T Lymphocyte Antigen 4-Modified Dendritic Cell Therapy for Asthmatic Mice Guided by the CCR7 Chemokine Receptor

Abstract
:1. Introduction
2. Results
2.1. Quality Control of Recombinant Adenoviruses (rAdVs)
2.2. Therapeutic Dendritic Cell (DC) Modification
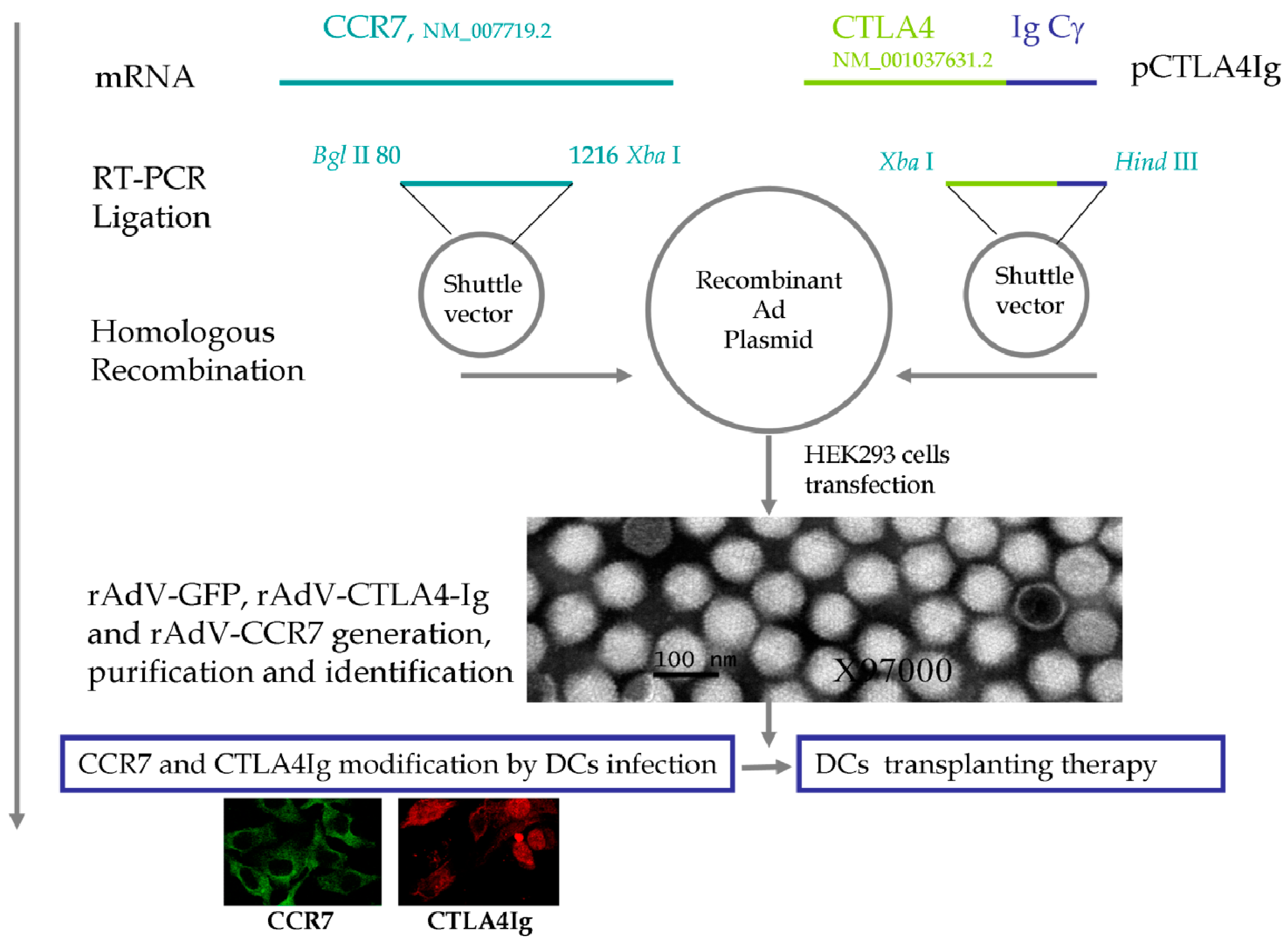

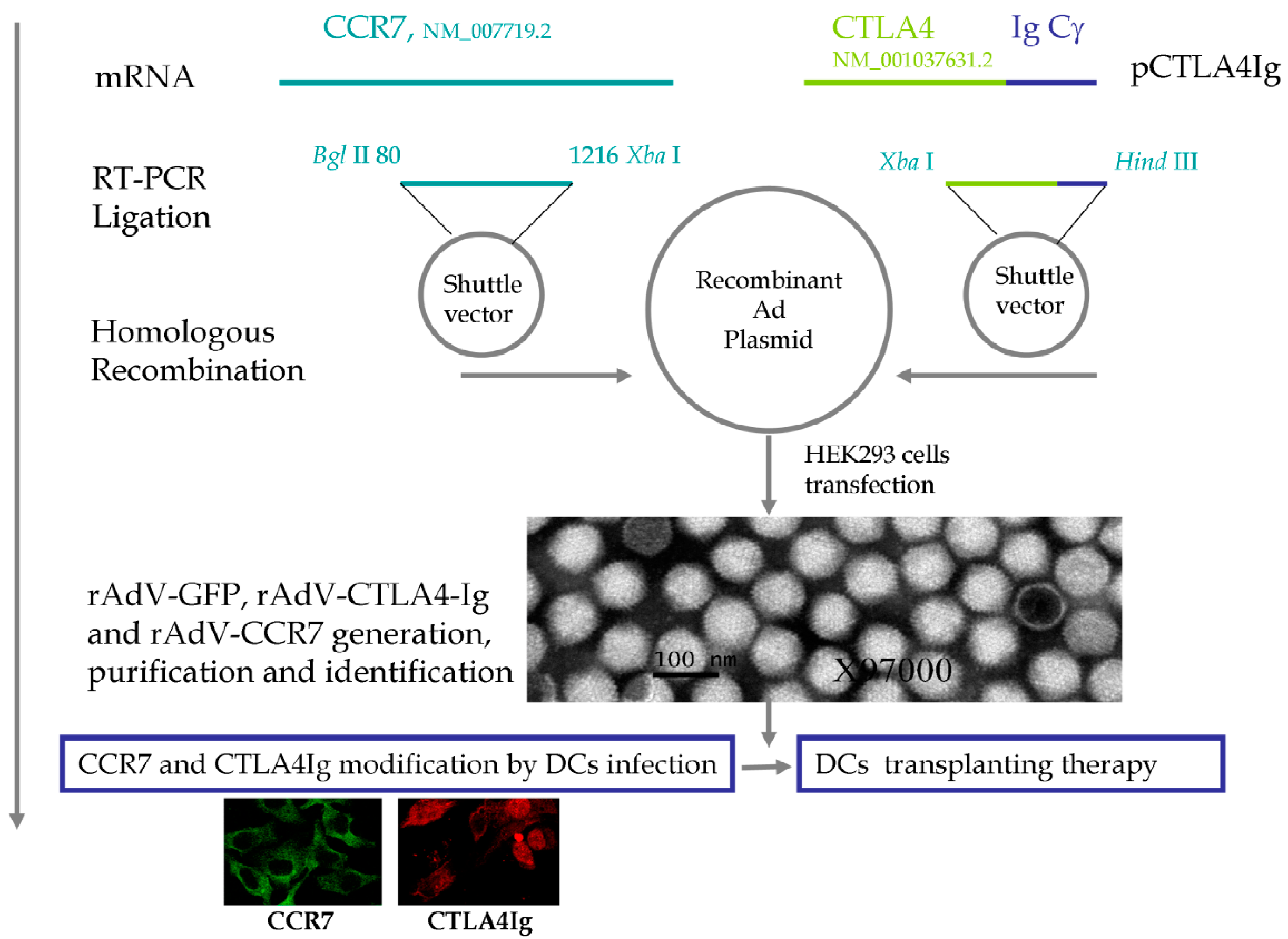{kind=link}
{kind=link}
{kind=link}
| Cellular Localization | CTLA4Ig (%) | CCR7 (%) | Both CTLA4Ig and CCR7 (%) |
|---|---|---|---|
| DC surface | |||
| rAdV-CTLA4Ig and rAdV-CCR7 | 10.86 ± 1.03 | 56.99 ± 1.42 | 10.04 ± 0.32 |
| rAdV-GFP | 0.00 ± 0.01 | 11.82 ± 0.73 | 0.00 ± 0.00 |
| OVA | 0.00 ± 0.00 | 10.19 ± 0.41 | 0.00 ± 0.01 |
| DC cytoplasm | |||
| rAdV-CTLA4Ig and rAdV-CCR7 | 80.00 ± 5.34 | 90.00 ± 4.52 | 75.00 ± 5.12 |
| rAdV-GFP | 0.00 ± 0.01 | 0.00 ± 0.01 | 0.00 ± 0.00 |
2.3. Surface Expression of DC Activation Markers after in Vitro Modification
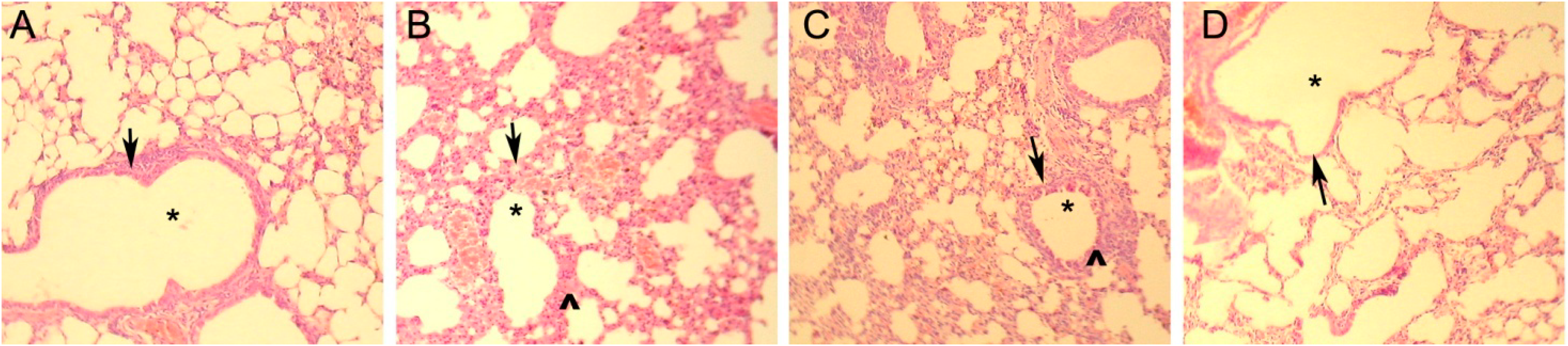
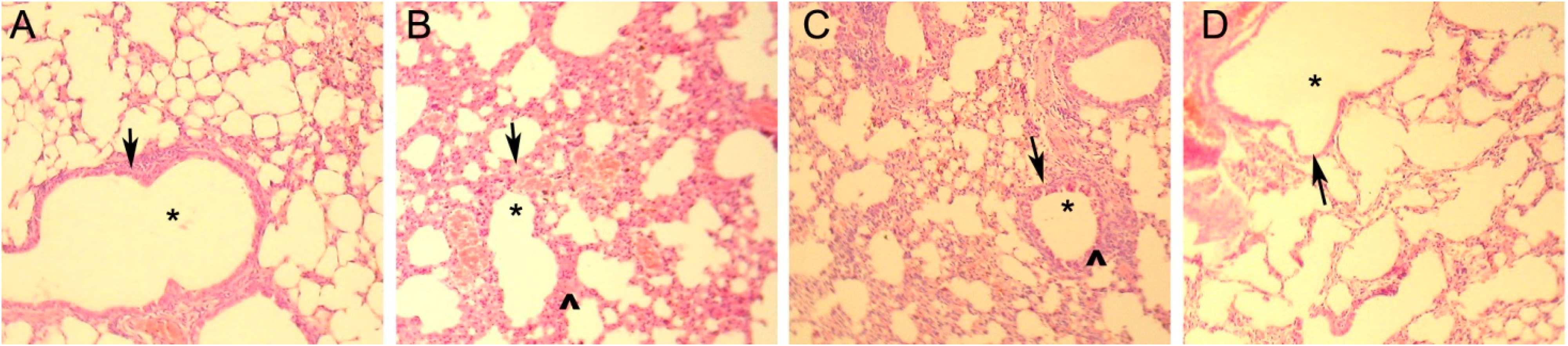
2.4. Mouse Asthma Model and Modified DC Therapy
| DC Activation Markers | GM-CSF and IL-4 | OVA | rAdV-GFP | rAdV-CTLA4Ig and rAdV-CCR7 |
|---|---|---|---|---|
| Induction duration (days) | 7 | 2 | 2 | 2 |
| CD40 (%) | 3.27 ± 0.21 | 4.43 ± 0.11 | 51.98 ± 4.55 | 19.59 ± 1.52 *,# |
| CD80 (%) | 63.74 ± 2.35 | 93.52 ± 5.22 | 96.41 ± 5.01 | 66.51 ± 2.21 *,# |
| CD86 (%) | 19.89 ± 1.10 | 30.87 ± 1.01 | 95.08 ± 4.66 | 36.90 ± 3.01 * |
| MHC II (%) | 52.50 ± 2.32 | 85.40 ± 2.33 | 98.56 ± 6.12 | 96.69 ± 6.01 |
| Symptoms and Pathology | Control | Asthma | rAdV-GFP | rAdV-CTLA4Ig and rAdV-CCR7 |
|---|---|---|---|---|
| (n = 8) | (n = 8) | (n = 8) | (n = 8) | |
| Asthma manifestations | ||||
| Dysphoria or asthenia | No | Yes | Yes | improved |
| Nodding with breathing | No | Yes | No | improved |
| Camponotus erect | No | Yes | No | improved |
| Forelimb shrinkage | No | Yes | No | improved |
| Gatism | No | Yes | No | improved |
| Respiratory rate (beats per minute) | 115 ± 13 | 180 ± 25 | 121 ± 16 | 120 ± 19 |
| Pathological damage of lung | ||||
| Bulging and bloodshot | No | Yes | No | improved |
| Inflammatory cell infiltration | No | Yes | Yes | improved |
| Bronchial tube pulmonary alveolus structure | Nomal | Pathological changes | Normal | Lesion improvement |
2.5. Cell Composition and Cytokine Changes
| Cell Classification and Cytokine Level | Control | Asthma | rAdV-GFP | rAdV-CTLA4Ig and rAdV-CCR7 |
|---|---|---|---|---|
| (n = 10) | (n = 10) | (n = 10) | (n = 10) | |
| BALF | ||||
| Total white blood cells (×104/mL) | 21.8 ± 3.25 | 155.93 ± 5.80 | 165.49 ± 7.00 | 18.89 ± 2.17 *,# |
| Eosinophils (×104 mL) | 0.45 ± 0.23 | 17.05 ± 1.59 | 20.74 ± 3.53 | 0.50 ± 0.24 *,# |
| Lymphocyte (×104 mL) | 10.94 ± 2.94 | 92.25 ± 6.90 | 91.13 ± 6.81 | 8.11 ± 1.12 *,# |
| Neutrophil (×104 mL) | 10.41 ± 2.47 | 46.63 ± 7.65 | 53.63 ± 5.21 | 10.28 ± 2.62 *,# |
| BALF | ||||
| IL-4 (pg/mL) | 93.28 ± 14.01 | 156.08 ± 51.78 | 161.18 ± 56.68 | 84.02 ± 17.30 *,# |
| IFNγ (pg/mL) | 162.16 ± 62.14 | 104.25 ± 14.99 | 98.12 ± 14.74 | 55.87 ± 12.8 *,# |
| Sera | ||||
| IL-4 (pg/mL) | 36.47 ± 1.46 | 39.82 ± 1.43 | 36.19 ± 2.94 | 35.92 ± 1.93 |
| IFNγ (pg/mL) | 65.27 ± 3.31 | 52.75 ± 1.93 | 48.78 ± 4.17 | 55.27 ± 3.31 |
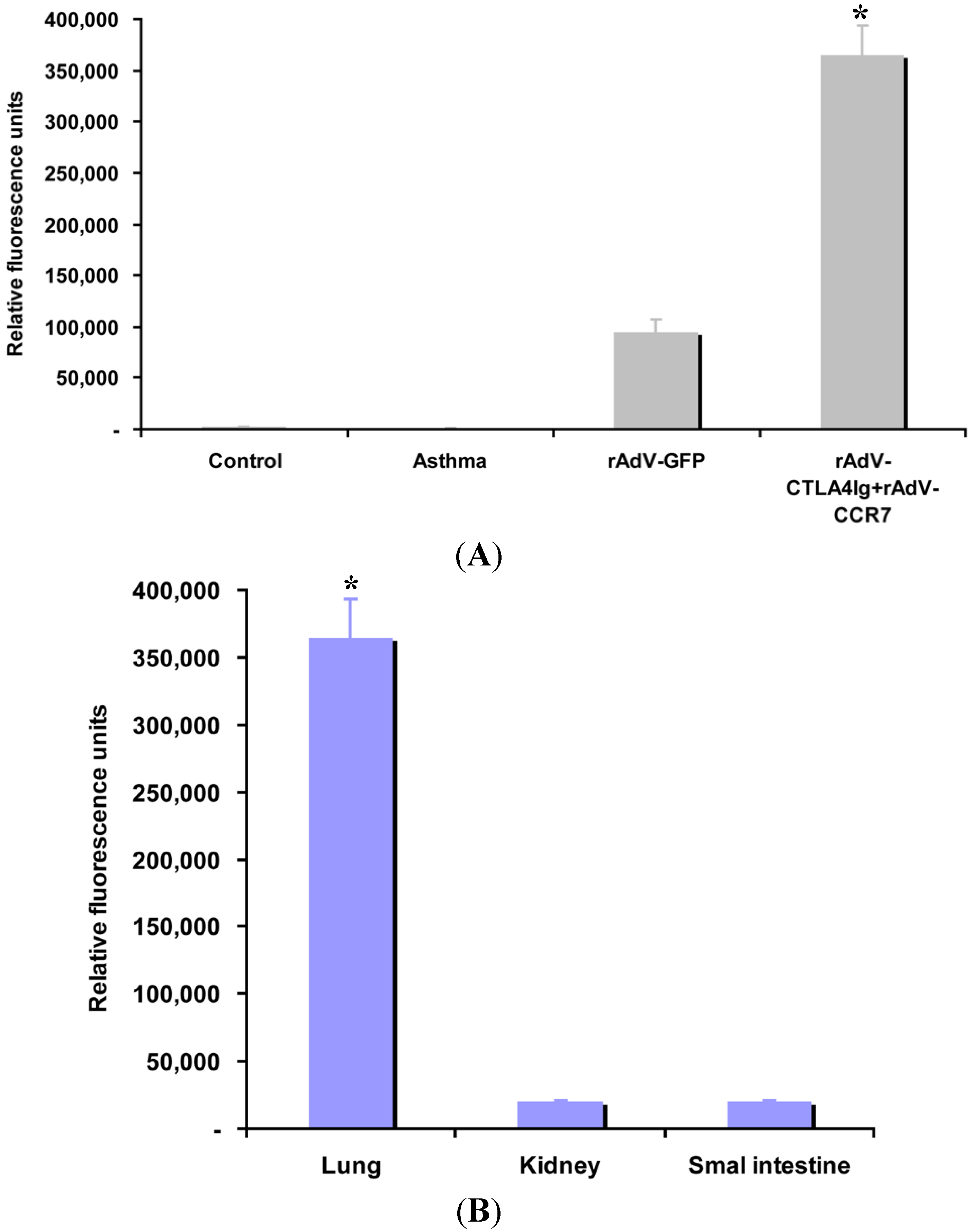
2.6. CCR7 Guides DC Migration
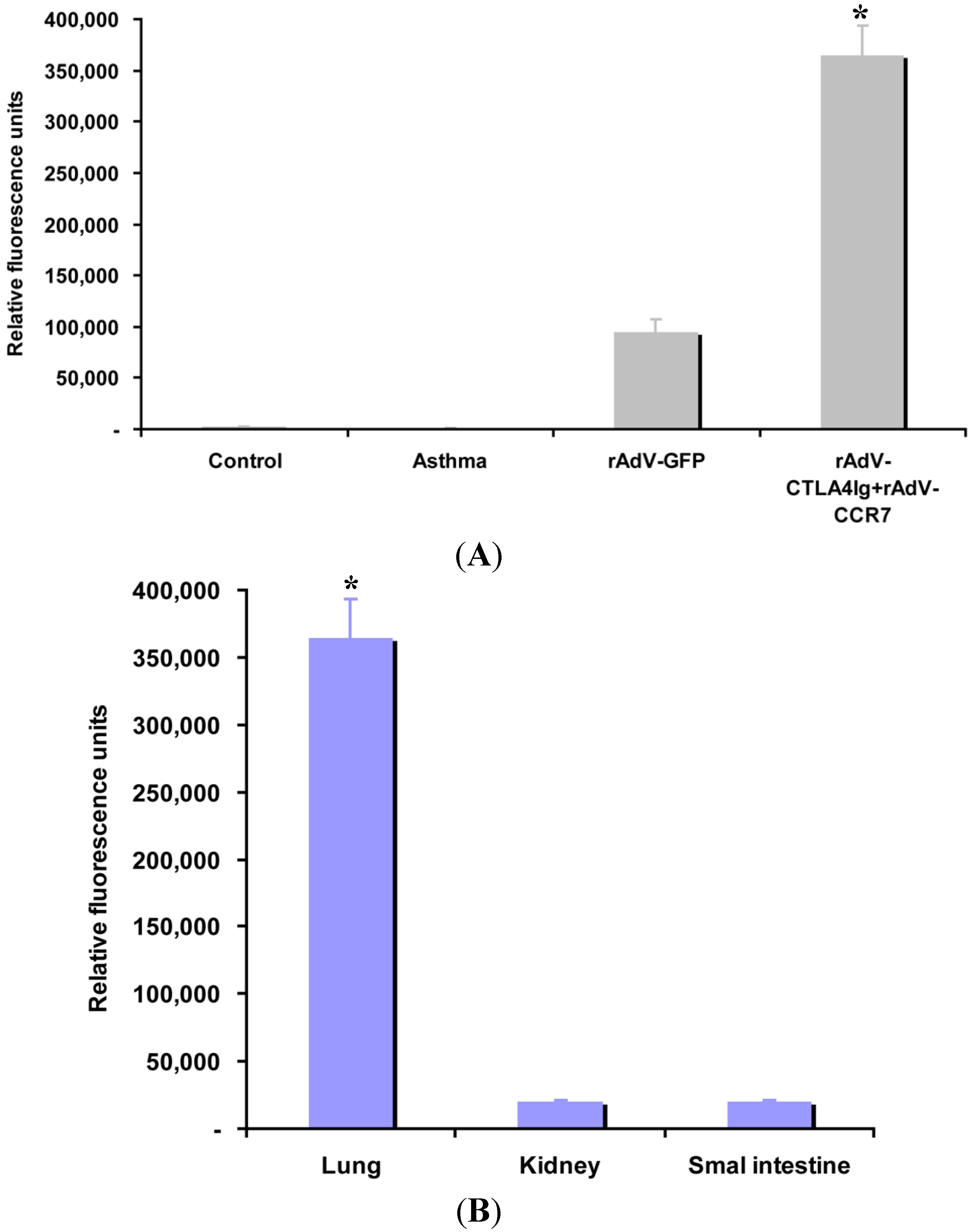
3. Discussion
4. Experimental Section
4.1. Mice
4.2. Recombinant CTLA4Ig and CCR7 Adenovirus Vectors
4.3. DC Isolation, Induction and Modification
4.4. Fluorescence-Activated Cell Sorting (FACS) Analysis
4.5. CTLA4Ig and CCR7 Expression Analysis
4.6. Allergen Sensitization, Challenge and Treatment
4.7. Cytokine Measurement and Cell Subset Analysis
4.8. Histopathology
4.9. CCR7 Guided DC Homing Analysis
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bush, A.; Saglani, S. Management of severe asthma in children. Lancet 2010, 376, 814–825. [Google Scholar] [CrossRef]
- Shin, Y.S.; Takeda, K.; Gelfand, E.W. Understanding asthma using animal models. Allergy Asthma Immunol. Res. 2009, 1, 10–18. [Google Scholar]
- Bartlett, N.W.; Walton, R.P.; Edwards, M.R.; Aniscenko, J.; Caramori, G.; Zhu, J.; Glanville, N.; Choy, K.J.; Jourdan, P.; Burnet, J.; et al. Mouse models of rhinovirus-induced disease and exacerbation of allergic airway inflammation. Nat. Med. 2008, 14, 199–204. [Google Scholar]
- Nials, A.T.; Uddin, S. Mouse models of allergic asthma: Acute and chronic allergen challenge. Dis. Model Mech. 2008, 1, 213–220. [Google Scholar] [CrossRef]
- Salvi, S.; Barnes, P.J. Chronic obstructive pulmonary disease in non-smokers. Lancet 2009, 374, 733–743. [Google Scholar] [CrossRef]
- Hurst, J.R.; Vestbo, J.; Anzueto, A.; Locantore, N.; Müllerova, H.; Tal-Singer, R.; Miller, B.; Lomas, D.A.; Agusti, A.; Macnee, W.; et al. Susceptibility to exacerbation in chronic obstructive pulmonary disease. N. Engl. J. Med. 2010, 363, 1128–1138. [Google Scholar] [CrossRef]
- NHLBI. Guidelines for the Diagnosis and Treatment of Asthma. Available online: http://www.nhlbi.nih.gov/guidelines/asthma/asthgdln.htm (accessed on 10 April 2014).
- Ying, L.; Fu, Z.; Luo, J.; Zhou, C.; Chen, Y.; Wang, L.; Liu, E. Cytotoxic T lymphocyte antigen 4 immunoglobulin modified dendritic cells attenuate allergic airway inflammation and hyperresponsiveness by regulating the development of T helper type 1 (Th1)/Th2 and Th2/regulatory T cell subsets in a murine model of asthma. Clin. Exp. Immunol. 2011, 165, 130–139. [Google Scholar] [CrossRef]
- Padrid, P.A.; Mathur, M.; Li, X.; Herrmann, K.; Qin, Y.; Cattamanchi, A.; Weinstock, J.; Elliott, D.; Sperling, A.I.; Bluestone, J.A.; et al. CTLA4Ig inhibits airway eosinophilia and hyperresponsiveness by regulating the development of Th1/Th2 subsets in a murine model of asthma. Am. J. Respir. Cell Mol. Biol. 1998, 18, 453–462. [Google Scholar] [CrossRef]
- Wilson, R.H.; Whitehead, G.S.; Nakano, H.; Free, M.E.; Kolls, J.K.; Cook, D.N. Allergic sensitization through the airway primes Th17-dependent neutrophilia and airway hyperresponsiveness. Am. J. Respir. Crit. Care Med. 2009, 180, 720–730. [Google Scholar] [CrossRef]
- Doe, C.; Bafadhel, M.; Siddiqui, S.; Desai, D.; Mistry, V.; Rugman, P.; McCormick, M.; Woods, J.; May, R.; Sleeman, M.A.; et al. Expression of the T helper 17-associated cytokines IL-17A and IL-17F in asthma and COPD. Chest 2010, 138, 1140–1147. [Google Scholar] [CrossRef]
- Al-Ramli, W.; Préfontaine, D.; Chouiali, F.; Martin, J.G.; Olivenstein, R.; Lemière, C.; Hamid, Q. T(H)17-associated cytokines (IL-17A and IL-17F) in severe asthma. J. Allergy Clin. Immunol. 2009, 123, 1185–1187. [Google Scholar] [CrossRef]
- Barczyk, A.; Pierzchala, W.; Sozañska, E. Interleukin-17 in sputum correlates with airway hyperresponsiveness to methacholine. Respir. Med. 2003, 97, 726–733. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Takahashi, D.; Hizawa, N.; Suzuki, S.; Matsukura, S.; Kokubu, F.; Maeda, Y.; Fukui, Y.; Konno, S.; Huang, S.K.; et al. IL-17F sequence variant (His161Arg) is associated with protection against asthma and antagonizes wild-type IL-17F activity. J. Allergy Clin. Immunol. 2006, 117, 795–801. [Google Scholar] [CrossRef]
- Scanlon, S.T.; McKenzie, A.N. Type 2 innate lymphoid cells: New players in asthma and allergy. Curr. Opin. Immunol. 2012, 24, 707–712. [Google Scholar] [CrossRef]
- Cazzola, M.; Page, C.P.; Calzetta, L.; Matera, M.G. Emerging anti-inflammatory strategies for COPD. Eur. Respir. J. 2012, 40, 724–741. [Google Scholar] [CrossRef]
- Hubbell, J.A.; Thomas, S.N.; Swartz, M.A. Materials engineering for immunomodulation. Nature 2009, 462, 449–460. [Google Scholar] [CrossRef]
- Chen, L. Co-inhibitory molecules of the B7-CD28 family in the control of T-cell immunity. Nat. Rev. Immunol. 2004, 4, 336–347. [Google Scholar] [CrossRef]
- Van der Merwe, P.A.; Dushek, O. Mechanisms for T cell receptor triggering. Nat. Rev. Immunol. 2011, 11, 47–55. [Google Scholar] [CrossRef]
- Goldrath, A.W.; Bevan, M.J. Selecting and maintaining a diverse T-cell repertoire. Nature 1999, 402, 255–262. [Google Scholar] [CrossRef]
- Jung, K.; Choi, I. Emerging co-signaling networks in T cell immune regulation. Immune Netw. 2013, 13, 184–193. [Google Scholar] [CrossRef]
- Vanneman, M.; Dranoff, G. Combining immunotherapy and targeted therapies in cancer treatment. Nat. Rev. Cancer 2012, 12, 237–251. [Google Scholar] [CrossRef]
- Deppong, C.M.; Green, J.M. Experimental advances in understanding allergic airway inflammation. Front. Biosci. 2013, 5, 167–180. [Google Scholar]
- Vinall, S.L.; Townsend, E.R.; Pettipher, R. A paracrine role for chemoattractant receptor-homologous molecule expressed on T helper type 2 cells (CRTH2) in mediating chemotactic activation of CRTH2+ CD4+ T helper type 2 lymphocytes. Immunology 2007, 121, 577–584. [Google Scholar] [CrossRef]
- Xu, Z.; Juan, V.; Ivanov, A.; Ma, Z.; Polakoff, D.; Powers, D.B.; Dubridge, R.B.; Wilson, K.; Akamatsu, Y. Affinity and cross-reactivity engineering of CTLA4-Ig to modulate T cell costimulation. J. Immunol. 2012, 189, 4470–4477. [Google Scholar] [CrossRef]
- Cecchinato, V.; Tryniszewska, E.; Ma, Z.M.; Vaccari, M.; Boasso, A.; Tsai, W.P.; Petrovas, C.; Fuchs, D.; Heraud, J.M.; Venzon, D.; et al. Immune activation driven by CTLA-4 blockade augments viral replication at mucosal sites in simian immunodeficiency virus infection. J. Immunol. 2008, 180, 5439–5447. [Google Scholar] [CrossRef]
- Abrams, J.R.; Lebwohl, M.G.; Guzzo, C.A.; Jegasothy, B.V.; Goldfarb, M.T.; Goffe, B.S.; Menter, A.; Lowe, N.J.; Krueger, G.; Brown, M.J.; et al. CTLA4Ig-mediated blockade of T-cell costimulation in patients with psoriasis vulgaris. J. Clin. Investig. 1999, 103, 1243–1252. [Google Scholar]
- Zhang, J.; Chen, C.; Chen, Y.; Wang, L.B.; Zhang, X.B.; Zhang, M.Z. Experimental study of CTLA4Ig gene-modified dendritic cells transfused neonatally inducing asthmatic immune tolerance in adult mice. Zhonghua Yi Xue Za Zhi 2012, 92, 848–852. [Google Scholar]
- Vander Lugt, B.; Tubo, N.J.; Nizza, S.T.; Boes, M.; Malissen, B.; Fuhlbrigge, R.C.; Kupper, T.S.; Campbell, J.J. CCR7 plays no appreciable role in trafficking of central memory CD4 T cells to lymph nodes. J. Immunol. 2013, 191, 3119–3127. [Google Scholar]
- Chen, E.J.; Shaffer, M.H.; Williamson, E.K.; Huang, Y.; Burkhardt, J.K. Ezrin and moesin are required for efficient T cell adhesion and homing to lymphoid organs. PLoS One 2013, 8, e52368. [Google Scholar]
- Bromley, S.K.; Thomas, S.Y.; Luster, A.D. Chemokine receptor CCR7 guides T cell exit from peripheral tissues and entry into afferent lymphatics. Nat. Immunol. 2005, 6, 895–901. [Google Scholar]
- Ritprajak, P.; Hashiguchi, M.; Azuma, M. Topical application of cream-emulsified CD86 siRNA ameliorates allergic skin disease by targeting cutaneous dendritic cells. Mol. Ther. 2008, 16, 1323–1330. [Google Scholar]
- Leake, I. Viral hepatitis: Subinfectious HCV exposure. Nat. Rev. Gastroenterol. Hepatol. 2014. [Google Scholar] [CrossRef]
- Thomson, A.W.; Knolle, P.A. Antigen-presenting cell function in the tolerogenic liver environment. Nat. Rev. Immunol. 2010, 10, 753–766. [Google Scholar] [CrossRef]
- Geissmann, F.; Gordon, S.; Hume, D.A.; Mowat, A.M.; Randolph, G.J. Unravelling mononuclear phagocyte heterogeneity. Nat. Rev. Immunol. 2010, 10, 453–460. [Google Scholar] [CrossRef]
- Manicassamy, S.; Pulendran, B. Dendritic cell control of tolerogenic responses. Immunol. Rev. 2011, 241, 206–227. [Google Scholar] [CrossRef]
- Ten Hacken, N.H.; Oosterhoff, Y.; Kauffman, H.F.; Guevarra, L.; Satoh, T.; Tollerud, D.J.; Postma, D.S. Elevated serum interferon-γ in atopic asthma correlates with increased airways responsiveness and circadian peak expiratory flow variation. Eur. Respir. J. 1998, 11, 312–316. [Google Scholar] [CrossRef]
- Sakai, K.; Yokoyama, A.; Kohno, N.; Hiwada, K. Effect of different sensitizing doses of antigen in a murine model of atopic asthma. Clin. Exp. Immunol. 1999, 118, 9–15. [Google Scholar] [CrossRef]
- Mitchell, C.; Provost, K.; Niu, N.; Homer, R.; Cohn, L. IFNγ acts on the airway epithelium to inhibit local and systemic pathology in allergic airway disease. J. Immunol. 2011, 187, 3815–3820. [Google Scholar] [CrossRef]
- Ray, A.; Khare, A.; Krishnamoorthy, N.; Qi, Z.; Ray, P. Regulatory T cells in many flavors control asthma. Mucosal Immunol. 2010, 3, 216–229. [Google Scholar] [CrossRef]
- Parulekar, A.D.; Boomer, J.S.; Patterson, B.M.; Yin-Declue, H.; Deppong, C.M.; Wilson, B.S.; Jarjour, N.N.; Castro, M.; Green, J.M. A randomized controlled trial to evaluate inhibition of T-cell costimulation in allergen-induced airway inflammation. Am. J. Respir. Crit. Care Med. 2013, 187, 494–501. [Google Scholar] [CrossRef]
- Inaba, K.; Inaba, M.; Romani, N.; Aya, H.; Deguchi, M.; Ikehara, S.; Muramatsu, S.; Steinman, R.M. Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 1992, 176, 1693–1702. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, Y.; Wang, Y.; Fu, Z. T Lymphocyte Antigen 4-Modified Dendritic Cell Therapy for Asthmatic Mice Guided by the CCR7 Chemokine Receptor. Int. J. Mol. Sci. 2014, 15, 15304-15319. https://doi.org/10.3390/ijms150915304
Chen Y, Wang Y, Fu Z. T Lymphocyte Antigen 4-Modified Dendritic Cell Therapy for Asthmatic Mice Guided by the CCR7 Chemokine Receptor. International Journal of Molecular Sciences. 2014; 15(9):15304-15319. https://doi.org/10.3390/ijms150915304
Chicago/Turabian StyleChen, Yan, Yongming Wang, and Zhou Fu. 2014. "T Lymphocyte Antigen 4-Modified Dendritic Cell Therapy for Asthmatic Mice Guided by the CCR7 Chemokine Receptor" International Journal of Molecular Sciences 15, no. 9: 15304-15319. https://doi.org/10.3390/ijms150915304
APA StyleChen, Y., Wang, Y., & Fu, Z. (2014). T Lymphocyte Antigen 4-Modified Dendritic Cell Therapy for Asthmatic Mice Guided by the CCR7 Chemokine Receptor. International Journal of Molecular Sciences, 15(9), 15304-15319. https://doi.org/10.3390/ijms150915304