
Mutations of Glucose-6-Phosphate Dehydrogenase Durham, Santa-Maria and A+ Variants Are Associated with Loss Functional and Structural Stability of the Protein

 ,
,  ,
, 
Abstract
:
1. Introduction
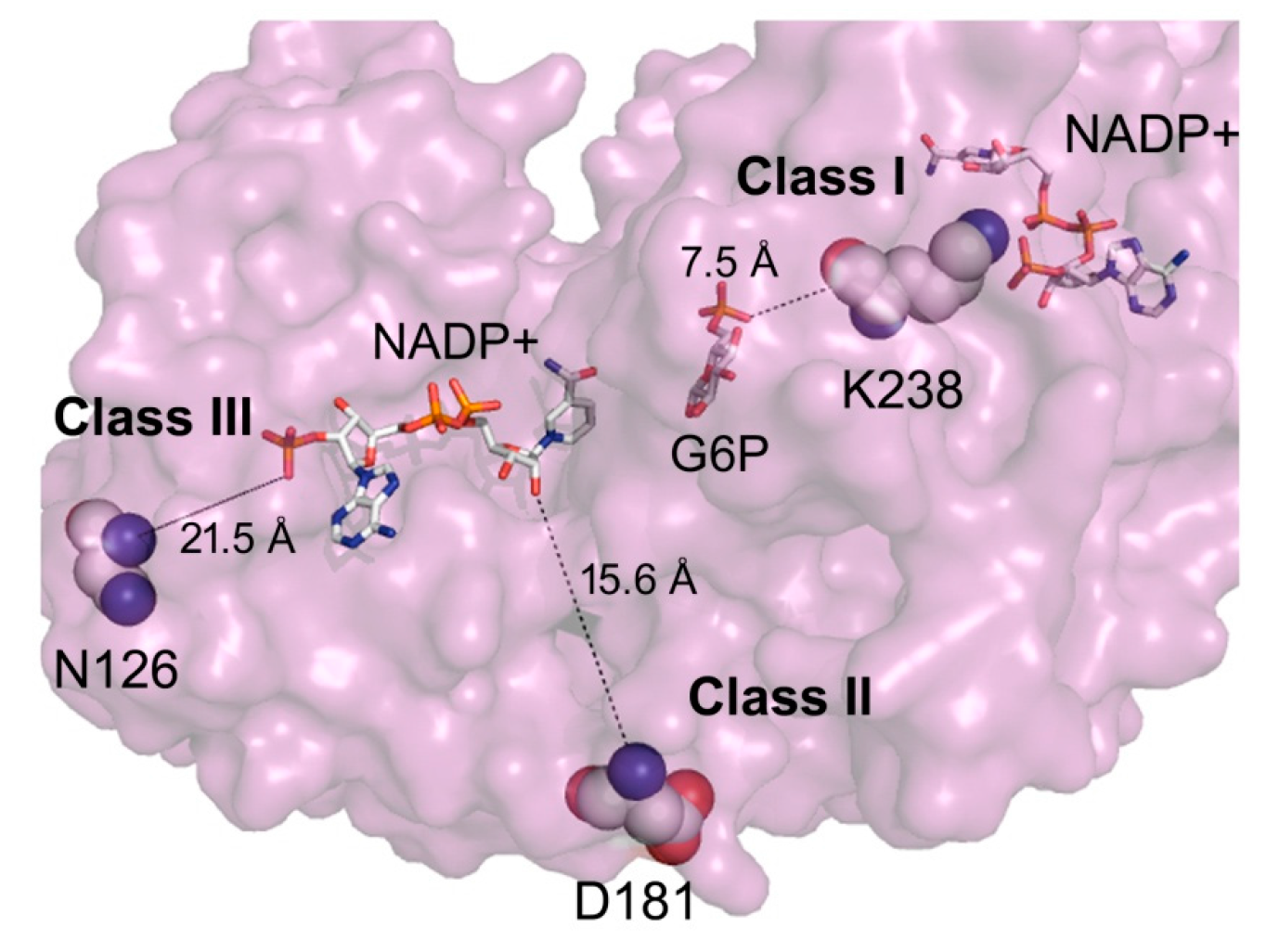
2. Results
2.1. Expression and Purification of Recombinant Human Glucose-6-Phosphate Dehydrogenase (G6PD) Enzymes
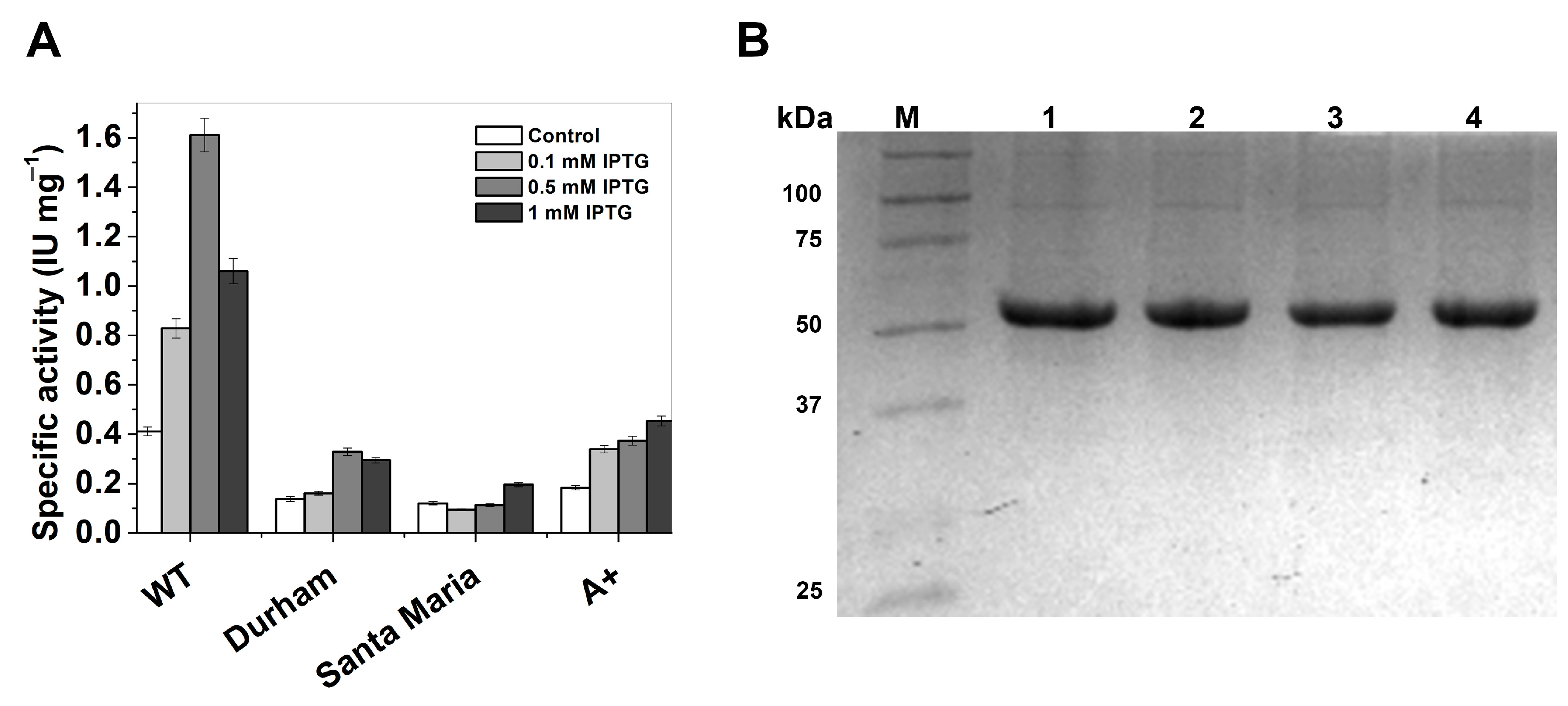

| G6PD | Total Protein (mg) | Specific Activity (IU·mg−1) | Total Activity (IU) | Yield (%) |
|---|---|---|---|---|
| WT | 1.95 | 224 | 436 | 61 |
| Durham | 1.15 | 71 | 81 | 21 |
| Santa-Maria | 1.41 | 71 | 100 | 32 |
| A+ | 1.81 | 114 | 373 | 43 |
2.2. Functional Characterization of Recombinant Human G6PD Enzymes
2.2.1. Steady-State Kinetics of G6PD Enzymes
| Kinetic Constants | WT-G6PD | Variants | ||
|---|---|---|---|---|
| Durham | Santa-Maria | A+ | ||
| Km G6P (µM) | 38.49 | 24.77 | 15.35 | 56.44 |
| Km NADP+ (µM) | 6.16 | 6.96 | 9.06 | 12.97 |
| kcat (s−1) | 233 | 71 | 71 | 114 |
| kcat/Km G6P (s−1·M−1) | 6.05 × 106 | 2.85 × 106 | 4.62 × 106 | 2.02 × 106 |
| kcat/Km NADP+ (s−1·M−1) | 37.82 × 106 | 10.20 × 106 | 7.83 × 106 | 8.78 × 106 |
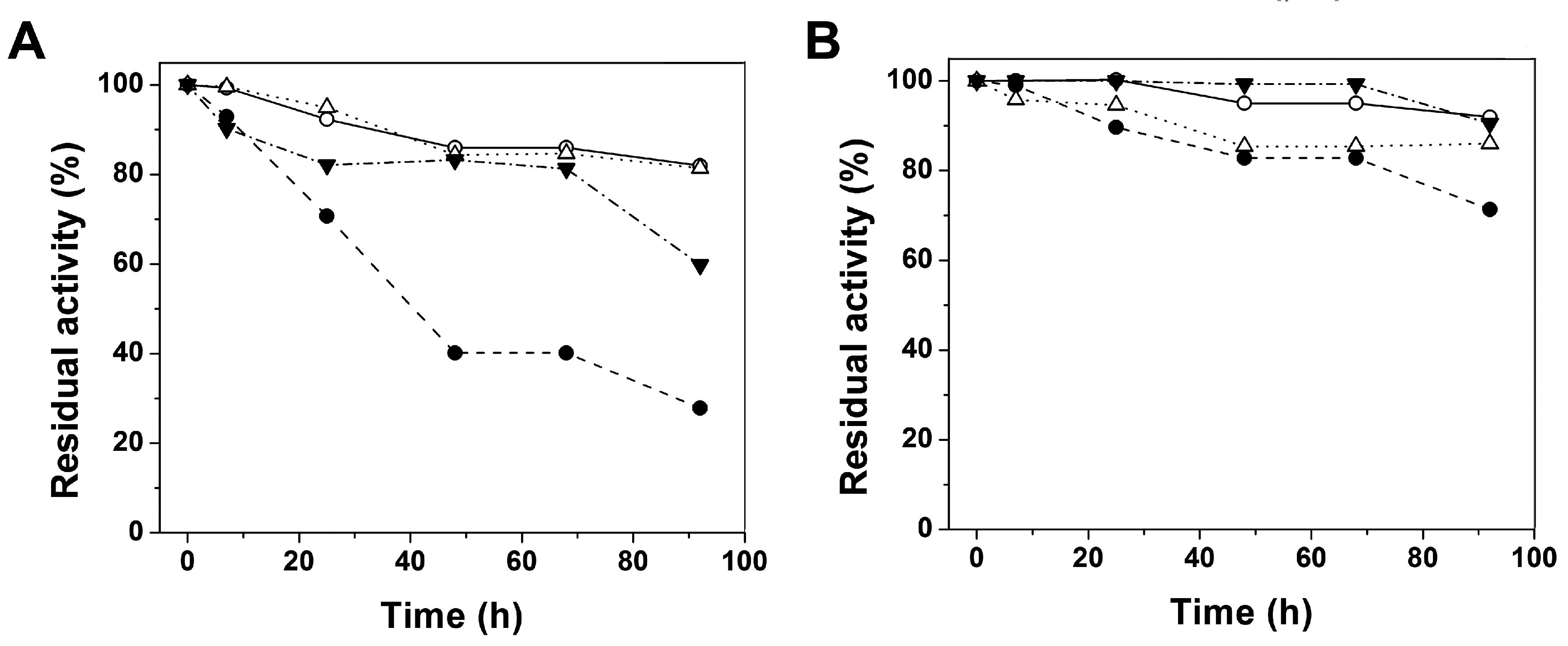
2.2.2. Stability Characterization of Recombinant Human G6PD Enzymes
| G6PD | T50 (°C) | Tm (°C) |
|---|---|---|
| WT | 48.8 ± 0.2 | 54.8 ± 0.3 |
| Durham | 40.3 ± 0.3 | 49.9 ± 0.4 |
| Santa-Maria | 45.5 ± 0.4 | 54.8 ± 0.3 |
| A+ | 45.6 ± 0.3 | 55.8 ± 0.4 |
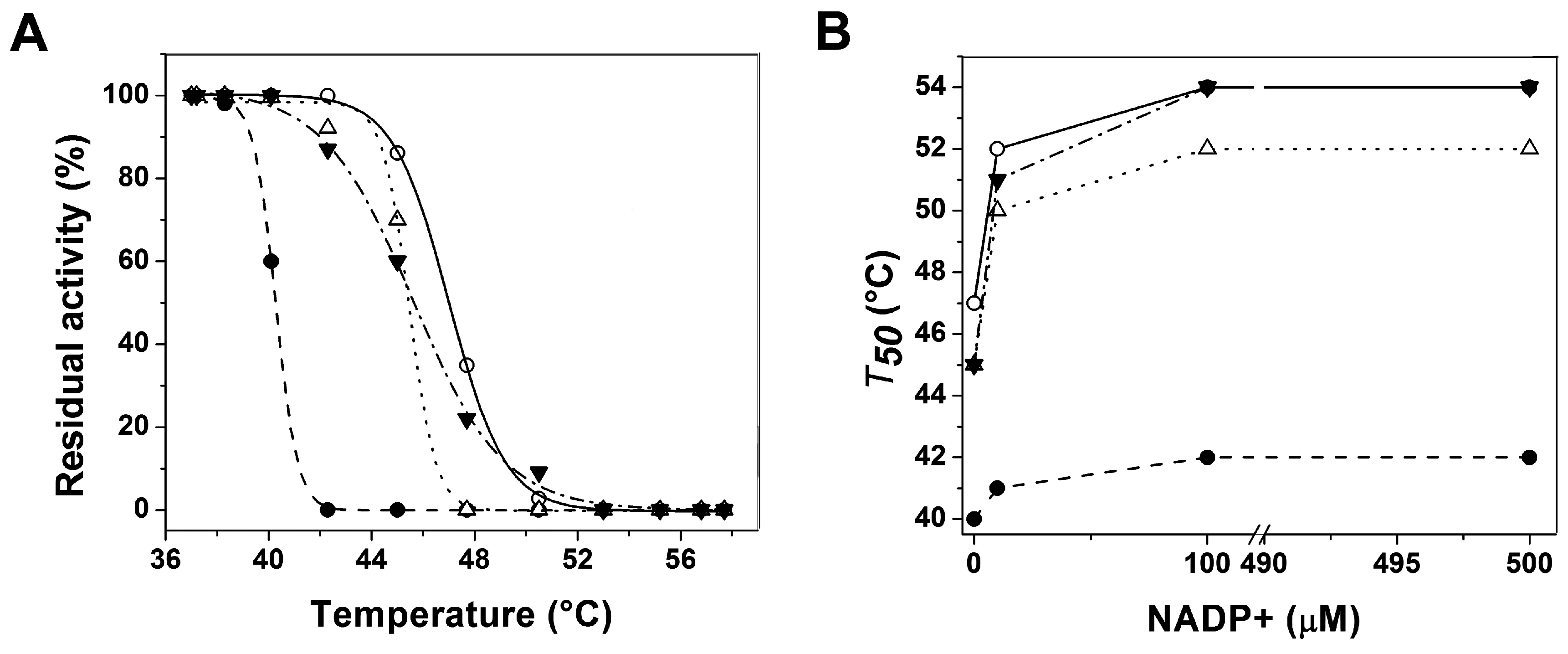
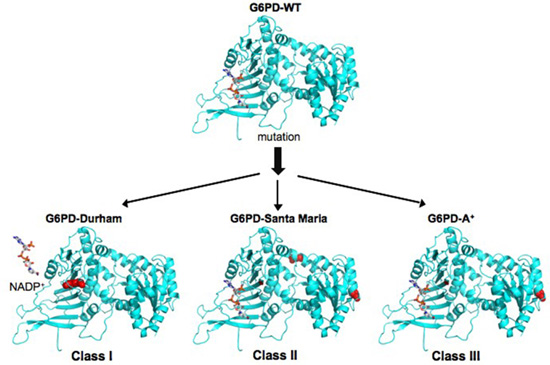
2.3. Structural Characterization of Recombinant Human G6PD Enzymes
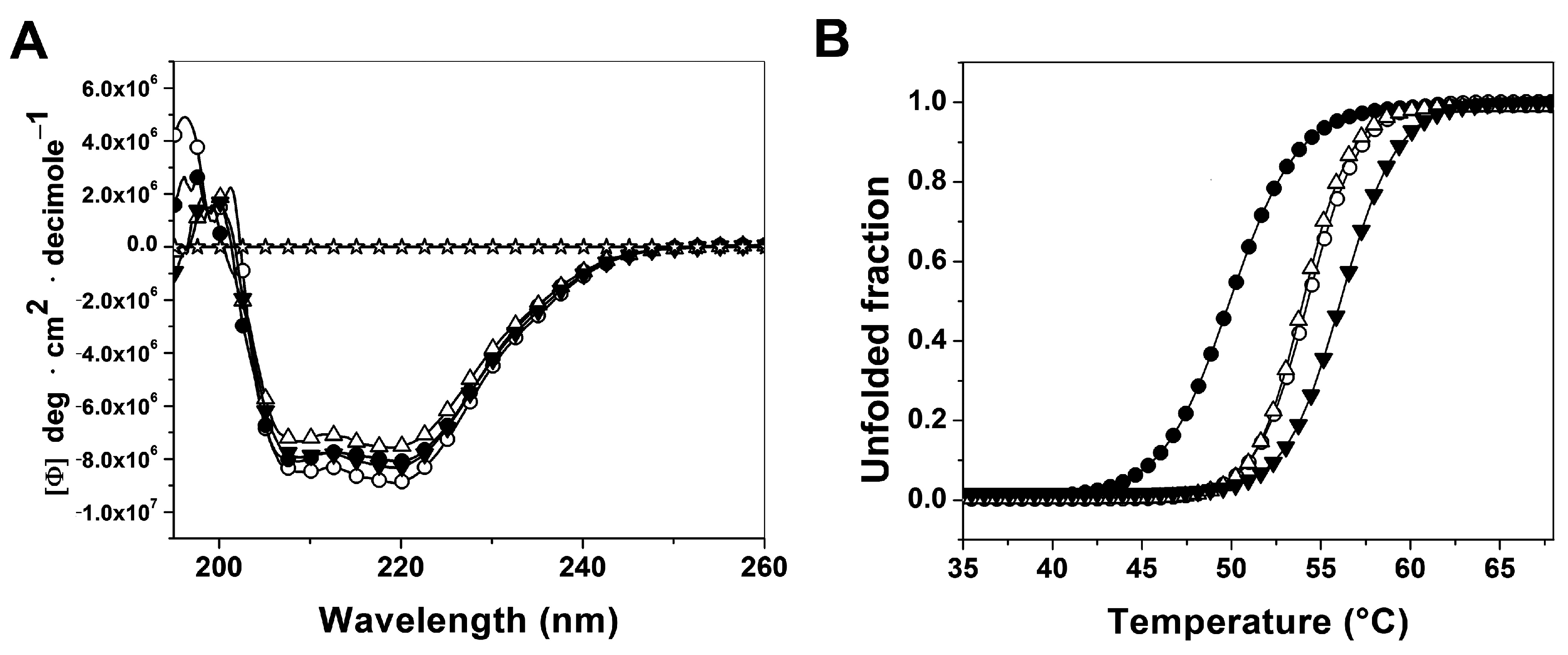
2.3.1. Analysis of Secondary Structure and Thermal Stability of Recombinant Human G6PD Enzymes

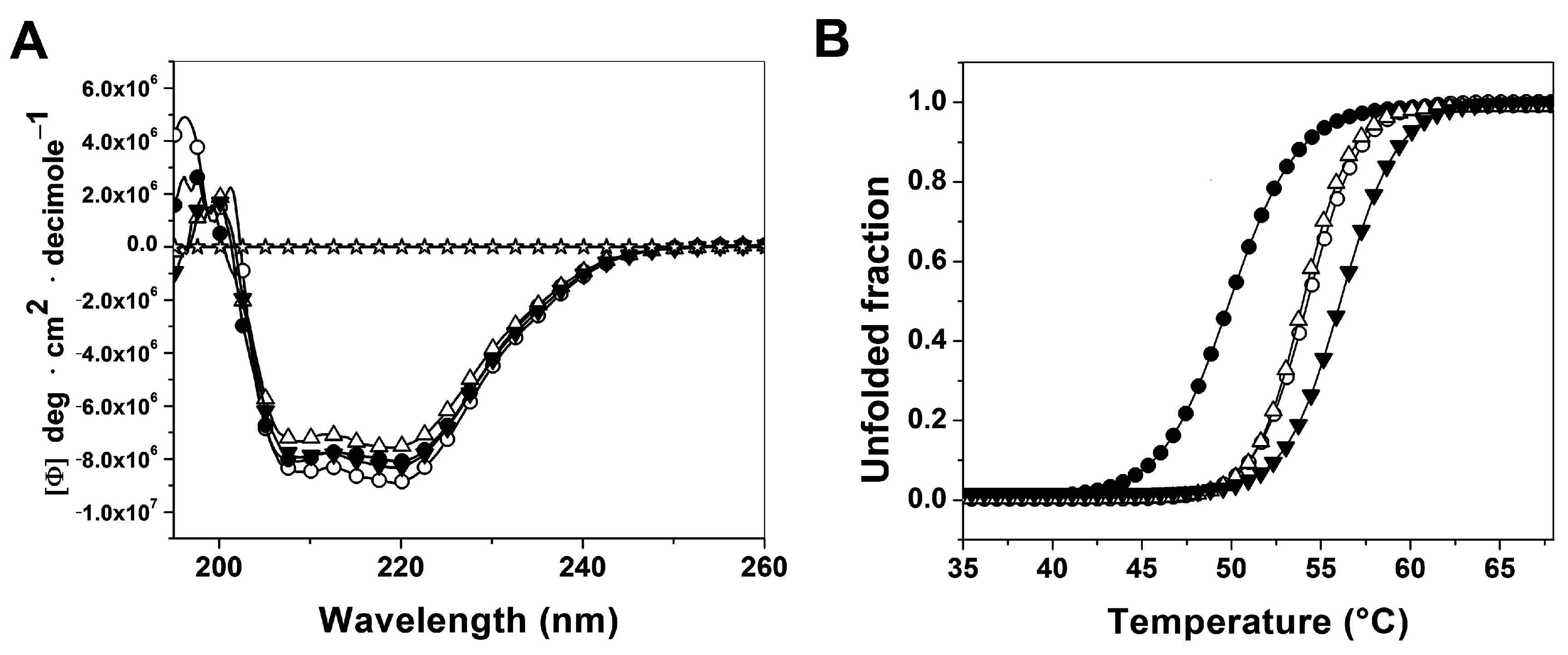
2.3.2. Analysis of Conformational Changes of Recombinant Human G6PD Enzymes
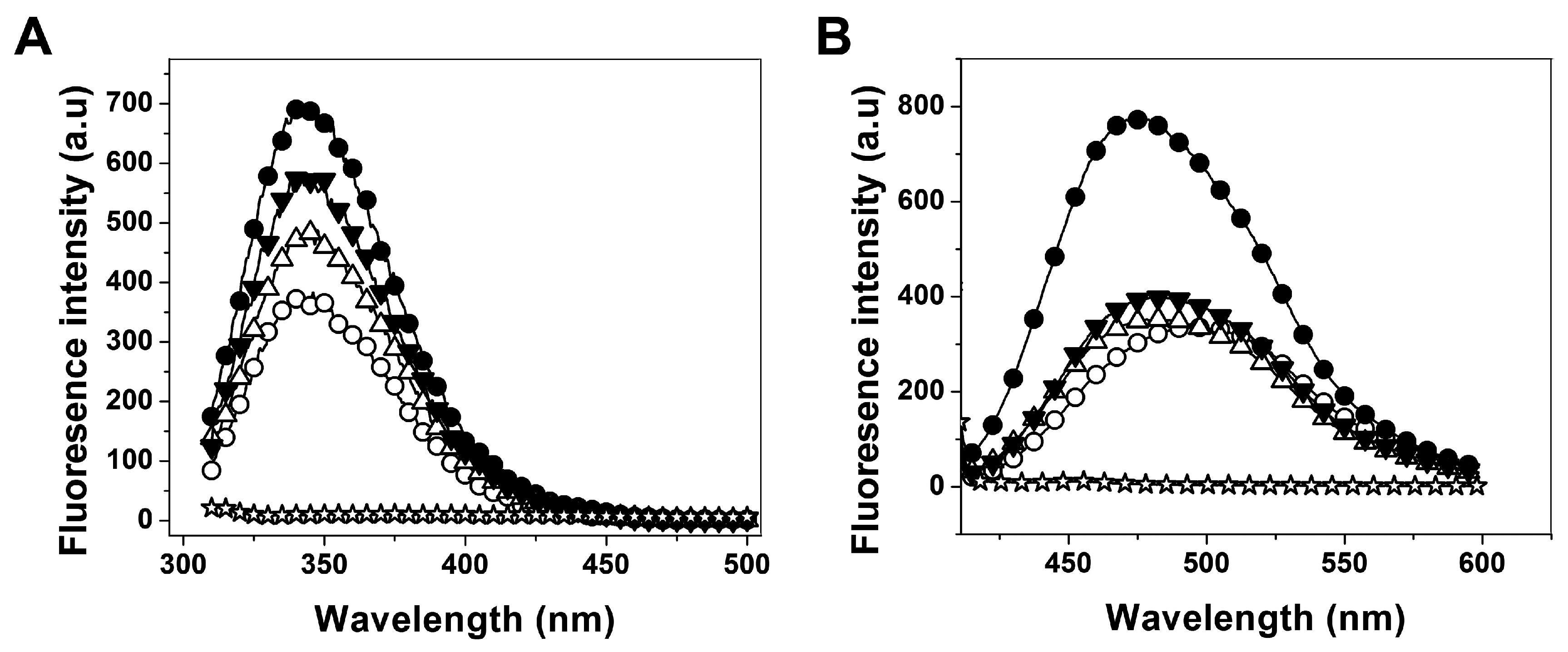
3. Discussion
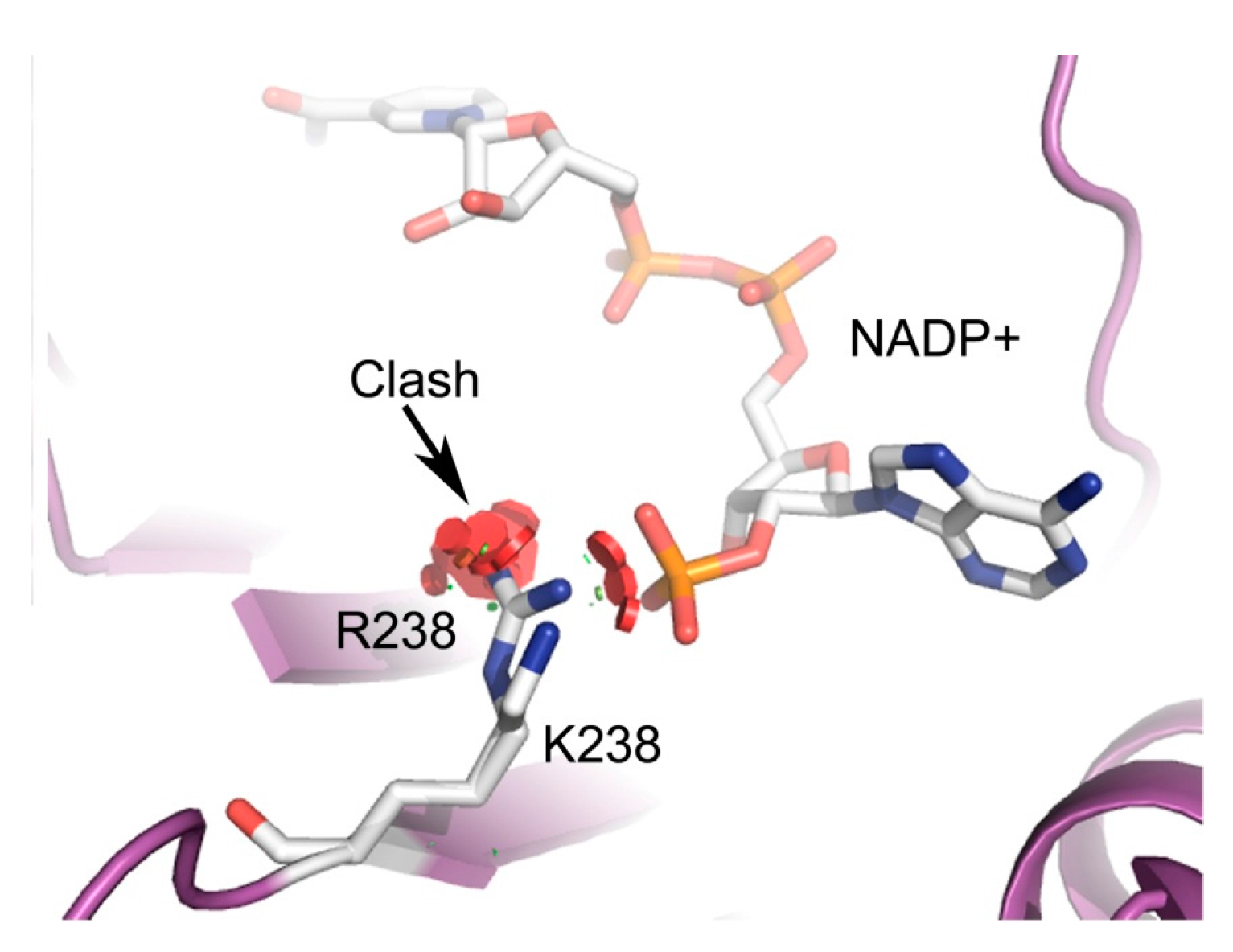
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4. Materials and Methods
4.1. Construction and Cloning of WT-G6PD and Durham, Santa-Maria and A+ Variants
| Primer Name | Primer Sequence |
|---|---|
| Durham K238R | 5′-CTCACCTTCAGGGACCC-3´ |
| 5´-GGGTCCCTGAAGGTGAG-3´ | |
| Santa-Maria N126D | 5´-AGCCACATGGATGCCCTCCAC-3´ |
| 5´-GTGGAGGGCATCCATGTGGCT-3´ | |
| Santa-Maria D181V | 5´-AGAGCTCTGTCGGCTGTCCA-3´ |
| 5´-TGGACAGCCGGACAGAGCTCT-3´ | |
| A+ forward N126D | 5´-AGCCACATGGATGCCCTCCAC-3´ |
| 5´-GTGGAGGGCATCCATGTGGCT-3´ |
4.2. Expression and Purification of Recombinant Human G6PD Enzymes
4.3. Functional Characterization of Recombinant Human G6PD Enzymes
4.3.1. Steady-State Kinetic Experiments
4.3.2. Thermal Inactivation Assays
4.4. Structural Characterization of Recombinant Human G6PD Enzymes
4.4.1. Analysis of Secondary Structure and Thermal Stability
4.4.2. Analysis of Conformational Changes of Recombinant Human G6PD Enzymes
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vulliamy, T.; Luzzatto, L.; Hirono, A.; Beutler, E. Hematologically important mutations: Glucose-6-phosphate dehydrogenase. Blood Cells Mol. Dis. 1997, 23, 302–313. [Google Scholar] [CrossRef]
- Pai, G.S.; Sprenkle, J.A.; Do, T.T.; Mareni, C.E.; Migeon, B.R. Localization of loci for hypoxanthine phosphoribosyltransferase and glucose-6-phosphate dehydrogenase and biochemical evidence of nonrandom X chromosome expression from studies of a human X-autosome translocation. Proc. Natl. Acad. Sci. USA 1980, 77, 2810–2813. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.J.; Bautista, J.M.; Gilsanz, F. G6PD deficiency: The genotype-phenotype association. Blood Rev. 2007, 5, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Cappellini, D.; Fiorelli, G. Glucose-6-phosphate dehydro-genase deficiency. Lancet 2008, 9606, 64–74. [Google Scholar] [CrossRef]
- Minucci, A.; Moradkhani, K.; Hwang, M.; Zuppi, C.; Giardina, B.; Capoluongo, P. Glucose-6-phosphate dehydrogenase (G6PD) mutations database: Review of the “old” and update of the new mutations. Blood Cells Mol. Dis. 2012, 48, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.T.; Chan, T.F.; Lam, V.; Engel, P. What is the role of the second “structural” NADP-binding site in human glucose-6-phosphate dehydrogenase? Protein Sci. 2008, 17, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Manzo, S.; Terrón-Hernández, J.; de la Mora-de la Mora, I.; González-Valdez, A.; Marcial-Quino, J.; García-Torres, I.; Vanoye-Carlo, A.; López-Velázquez, G.; Hernández-Alcantara, G.; Oria-Hernández, J.; et al. The stability of G6PD is affected by mutations with different clinical phenotypes. Int. J. Mol. Sci. 2014, 15, 21179–21201. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.; Ware, R.; Forman, R.; Westwood, B.; Beutler, E. Glucose-6phosphate dehydrogenase Durham: A de novo mutation associated chronic hemolytic anemia. J. Pediatr. 1997, 131, 284–287. [Google Scholar] [CrossRef]
- Vaca, G.; Arámbula, E.; Monsalvo, A.; Medina, C.; Nuñez, C.; Sandoval, L.; López-Guido, B. Glucose-6-phosphate dehydrogenase (G6PD) mutations in Mexico: Four new G6PD variants. Blood Cells Mol. Dis. 2003, 31, 112–120. [Google Scholar] [CrossRef]
- Saenz, G.F.; Chaves, M.; Berrantes, A.; Elizondo, J.; Montero, A.G.; Yoshida, A. A glucose-6-phosphate dehydrogenase variant, Gd(-) Santamaria found in Costa Rica. Acta Haematol. 1984, 72, 37–40. [Google Scholar] [PubMed]
- Beutler, E.; Kuhl, W.; German, F.; Sdenz, R.; Rodriguez, W.R. Mutation analysis of glucose-6-phosphate dehydrogenase (G6PD) variants in Costa Rica. Hum. Genet. 1991, 87, 462–464. [Google Scholar] [CrossRef] [PubMed]
- Citadella, R.; Civitelli, D.; Manna, I.; Azzia, N.; di Cataldos, A.; Schiliroa, G.; Brancatis, C. Genetic heterogeneity of glucose-6-phosphate dehydrogenase deficiency in south-east Sicily. Ann. Hum. Genet. 1997, 61, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Vaca, G.; Arambula, E.; Esparza, A. Molecular heterogeneity of glucose-6-phosphate dehydrogenase deficiency in Mexico: Overall results of a 7-year project. Blood Cells Mol. Dis. 2002, 28, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E.; Vulliamy, T.J. Hematologically important mutations: Glucose-6-phosphate dehydrogenase. Blood Cells Mol. Dis. 2002, 28, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, T.; Yoneyama, Y.; Miwa, S.; Yoshida, A. A single nucleotide base transi- tion is the basis of the common human glucose-6-phosphate dehydrogenase variant A(+). Genomics 1987, 1, 228–231. [Google Scholar] [CrossRef]
- WHO Working Group. Glucose-6-phosphate dehydrogenase deficiency. Bull. World Health Organ. 1989, 67, 601–611. [Google Scholar]
- Kotaka, M.; Gover, S.; Vandeputte-Rutten, L.; Au, S.W.N.; Lam, V.M.S.; Adams, M.J. Structural studies of glucose-6-phosphate and NADP+ binding to human glucose-6-phosphate dehydrogenase. Acta Crystallogr. 2005, D61, 495–504. [Google Scholar] [CrossRef] [PubMed]
- The PyMOL Molecular Graphics System; DeLano Scientific: Palo Alto, CA, USA, 2002.
- Wang, X.T.; Lam, V.M.S.; Engel, P.C. Functional properties of two mutants of human glucose 6-phosphate dehydrogenase, R393G and R393H, corresponding to the clinical variants G6PD Wisconsin and Nashville. Biochim. Biophys. Acta 2006, 1762, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Choi, M.Y.; Au, S.W.; Au, D.M.; Lam, V.M.S.; Engel, P.C. Purification and detailed study of two clinically different human glucose 6-phosphate dehydrogenase variants, G6PD (Plymouth) and G6PD (Mahidol): Evidence for defective protein folding as the basis of disease. Mol. Genet. Metab. 2008, 93, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Enríquez-Flores, S.; Rodríguez-Romero, A.; Hernández-Alcántara, G.; Oria-Hernéndez, J.; Gutiérrez-Castrellón, P.; Pérez-Hernández, G.; de la Mora-de la Mora, I.; Castillo-Villanueva, A.; García-Torres, I.; Méndez, S.T.; et al. Determining the molecular mechanism of inactivation by chemical modification of triosephosphate isomerase from the human parasite Giardia lamblia: A study for antiparasitic drug design. Proteins 2011, 79, 2711–2724. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.T.; Lam, V.M.; Engel, P.C. Marked decrease in specific activity contributes to disease phenotype in two human glucose-6-phosphate dehydrogenase mutants, G6PDUnion and G6PDAndalus. Hum. Mutat. 2005, 26, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Au, S.W.N.; Gover, S.; Lam, V.M.S.; Adams, M. Human glucose-6-phosphate dehydrogenase: The crystal structure reveals a structural NADP+ molecule and provides insights into enzyme deficiency. Structure 2000, 8, 293–303. [Google Scholar] [CrossRef]
- Chotia, C. The Nature of the Accessible and Buried Surfaces in Proteins. J. Mol. Biol. 1976, 105, 1–12. [Google Scholar] [CrossRef]
- Gómez-Manzo, S.; Terrón-Hernández, J.; de la Mora-de la Mora, I.; García Torres, I.; López-Velázquez, G.; Reyes-Vivas, H.; Oria-Hernández, J. Cloning, expression, purification and characterization of His-tagged human glucose-6-phosphate dehydrogenase: A simplified method for protein yield. Protein J. 2013, 32, 585–592. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Manzo, S.; Marcial-Quino, J.; Vanoye-Carlo, A.; Enríquez-Flores, S.; De la Mora-De la Mora, I.; González-Valdez, A.; García-Torres, I.; Martínez-Rosas, V.; Sierra-Palacios, E.; Lazcano-Pérez, F.; et al. Mutations of Glucose-6-Phosphate Dehydrogenase Durham, Santa-Maria and A+ Variants Are Associated with Loss Functional and Structural Stability of the Protein. Int. J. Mol. Sci. 2015, 16, 28657-28668. https://doi.org/10.3390/ijms161226124
Gómez-Manzo S, Marcial-Quino J, Vanoye-Carlo A, Enríquez-Flores S, De la Mora-De la Mora I, González-Valdez A, García-Torres I, Martínez-Rosas V, Sierra-Palacios E, Lazcano-Pérez F, et al. Mutations of Glucose-6-Phosphate Dehydrogenase Durham, Santa-Maria and A+ Variants Are Associated with Loss Functional and Structural Stability of the Protein. International Journal of Molecular Sciences. 2015; 16(12):28657-28668. https://doi.org/10.3390/ijms161226124
Chicago/Turabian StyleGómez-Manzo, Saúl, Jaime Marcial-Quino, America Vanoye-Carlo, Sergio Enríquez-Flores, Ignacio De la Mora-De la Mora, Abigail González-Valdez, Itzhel García-Torres, Víctor Martínez-Rosas, Edgar Sierra-Palacios, Fernando Lazcano-Pérez, and et al. 2015. "Mutations of Glucose-6-Phosphate Dehydrogenase Durham, Santa-Maria and A+ Variants Are Associated with Loss Functional and Structural Stability of the Protein" International Journal of Molecular Sciences 16, no. 12: 28657-28668. https://doi.org/10.3390/ijms161226124
APA StyleGómez-Manzo, S., Marcial-Quino, J., Vanoye-Carlo, A., Enríquez-Flores, S., De la Mora-De la Mora, I., González-Valdez, A., García-Torres, I., Martínez-Rosas, V., Sierra-Palacios, E., Lazcano-Pérez, F., Rodríguez-Bustamante, E., & Arreguin-Espinosa, R. (2015). Mutations of Glucose-6-Phosphate Dehydrogenase Durham, Santa-Maria and A+ Variants Are Associated with Loss Functional and Structural Stability of the Protein. International Journal of Molecular Sciences, 16(12), 28657-28668. https://doi.org/10.3390/ijms161226124