
Andrographolide Inhibits Ovariectomy-Induced Bone Loss via the Suppression of RANKL Signaling Pathways


Abstract
:1. Introduction
2. Results
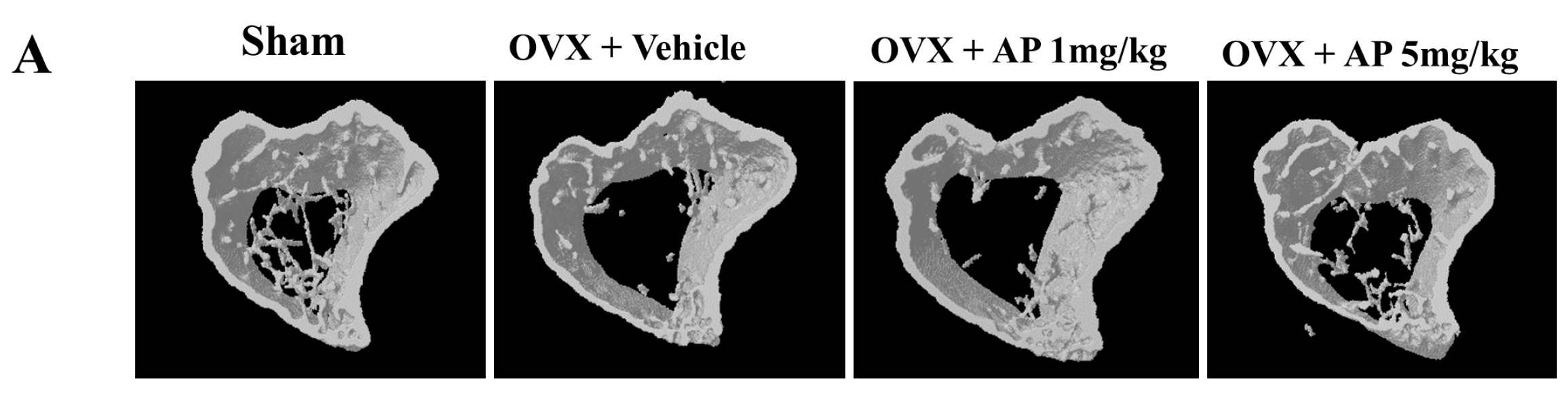
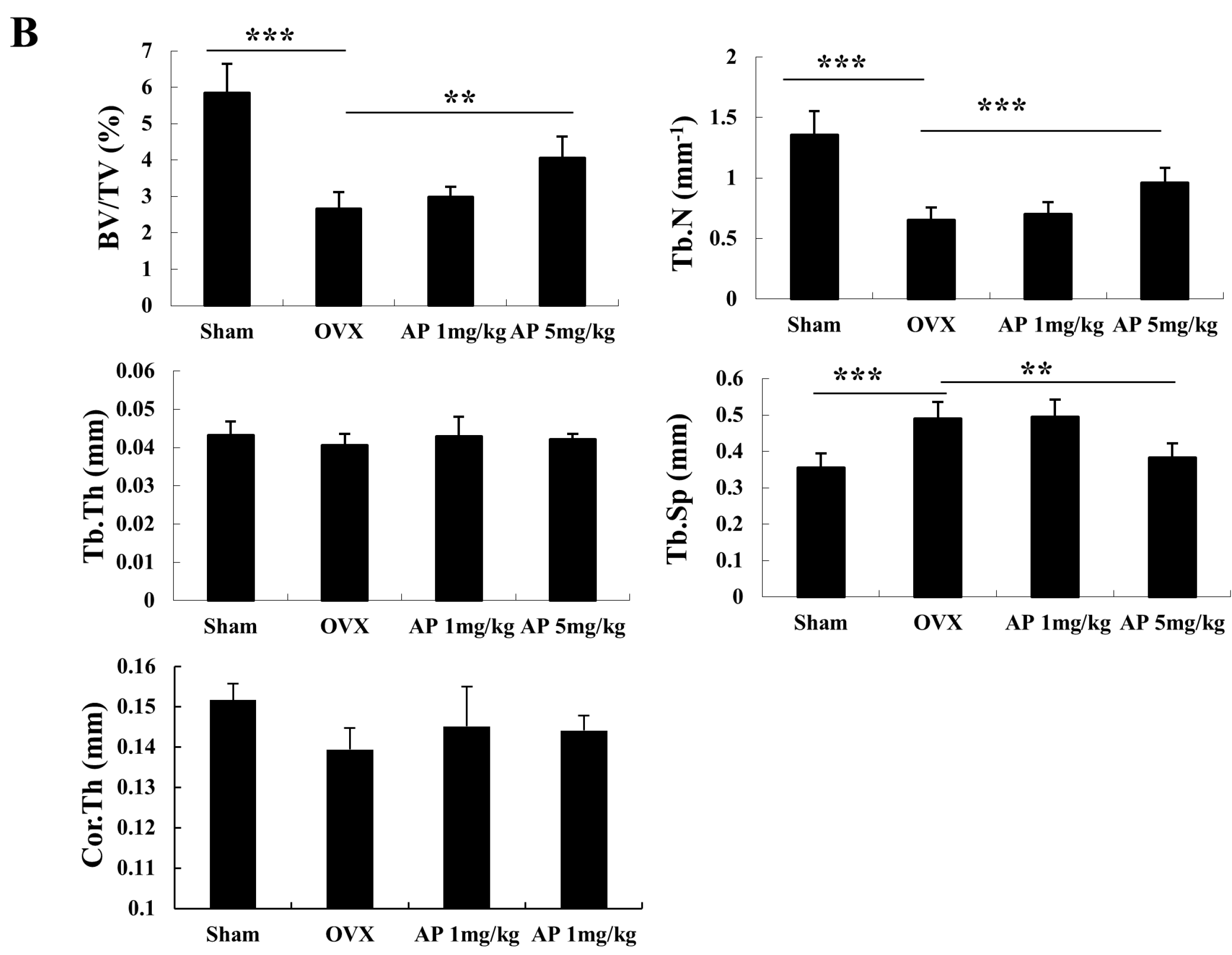
2.1. Andrographolide (AP) Inhibits Ovariectomy-Induced Bone Loss in Vivo
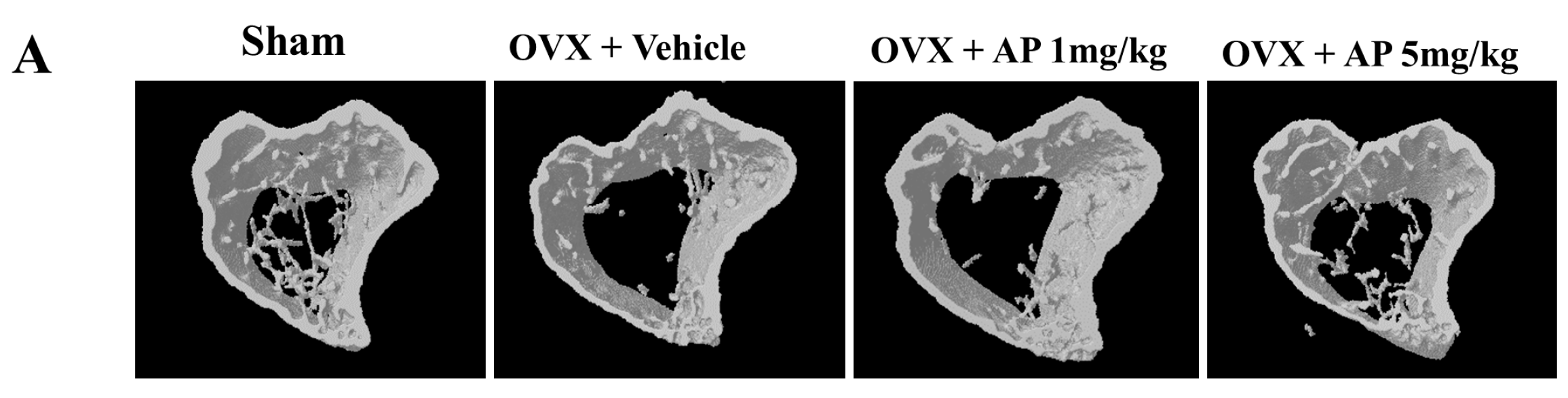
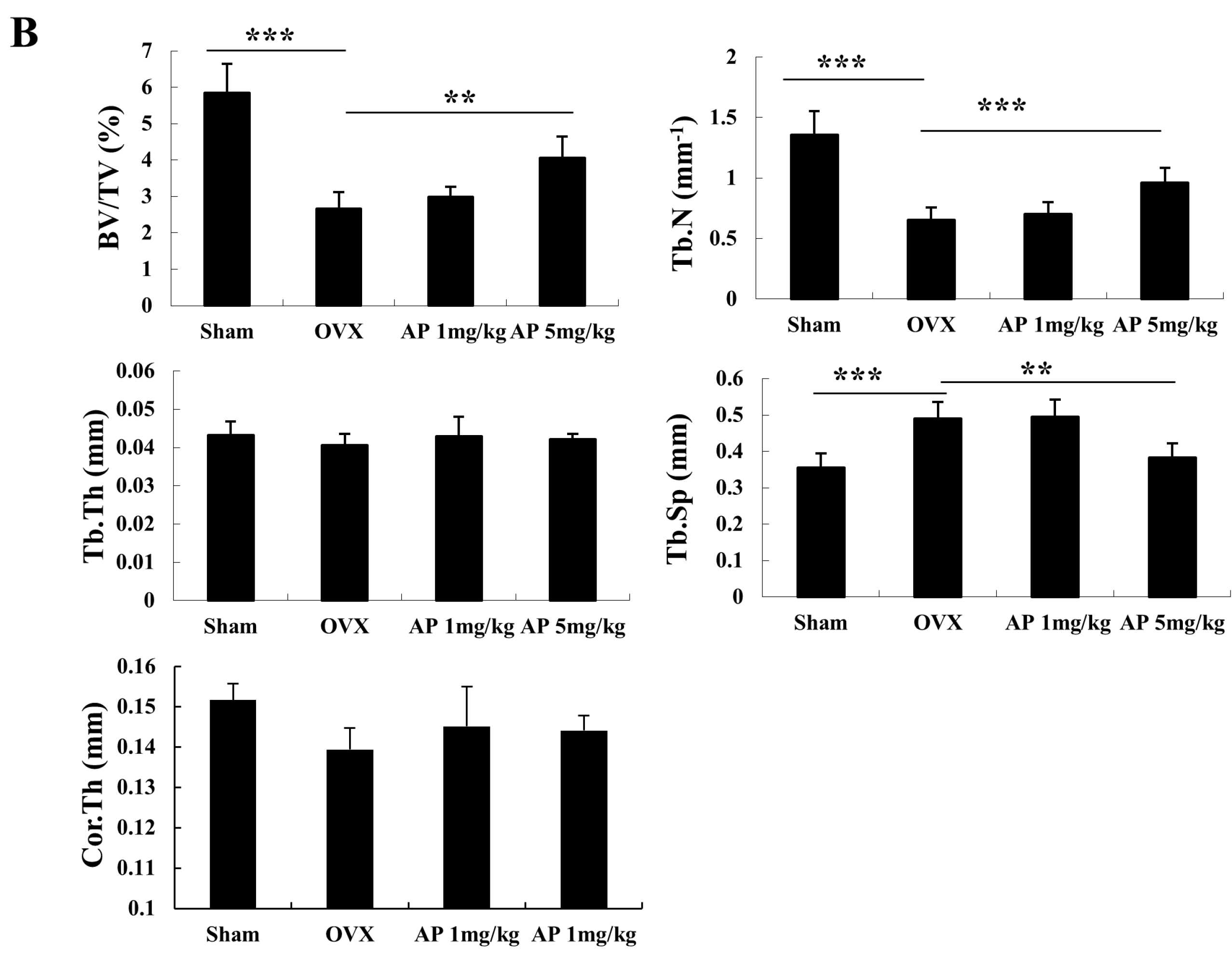
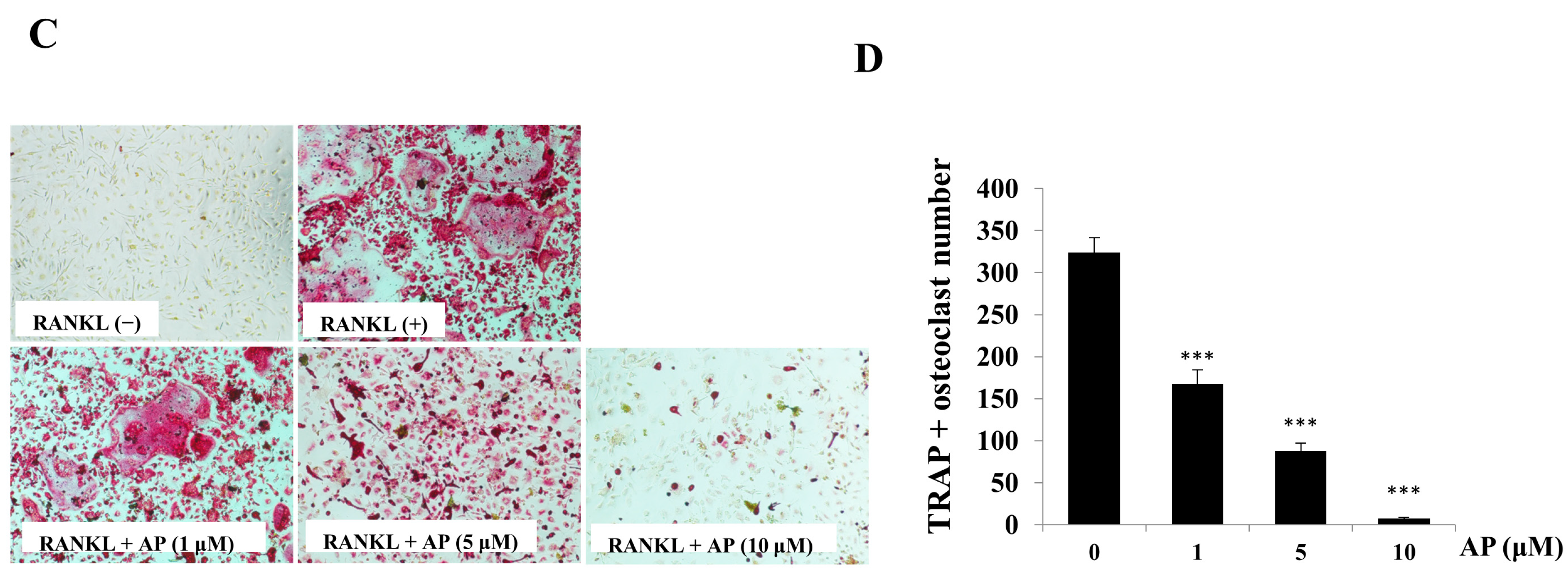
2.2. AP Inhibits Receptor Activator of Nuclear Factor-κB Ligand (RANKL)-Induced Osteoclastogenesis
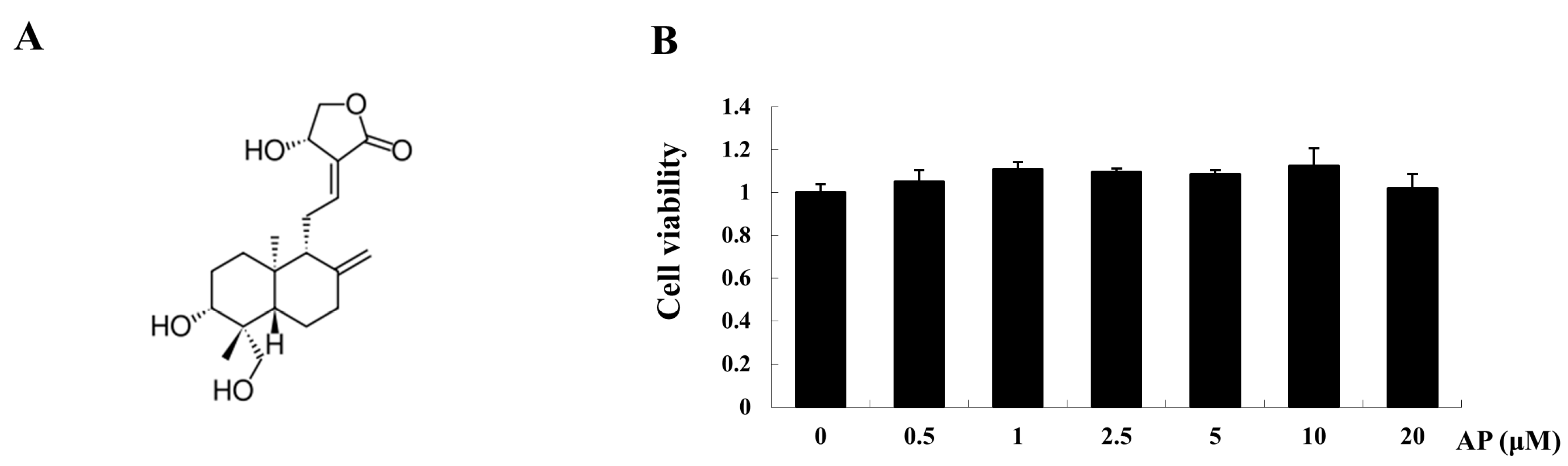

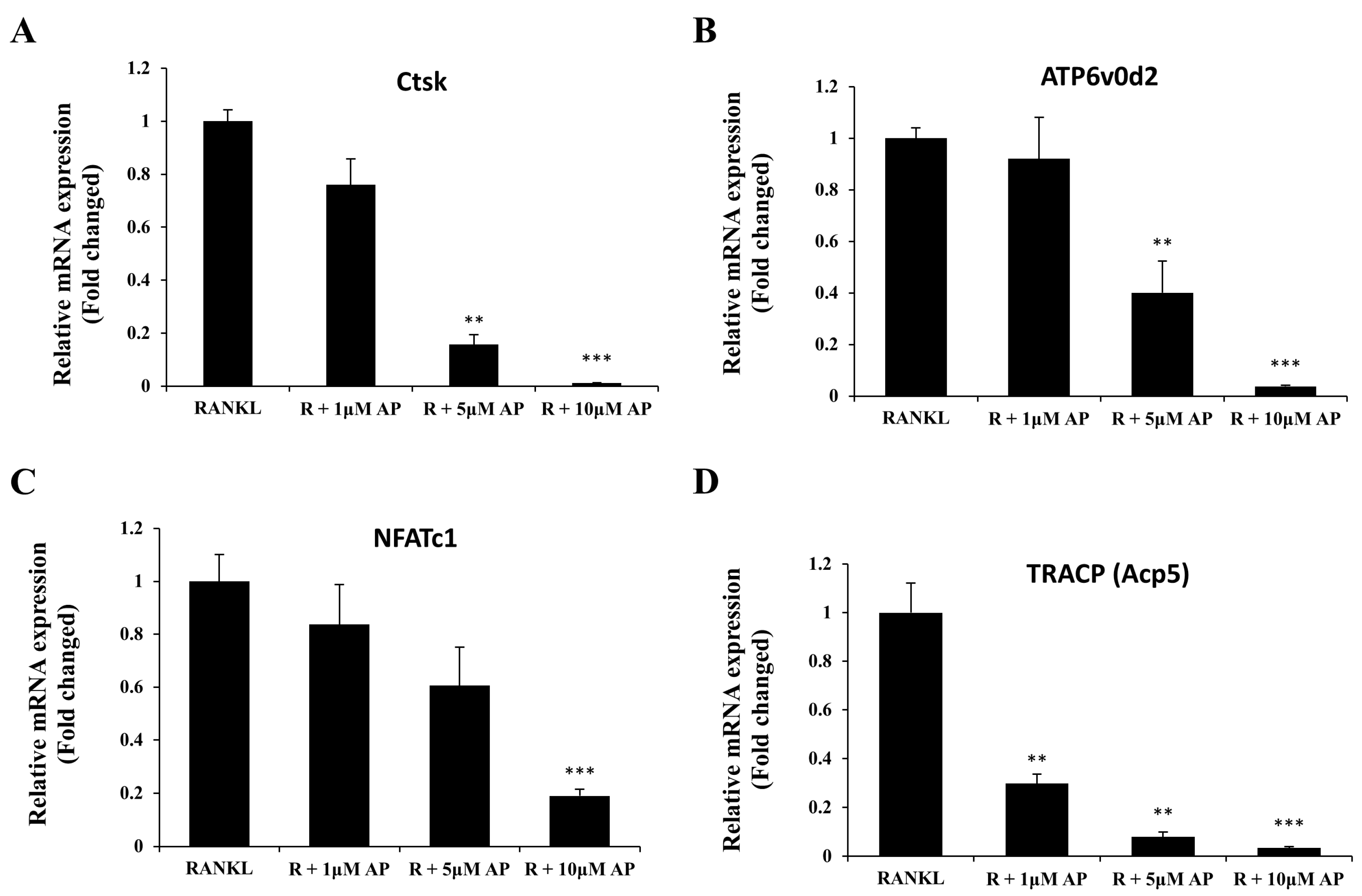
2.3. AP Suppresses Osteoclast-Related Gene Expression

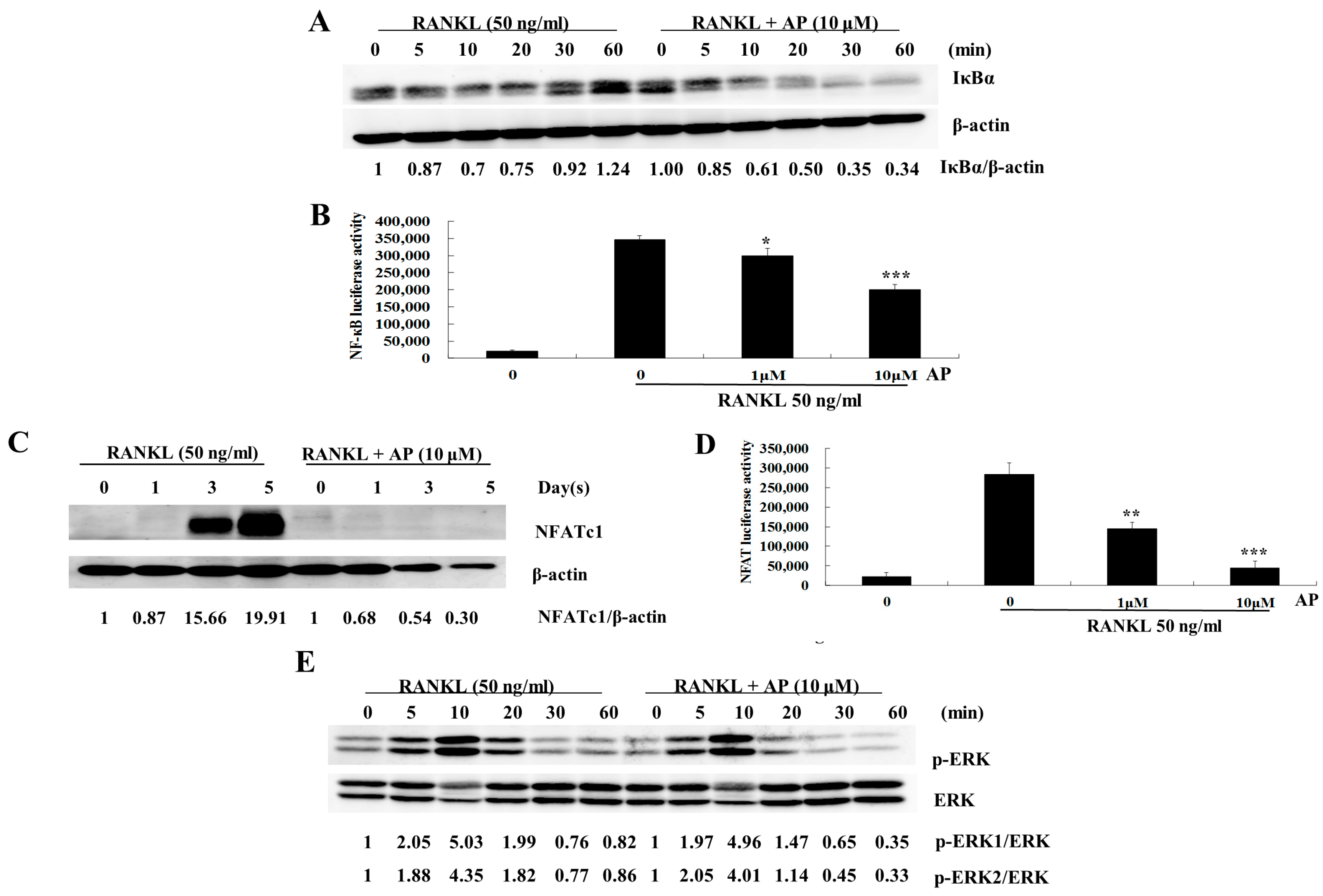
2.4. AP Suppresses RANKL-Induced Nuclear Factor-κB (NF-κB) Activation
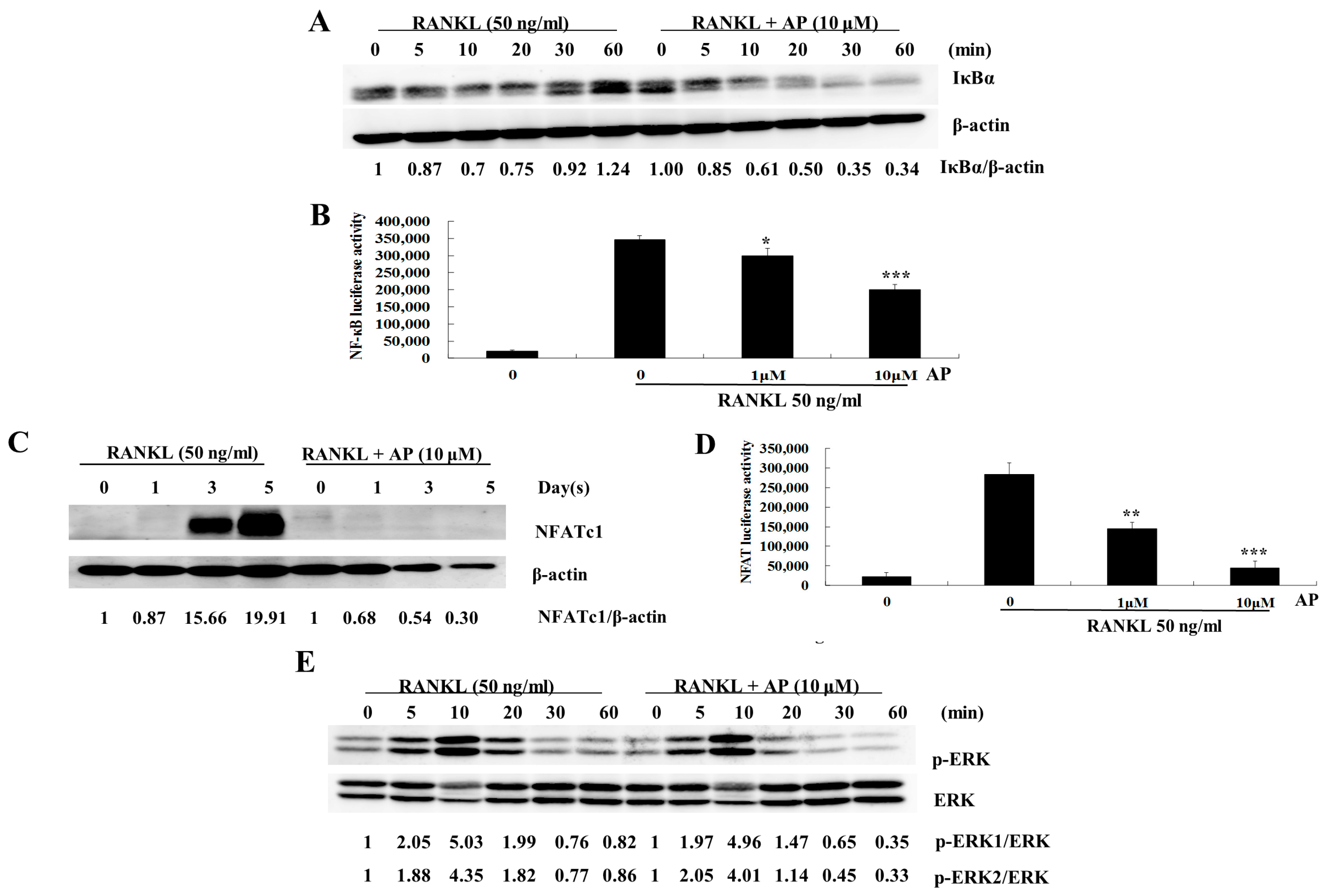
2.5. AP Suppresses RANKL-Induced Nuclear Factor of Activated T-Cells (NFATc1) Activation
2.6. AP Does not Affect RANKL-Induced Extracellular Signal-Regulated Kinases (ERK) Phosphorylation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Reagents and Antibodies
4.3. In Vitro Osteoclastogenesis Assay
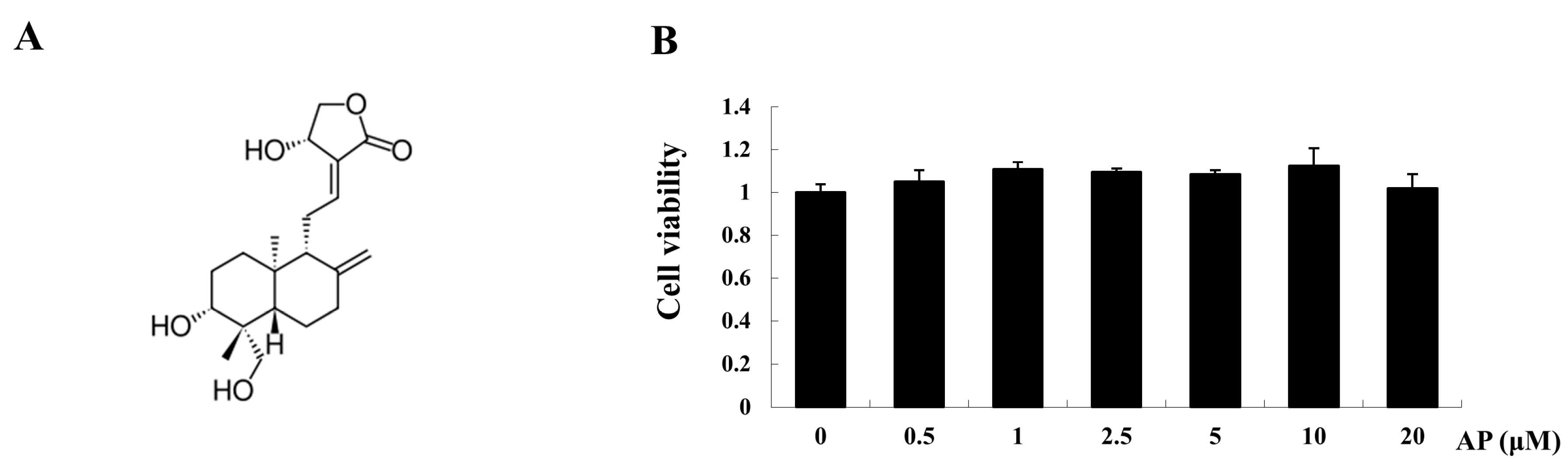
4.4. Cytotoxicity Assay
4.5. Real Time Polymerase Chain Reaction (Real-Time PCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA | Primer | Sequences (5ʹ–3ʹ) | Product (bp) |
|---|---|---|---|
| Ctsk | Forward | GGGAGAAAAACCTGAAGC | 350 |
| Reverse | ATTCTGGGGACTCAGAGC | ||
| ATP6v0d2 | Forward | GTGAGACCTTGGAAGACCTGAA | 176 |
| Reverse | GAGAAATGTGCTCAGGGGCT | ||
| NFATc1 | Forward | CAACGCCCTGACCACCGATAG | 392 |
| Reverse | GGCTGCCTTCCGTCTCATAGT | ||
| TRACP (Acp5) | Forward | TGTGGCCATCTTTATGCT | 462 |
| Reverse | GTCATTTCTTTGGGGCTT | ||
| GAPDH | Forward | ACCACAGTCCATGCCATCAC | 452 |
| Reverse | TCCACCACCCTGTTGCTGTA |
4.6. Western Blot Assay
4.7. Luciferase Reporter Gene Assay for NF-κB and NFAT
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Blume, S.W.; Curtis, J.R. Medical costs of osteoporosis in the elderly medicare population. Osteoporos. Int. 2011, 22, 1835–1844. [Google Scholar] [CrossRef] [PubMed]
- Hadjidakis, D.J.; Androulakis, I.I. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.C.; Xu, J.; Zheng, M.H. Interaction between osteoblast and osteoclast: Impact in bone disease. Histol. Histopathol. 2004, 19, 1325–1344. [Google Scholar] [PubMed]
- Gamble, C.L. Osteoporosis: Making the diagnosis in patients at risk for fracture. Geriatrics 1995, 50, 24–26, 29–30, 33. [Google Scholar] [PubMed]
- Kassem, M.; Abdallah, B.M.; Saeed, H. Osteoblastic cells: Differentiation and trans-differentiation. Arch. Biochem. Biophys. 2008, 473, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, N.; Takahashi, N.; Akatsu, T.; Tanaka, H.; Sasaki, T.; Nishihara, T.; Koga, T.; Martin, T.J.; Suda, T. Origin of osteoclasts: Mature monocytes and macrophages are capable of differentiating into osteoclasts under a suitable microenvironment prepared by bone marrow-derived stromal cells. Proc. Natl. Acad. Sci. USA 1990, 87, 7260–7264. [Google Scholar] [CrossRef] [PubMed]
- Gay, C.V.; Gilman, V.R.; Sugiyama, T. Perspectives on osteoblast and osteoclast function. Poult. Sci. 2000, 79, 1005–1008. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.R.; Mundy, G.R.; Clark, S.; Wang, E.A.; Kuehl, T.J.; Stanley, E.R.; Roodman, G.D. Effects of human recombinant CSF-GM and highly purified CSF-1 on the formation of multinucleated cells with osteoclast characteristics in long-term bone marrow cultures. J. Bone Miner. Res. 1986, 1, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Akbar, S. Andrographis paniculata: A review of pharmacological activities and clinical effects. Altern. Med. Rev. 2011, 16, 66–77. [Google Scholar] [PubMed]
- Zhang, Q.Q.; Ding, Y.; Lei, Y.; Qi, C.L.; He, X.D.; Lan, T.; Li, J.C.; Gong, P.; Yang, X.; Geng, J.G.; et al. Andrographolide suppress tumor growth by inhibiting TLR4/NF-κB signaling activation in insulinoma. Int. J. Biol. Sci. 2014, 10, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ghefreh, A.A.; Canatan, H.; Ezeamuzie, C.I. In vitro and in vivo anti-inflammatory effects of andrographolide. Int. Immunopharmacol. 2009, 9, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.J.; Zheng, X.W.; Feng, T.; Huang, J.; Chen, J.; Wu, Y.Y.; Zhou, L.M.; Tu, W.W.; Li, H. Andrographolide exerted its antimicrobial effects by upregulation of human β-defensin-2 induced through p38 MAPK and NF-κB pathway in human lung epithelial cells. Can. J. Physiol. Pharmacol. 2012, 90, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.S.; Loh, X.Y.; Wijaya, H.; Wang, J.; Lin, Q.; Lam, Y.; Wong, W.S.; Mok, Y.K. Specificity and inhibitory mechanism of andrographolide and its analogues as antiasthma agents on NF-κB p50. J. Nat. Prod. 2015, 78, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.J.; Wong, W.S.; Wong, P.T.; Bian, J.S. Neuroprotective effects of andrographolide in a rat model of permanent cerebral ischaemia. Br. J. Pharmacol. 2010, 161, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Al Batran, R.; Al-Bayaty, F.H.; Al-Obaidi, M.M. In-vivo effect of andrographolide on alveolar bone resorption induced by porphyromonas gingivalis and its relation with antioxidant enzymes. Biomed. Res. Int. 2013, 2013, 276329. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.J.; Li, H.W.; Liu, G.W.; Qu, X.H.; Tian, B.; Yan, W.; Lin, Z.; Tang, T.T.; Qin, A.; Dai, K.R. Andrographolide suppresses rankl-induced osteoclastogenesis in vitro and prevents inflammatory bone loss in vivo. Br. J. Pharmacol. 2014, 171, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Qu, X.; Yan, W.; Li, H.; Liu, G.; Liu, X.; Tang, T.; Qin, A.; Dai, K. Andrographolide prevents human breast cancer-induced osteoclastic bone loss via attenuated rankl signaling. Breast Cancer Res. Treat. 2014, 144, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Xiu, Y.; Li, J.; Xing, L.; Yao, Z. NF-κB-mediated regulation of osteoclastogenesis. Endocrinol. Metab. 2015, 30, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef]
- Kim, K.; Lee, S.H.; Ha Kim, J.; Choi, Y.; Kim, N. NFATc1 induces osteoclast fusion via up-regulation of Atp6v0d2 and the dendritic cell-specific transmembrane protein (DC-STAMP). Mol. Endocrinol. 2008, 22, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Sudo, T.; Saito, T.; Osada, H.; Tsujimoto, M. Involvement of p38 mitogen-activated protein kinase signaling pathway in osteoclastogenesis mediated by receptor activator of NF-κB ligand (RANKL). J. Biol. Chem. 2000, 275, 31155–31161. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.; Lacey, D.L.; Dunstan, C.R.; Solovyev, I.; Colombero, A.; Timms, E.; Tan, H.L.; Elliott, G.; Kelley, M.J.; Sarosi, I.; et al. Tumor necrosis factor receptor family member rank mediates osteoclast differentiation and activation induced by osteoprotegerin ligand. Proc. Natl. Acad. Sci. USA 1999, 96, 3540–3545. [Google Scholar] [CrossRef] [PubMed]
- Weitzmann, M.N.; Pacifici, R. Estrogen deficiency and bone loss: An inflammatory tale. J. Clin. Investig. 2006, 116, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Rosenberg, E.; de Papp, A.E.; Duong, L.T. The osteoclast, bone remodelling and treatment of metabolic bone disease. Eur. J. Clin. Investig. 2012, 42, 1332–1341. [Google Scholar] [CrossRef] [PubMed]
- Ang, E.; Liu, Q.; Qi, M.; Liu, H.G.; Yang, X.; Chen, H.; Zheng, M.H.; Xu, J. Mangiferin attenuates osteoclastogenesis, bone resorption, and rankl-induced activation of NF-κB and ERK. J. Cell. Biochem. 2011, 112, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.K.; Ke, K.; Sul, O.J.; Kim, H.J.; Kim, S.H.; Lee, M.H.; Kim, H.J.; Kim, S.Y.; Chung, H.T.; Choi, H.S. Curcumin protects against ovariectomy-induced bone loss and decreases osteoclastogenesis. J. Cell. Biochem. 2011, 112, 3159–3166. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Sun, X.; Ma, J.; Ma, X.; Zhao, B.; Zhang, Y.; Tian, P.; Li, Y.; Han, Z. Naringin prevents ovariectomy-induced osteoporosis and promotes osteoclasts apoptosis through the mitochondria-mediated apoptosis pathway. Biochem. Biophys. Res. Commun. 2014, 452, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, E.F.; Hodgson, S.F.; Eastell, R.; Cedel, S.L.; O’Fallon, W.M.; Riggs, B.L. Cancellous bone remodeling in type I (postmenopausal) osteoporosis: Quantitative assessment of rates of formation, resorption, and bone loss at tissue and cellular levels. J. Bone Miner. Res. 1990, 5, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Yavropoulou, M.P.; Yovos, J.G. Osteoclastogenesis—Current knowledge and future perspectives. J. Musculoskelet. Neuronal Interact. 2008, 8, 204–216. [Google Scholar] [PubMed]
- Feng, X.; McDonald, J.M. Disorders of bone remodeling. Annu. Rev. Pathol. 2011, 6, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Iotsova, V.; Caamano, J.; Loy, J.; Yang, Y.; Lewin, A.; Bravo, R. Osteopetrosis in mice lacking NF-κB1 and NF-κB2. Nat. Med. 1997, 3, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Franzoso, G.; Carlson, L.; Xing, L.; Poljak, L.; Shores, E.W.; Brown, K.D.; Leonardi, A.; Tran, T.; Boyce, B.F.; Siebenlist, U. Requirement for NF-κB in osteoclast and B-cell development. Genes Dev. 1997, 11, 3482–3496. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.F.; Ye, B.Q.; Li, Y.D.; Wang, J.G.; He, X.J.; Lin, X.; Yao, X.; Ma, D.; Slungaard, A.; Hebbel, R.P.; et al. Andrographolide attenuates inflammation by inhibition of NF-κB activation through covalent modification of reduced cysteine 62 of p50. J. Immunol. 2004, 173, 4207–4217. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wang, X.; Liu, Y.; He, A.; Jia, R. Nfatc1: Functions in osteoclasts. Int. J. Biochem. Cell Biol. 2010, 42, 576–579. [Google Scholar] [CrossRef] [PubMed]
- Aliprantis, A.O.; Ueki, Y.; Sulyanto, R.; Park, A.; Sigrist, K.S.; Sharma, S.M.; Ostrowski, M.C.; Olsen, B.R.; Glimcher, L.H. NFATc1 in mice represses osteoprotegerin during osteoclastogenesis and dissociates systemic osteopenia from inflammation in cherubism. J. Clin. Investig. 2008, 118, 3775–3789. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, R. Estrogen, cytokines, and pathogenesis of postmenopausal osteoporosis. J. Bone Miner. Res. 1996, 11, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Cenci, S.; Weitzmann, M.N.; Roggia, C.; Namba, N.; Novack, D.; Woodring, J.; Pacifici, R. Estrogen deficiency induces bone loss by enhancing T-cell production of TNF-α. J. Clin. Investig. 2000, 106, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tan, J.W.; Huang, L.; Gao, X.H.; Laird, R.; Liu, D.; Wysocki, S.; Zheng, M.H. Cloning, sequencing, and functional characterization of the rat homologue of receptor activator of NF-κB ligand. J. Bone Miner. Res. 2000, 15, 2178–2186. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Steer, J.H.; Joyce, D.A.; Yip, K.H.; Zheng, M.H.; Xu, J. 12-O-tetradecanoylphorbol-13-acetate (TPA) inhibits osteoclastogenesis by suppressing rankl-induced NF-κB activation. J. Bone Miner. Res. 2003, 18, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- van der Kraan, A.G.; Chai, R.C.; Singh, P.P.; Lang, B.J.; Xu, J.; Gillespie, M.T.; Price, J.T.; Quinn, J.M. Hsp90 inhibitors enhance differentiation and MITF (microphthalmia transcription factor) activity in osteoclast progenitors. Biochem. J. 2013, 451, 235–244. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Liu, Q.; Zhou, L.; Yuan, J.B.; Lin, X.; Zeng, R.; Liang, X.; Zhao, J.; Xu, J. Andrographolide Inhibits Ovariectomy-Induced Bone Loss via the Suppression of RANKL Signaling Pathways. Int. J. Mol. Sci. 2015, 16, 27470-27481. https://doi.org/10.3390/ijms161126039
Wang T, Liu Q, Zhou L, Yuan JB, Lin X, Zeng R, Liang X, Zhao J, Xu J. Andrographolide Inhibits Ovariectomy-Induced Bone Loss via the Suppression of RANKL Signaling Pathways. International Journal of Molecular Sciences. 2015; 16(11):27470-27481. https://doi.org/10.3390/ijms161126039
Chicago/Turabian StyleWang, Tao, Qian Liu, Lin Zhou, Jin Bo Yuan, Xixi Lin, Rong Zeng, Xiaonan Liang, Jinmin Zhao, and Jiake Xu. 2015. "Andrographolide Inhibits Ovariectomy-Induced Bone Loss via the Suppression of RANKL Signaling Pathways" International Journal of Molecular Sciences 16, no. 11: 27470-27481. https://doi.org/10.3390/ijms161126039
APA StyleWang, T., Liu, Q., Zhou, L., Yuan, J. B., Lin, X., Zeng, R., Liang, X., Zhao, J., & Xu, J. (2015). Andrographolide Inhibits Ovariectomy-Induced Bone Loss via the Suppression of RANKL Signaling Pathways. International Journal of Molecular Sciences, 16(11), 27470-27481. https://doi.org/10.3390/ijms161126039