediatr. Perinat. Epidemiol. 2013, 27, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.; Shenassa, E.D.; Mendola, P.; Wu, T.; Schoendorf, K. Allostatic load may not be associated with chronic stress in pregnant women, NHANES 1999–2006. Ann. Epidemiol. 2013, 23, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.F.; Fraser, A.M.; Hunter, C.Y.; Ward, R.H.; Varner, M.W. The risk of preterm birth across generations. Obstet. Gynecol. 1997, 90, 63–67. [Google Scholar] [CrossRef]
- Coussons-Read, M.E.; Lobel, M.; Carey, J.C.; Kreither, M.O.; D’Anna, K.; Argys, L.; Ross, R.G.; Brandt, C.; Cole, S. The occurrence of preterm delivery is linked to pregnancy-specific distress and elevated inflammatory markers across gestation. Brain Behav. Immun. 2012, 26, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Crews, D.; Gillette, R.; Scarpino, S.V.; Manikkam, M.; Savenkova, M.I.; Skinner, M.K. Epigenetic transgenerational inheritance of altered stress responses. Proc. Natl. Acad. Sci. USA 2012, 109, 9143–9148. [Google Scholar] [CrossRef] [PubMed]
- Zucchi, F.C.; Yao, Y.; Ward, I.D.; Ilnytskyy, Y.; Olson, D.M.; Benzies, K.; Kovalchuk, I.; Kovalchuk, O.; Metz, G.A.S. Maternal stress induces epigenetic signatures of psychiatric and neurological diseases in the offspring. PLoS ONE 2013, 8, e56967. [Google Scholar] [CrossRef] [PubMed]
- Kaati, G.; Bygren, L.O.; Edvinsson, S. Cardiovascular and diabetes mortality determined by nutrition during parents’ and grandparents’ slow growth period. Eur. J. Hum. Genet. 2002, 10, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Franklin, T.B.; Linder, N.; Russig, H.; Thony, B.; Mansuy, I.M. Influence of early stress on social abilities and serotonergic functions across generations in mice. PLoS ONE 2011, 6, e21842. [Google Scholar] [CrossRef] [PubMed]
- Weiss, I.C.; Franklin, T.B.; Vizi, S.; Mansuy, I.M. Inheritable effect of unpredictable maternal separation on behavioral responses in mice. Front. Behav. Neurosci. 2011, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Ward, I.D.; Zucchi, F.C.; Robbins, J.C.; Falkenberg, E.A.; Olson, D.M.; Benzies, K.; Metz, G.A. Transgenerational programming of maternal behaviour by prenatal stress. BMC Pregnancy Childbirth 2013, 13, S9. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, J.; Parker, B.; Soeken, K.; Bullock, L. Assessing for abuse during pregnancy. Severity and frequency of injuries and associated entry into prenatal care. JAMA 1992, 267, 3176–3178. [Google Scholar] [CrossRef] [PubMed]
- Paris, J.J.; Frye, C.A. Gestational exposure to variable stressors produces decrements in cognitive and neural development of juvenile male and female rats. Curr. Top. Med. Chem. 2011, 11, 1706–1713. [Google Scholar] [PubMed]
- Herbison, A.E. Physiological roles for the neurosteroid allopregnanolone in the modulation of brain function during pregnancy and parturition. Prog. Brain Res. 2001, 133, 39–47. [Google Scholar] [PubMed]
- Chin, V.S.; van Skike, C.E.; Berry, R.B.; Kirk, R.E.; Diaz-Granados, J.; Matthews, D.B. Effect of acute ethanol and acute allopregnanolone on spatial memory in adolescent and adult rats. Alcohol 2011, 45, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Paris, J.J.; Frye, C.A. Juvenile offspring of rats exposed to restraint stress in late gestation have impaired cognitive performance and dysregulated progestogen formation. Stress 2011, 14, 23–32. [Google Scholar] [PubMed]
- McCreary, J.K.; Truica, L.S.; Friesen, B.; Yao, Y.; Olson, D.M.; Cross, A.R.; Metz, G.A.S. Recurrent prenatal stress exacerbates stress sensitivity and vulnerability to mental illness across generations. 2015; (submitted). [Google Scholar]
- Knudson, A.G., Jr. Mutation and cancer: Statistical study of retinoblastoma. Proc. Natl. Acad. Sci. USA 1971, 68, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.M.; (University of Alberta); McCreary, J.K.; (University of Lethbridge); Verstraeten, B.S.E.; (University of Alberta); Metz, G.A.S.; (University of Lethbridge). Personal communication, 2015.
- Yang, K.; Jones, S.A.; Challis, J.R. Changes in glucocorticoid receptor number in the hypothalamus and pituitary of the sheep fetus with gestational age and after adrenocorticotropin treatment. Endocrinology 1990, 126, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Charil, A.; Laplante, D.P.; Vaillancourt, C.; King, S. Prenatal stress and brain development. Brain Res. Rev. 2010, 65, 56–79. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Tsai, S.W.; Nguyen, T.T.; Plevyak, M.; Padbury, J.F.; Rubin, L.P. Inhibition of placental 11β-hydroxysteroid dehydrogenase type 2 by catecholamines via α-adrenergic signaling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R1966–R1974. [Google Scholar] [PubMed]
- Newnham, J.P. Is prenatal glucocorticoid administration another origin of adult disease? Clin. Exp. Pharmacol. Physiol. 2001, 28, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Seckl, J.R. Glucocorticoids, developmental “programming” and the risk of affective dysfunction. Prog. Brain Res. 2008, 167, 17–34. [Google Scholar] [PubMed]
- Kossintseva, I.; Wong, S.; Johnstone, E.; Guilbert, L.; Olson, D.M.; Mitchell, B.F. Proinflammatory cytokines inhibit human placental 11β-hydroxysteroid dehydrogenase type 2 activity through Ca2+ and cAMP pathways. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E282–E288. [Google Scholar] [CrossRef] [PubMed]
- Potestio, F.A.; Zakar, T.; Olson, D.M. Glucocorticoids stimulate prostaglandin synthesis in human amnion cells by a receptor-mediated mechanism. J. Clin. Endocrinol. Metab. 1988, 67, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Alfaidy, N.; Xiong, Z.G.; Myatt, L.; Lye, S.J.; MacDonald, J.F.; Challis, J.R. Prostaglandin F2α potentiates cortisol production by stimulating 11β-hydroxysteroid dehydrogenase 1: A novel feedback loop that may contribute to human labor. J. Clin. Endocrinol. Metab. 2001, 86, 5585–5592. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.; Seckl, J. Glucocorticoids, prenatal stress and the programming of disease. Horm. Behav. 2011, 59, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Migicovsky, Z.; Kovalchuk, I. Epigenetic memory in mammals. Front. Genet. 2011, 2, 28. [Google Scholar] [CrossRef] [PubMed]
- Zucchi, F.C.; Yao, Y.; Metz, G.A. The secret language of destiny: Stress imprinting and transgenerational origins of disease. Front. Genet. 2012, 3, 96. [Google Scholar] [CrossRef] [PubMed]
- Babenko, O.; Golubov, A.; Ilnytskyy, Y.; Kovalchuk, I.; Metz, G.A. Genomic and epigenomic responses to chronic stress involve miRNA-mediated programming. PLoS ONE 2012, 7, e29441. [Google Scholar] [CrossRef] [PubMed]
- Babenko, O.; Kovalchuk, I.; Metz, G.A. Stress-induced perinatal and transgenerational epigenetic programming of brain development and mental health. Neurosci. Biobehav. Rev. 2015, 48, 70–91. [Google Scholar] [CrossRef] [PubMed]
- Challis, J.R.; Lockwood, C.J.; Myatt, L.; Norman, J.E.; Strauss, J.F., 3rd; Petraglia, F. Inflammation and pregnancy. Reprod. Sci. 2009, 16, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Dey, S.K.; Fisher, S.J. Preterm labor: One syndrome, many causes. Science 2014, 345, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Breen, K.; Brown, A.; Burd, I.; Chai, J.; Friedman, A.; Elovitz, M.A. TLR-4-dependent and -independent mechanisms of fetal brain injury in the setting of preterm birth. Reprod. Sci. 2012, 19, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Hoang, M.; Potter, J.A.; Gysler, S.M.; Han, C.S.; Guller, S.; Norwitz, E.R.; Abrahams, V.M. Human fetal membranes generate distinct cytokine profiles in response to bacterial Toll-like receptor and nod-like receptor agonists. Biol. Reprod. 2014, 90, 39. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Campo, P.; Dominguez, F.; Coloma, J.; Meseguer, M.; Remohi, J.; Pellicer, A.; Simón, C. Hormonal and embryonic regulation of chemokines IL-8, MCP-1 and RANTES in the human endometrium during the window of implantation. Mol. Hum. Reprod. 2002, 8, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Van Mourik, M.S.; Macklon, N.S.; Heijnen, C.J. Embryonic implantation: Cytokines, adhesion molecules, and immune cells in establishing an implantation environment. J. Leukoc. Biol. 2009, 85, 4–19. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Stress, adaptation, and disease. Allostasis and allostatic load. Ann. N. Y. Acad. Sci. 1998, 840, 33–44. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef]
- Shannon, M.; King, T.L.; Kennedy, H.P. Allostasis: A theoretical framework for understanding and evaluating perinatal health outcomes. J. Obstet. Gynecol. Neonatal Nurs. 2007, 36, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Elenkov, I.J.; Webster, E.L.; Torpy, D.J.; Chrousos, G.P. Stress, corticotropin-releasing hormone, glucocorticoids, and the immune/inflammatory response: Acute and chronic effects. Ann. N. Y. Acad. Sci. 1999, 876, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Djuric, Z.; Bird, C.E.; Furumoto-Dawson, A.; Rauscher, G.H.; Ruffin, M.T.T.; Stowe, R.P.; Tucker, K.L.; Masi, C.M. Biomarkers of Psychological Stress in Health Disparities Research. Open Biomark. J. 2008, 1, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Daskalakis, N.P.; Bagot, R.C.; Parker, K.J.; Vinkers, C.H.; de Kloet, E.R. The three-hit concept of vulnerability and resilience: Toward understanding adaptation to early-life adversity outcome. Psychoneuroendocrinology 2013, 38, 1858–1873. [Google Scholar] [CrossRef] [PubMed]
- Saban, K.L.; Mathews, H.L.; DeVon, H.A.; Janusek, L.W. Epigenetics and social context: Implications for disparity in cardiovascular disease. Aging Dis. 2014, 5, 346–355. [Google Scholar] [PubMed]
- Li, J.; Zhang, Y.; Li, D.; Liu, Y.; Chu, D.; Jiang, X. Small non-coding RNAs transfer through mammalian placenta and directly regulate fetal gene expression. Protein Cell 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Misra, D.P.; Straughen, J.K.; Slaughter-Acey, J.C. Allostatic load and health: Can perinatal epidemiology lead the way forward? Paediatr. Perinat. Epidemiol. 2013, 27, 507–508. [Google Scholar] [CrossRef] [PubMed]
- Nordling, C.O. A new theory on cancer-inducing mechanism. Br. J. Cancer 1953, 7, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Spagnoli, C.; Cilio, M.R.; Pavlidis, E.; Pisani, F. Symptomatic neonatal seizures followed by febrile status epilepticus: The two-hit hypothesis for the subsequent development of epilepsy. J. Child Neurol. 2015, 30, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.J.; Monks, S.; Murphy, K.C.; Kirov, G.; O'Donovan, M.C.; Owen, M.J. Schizophrenia two-hit hypothesis in velo-cardio facial syndrome. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2013, 162B, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Racicot, K.; Kwon, J.Y.; Aldo, P.; Silasi, M.; Mor, G. Understanding the complexity of the immune system during pregnancy. Am. J. Reprod. Immunol. 2014, 72, 107–116. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Interacting mediators of allostasis and allostatic load: Towards an understanding of resilience in aging. Metabolism 2003, 52, 10–16. [Google Scholar] [CrossRef]
- Seeman, T.E.; Singer, B.H.; Rowe, J.W.; Horwitz, R.I.; McEwen, B.S. Price of adaptation-allostatic load and its health consequences. MacArthur studies of successful aging. Arch. Intern. Med. 1997, 157, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.C.; Halfon, N. Racial and ethnic disparities in birth outcomes: A life-course perspective. Matern. Child Health J. 2003, 7, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Latendresse, G. The interaction between chronic stress and pregnancy: Preterm birth from a biobehavioral perspective. J. Midwifery Women’s Health 2009, 54, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Power, M.L.; Schulkin, J. Maternal obesity, metabolic disease, and allostatic load. Physiol. Behav. 2012, 106, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.E.; Harville, E.W. Allostatic load and birth outcomes among white and black women in New Orleans. Matern. Child Health J. 2013, 17, 1025102–1025109. [Google Scholar] [CrossRef] [PubMed]
- Merkin, S.S.; Karlamangla, A.; Roux, A.V.; Shrager, S.; Seeman, T.E. Life course socioeconomic status and longitudinal accumulation of allostatic load in adulthood: Multi-ethnic study of atherosclerosis. Am. J. Public Health 2014, 104, e48–e55. [Google Scholar] [CrossRef] [PubMed]
- Koopman, C.; Angell, K.; Turner-Cobb, J.M.; Kreshka, M.A.; Donnelly, P.; McCoy, R.; Turkseven, A.; Graddy, K.; Giese-Davis, J.; Spiegel, D. Distress, coping, and social support among rural women recently diagnosed with primary breast cancer. Breast J. 2001, 7, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Birkhoff, G. Lattice Theory, 4th ed.; American Mathematical Society Colloquium Publications: Providence, RI, USA, 1979. [Google Scholar]
- Wille, R. Line diagrams of hierarchical concept systems. Int. Classif. 1984, 11, 77–86. [Google Scholar]
- Walker, L.O.; Avant, K.C. Concept analysis. In Strategies for Theory Construction in Nursing, 5th ed.; Prentice Hall: Upper Sadle River, NJ, USA, 2011; pp. 157–179. [Google Scholar]
- Erickson, Z.T.; Falkenberg, E.A.; Metz, G.A. Lifespan psychomotor behaviour profiles of multigenerational prenatal stress and artificial food dye effects in rats. PLoS ONE 2014, 9, e92132. [Google Scholar] [CrossRef] [PubMed]
- Karagiannides, I.; Golovatscka, V.; Bakirtzi, K.; Sideri, A.; Salas, M.; Stavrakis, D.; Polytarchou, C.; Iliopoulos, D.; Pothoulakis, C.; Bradesi, S. Chronic unpredictable stress regulates visceral adipocyte-mediated glucose metabolism and inflammatory circuits in male rats. Physiol. Rep. 2014, 2, e00284. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olson, D.M.; Severson, E.M.; Verstraeten, B.S.E.; Ng, J.W.Y.; McCreary, J.K.; Metz, G.A.S. Allostatic Load and Preterm Birth. Int. J. Mol. Sci. 2015, 16, 29856-29874. https://doi.org/10.3390/ijms161226209
Olson DM, Severson EM, Verstraeten BSE, Ng JWY, McCreary JK, Metz GAS. Allostatic Load and Preterm Birth. International Journal of Molecular Sciences. 2015; 16(12):29856-29874. https://doi.org/10.3390/ijms161226209
Chicago/Turabian StyleOlson, David M., Emily M. Severson, Barbara S. E. Verstraeten, Jane W. Y. Ng, J. Keiko McCreary, and Gerlinde A. S. Metz. 2015. "Allostatic Load and Preterm Birth" International Journal of Molecular Sciences 16, no. 12: 29856-29874. https://doi.org/10.3390/ijms161226209
APA StyleOlson, D. M., Severson, E. M., Verstraeten, B. S. E., Ng, J. W. Y., McCreary, J. K., & Metz, G. A. S. (2015). Allostatic Load and Preterm Birth. International Journal of Molecular Sciences, 16(12), 29856-29874. https://doi.org/10.3390/ijms161226209