
Apigenin-7-Glycoside Prevents LPS-Induced Acute Lung Injury via Downregulation of Oxidative Enzyme Expression and Protein Activation through Inhibition of MAPK Phosphorylation


 ,
,
Abstract
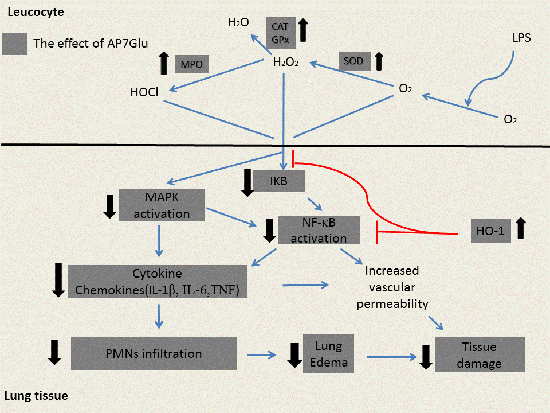
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
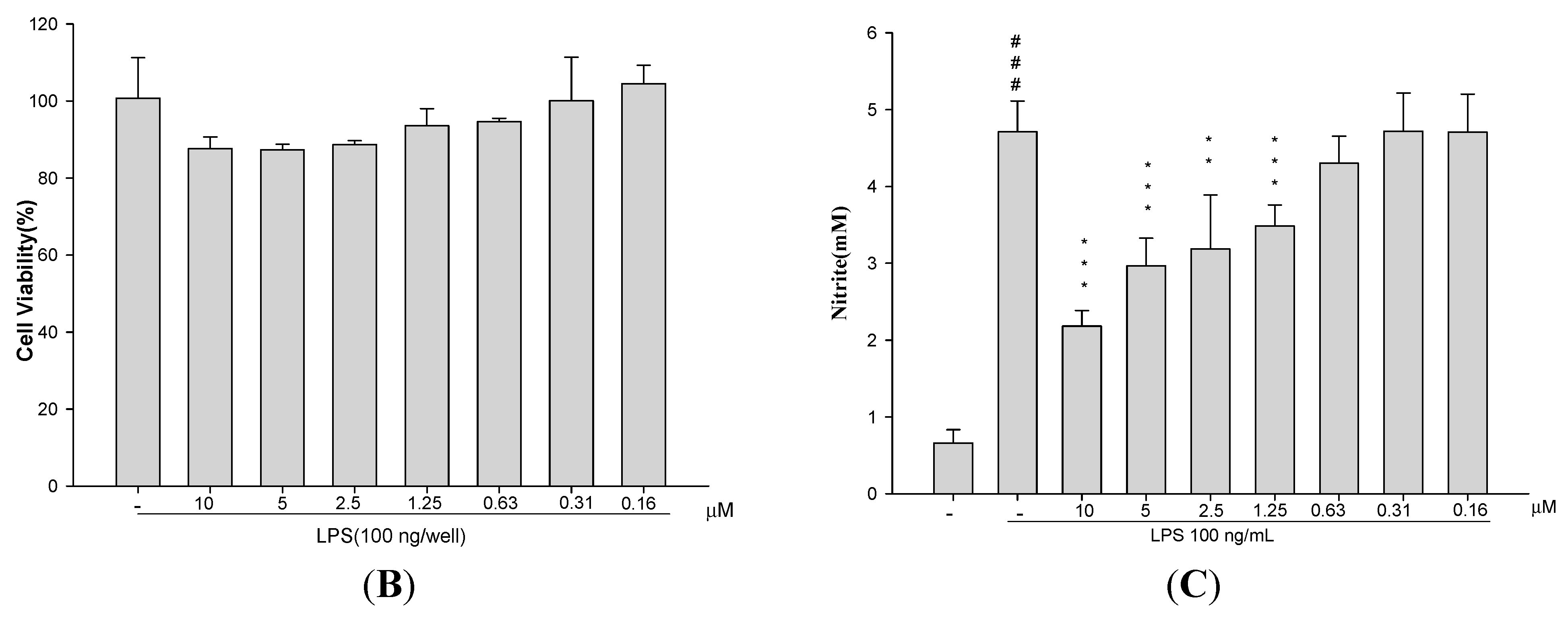
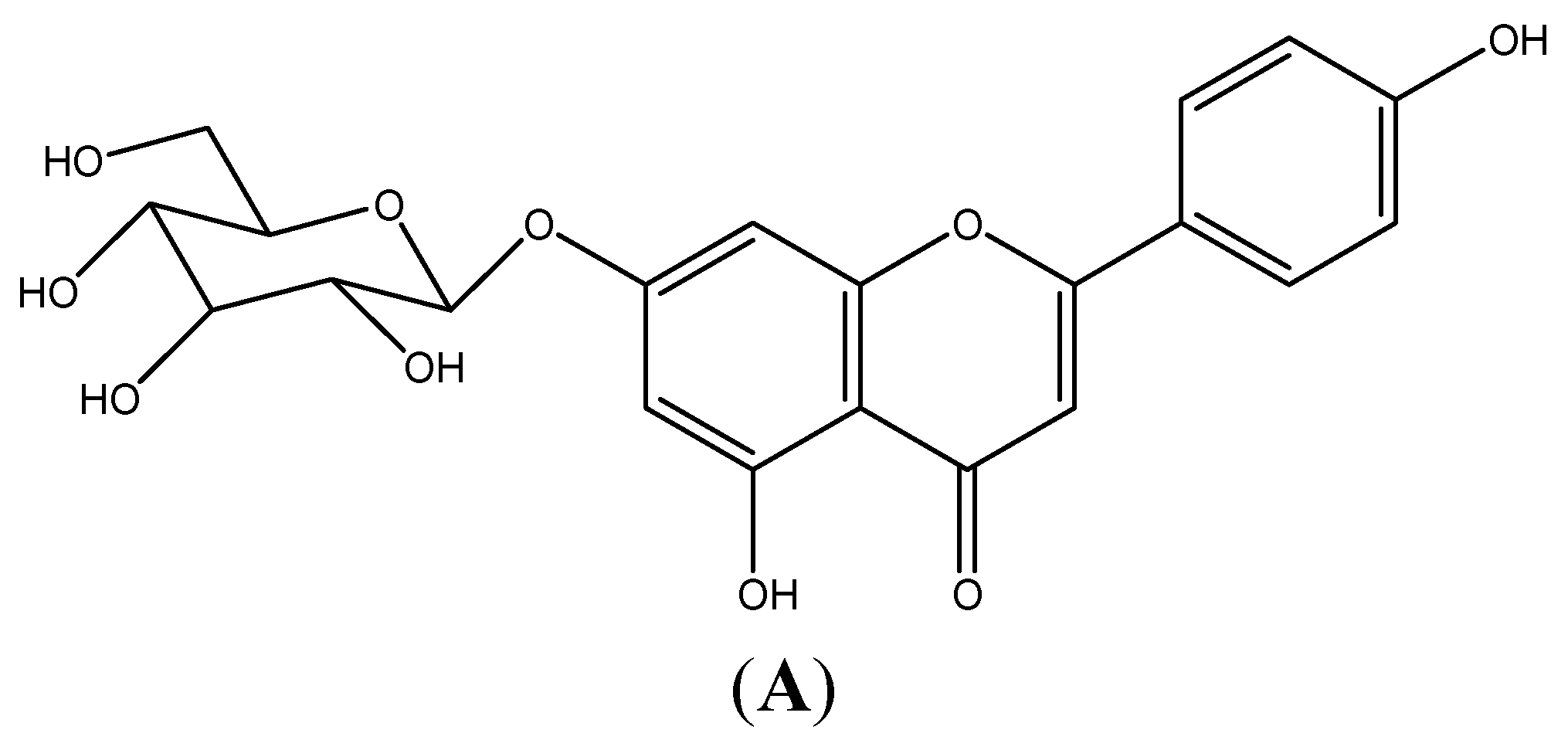
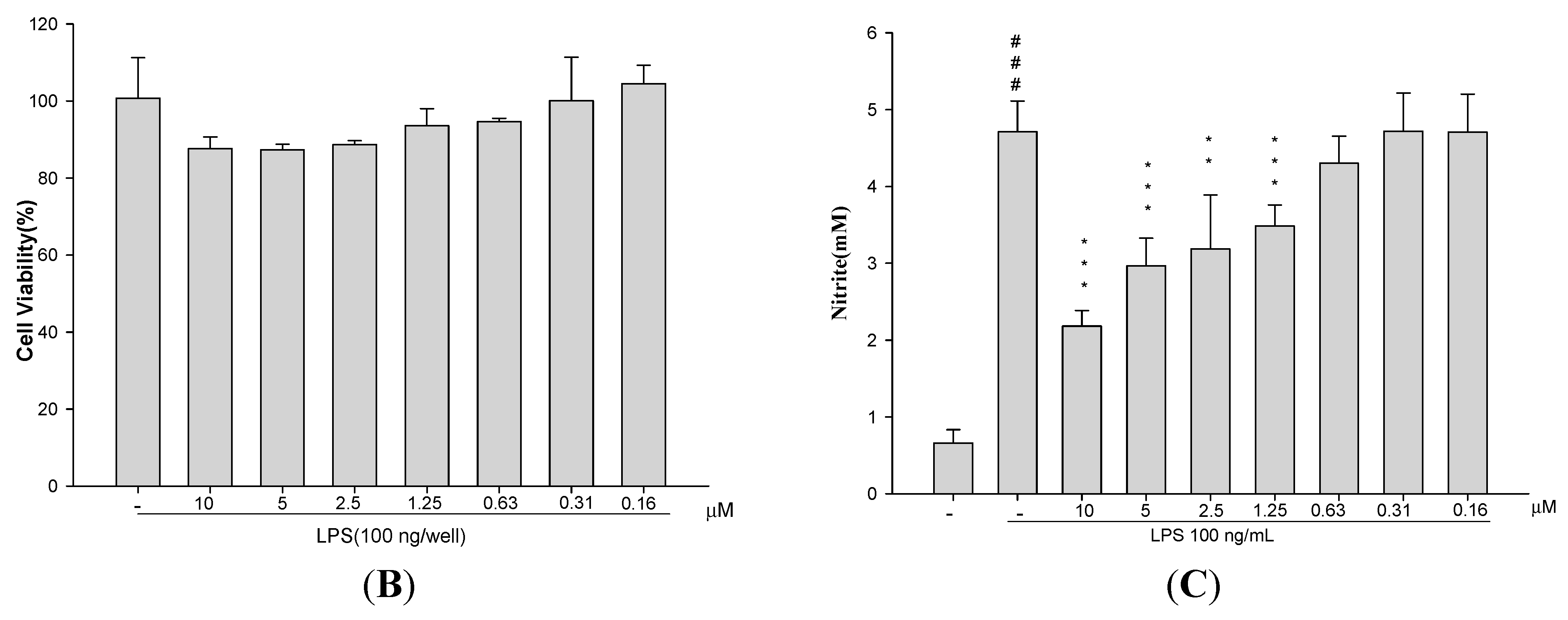
2.1. Cytotoxicity
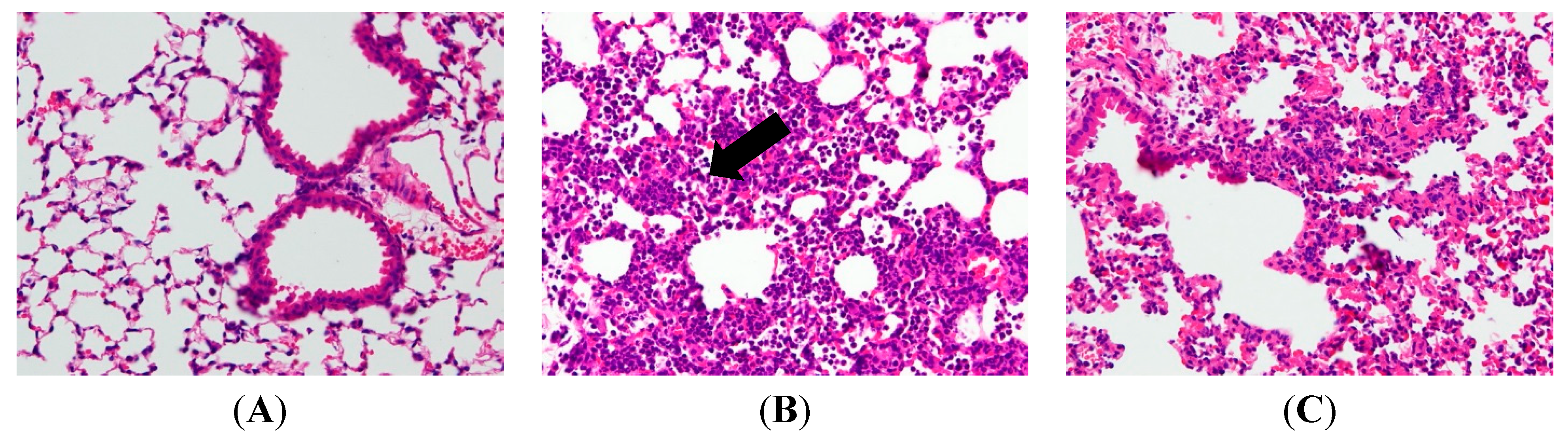
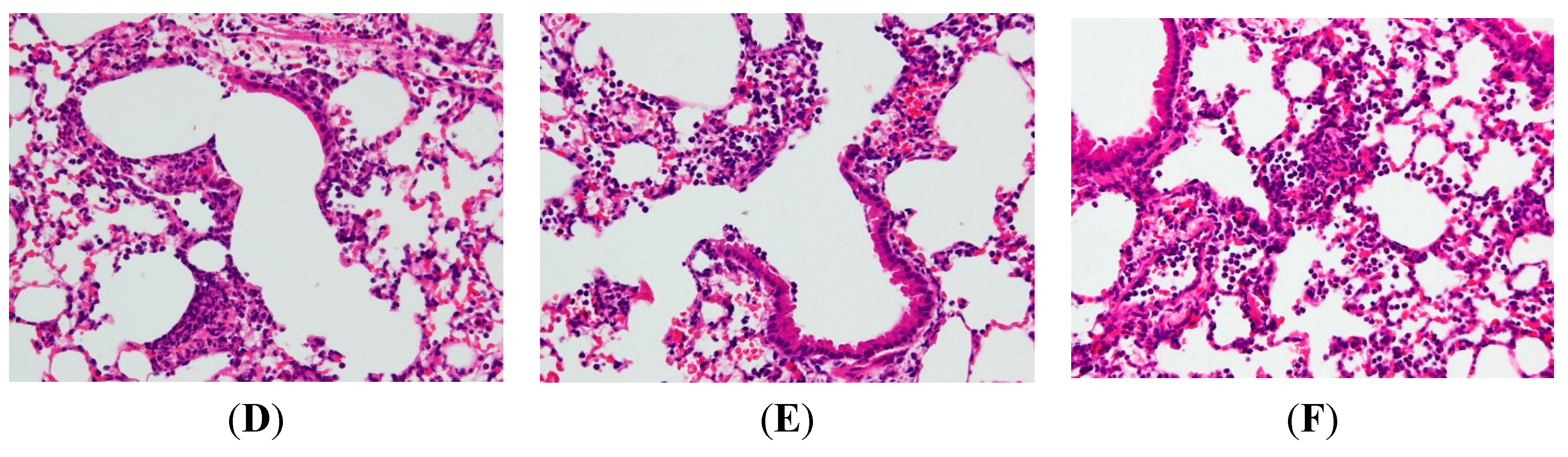
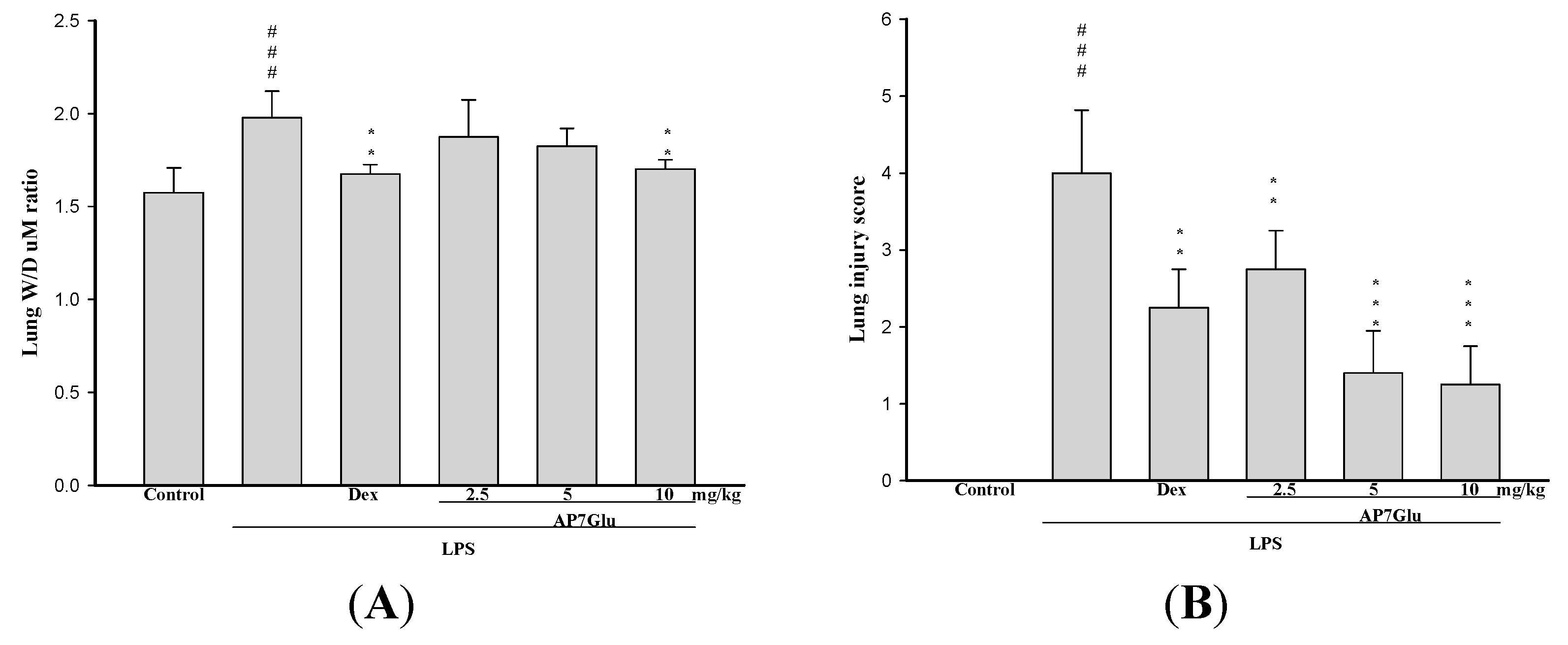
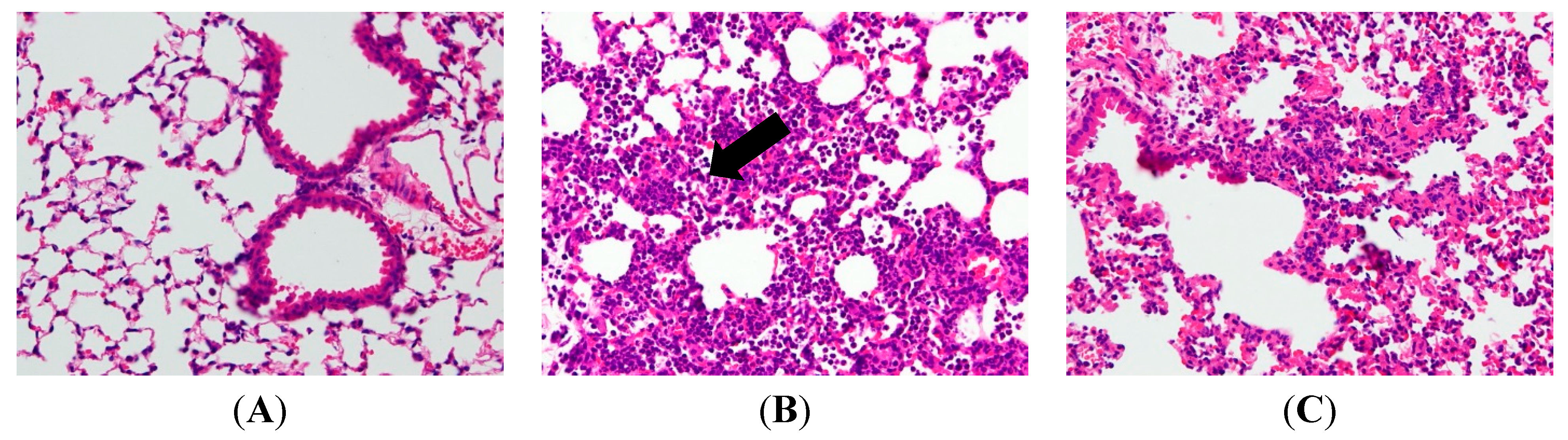
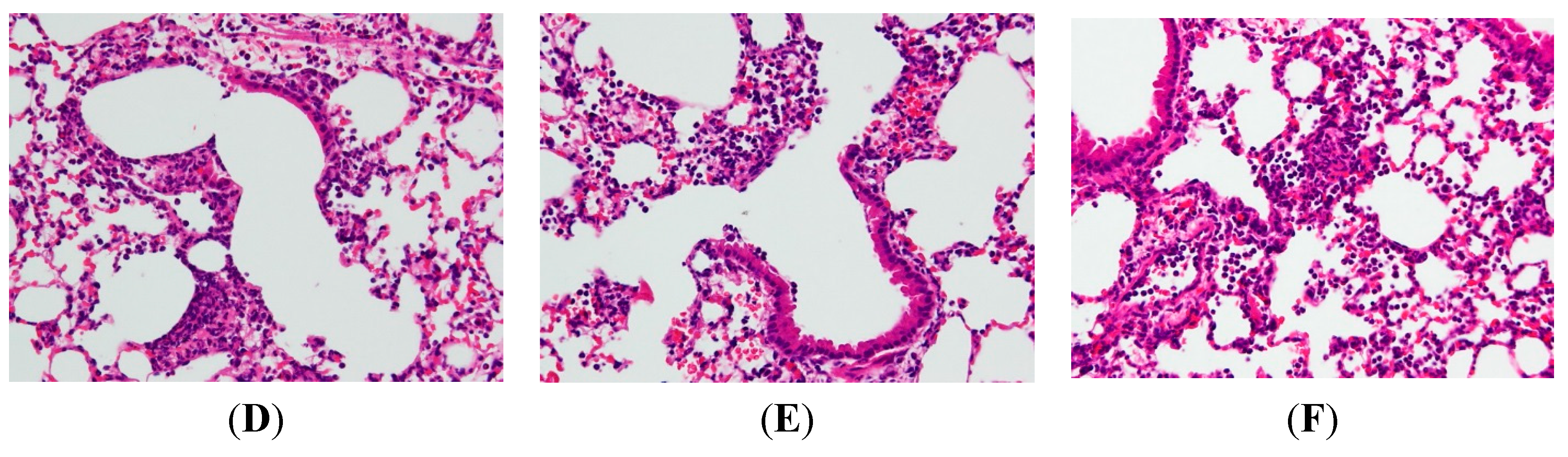
2.2. AP7Glu Attenuates Pulmonary Inflammation in LPS-Induced Acute Lung Injury (ALI)
2.3. AP7Glu Attenuates Pulmonary Edema in LPS-Induced Acute Lung Injury (ALI)
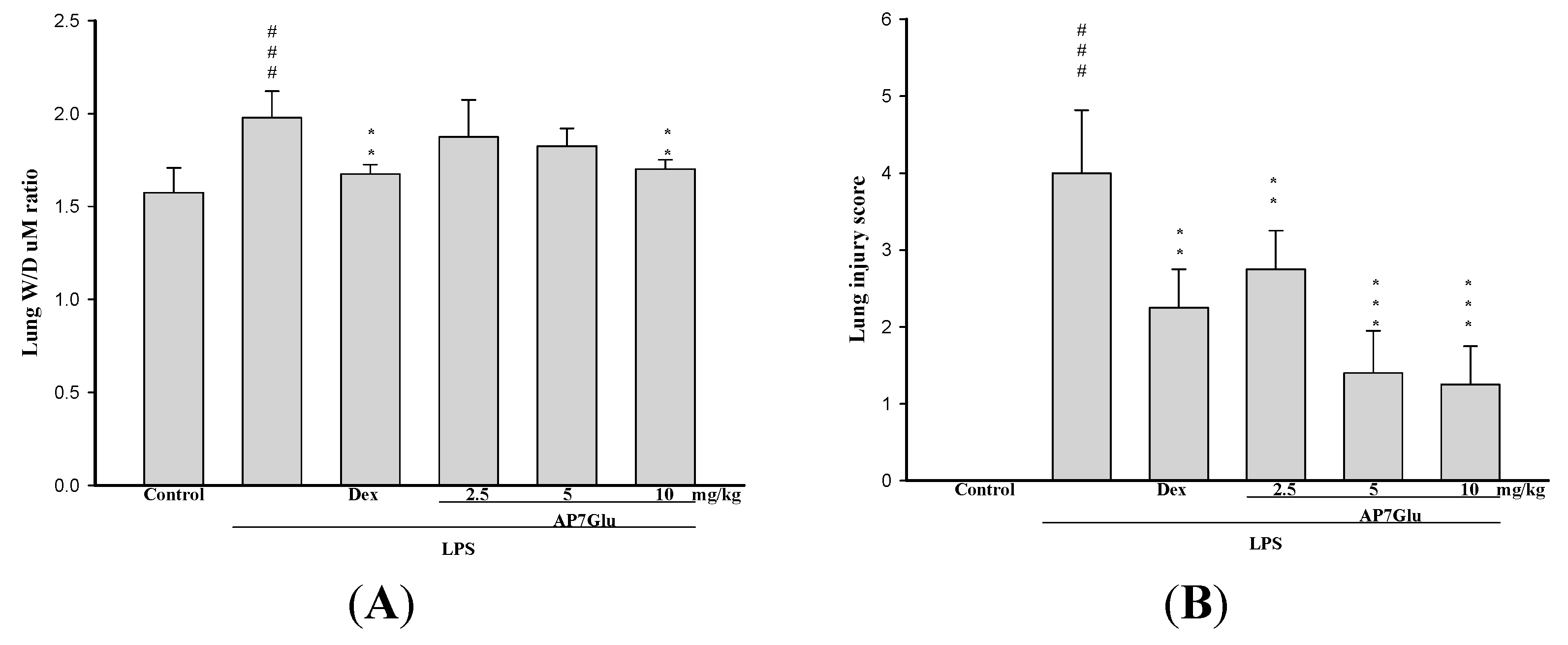
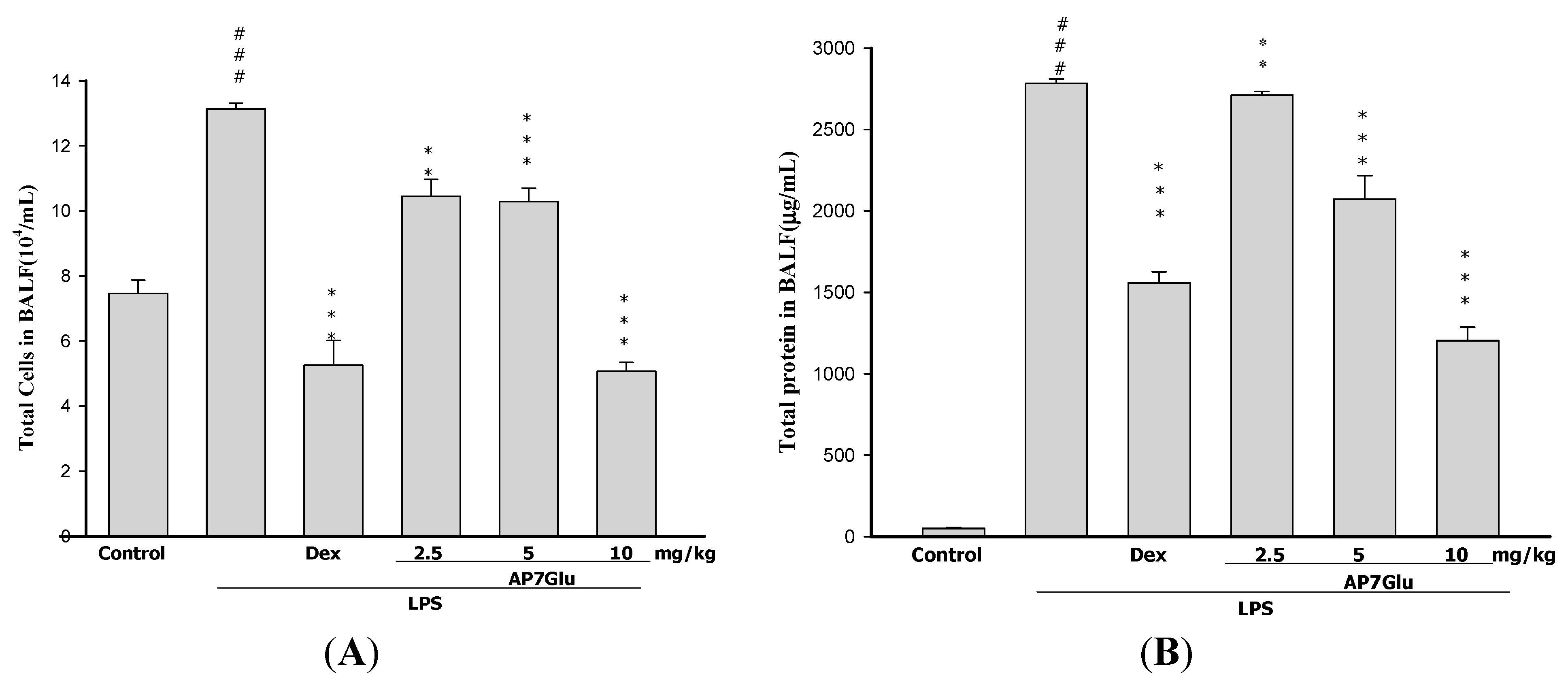
2.4. AP7Glu Reduces Cellular Counts and Proteins in BALF

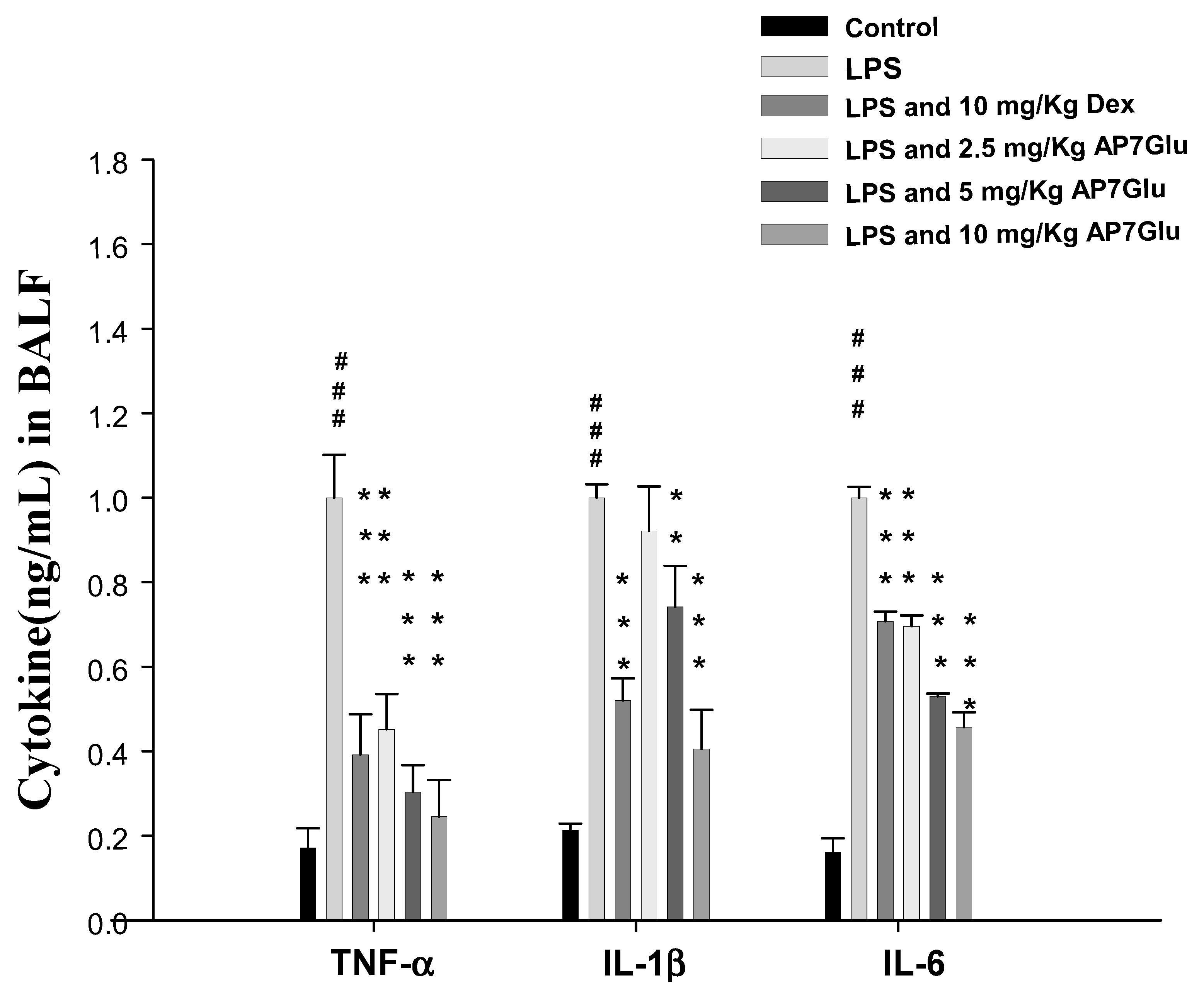
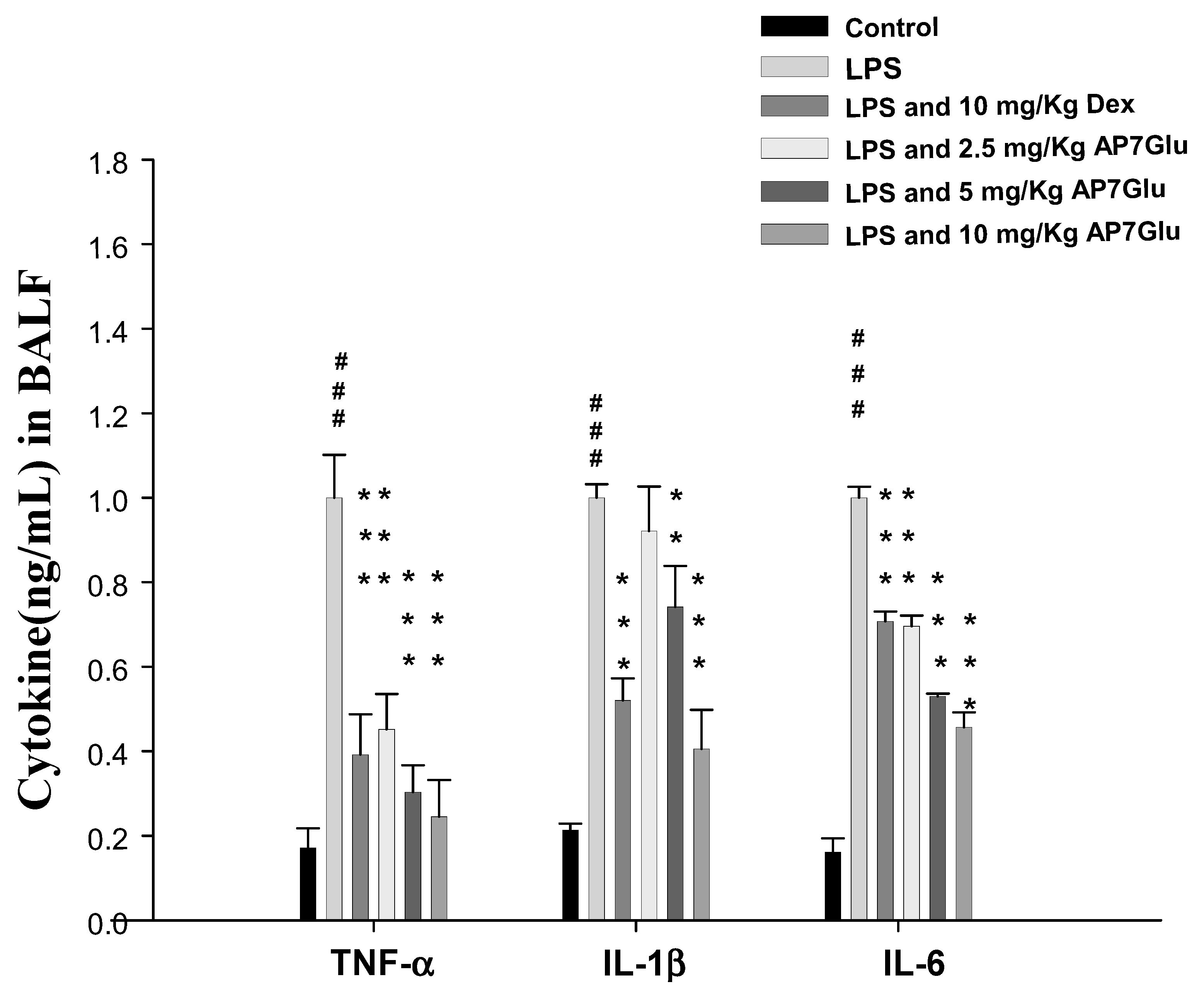
2.5. AP7Glu Downregulates TNF-α, IL-6, and IL-1β in BALF
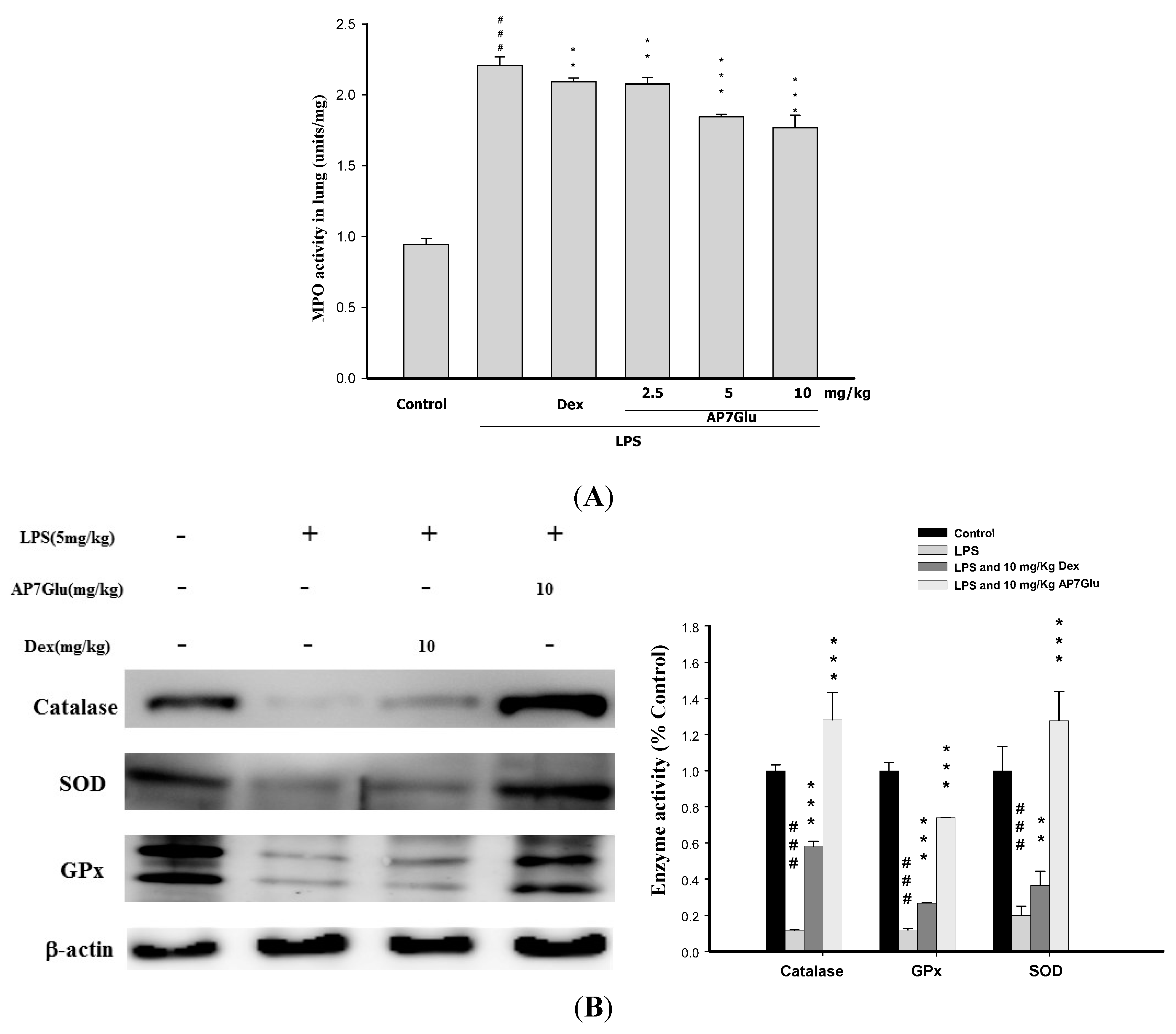
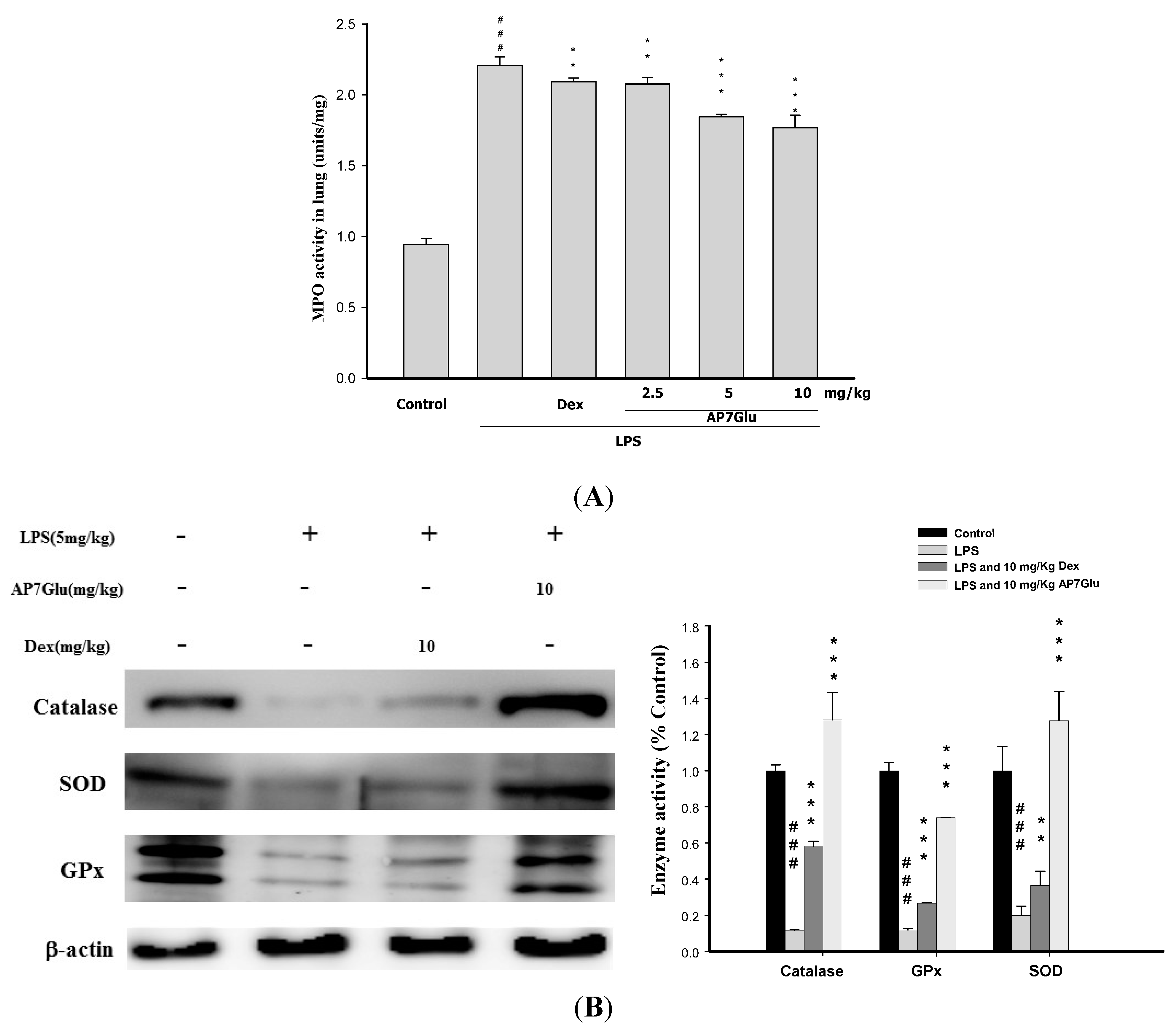
2.6. Effects of AP7Glu on MPO Activity and Antioxidative Enzymes in LPS Induced ALI
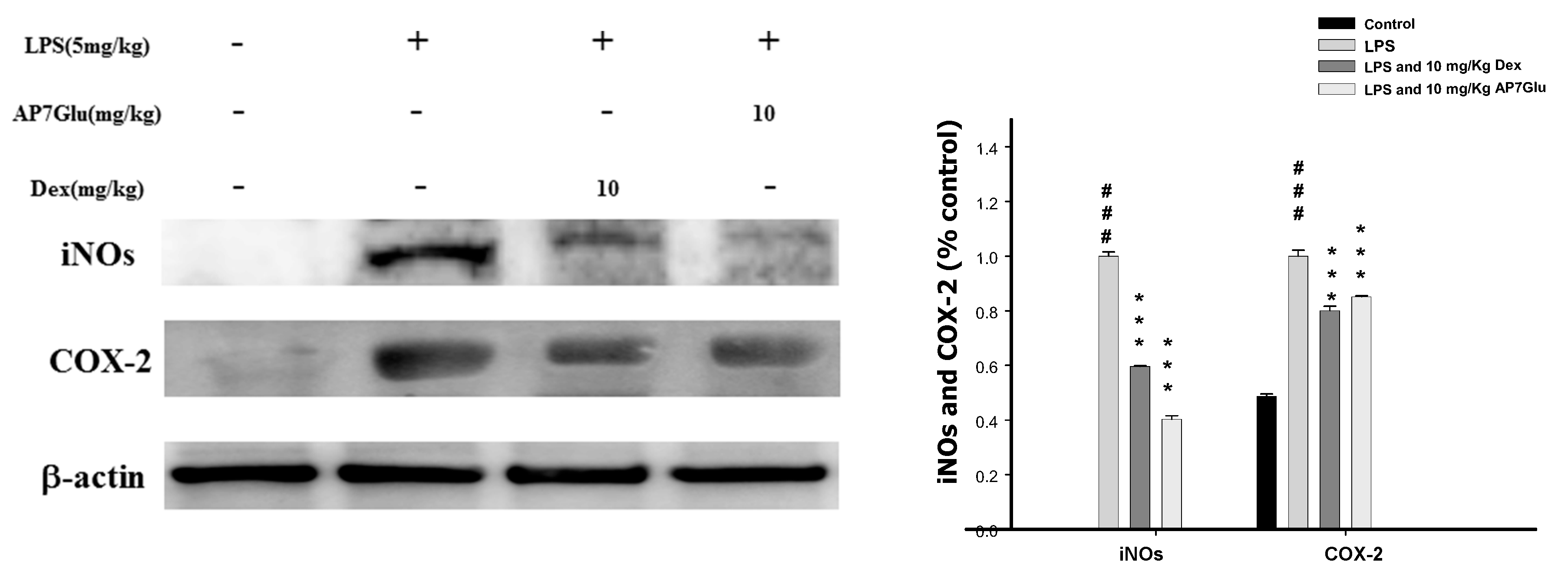
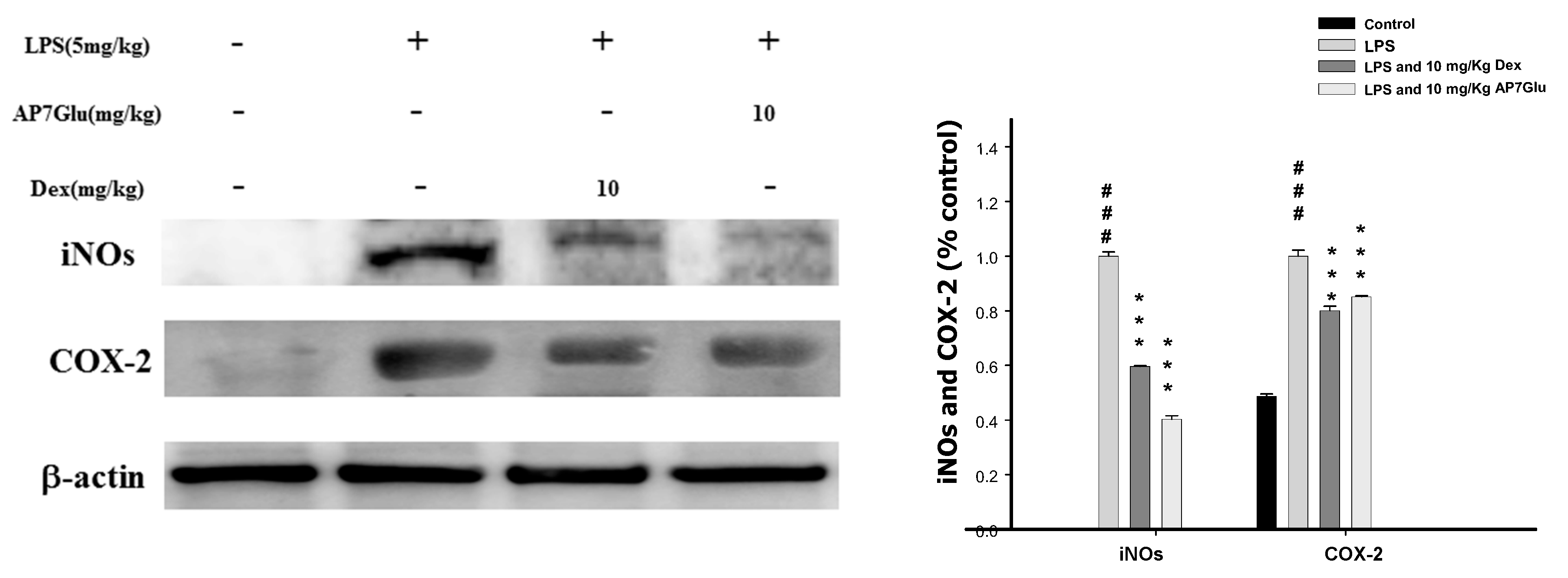
2.7. Inhibition of LPS Induced iNOs and COX-2 Proteins by AP7Glu in Lung Tissue
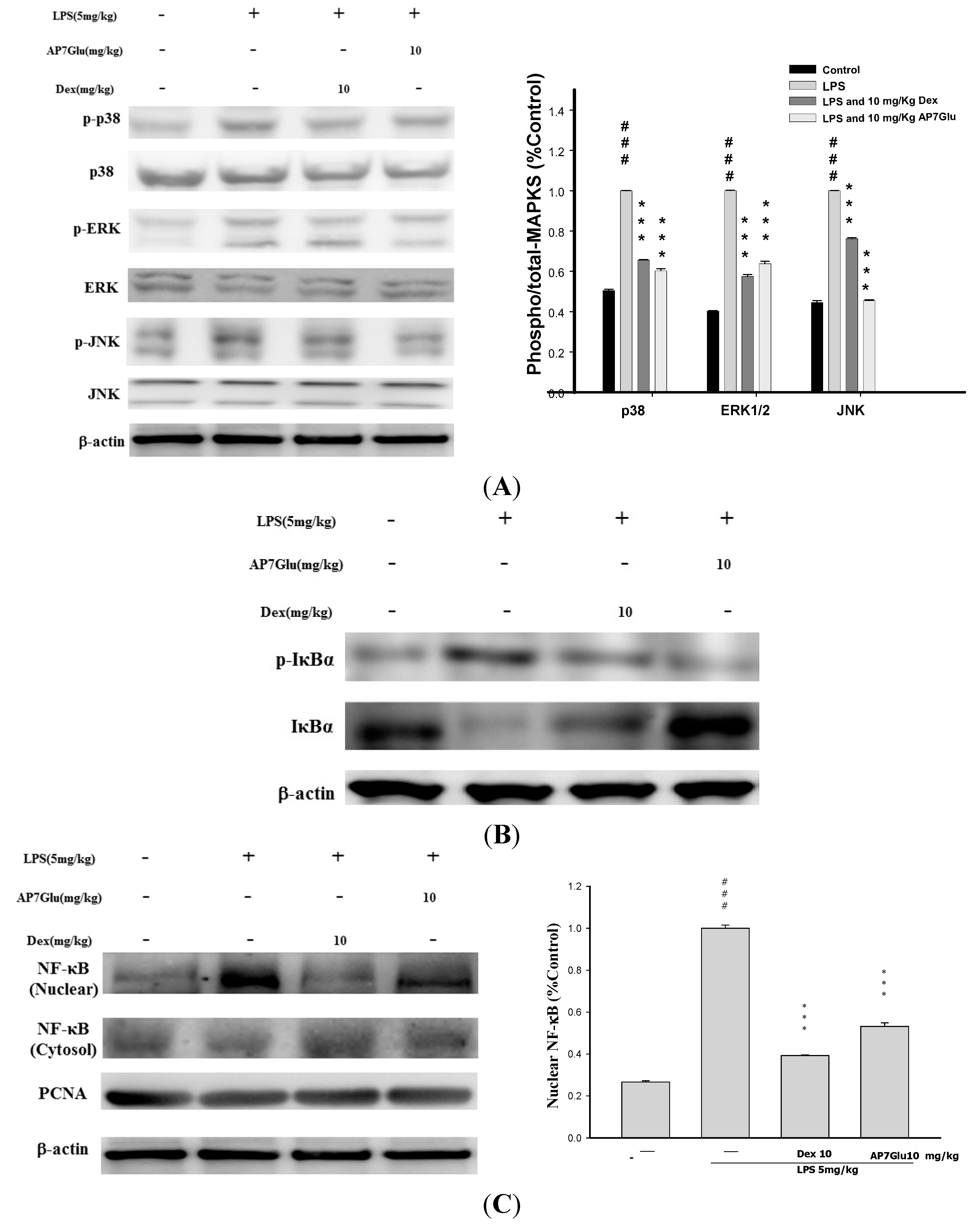
2.8. Effects of AP7Glu on MAPK, IκB and NF-κB Activation in LPS Induced ALI
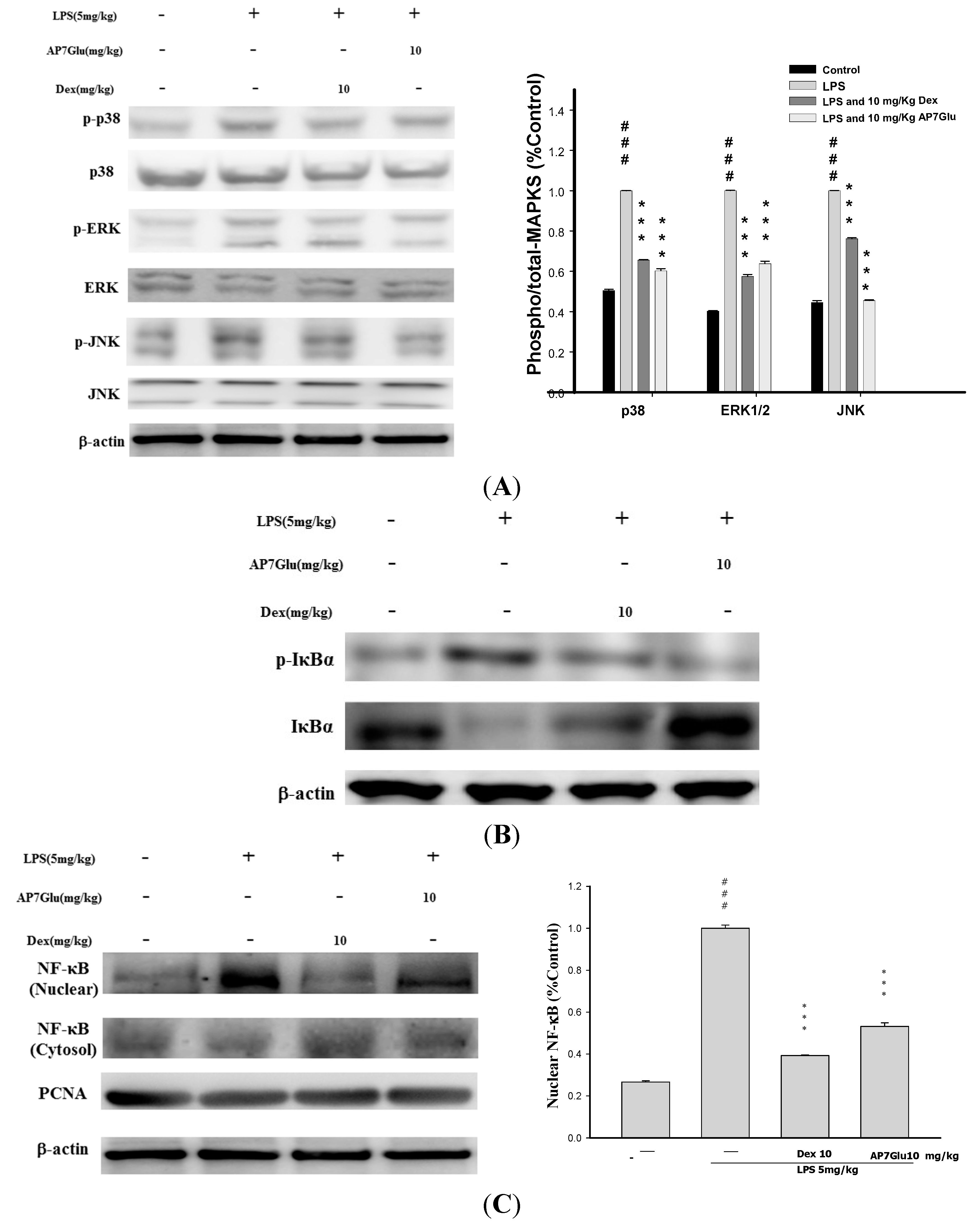
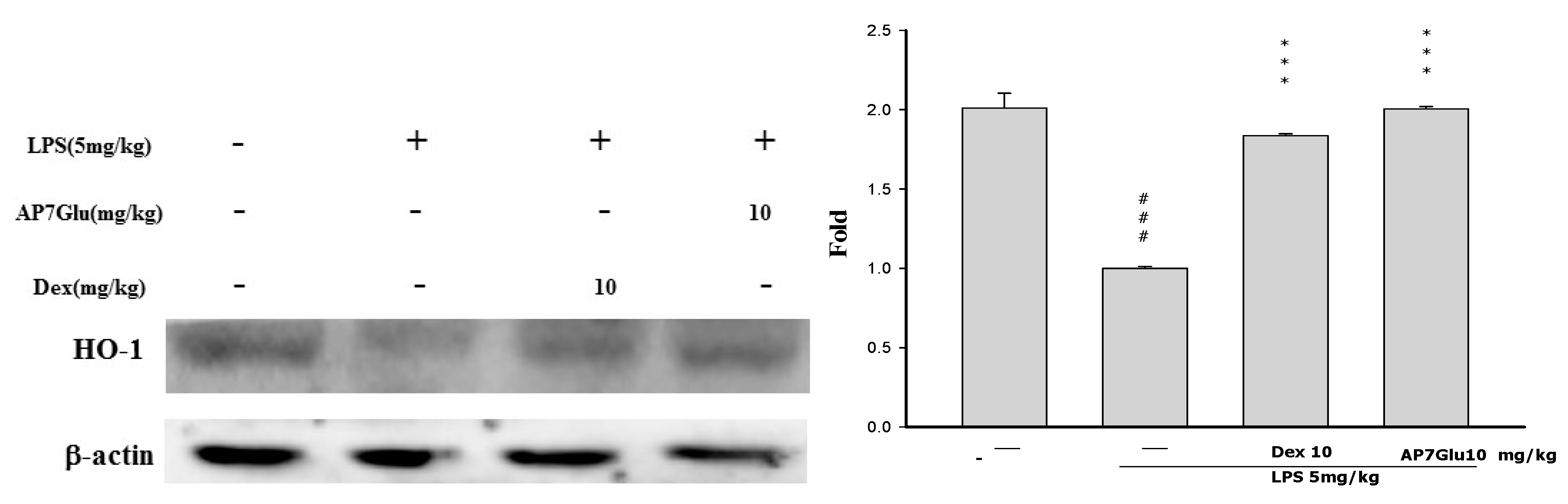
2.9. Effects of AP7Glu on HO-1 Expression in LPS-Induced ALI
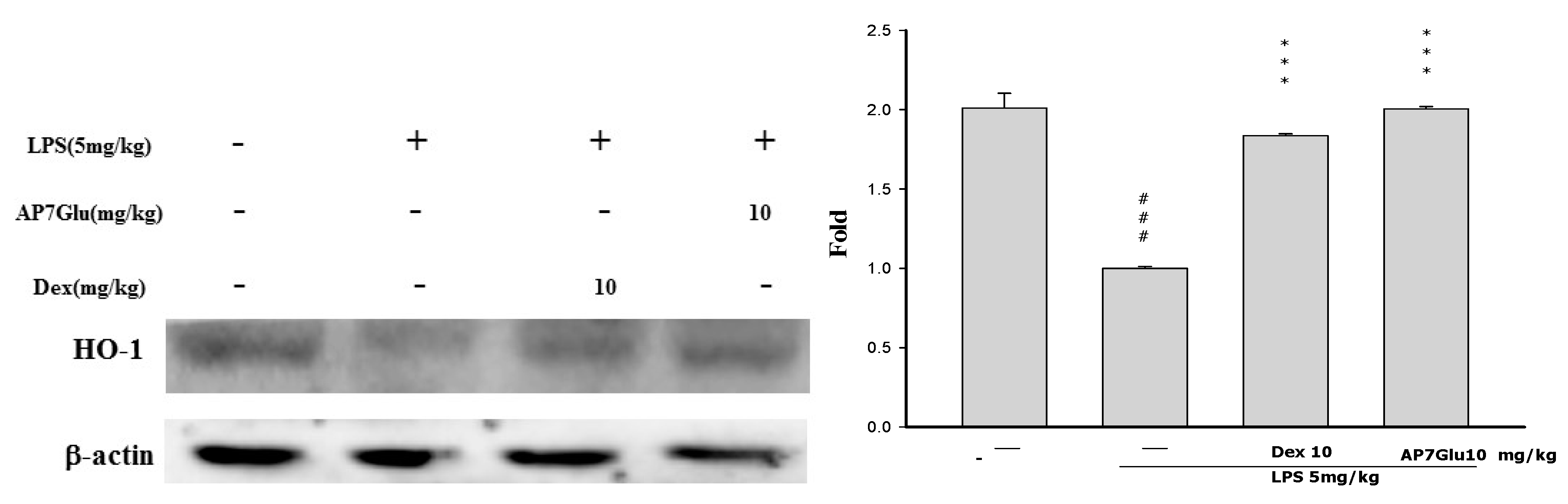
3. Discussion
4. Experimental Section
4.1. Cell Culture
4.2. Cytotoxicity and NO Production
4.3. Animals
4.4. Model of LPS Induced ALI
4.5. Bronchoalveolar Lavage Fluid (BALF), Total Cell Count and Protein Analysis
4.6. TNF-α, IL-6, and IL-1β Cytokines in BALF
4.7. Myeloperoxidase (MPO) Activity Assay
4.8. Lung Wet/Dry Weight Ratio
4.9. H&E Staining
4.10. Western Blot Analysis of Lung Tissue
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Annapurna, A.; Ansari, M.A.; Manjunath, P.M. Partial role of multiple pathways in infarct size limiting effect of quercetin and rutin against cerebral ischemia-reperfusion injury in rats. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 491–500. [Google Scholar] [PubMed]
- Huang, K.L.; Chen, C.S.; Hsu, C.W.; Li, M.H.; Chang, H.; Tsai, S.H.; Chu, S.J. Therapeutic effects of baicalin on lipopolysaccharide-induced acute lung injury in rats. Am. J. Chin. Med. 2008, 36, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Lu, T.H.; Su, C.C.; Lay, I.S.; Lin, H.Y.; Fang, K.M.; Ho, T.J.; Chen, K.L.; Su, Y.C.; Chiang, W.C.; et al. Lotus leaf (Nelumbo nucifera) and its active constituents prevent inflammatory responses in macrophages via JNK/NF-κB signaling pathway. Am. J. Chin. Med. 2014, 42, 869–889. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.L.; Lin, Y.C.; Wang, H.M.; Chou, T.C. Baicalein, an active component of Scutellaria baicalensis, protects against lipopolysaccharide-induced acute lung injury in rats. J. Ethnopharmacol. 2014, 153, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Park, S.W.; Kim, H.J.; Kwak, J.H.; Lee, D.U.; Chang, K.C. Dehydrocostuslactone inhibits LPS-induced inflammation by p38MAPK-dependent induction of hemeoxygenase-1 in vitro and improves survival of mice in CLP-induced sepsis in vivo. Int. Immunopharmacol. 2014, 22, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Du, L.; Zhao, L.; Shang, R.; Liu, D.; Jing, Q.; Liang, J.; Ren, Y. The total alkaloids of Aconitum tanguticum protect against lipopolysaccharide-induced acute lung injury in rats. J. Ethnopharmacol. 2014, 155, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xie, J.Y.; Li, H.; Zhang, Y.Y.; Cao, J.; Cheng, Z.H.; Chen, D.F. Viola yedoensis liposoluble fraction ameliorates lipopolysaccharide-induced acute lung injury in mice. Am. J. Chin. Med. 2012, 40, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Choi, R.J.; Shehzad, O.; Kim, H.P.; Islam, M.N.; Choi, J.S.; Kim, Y.S. Molecular mechanism of capillarisin-mediated inhibition of MyD88/TIRAP inflammatory signaling in in vitro and in vivo experimental models. J. Ethnopharmacol. 2013, 145, 626–637. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, N.; Shukla, S.; Srivastava, J.K.; Gupta, S. Chamomile: An anti-inflammatory agent inhibits inducible nitric oxide synthase expression by blocking RelA/p65 activity. Int. J. Mol. Med. 2010, 26, 935–940. [Google Scholar] [PubMed]
- Bhaskaran, N.; Srivastava, J.K.; Shukla, S.; Gupta, S. Chamomile confers protection against hydrogen peroxide-induced toxicity through activation of Nrf2-mediated defense response. Phytother. Res. 2013, 27, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, H.; Nie, Y.C.; Chen, J.L.; Su, W.W.; Li, P.B. Naringin attenuates acute lung injury in LPS-treated mice by inhibiting NF-κB pathway. Int. Immunopharmacol. 2011, 11, 1606–1612. [Google Scholar] [CrossRef] [PubMed]
- Araz, O.; Demirci, E.; Yilmazel Ucar, E.; Calik, M.; Pulur, D.; Karaman, A.; Yayla, M.; Altun, E.; Halici, Z.; Akgun, M. Comparison of reducing effect on lung injury of dexamethasone and bosentan in acute lung injury: An experimental study. Multidiscip. Respir. Med. 2013, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Kim, D.H.; Park, M.H.; Kim, M.K.; Kim, N.D.; Kim, C.M.; Tanaka, T.; Yokozawa, T.; Chung, H.Y.; Moon, H.R. Chinese prescription Kangen-karyu and Salviae Miltiorrhizae Radix improve age-related oxidative stress and inflammatory response through the PI3K/Akt or MAPK pathways. Am. J. Chin. Med. 2014, 42, 987–1005. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Suangkaew, N.; Morinaga, O.; Kariyazono, H.; Oiso, S.; Shoyama, Y. Eriobotryae folium extract suppresses LPS-induced iNOS and COX-2 expression by inhibition of NF-κB and MAPK activation in murine macrophages. Am. J. Chin. Med. 2010, 38, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.; Milbradt, R. Skin anti-inflammatory activity of apigenin-7-glucoside in rats. Arzneim.-Forsch. 1993, 43, 370–372. [Google Scholar]
- Lewin, G.; Maciuk, A.; Moncomble, A.; Cornard, J.P. Enhancement of the water solubility of flavone glycosides by disruption of molecular planarity of the aglycone moiety. J. Nat. Prod. 2013, 76, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, N.; Shukla, S.; Kanwal, R.; Srivastava, J.K.; Gupta, S. Induction of heme oxygenase-1 by chamomile protects murine macrophages against oxidative stress. Life Sci. 2012, 90, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Hosakote, Y.M.; Liu, T.; Castro, S.M.; Garofalo, R.P.; Casola, A. Respiratory syncytial virus induces oxidative stress by modulating antioxidant enzymes. Am. J. Respir. Cell Mol. Biol. 2009, 41, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Barquero, L.; Villegas, I.; Sanchez-Calvo, J.M.; Talero, E.; Sanchez-Fidalgo, S.; Motilva, V.; Alarcon de la Lastra, C. Curcumin, a Curcuma longa constituent, acts on MAPK p38 pathway modulating COX-2 and iNOS expression in chronic experimental colitis. Int. Immunopharmacol. 2007, 7, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Yin, H.; Fang, M.; Xiang, Y.; Yuan, C.L.; Zheng, G.Y.; Yang, H.; Xiong, P.; Chen, G.; Gong, F.L.; et al. Heme oxygenase-1 upregulation significantly inhibits TNF-α and Hmgb1 releasing and attenuates lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2008, 8, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Pawlowska, E.; Wysokinski, D.; Tokarz, P.; Piastowska-Ciesielska, A.; Szczepanska, J.; Blasiak, J. Dexamethasone and 1,25-dihydroxyvitamin D3 reduce oxidative stress-related DNA damage in differentiating osteoblasts. Int. J. Mol. Sci. 2014, 15, 16649–16664. [Google Scholar] [CrossRef] [PubMed]
- Niu, N.; Li, B.; Hu, Y.; Li, X.; Li, J.; Zhang, H. Protective effects of scoparone against lipopolysaccharide-induced acute lung injury. Int. Immunopharmacol. 2014, 23, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.H.; Lee, J.H.; Kwon, D.Y. Apigenin inhibits release of inflammatory mediators by blocking the NF-κB activation pathways in the HMC-1 cells. Immunopharmacol. Immunotoxicol. 2011, 33, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Hung, M.C. Beyond NF-κB activation: Nuclear functions of IκB kinase alpha. J. Biomed. Sci. 2013, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Koppula, S.; Kim, W.J.; Jiang, J.; Shim, D.W.; Oh, N.H.; Kim, T.J.; Kang, T.B.; Lee, K.H. Carpesium macrocephalum attenuates lipopolysaccharide-induced inflammation in macrophages by regulating the NF-κ/IκB-α, Akt, and STAT signaling pathways. Am. J. Chin. Med. 2013, 41, 927–943. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Wu, C.F.; Huang, H.W.; Jao, Y.C.; Yen, G.C. Naturally occurring flavonoids attenuate high glucose-induced expression of proinflammatory cytokines in human monocytic THP-1 cells. Mol. Nutr. Food Res. 2009, 53, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Ferrell, J.E., Jr.; Bhatt, R.R. Mechanistic studies of the dual phosphorylation of mitogen-activated protein kinase. J. Biol. Chem. 1997, 272, 19008–19016. [Google Scholar] [CrossRef] [PubMed]
- Kuan, Y.H.; Lin, R.H.; Tsao, L.T.; Chen, Y.L.; Tzeng, C.C.; Wang, J.P. Inhibition of phospholipase D activation by CYL-26z in formyl peptide-stimulated neutrophils involves the blockade of RhoA activation. Biochem. Pharmacol. 2005, 70, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.C.; Peng, W.H.; Chiu, T.H.; Lai, S.C.; Lee, C.Y. Anti-inflammatory effects of Scoparia dulcis L. and betulinic acid. Am. J. Chin. Med. 2011, 39, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Simeonova, R.; Kondeva-Burdina, M.; Vitcheva, V.; Krasteva, I.; Manov, V.; Mitcheva, M. Protective effects of the apigenin-O/C-diglucoside saponarin from Gypsophila trichotoma on carbone tetrachloride-induced hepatotoxicity in vitro/in vivo in rats. Phytomedicine 2014, 21, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Victor, V.M.; Rocha, M.; Esplugues, J.V.; de la Fuente, M. Role of free radicals in sepsis: Antioxidant therapy. Curr. Pharm. Des. 2005, 11, 3141–3158. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Choi, H.J.; Lee, D.S.; Oh, H.; Kim, Y.C.; Moon, J.Y.; Park, W.H.; Park, S.D.; Kim, J.E. Amomum Tsao-ko suppresses lipopolysaccharide-induced inflammatory responses in RAW264.7 macrophages via Nrf2-dependent heme oxygenase-1 expression. Am. J. Chin. Med. 2014, 42, 1229–1244. [Google Scholar] [CrossRef] [PubMed]
- Yayeh, T.; Hong, M.; Jia, Q.; Lee, Y.C.; Kim, H.J.; Hyun, E.; Kim, T.W.; Rhee, M.H. Pistacia chinensis inhibits NO production and upregulates HO-1 induction via PI-3K/Akt pathway in LPS stimulated macrophage cells. Am. J. Chin. Med. 2012, 40, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Takashima, K.; Matsushima, M.; Hashimoto, K.; Nose, H.; Sato, M.; Hashimoto, N.; Hasegawa, Y.; Kawabe, T. Protective effects of intratracheally administered quercetin on lipopolysaccharide-induced acute lung injury. Respir. Res. 2014, 15, 150. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zhang, H.; Meng, X.; Wang, F.; Wang, P. Aloe-emodin from rhubarb (Rheum rhabarbarum) inhibits lipopolysaccharide-induced inflammatory responses in RAW264.7 macrophages. J. Ethnopharmacol. 2014, 153, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, H.; Yang, T.; Ye, Y.; Shan, J.; Yin, Z.; Luo, L. Chlorogenic acid protects mice against lipopolysaccharide-induced acute lung injury. Injury 2010, 41, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Yang, J.J.; Yang, M.L.; Li, Y.C.; Kuan, Y.H. Rutin decreases lipopolysaccharide-induced acute lung injury via inhibition of oxidative stress and the MAPK-NF-κB pathway. Free Radic. Biol. Med. 2014, 69, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Bani, D.; Masini, E.; Bello, M.G.; Bigazzi, M.; Sacchi, T.B. Relaxin protects against myocardial injury caused by ischemia and reperfusion in rat heart. Am. J. Pathol. 1998, 152, 1367–1376. [Google Scholar] [PubMed]
- Zhou, E.; Li, Y.; Wei, Z.; Fu, Y.; Lei, H.; Zhang, N.; Yang, Z.; Xie, G. Schisantherin A protects lipopolysaccharide-induced acute respiratory distress syndrome in mice through inhibiting NF-κB and MAPKs signaling pathways. Int. Immunopharmacol. 2014, 22, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Parsey, M.V.; Tuder, R.M.; Abraham, E. Neutrophils are major contributors to intraparenchymal lung IL-1 beta expression after hemorrhage and endotoxemia. J. Immunol. 1998, 160, 1007–1013. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.-C.; Ho, Y.-L.; Hsieh, W.-T.; Huang, S.-S.; Chang, Y.-S.; Huang, G.-J. Apigenin-7-Glycoside Prevents LPS-Induced Acute Lung Injury via Downregulation of Oxidative Enzyme Expression and Protein Activation through Inhibition of MAPK Phosphorylation. Int. J. Mol. Sci. 2015, 16, 1736-1754. https://doi.org/10.3390/ijms16011736
Li K-C, Ho Y-L, Hsieh W-T, Huang S-S, Chang Y-S, Huang G-J. Apigenin-7-Glycoside Prevents LPS-Induced Acute Lung Injury via Downregulation of Oxidative Enzyme Expression and Protein Activation through Inhibition of MAPK Phosphorylation. International Journal of Molecular Sciences. 2015; 16(1):1736-1754. https://doi.org/10.3390/ijms16011736
Chicago/Turabian StyleLi, Kun-Cheng, Yu-Ling Ho, Wen-Tsong Hsieh, Shyh-Shyun Huang, Yuan-Shiun Chang, and Guan-Jhong Huang. 2015. "Apigenin-7-Glycoside Prevents LPS-Induced Acute Lung Injury via Downregulation of Oxidative Enzyme Expression and Protein Activation through Inhibition of MAPK Phosphorylation" International Journal of Molecular Sciences 16, no. 1: 1736-1754. https://doi.org/10.3390/ijms16011736
APA StyleLi, K.-C., Ho, Y.-L., Hsieh, W.-T., Huang, S.-S., Chang, Y.-S., & Huang, G.-J. (2015). Apigenin-7-Glycoside Prevents LPS-Induced Acute Lung Injury via Downregulation of Oxidative Enzyme Expression and Protein Activation through Inhibition of MAPK Phosphorylation. International Journal of Molecular Sciences, 16(1), 1736-1754. https://doi.org/10.3390/ijms16011736