
Responses to Oxidative and Heavy Metal Stresses in Cyanobacteria: Recent Advances

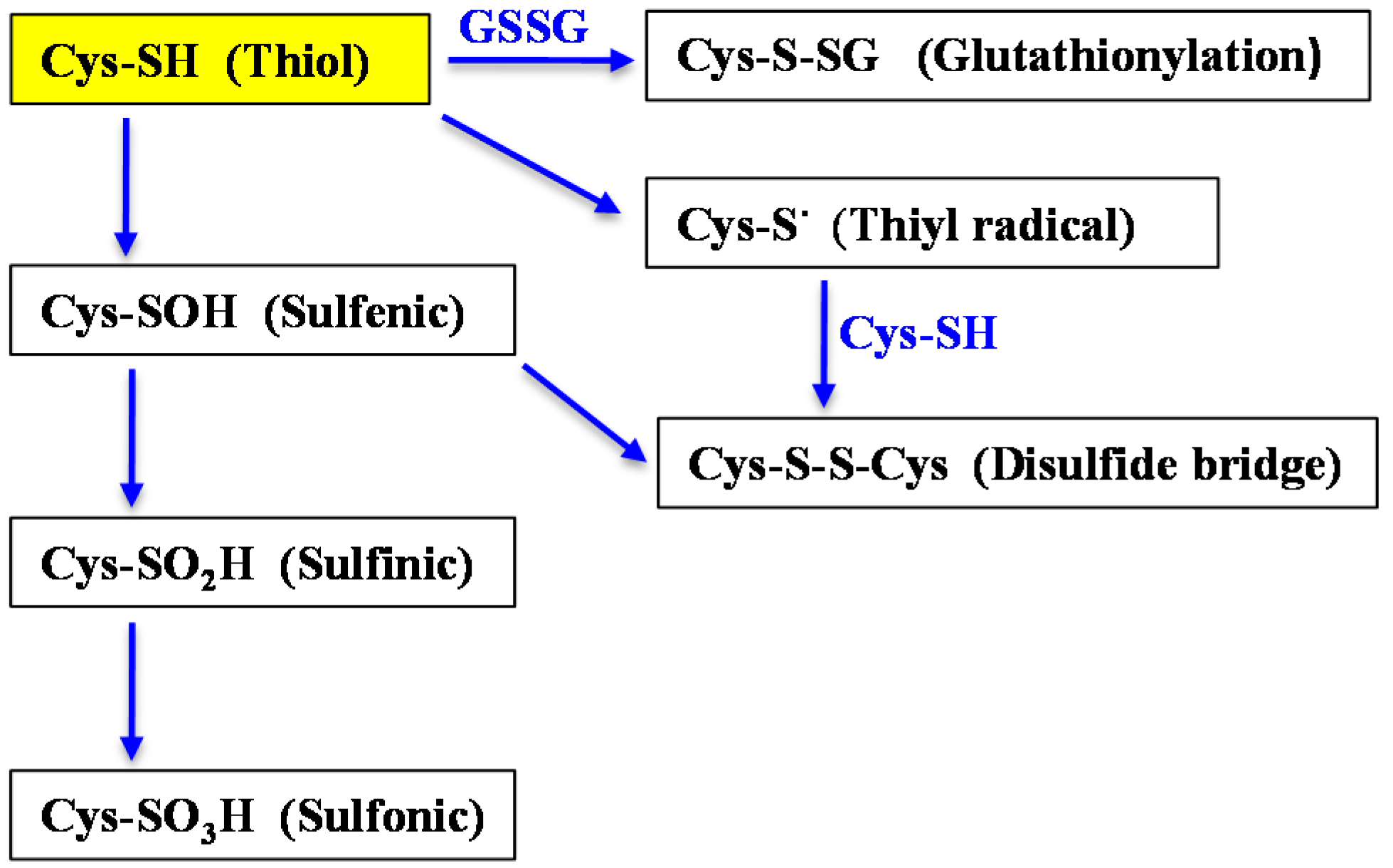
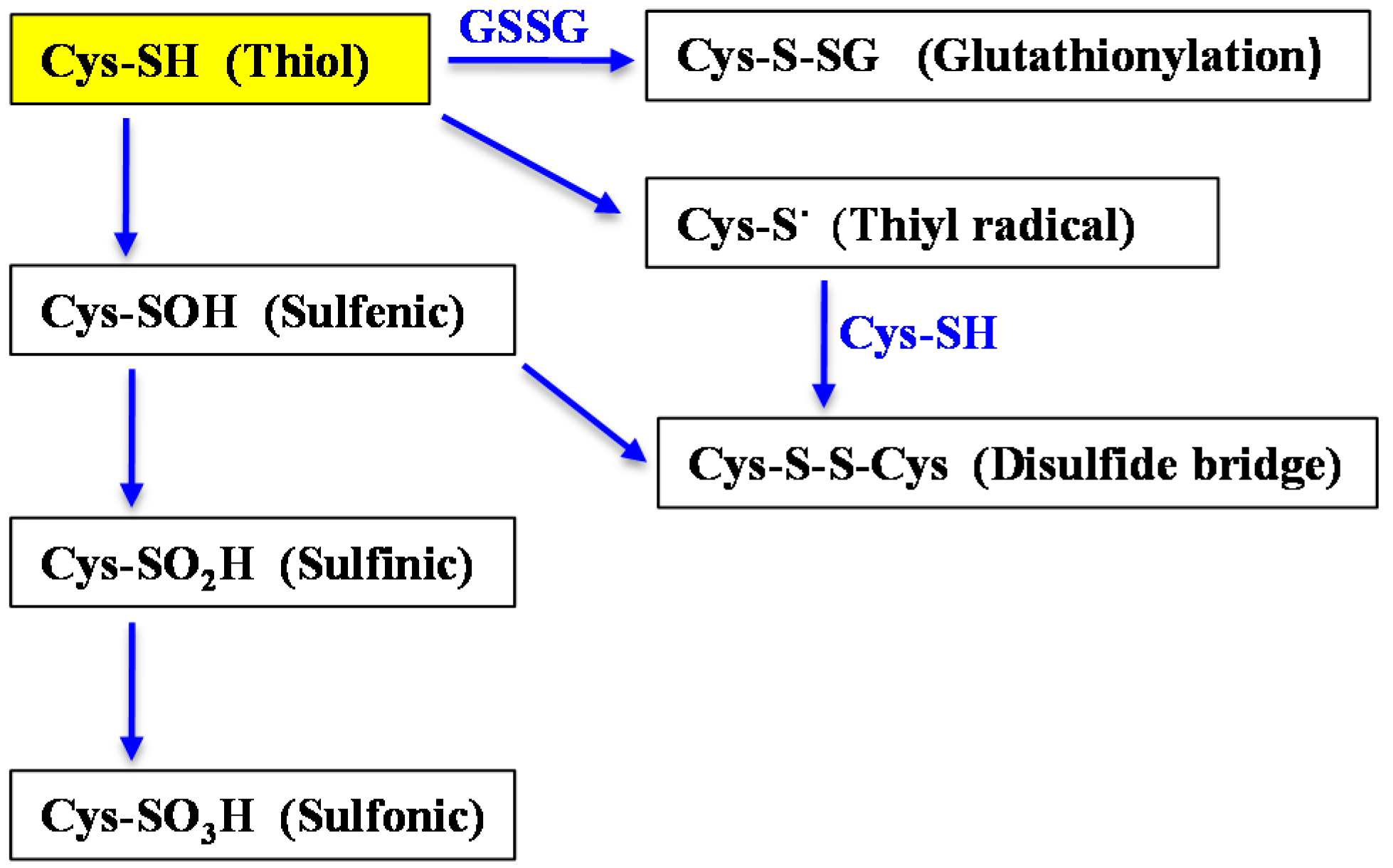{kind=link}
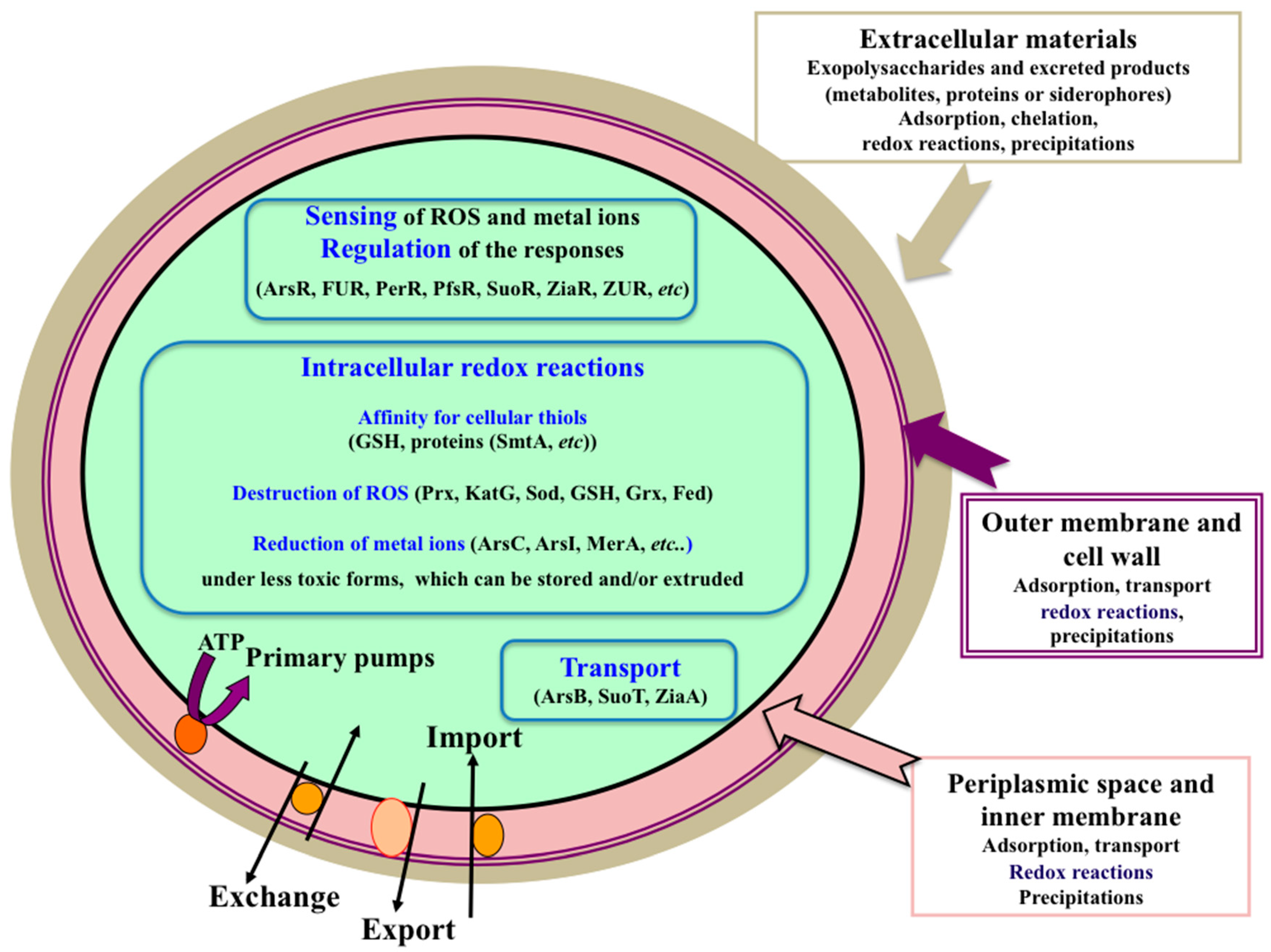{kind=link}
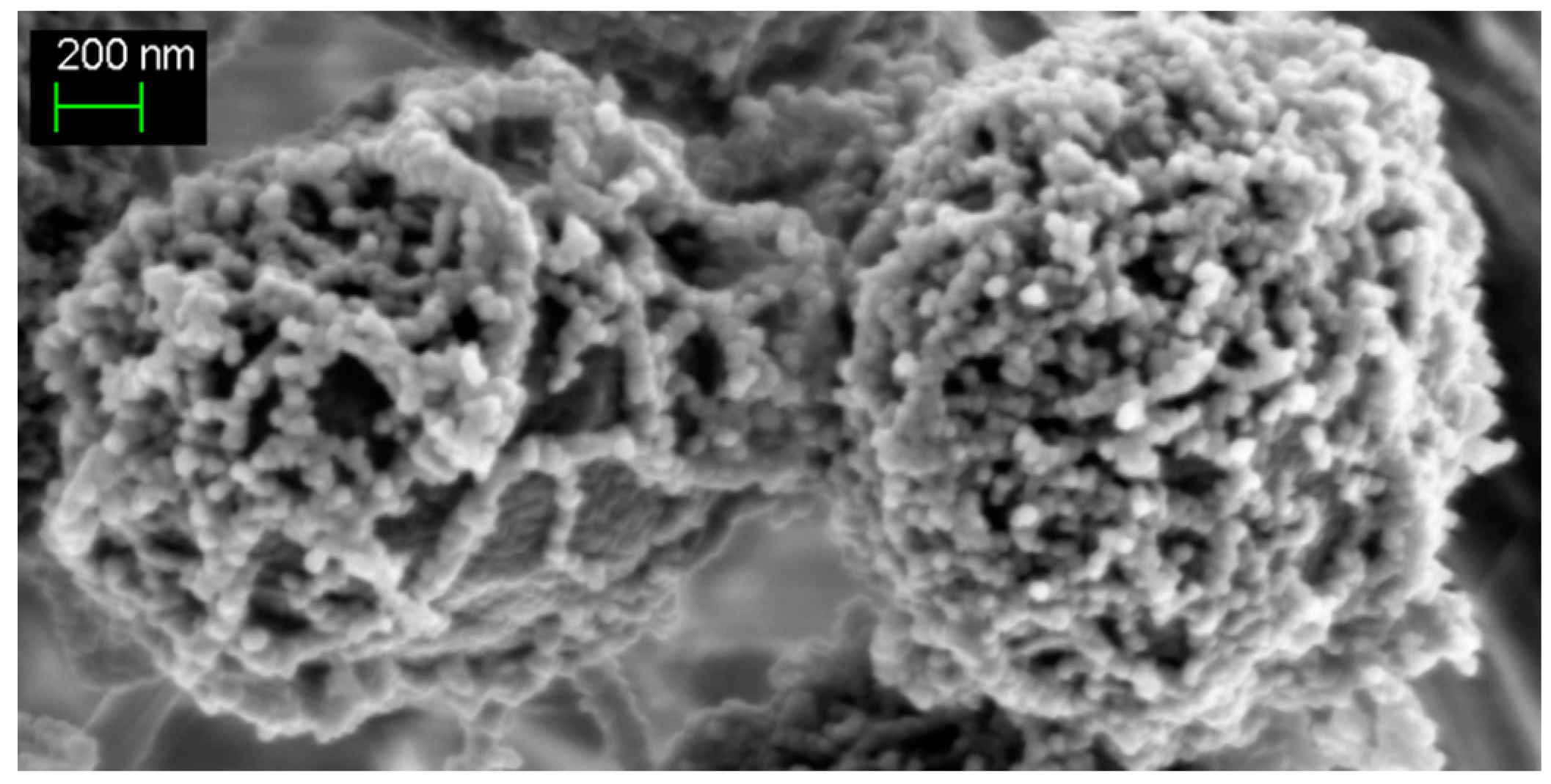{kind=link}
Abstract
:1. Introduction
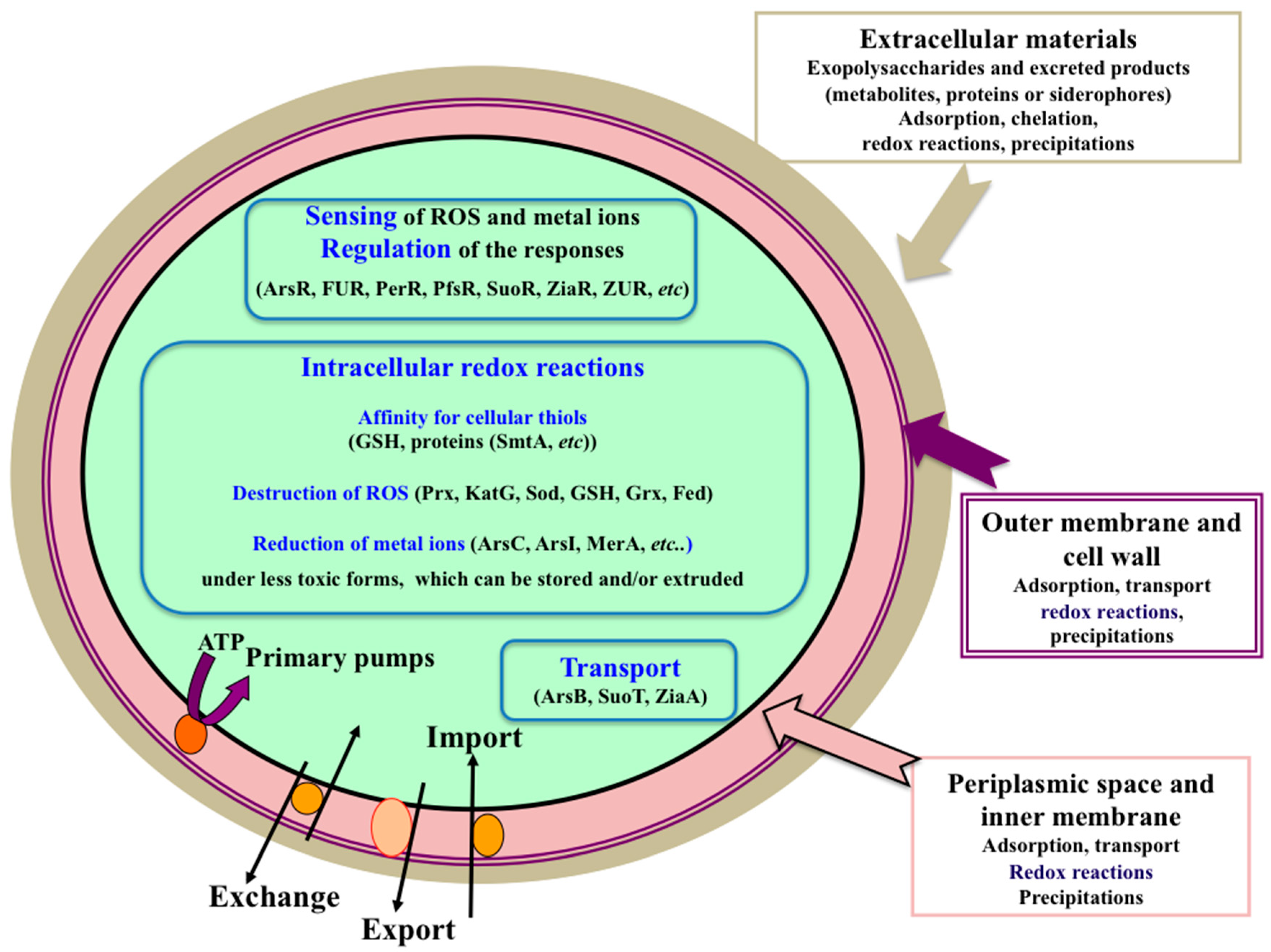
2. The Extracellular Mantle of Exopolysaccharides Constitutes the First Protective Barrier against Metal Stresses (Cd, Co, Fe, and CeO2 and TiO2 Nanoparticles)
3. Metallothionein and/or Efflux ATPases Operate in Zinc Homeostasis and Cadmium Tolerance
4. Transport Systems and Redox Enzymes Operate in the Defense against Arsenic
5. The Mercuric Reductase-Like Enzyme Plays a Prominent Role in the Reduction of, and Protection against, both Mercury and Uranium
6. Glutathione Protects Cells against Oxidative and Metal Stresses
7. The Glutaredoxin Enzymes Operate in the Protection against Oxidative and Metal Stresses
8. The Ferredoxin Enzymes Are Involved in the Tolerance to Oxidative and Metal Stresses
9. The Fur-Like Regulators Play a Crucial Role in Metal Homeostasis and the Tolerance to Oxidative Stress and Heavy Metals
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schopf, W.J. The paleobiological record of photosynthesis. Photosynth. Res. 2011, 107, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Archibald, J.M. The puzzle of plastid evolution. Curr. Biol. 2009, 19, R81–R88. [Google Scholar] [CrossRef] [PubMed]
- Jansson, C.; Northen, T. Calcifying cyanobacteria—The potential of biomineralization for carbon capture and storage. Curr. Opin. Biotechnol. 2010, 21, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Rosgaard, L.; de Porcellinis, A.J.; Jacobsen, J.H.; Frigaard, N.U.; Sakuragi, Y. Bioengineering of carbon fixation, biofuels, and biochemicals in cyanobacteria and plants. J. Biotechnol. 2012, 162, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Cassier-Chauvat, C.; Veaudor, T.; Chauvat, F. Advances in the function and regulation of hydrogenase in the cyanobacterium Synechocystis PCC6803. Int. J. Mol. Sci. 2014, 15, 19938–19951. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Kaneko, T.; Hirosawa, M.; Miyajima, N.; Tabata, S. CyanoBase, a www database containing the complete nucleotide sequence of the genome of Synechocystis sp. strain PCC6803. Nucleic Acids Res. 1998, 26, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Grigorieva, G.; Shestakov, S. Transformation in the cyanobacterium Synechocystis sp 6803. FEMS Microbiol. Lett. 1982, 13, 367–370. [Google Scholar] [CrossRef]
- Marraccini, P.; Bulteau, S.; Cassier-Chauvat, C.; Mermet-Bouvier, P.; Chauvat, F. A conjugative plasmid vector for promoter analysis in several cyanobacteria of the genera Synechococcus and Synechocystis. Plant Mol. Biol. 1993, 23, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Mermet-Bouvier, P.; Chauvat, F. A conditional expression vector for the cyanobacteria Synechocystis sp. PCC6803 and PCC6714 or Synechococcus sp. PCC7942 and PCC6301. Curr. Microbiol. 1994, 28, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Poncelet, M.; Cassier-Chauvat, C.; Leschelle, X.; Bottin, H.; Chauvat, F. Targeted deletion and mutational analysis of the essential [2Fe–2S] plant-like ferredoxin in Synechocystis PCC6803 by plasmid shuffling. Mol. Microbiol. 1998, 28, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Ramos, M.; Jittawuttipoka, T.; Saenkham, P.; Czarnecka-Kwasiborski, A.; Bottin, H.; Cassier-Chauvat, C.; Chauvat, F. Engineering Synechocystis PCC6803 for hydrogen production: Influence on the tolerance to oxidative and sugar stresses. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Hess, W.R. Cyanobacterial genomics for ecology and biotechnology. Curr. Opin. Microbiol. 2011, 14, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Narainsamy, K.; Marteyn, B.; Sakr, S.; Cassier-Chauvat, C.; Chauvat, F. Genomics of the pleïotropic glutathione system in cyanobacteria. In Advances in Botanical Research; Chauvat, F., Cassier-Chauvat, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 65, pp. 157–188. [Google Scholar]
- Chauvat, F.; de Vries, L.; van der Ende, A.; van Arkel, G.A. A host-vector system for gene cloning in the cyanobacterium Synechocystis PCC6803. Mol. Gen. Genet. 1986, 204, 185–191. [Google Scholar] [CrossRef]
- Hagemann, M. Genomics of salt acclimation: Synthesis of compatible solutes among cyanobacteria. In Advances in Botanical Research; Chauvat, F., Cassier-Chauvat, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 65, pp. 27–55. [Google Scholar]
- Kirilovsky, D.; Kerfeld, C.A. Structural, mechanistic and genomic insights into OCP-mediated photoprotection. In Advances in Botanical Research; Chauvat, F., Cassier-Chauvat, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 65, pp. 1–26. [Google Scholar]
- Mejean, A.; Ploux, O. A genomic view of secondary metabolite production in cyanobacteria. In Advances in Botanical Research; Chauvat, F., Cassier-Chauvat, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; Volume 65, pp. 189–234. [Google Scholar]
- Cassier-Chauvat, C.; Chauvat, F. Cell division in cyanobacteria. In The Cell Biology of Cyanobacteria; Flores, E., Herrero, A., Eds.; Caister Academic Press: Norfolk, UK, 2014; pp. 7–27. [Google Scholar]
- Maldener, I.; Summers, M.L.; Sukenik, A. Cellular differentiation in filamentous cyanobacteria. In The Cell Biology of Cyanobacteria; Flores, E., Herrero, A., Eds.; Caister Academic Press: Norfolk, UK, 2014; pp. 263–291. [Google Scholar]
- Narainsamy, K.; Cassier-Chauvat, C.; Junot, C.; Chauvat, F. High performance analysis of the cyanobacterial metabolism via liquid chromatography coupled to a LTQ-orbitrap mass spectrometer: Evidence that glucose reprograms the whole carbon metabolism and triggers oxidative stress. Metabolomics 2011, 9, 21–32. [Google Scholar] [CrossRef]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Chardonnet, S.; Sakr, S.; Cassier-Chauvat, C.; le Marechal, P.; Chauvat, F.; Lemaire, S.D.; Decottignies, P. First proteomic study of S-glutathionylation in cyanobacteria. J. Proteome Res. 2014. [Google Scholar] [CrossRef]
- Marteyn, B.; Sakr, S.; Farci, S.; Bedhomme, M.; Chardonnet, S.; Decottignies, P.; Lemaire, S.D.; Cassier-Chauvat, C.; Chauvat, F. The Synechocystis PCC6803 MerA-like enzyme operates in the reduction of both mercury and uranium under the control of the glutaredoxin 1 enzyme. J. Bacteriol. 2013, 195, 4138–4145. [Google Scholar] [CrossRef] [PubMed]
- Sakr, S.; Dutheil, J.; Saenkham, P.; Bottin, H.; Leplat, C.; Ortega-Ramos, M.; Aude, J.C.; Chapuis, V.; Guedeney, G.; Decottignies, P.; et al. The activity of the Synechocystis PCC6803 AbrB2 regulator of hydrogen production can be post-translationally controlled through glutathionylation. Int. J. Hydrog. Energy 2013, 38, 13547–13555. [Google Scholar] [CrossRef]
- Waldron, K.J.; Robinson, N.J. How do bacterial cells ensure that metalloproteins get the correct metal? Nat. Rev. Microbiol. 2009, 7, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kranzler, C.; Rudolf, M.; Keren, N.; Schleiff, E. Iron in cyanobacteria. Adv. Bot. Res. 2013, 65, 57–105. [Google Scholar]
- Blindauer, C.A. Zinc-handling in cyanobacteria: An update. Chem. Biodivers. 2008, 5, 1990–2013. [Google Scholar] [CrossRef] [PubMed]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar]
- Song, Q.; Li, J. Environmental effects of heavy metals derived from the e-waste recycling activities in China: A systematic review. Waste Manag. 2014, 34, 2587–2594. [Google Scholar] [CrossRef] [PubMed]
- Chetelat, J.; Amyot, M.; Arp, P.; Blais, J.M.; Depew, D.; Emmerton, C.A.; Evans, M.; Gamberg, M.; Gantner, N.; Girard, C.; et al. Mercury in freshwater ecosystems of the Canadian Arctic: Recent advances on its cycling and fate. Sci. Total Environ. 2014. [Google Scholar] [CrossRef]
- Gregoire, D.S.; Poulain, A.J. A little bit of light goes a long way: The role of phototrophs on mercury cycling. Metallomics 2014, 6, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. The Mismetallation of enzymes during oxidative stress. J. Biol. Chem. 2014, 289, 28121–28128. [Google Scholar] [CrossRef] [PubMed]
- Heindl, J.E.; Wang, Y.; Heckel, B.C.; Mohari, B.; Feirer, N.; Fuqua, C. Mechanisms and regulation of surface interactions and biofilm formation in Agrobacterium. Front. Plant Sci. 2014, 5, 176. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Colica, G.; Micheletti, E. Exopolysaccharide-producing cyanobacteria in heavy metal removal from water: Molecular basis and practical applicability of the biosorption process. Appl. Microbiol. Biotechnol. 2011, 92, 697–708. [Google Scholar]
- Jittawuttipoka, T.; Planchon, M.; Spalla, O.; Benzerara, K.; Guyot, F.; Cassier-Chauvat, C.; Chauvat, F. Multidisciplinary evidences that Synechocystis PCC6803 exopolysaccharides operate in cell sedimentation and protection against salt and metal stresses. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Planchon, M.; Jittawuttipoka, T.; Cassier-Chauvat, C.; Guyot, F.; Chauvat, F.; Spalla, O. Influence of exopolysaccharides on the electrophoretic properties of the model cyanobacterium Synechocystis. Colloids Surf. 2013, 110, 171–177. [Google Scholar] [CrossRef]
- Houot, L.; Floutier, M.; Marteyn, B.; Michaut, M.; Picciocchi, A.; Legrain, P.; Aude, J.C.; Cassier-Chauvat, C.; Chauvat, F. Cadmium triggers an integrated reprogramming of the metabolism of Synechocystis PCC6803, under the control of the Slr1738 regulator. BMC Genomics 2007, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Barras, F.; Fontecave, M. Cobalt stress in Escherichia coli and Salmonella enterica: Molecular bases for toxicity and resistance. Metallomics 2011, 3, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Zeyons, O.; Thill, A.; Chauvat, F.; Menguy, N.; Cassier-Chauvat, C.; Orear, C.; Daraspe, J.; Auffan, M.; Rose, J.; Spalla, O.; et al. Direct and indirect CeO2 nanoparticles toxicity for Escherichia coli and Synechocystis. Nanotoxicology 2009, 3, 284–295. [Google Scholar] [CrossRef]
- Planchon, M.; Jittawuttipoka, T.; Cassier-Chauvat, C.; Guyot, F.; Gelabert, A.; Benedetti, M.F.; Chauvat, F.; Spalla, O. Exopolysaccharides protect Synechocystis against the deleterious effects of Titanium dioxide nanoparticles in natural and artificial waters. J. Colloid Interface Sci. 2013, 405, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Von Moos, N.; Slaveykova, V.I. Oxidative stress induced by inorganic nanoparticles in bacteria and aquatic microalgae—State of the art and knowledge gaps. Nanotoxicology 2014, 8, 605–630. [Google Scholar]
- Turner, J.S.; Morby, A.P.; Whitton, B.A.; Gupta, A.; Robinson, N.J. Construction of Zn2+/Cd2+ hypersensitive cyanobacterial mutants lacking a functional metallothionein locus. J. Biol. Chem. 1993, 268, 4494–4498. [Google Scholar] [PubMed]
- Blindauer, C.A. Bacterial metallothioneins: Past, present, and questions for the future. J. Biol. Inorg. Chem. 2011, 16, 1011–1024. [Google Scholar] [CrossRef] [PubMed]
- Huckle, J.W.; Morby, A.P.; Turner, J.S.; Robinson, N.J. Isolation of a prokaryotic metallothionein locus and analysis of transcriptional control by trace-metal ions. Mol. Microbiol. 1993, 7, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Morby, A.P.; Turner, J.S.; Huckle, J.W.; Robinson, N.J. SmtB is a metal-dependent repressor of the cyanobacterial metallothionein gene smtA—Identification of a Zn inhibited DNA-protein complex. Nucleic Acids Res. 1993, 21, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Thelwell, C.; Robinson, N.J.; Turner-Cavet, J.S. An SmtB-like repressor from Synechocystis PCC6803 regulates a zinc exporter. Proc. Natl. Acad. Sci. USA 1998, 95, 10728–10733. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Dominguez, M.; Lopez-Maury, L.; Florencio, F.J.; Reyes, J.C. A gene cluster involved in metal homeostasis in the cyanobacterium Synechocystis sp. strain PCC6803. J. Bacteriol. 2000, 182, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhu, Y.; Song, Z.; Wang, J.; Zhang, W. An orphan response regulator Sll0649 involved in cadmium tolerance and metal homeostasis in photosynthetic Synechocystis sp. PCC6803. J. Proteomics 2014, 103, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Nagy, C.I.; Vass, I.; Rakhely, G.; Vass, I.Z.; Toth, A.; Duzs, A.; Peca, L.; Kruk, J.; Kos, P.B. Coregulated genes link sulfide:quinone oxidoreductase and arsenic metabolism in Synechocystis sp. strain PCC6803. J. Bacteriol. 2014, 196, 3430–3440. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Riego, A.M.; Lopez-Maury, L.; Florencio, F.J. Genomic responses to arsenic in the cyanobacterium Synechocystis sp. PCC6803. PLoS One 2014, 9, e96826. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Maury, L.; Florencio, F.J.; Reyes, J.C. Arsenic sensing and resistance system in the cyanobacterium Synechocystis sp. strain PCC6803. J. Bacteriol. 2003, 185, 5363–5371. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Haile, J.D.; Kennelly, P.J. An arsenate reductase from Synechocystis sp. strain PCC6803 exhibits a novel combination of catalytic characteristics. J. Bacteriol. 2003, 185, 6780–6789. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Chung, J.S.; Sutton, R.B.; Lee, J.S.; Lopez-Maury, L.; Lee, S.Y.; Florencio, F.J.; Lin, T.; Zabet-Moghaddam, M.; Wood, M.J.; et al. Redox, mutagenic and structural studies of the glutaredoxin/arsenate reductase couple from the cyanobacterium Synechocystis sp. PCC6803. Biochim. Biophys. Acta 2012, 1824, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Maury, L.; Sanchez-Riego, A.M.; Reyes, J.C.; Florencio, F.J. The glutathione/glutaredoxin system is essential for arsenate reduction in Synechocystis sp. strain PCC6803. J. Bacteriol. 2009, 191, 3534–3543. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.X.; Chen, J.; Qin, J.; Sun, G.X.; Rosen, B.P.; Zhu, Y.G. Biotransformation and volatilization of arsenic by three photosynthetic cyanobacteria. Plant Physiol. 2011, 156, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Wall, J.D.; Krumholz, L.R. Uranium reduction. Ann. Rev. Microbiol. 2006, 60, 149–166. [Google Scholar] [CrossRef]
- Lefebvre, D.D.; Kelly, D.; Budd, K. Biotransformation of Hg(II) by cyanobacteria. Appl. Environ. Microbiol. 2007, 73, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.C.; Pakrasi, H.B. Glutathione in Synechocystis 6803: A closer look into the physiology of a ΔgshB mutant. Plant Signal. Behav. 2011, 6, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Marteyn, B.; Domain, F.; Legrain, P.; Chauvat, F.; Cassier-Chauvat, C. The thioredoxin reductase-glutaredoxins-ferredoxin crossroad pathway for selenate tolerance in Synechocystis PCC6803. Mol. Microbiol. 2009, 71, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Thorsen, M.; Jacobson, T.; Vooijs, R.; Navarrete, C.; Bliek, T.; Schat, H.; Tamas, M.J. Glutathione serves an extracellular defence function to decrease arsenite accumulation and toxicity in yeast. Mol. Microbiol. 2012, 84, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Kong, X.L. Glutathione: A key component of the cytoplasmic labile iron pool. Biometals 2011, 24, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.C.; Pakrasi, H.B. Essential role of glutathione in acclimation to environmental and redox perturbations in the cyanobacterium Synechocystis sp. PCC6803. Plant Physiol. 2010, 154, 1672–1685. [Google Scholar] [CrossRef] [PubMed]
- Zaffagnini, M.; Bedhomme, M.; Marchand, C.H.; Morisse, S.; Trost, P.; Lemaire, S.D. Redox regulation in photosynthetic organisms: Focus on glutathionylation. Antioxid. Redox Signal. 2012, 16, 567–586. [Google Scholar] [CrossRef] [PubMed]
- Lillig, C.H.; Berndt, C. Glutaredoxins in thiol/disulfide exchange. Antioxid. Redox Signal. 2012, 18, 1654–1665. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N.; Couturier, J.; Johnson, M.K.; Jacquot, J.P. Glutaredoxins: Roles in iron homeostasis. Trends Biochem. Sci. 2010, 35, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Zaffagnini, M.; Bedhomme, M.; Lemaire, S.D.; Trost, P. The emerging roles of protein glutathionylation in chloroplasts. Plant Sci. 2012, 185–186, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Riego, A.M.; Lopez-Maury, L.; Florencio, F.J. Glutaredoxins are essential for stress adaptation in the cyanobacterium Synechocystis sp. PCC6803. Front. Plant Sci. 2013, 4, 428. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, Q.; Zhang, L.; Li, H.; Cui, Y.; Wu, Q. Identification of novel targets of cyanobacterial glutaredoxin. Arch. Biochem. Biophys. 2007, 458, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Florencio, F.J.; Perez-Perez, M.E.; Lopez-Maury, L.; Mata-Cabana, A.; Lindahl, M. The diversity and complexity of the cyanobacterial thioredoxin systems. Photosynth. Res. 2006, 89, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Picciocchi, A.; Saguez, C.; Boussac, A.; Cassier-Chauvat, C.; Chauvat, F. CGFS-type monothiol glutaredoxins from the cyanobacterium Synechocystis PCC6803 and other evolutionary distant model organisms possess a glutathione-ligated [2Fe–2S] cluster. Biochemistry 2007, 46, 15018–15026. [Google Scholar] [CrossRef] [PubMed]
- Iwema, T.; Picciocchi, A.; Traore, D.A.; Ferrer, J.L.; Chauvat, F.; Jacquamet, L. Structural basis for delivery of the intact [Fe2S2] cluster by monothiol glutaredoxin. Biochemistry 2009, 48, 6041–6043. [Google Scholar] [CrossRef] [PubMed]
- Lillig, C.H.; Berndt, C.; Holmgren, A. Glutaredoxin systems. Biochim. Biophys. Acta 2008, 1780, 1304–1317. [Google Scholar] [CrossRef] [PubMed]
- Herrero, E.; Belli, G.; Casa, C. Structural and functional diversity of glutaredoxins in yeast. Curr. Protein Pept. Sci. 2010, 11, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Cassier-Chauvat, C.; Chauvat, F. Function and regulation of ferredoxins in the cyanobacterium, Synechocystis PCC6803: Recent advances. Life 2014, 4, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Gutekunst, K.; Chen, X.; Schreiber, K.; Kaspar, U.; Makam, S.; Appel, J. The bidirectional NiFe-hydrogenase in Synechocystis sp. PCC6803 is reduced by flavodoxin and ferredoxin and is essential under mixotrophic, nitrate-limiting conditions. J. Biol. Chem. 2014, 289, 1930–1937. [Google Scholar] [CrossRef] [PubMed]
- Mustila, H.; Allahverdiyeva, Y.; Isojarvi, J.; Aro, E.M.; Eisenhut, M. The bacterial-type [4Fe–4S] ferredoxin 7 has a regulatory function under photooxidative stress conditions in the cyanobacterium Synechocystis sp. PCC6803. Biochim. Biophys. Acta 2014, 1837, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Behrenfeld, M.J.; Milligan, A.J. Photophysiological expressions of iron stress in phytoplankton. Ann. Rev. Mar. Sci. 2013, 5, 217–246. [Google Scholar] [CrossRef] [PubMed]
- Fillat, M.F. The FUR (ferric uptake regulator) superfamily: Diversity and versatility of key transcriptional regulators. Arch. Biochem. Biophys. 2014, 546, 41–52. [Google Scholar] [CrossRef]
- Cheng, D.; He, Q.F. PfsR is a key regulator of iron homeostasis in Synechocystis PCC6803. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Krynicka, V.; Tichy, M.; Krafl, J.; Yu, J.; Kana, R.; Boehm, M.; Nixon, P.J.; Komenda, J. Two essential FtsH proteases control the level of the Fur repressor during iron deficiency in the cyanobacterium Synechocystis sp. PCC6803. Mol. Microbiol. 2014, 94, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ishizuka, T.; Katayama, M.; Kanehisa, M.; Bhattacharyya-Pakrasi, M.; Pakrasi, H.B.; Ikeuchi, M. Response to oxidative stress involves a novel peroxiredoxin gene in the unicellular cyanobacterium Synechocystis sp. PCC6803. Plant Cell Physiol. 2004, 45, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Singh, A.K.; McIntyre, L.M.; Sherman, L.A. Differential gene expression in response to hydrogen peroxide and the putative PerR regulon of Synechocystis sp. strain PCC6803. J. Bacteriol. 2004, 186, 3331–3345. [Google Scholar] [CrossRef] [PubMed]
- Garcin, P.; Delalande, O.; Zhang, J.Y.; Cassier-Chauvat, C.; Chauvat, F.; Boulard, Y. A transcriptional-switch model for Slr1738-controlled gene expression in the cyanobacterium Synechocystis. BMC Struct. Biol. 2012, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.T.; Maldonado, M.T. Biogeochemistry of cadmium and its release to the environment. Met. Ions Life Sci. 2013, 11, 31–62. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cassier-Chauvat, C.; Chauvat, F. Responses to Oxidative and Heavy Metal Stresses in Cyanobacteria: Recent Advances. Int. J. Mol. Sci. 2015, 16, 871-886. https://doi.org/10.3390/ijms16010871
Cassier-Chauvat C, Chauvat F. Responses to Oxidative and Heavy Metal Stresses in Cyanobacteria: Recent Advances. International Journal of Molecular Sciences. 2015; 16(1):871-886. https://doi.org/10.3390/ijms16010871
Chicago/Turabian StyleCassier-Chauvat, Corinne, and Franck Chauvat. 2015. "Responses to Oxidative and Heavy Metal Stresses in Cyanobacteria: Recent Advances" International Journal of Molecular Sciences 16, no. 1: 871-886. https://doi.org/10.3390/ijms16010871
APA StyleCassier-Chauvat, C., & Chauvat, F. (2015). Responses to Oxidative and Heavy Metal Stresses in Cyanobacteria: Recent Advances. International Journal of Molecular Sciences, 16(1), 871-886. https://doi.org/10.3390/ijms16010871