Expression of S100A6 in Rat Hippocampus after Traumatic Brain Injury Due to Lateral Head Acceleration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
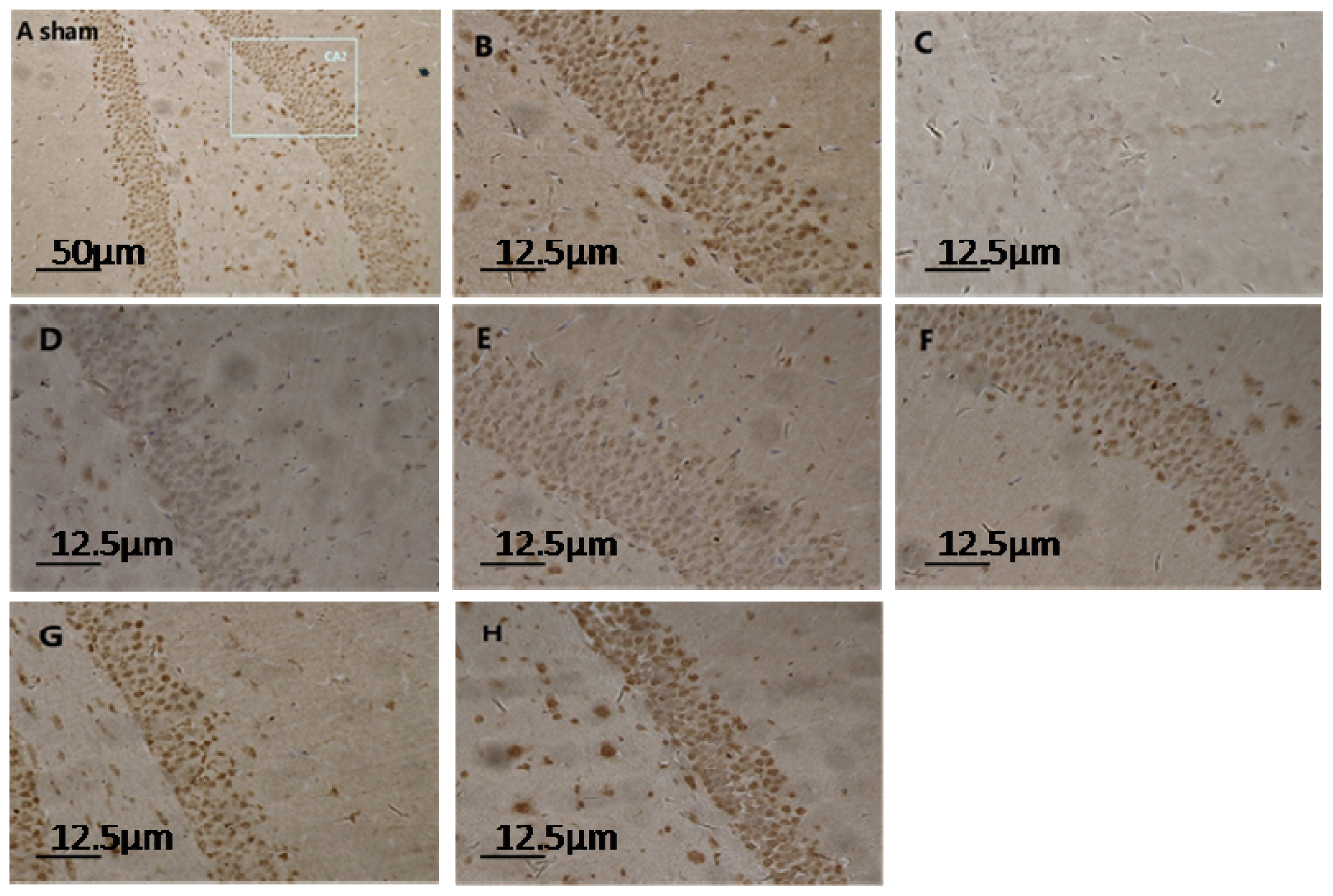
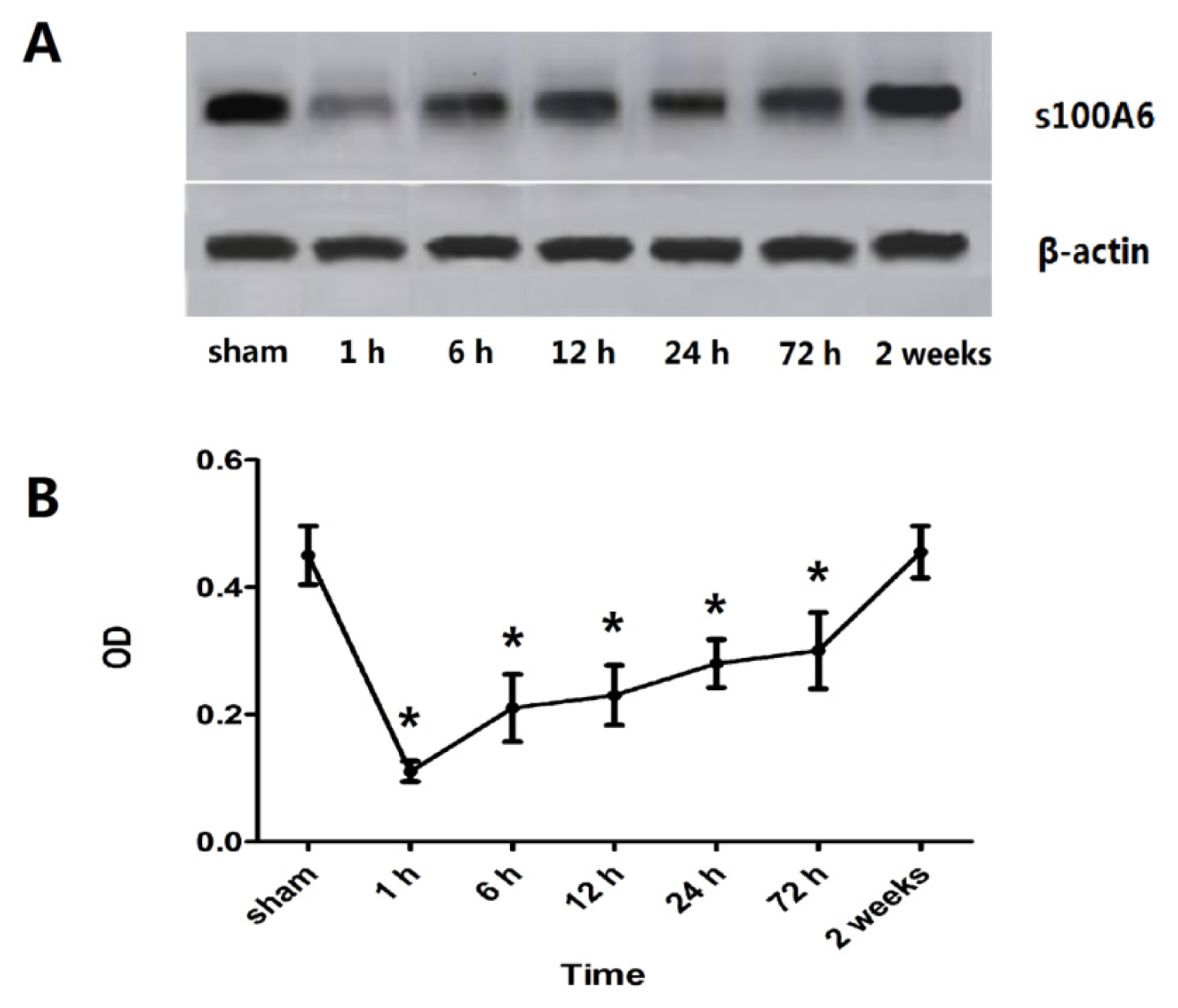
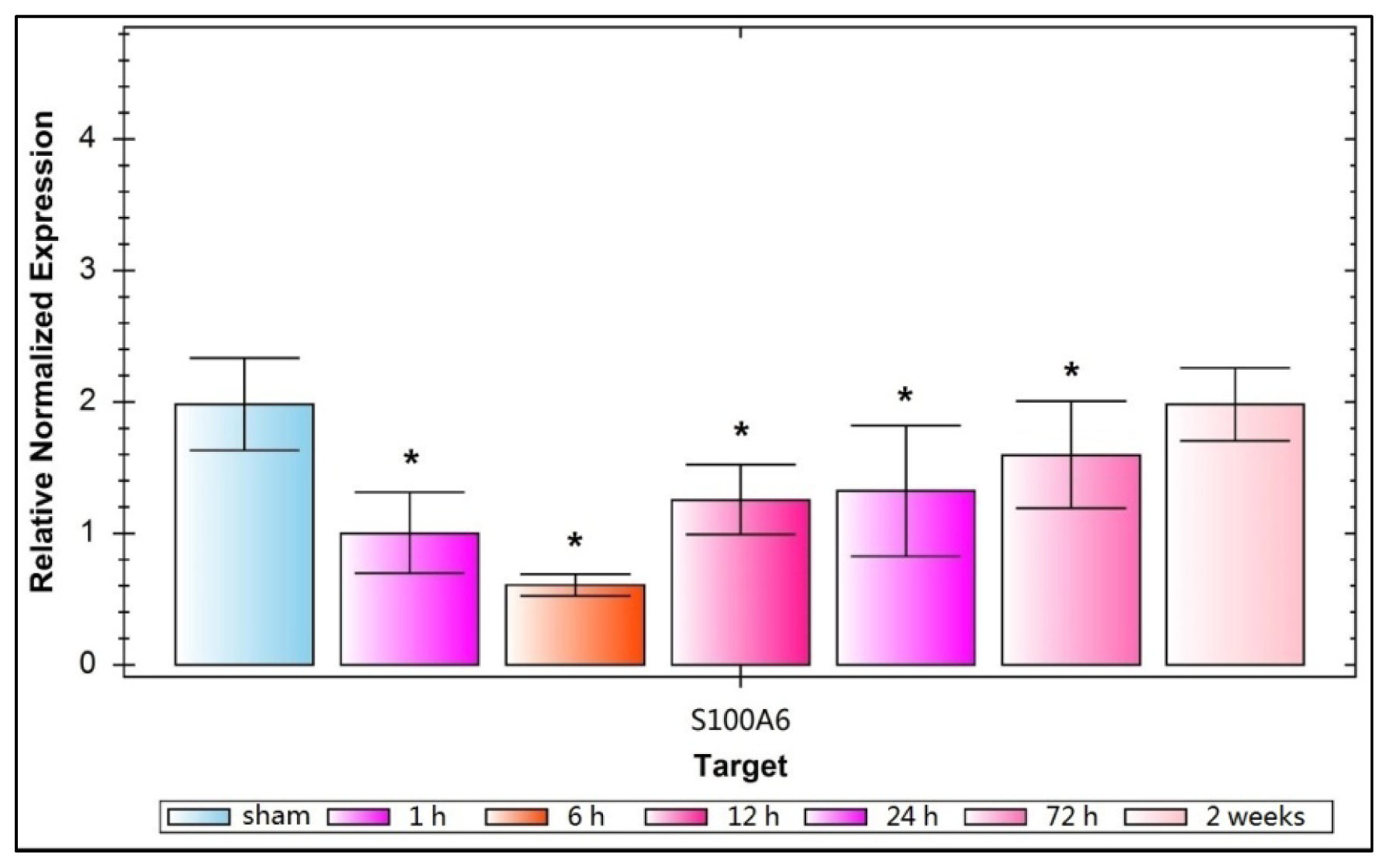
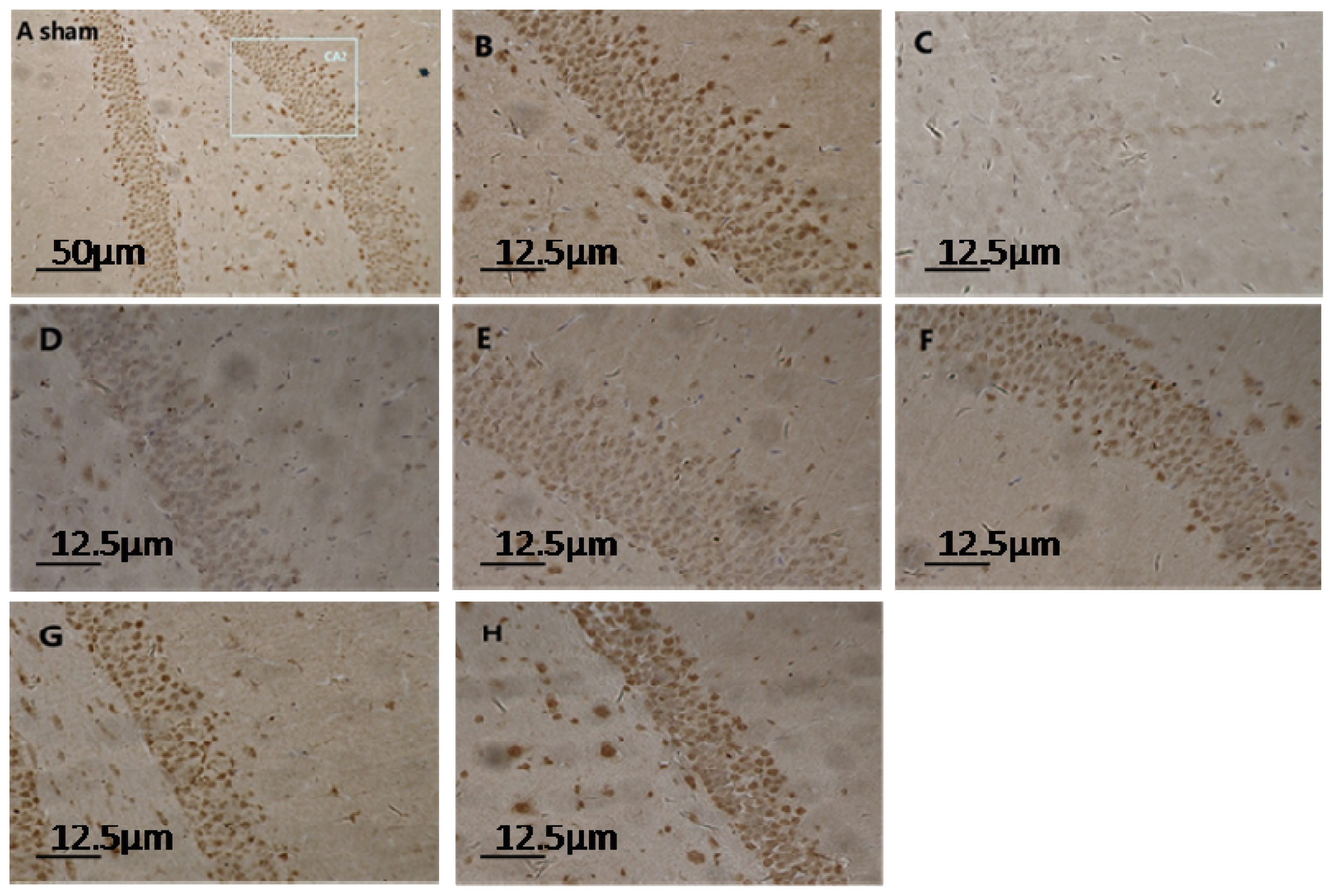
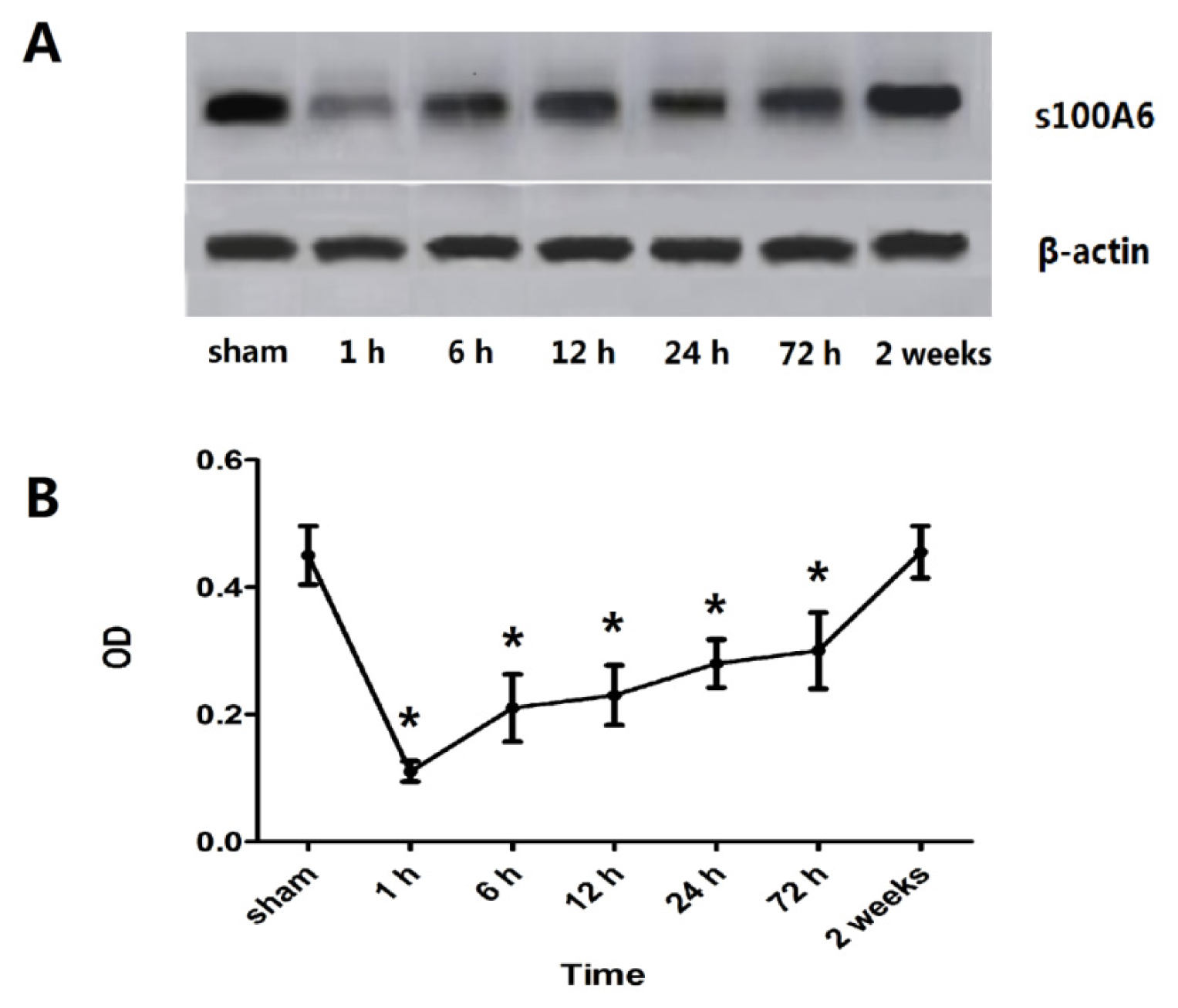
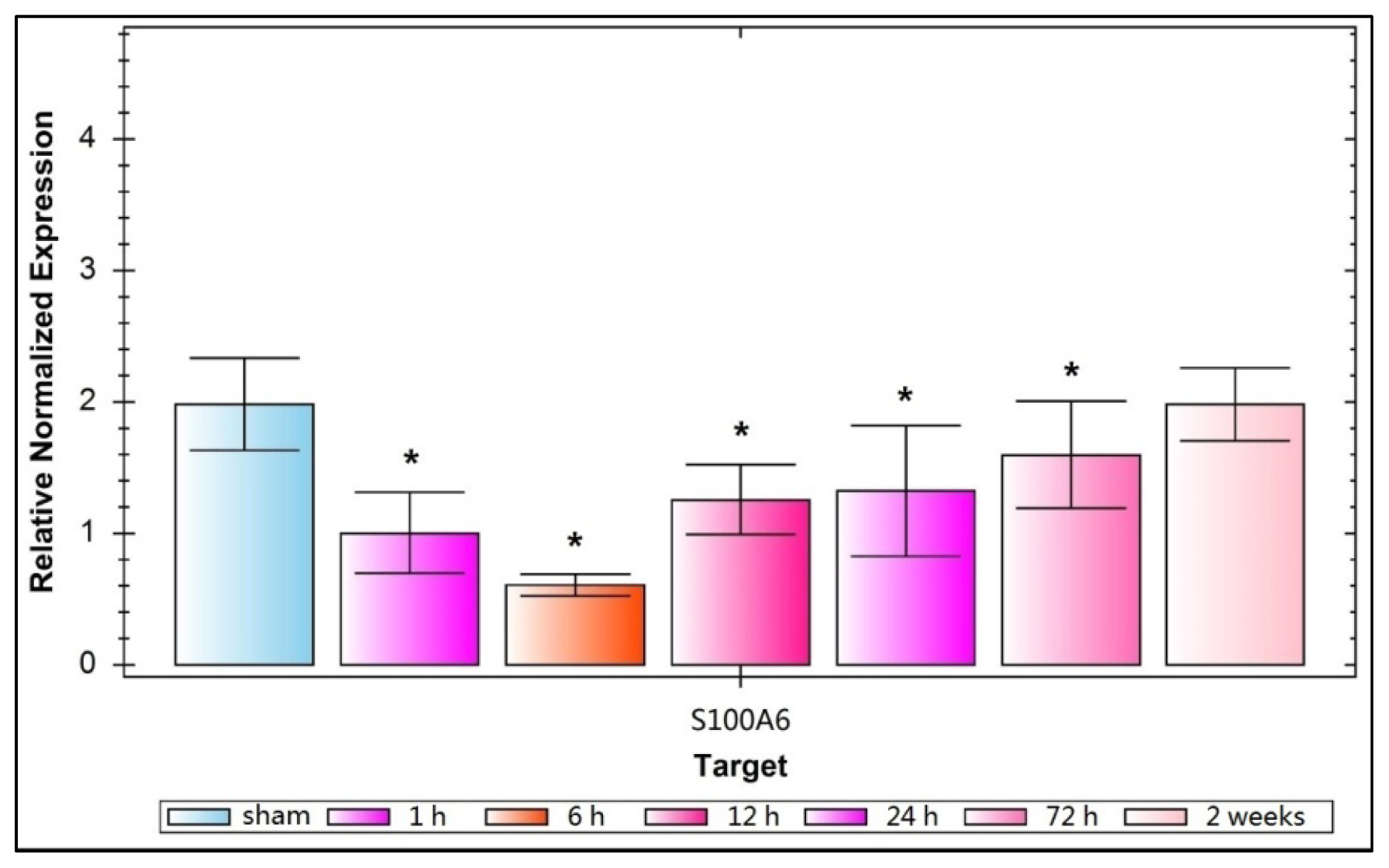
2.1. Expression of Hippocampal S100A6: Immunohistochemical Staining, Protein, and mRNA Levels
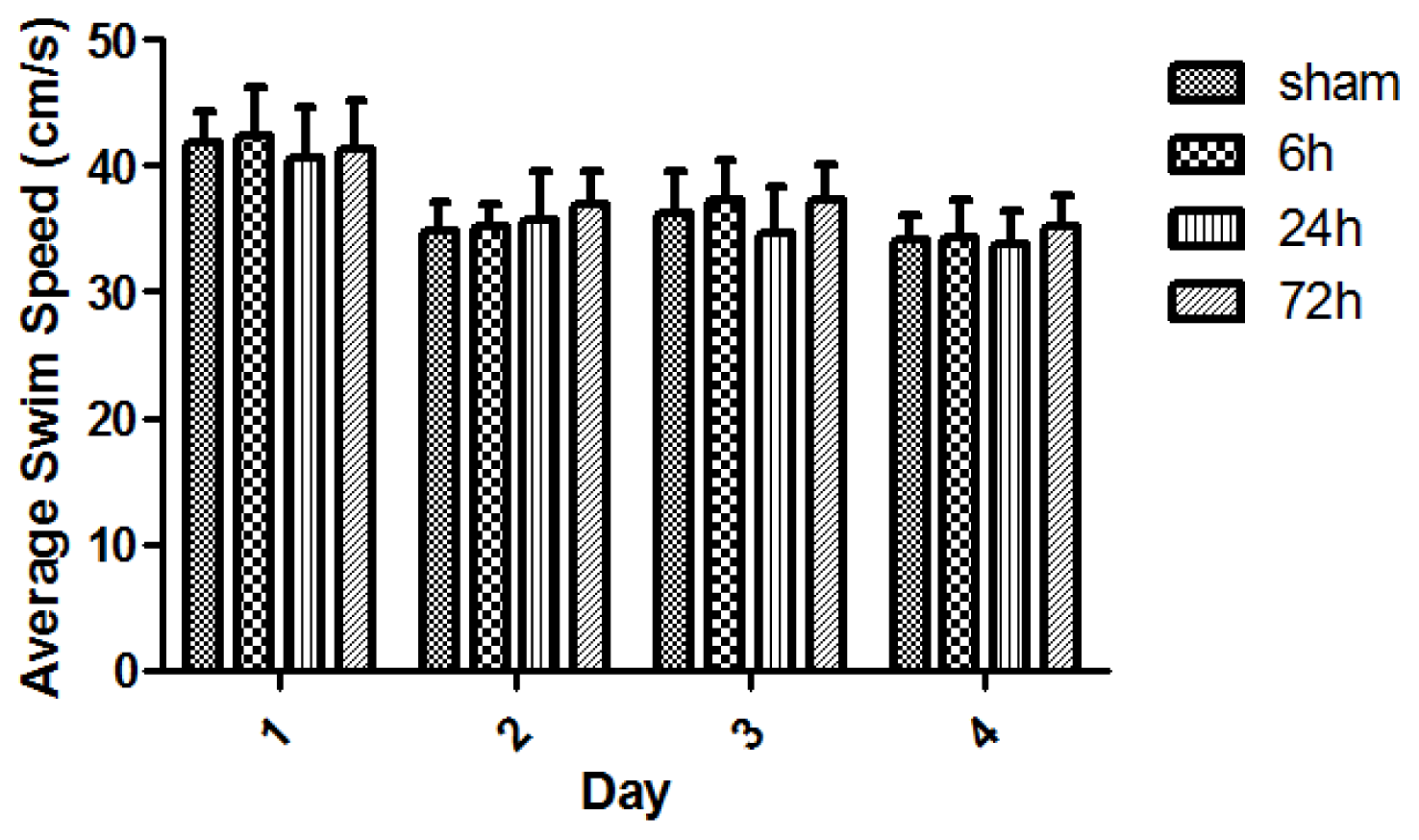
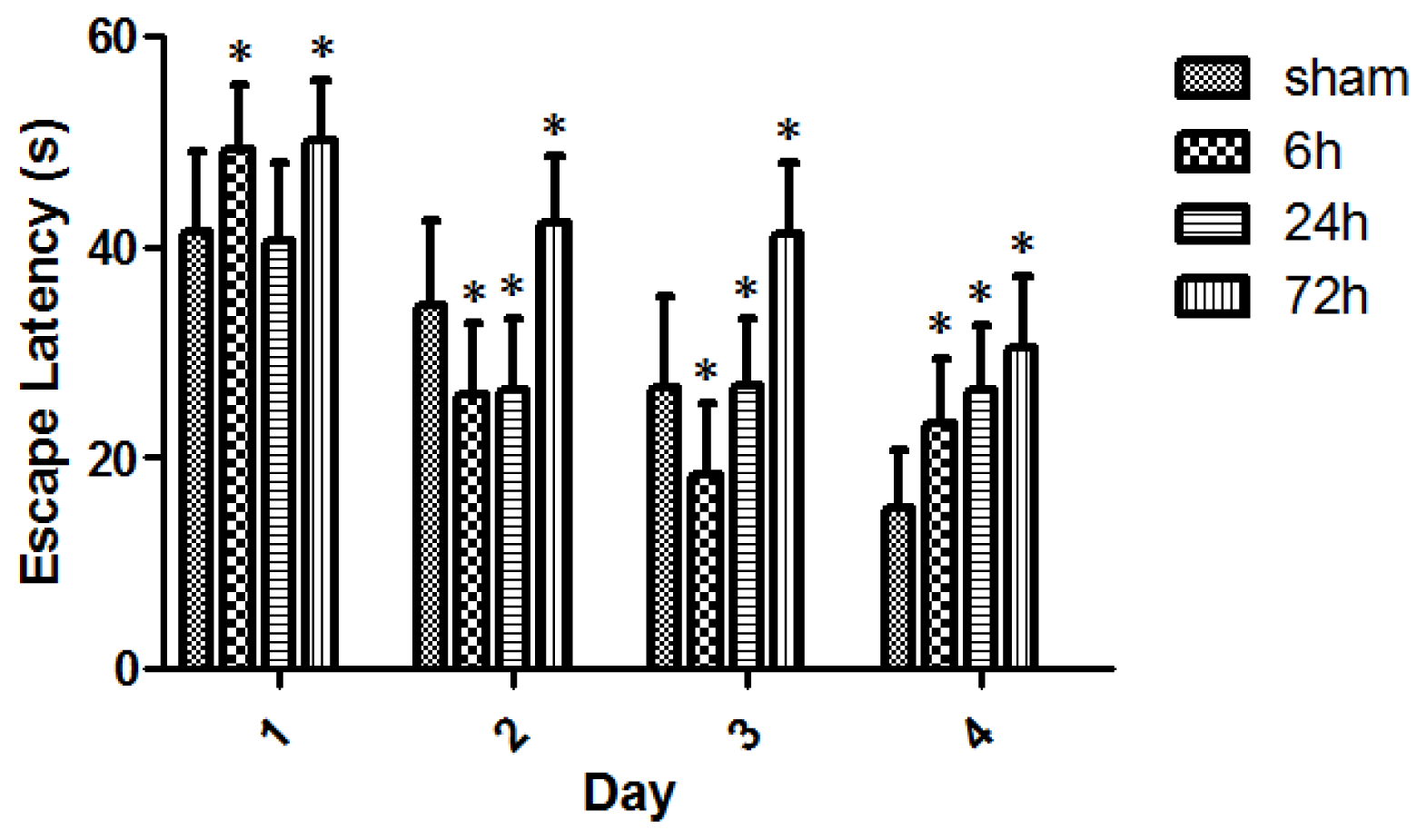
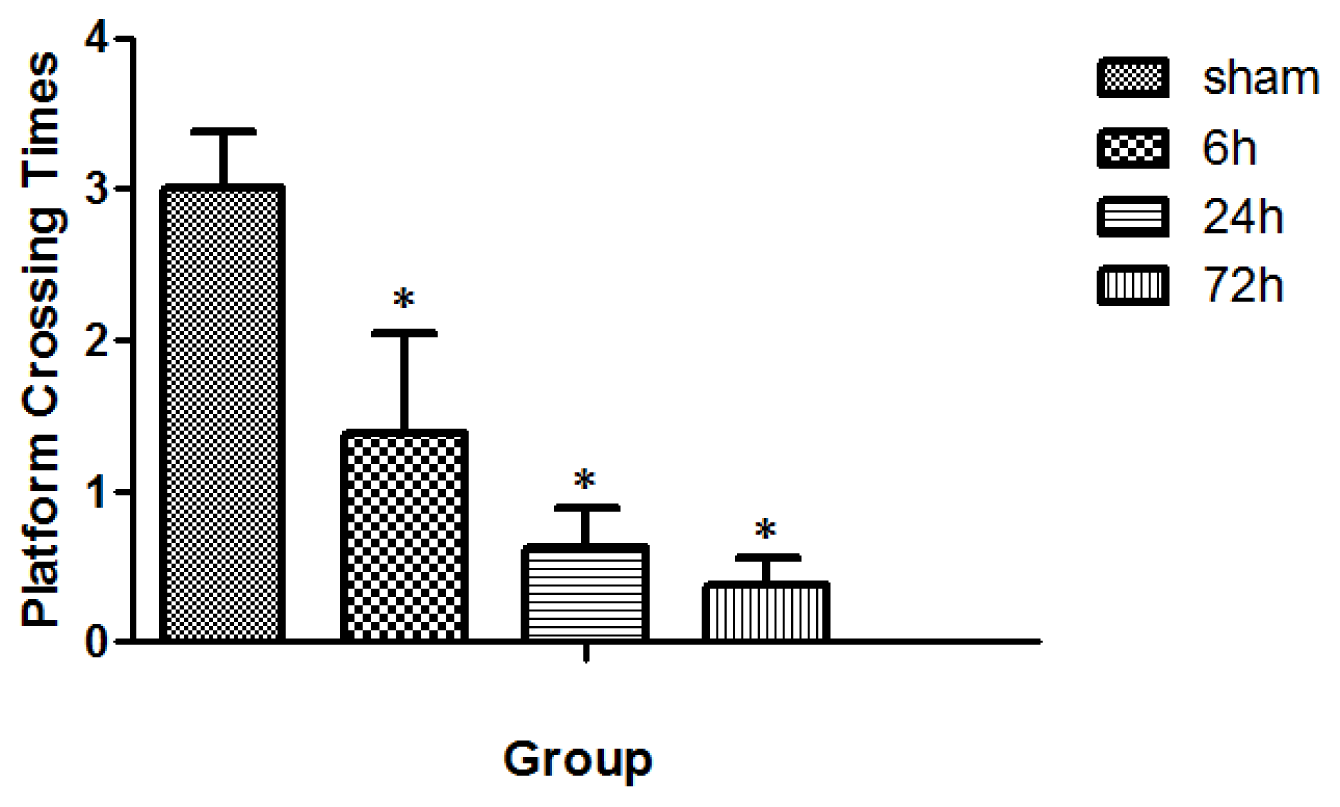
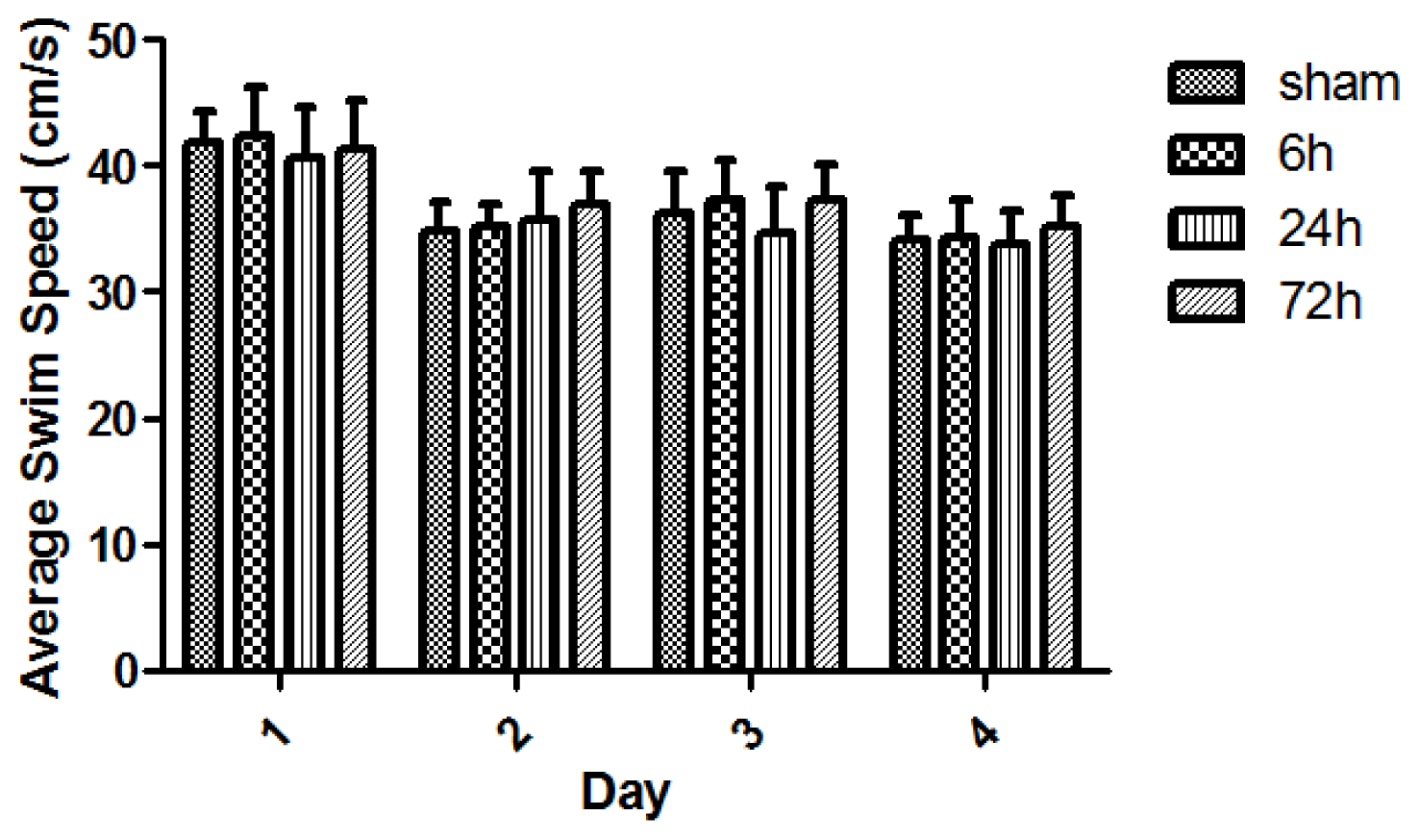
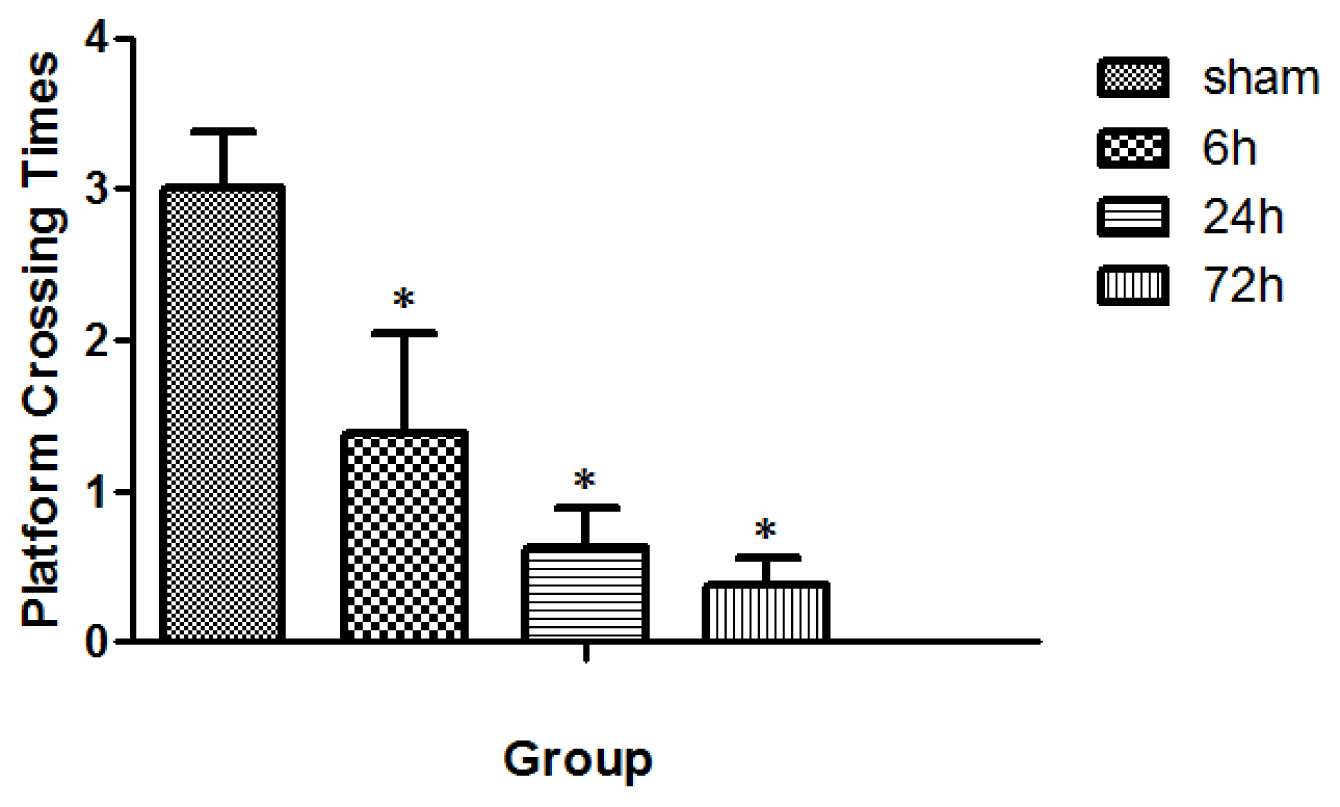
2.2. Learning and Memory Dysfunction after TBI
3. Discussion
4. Materials and Methods
4.1. Animals
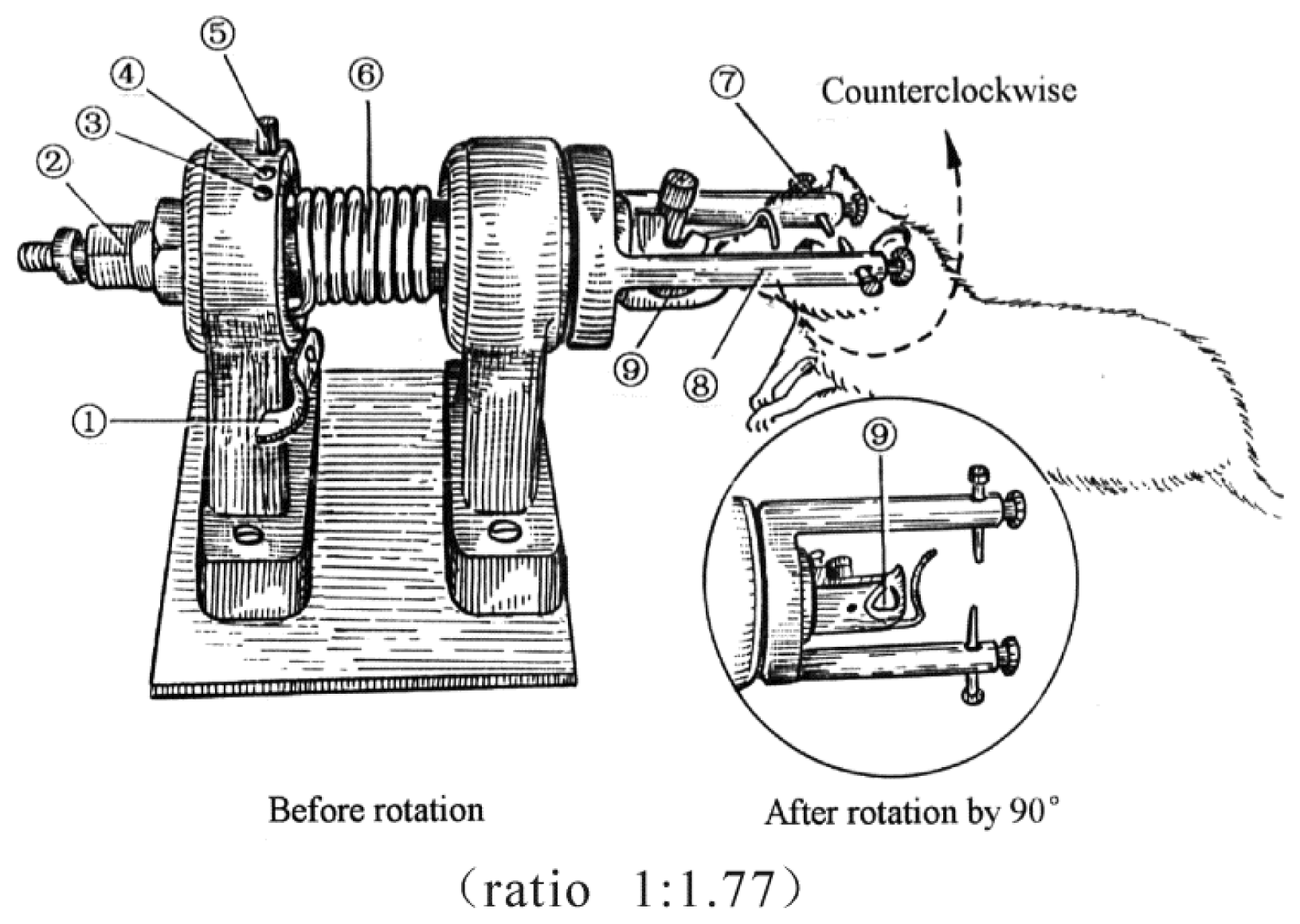
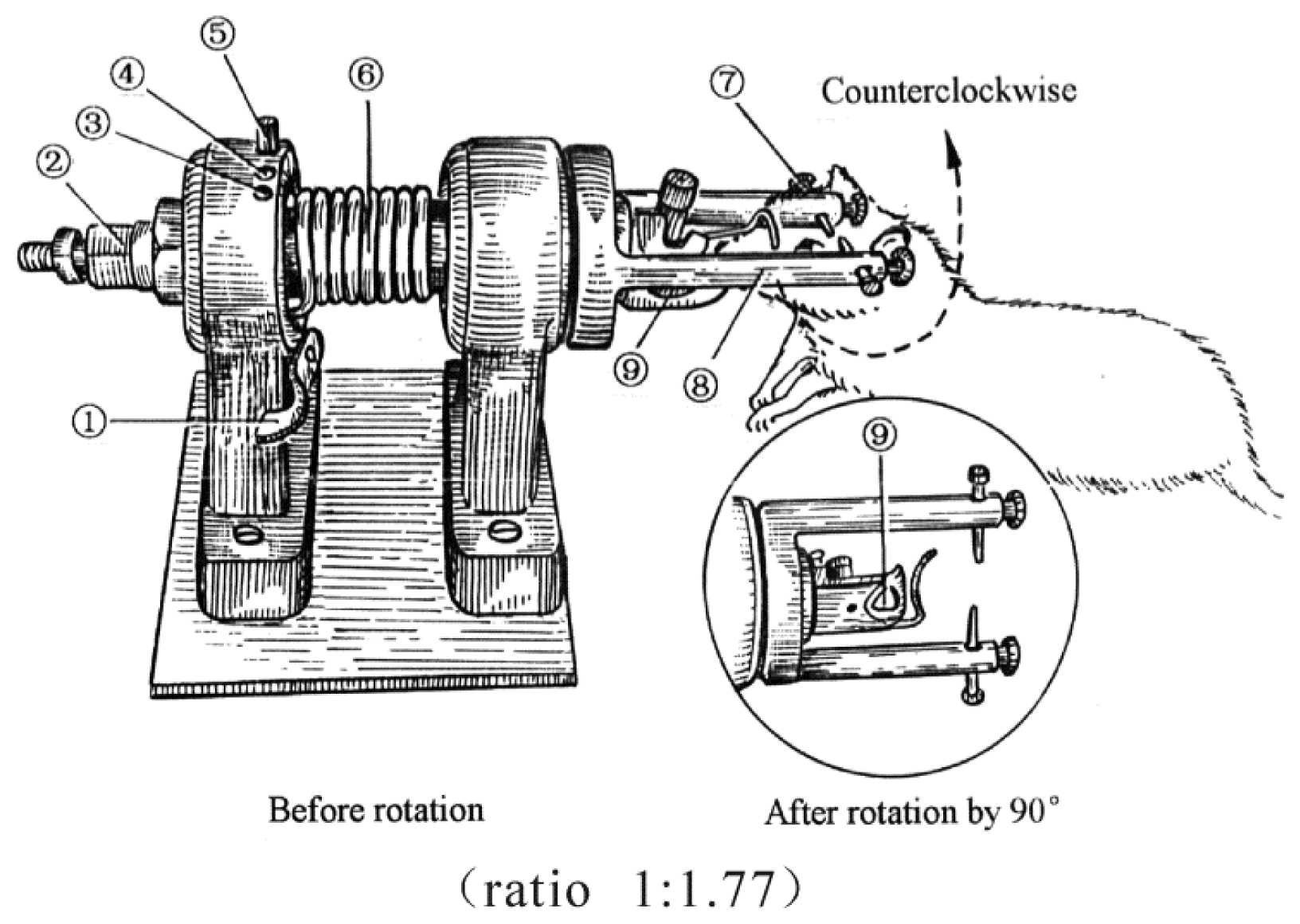
4.2. TBI Model and Grouping of Animals
4.3. Immunohistochemical Staining
4.4. Western Blot
4.5. RT-PCR
4.6. MWM
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bayir, H.; Clark, R.S.; Kochanek, P.M. Promising strategies to minimize secondary brain injury after head trauma. Crit. Care Med 2003, 31, 1–14. [Google Scholar]
- Dearden, N.M.; Gibson, J.S.; McDowall, D.G.; Gibson, R.M.; Cameron, M.M. Effect of high-dose dexamethasone on outcome from severe head injury. J. Neurosurg 1986, 64, 81–88. [Google Scholar]
- Hoskison, M.M.; Moore, A.N.; Hu, B.; Orsi, S.; Kobori, N.; Dash, P.K. Persistent working memory dysfunction following traumatic brain injury: Evidence for a time-dependent mechanism. Neuroscience 2009, 159, 483–491. [Google Scholar]
- White, N.M.; McDonald, R.J. Multiple parallel memory systems in the brain of the rat. Neurobiol. Learn. Mem 2002, 77, 125–184. [Google Scholar]
- Xiao-Sheng, H.; Sheng-Yu, Y.; Xiang, Z.; Fu, L.A.; Shuang, W. Calcium overloading in traumatic axonal injury by lateral head rotation: A morphological evidence in a rat model. J. Clin. Neurosci 2004, 11, 402–407. [Google Scholar]
- Eckert, R.L.; Broome, A.M.; Ruse, M.; Robinson, N.; Ryan, D.; Lee, K. S100 Proteins in the Epidermis. J. Investig. Dermatol 2004, 123, 23–33. [Google Scholar]
- Bian, X.X.; Yuan, X.S.; Qi, C.P. Effect of recombinant human erythropoietin on serum S100B protein and interleukin-6 levels after traumatic brain injury in the rat. Neurol. Med. Chir. (Tokyo) 2010, 50, 361–366. [Google Scholar]
- Schiavi, P.; Laccarino, C.; Servadei, F. The value of the calcium binding protein S100 in the management of patients with traumatic brain injury. Acta Biomed 2012, 83, 5–20. [Google Scholar]
- Courtois-Coutry, N.; le Moellic, C.; Boulkroun, S.; Fay, M.; Cluzeaud, F.; Escoubet, B.; Farman, N.; Blot-Chabaud, M. Calcyclin is an early vasopressin-induced gene in the renal collecting duct: Role in the long-term regulation of ion transport. J. Biol. Chem 2002, 277, 25728–25734. [Google Scholar]
- Okazaki, K.; Niki, I.; Iino, S.; Kobayashi, S.; Hidaka, H. A role of calcyclin, a Ca(2+)-binding protein, on the Ca(2+)-dependent insulin release from the pancreatic beta cell. J. Biol. Chem 1994, 269, 6149–6152. [Google Scholar]
- Filipek, A.; Jastrzebska, B.; Nowotny, M.; Kuznicki, J. CacyBP/SIP, a calcyclin and Siah-1-interacting protein, binds EF-hand proteins of the S100 family. J. Biol. Chem 2002, 277, 28848–28852. [Google Scholar]
- Leclerc, E.; Fritz, G.; Weibel, M.; Heizmann, C.W.; Galichet, A. S100B and S100A6 differentially modulate cell survival by interacting with distinct RAGE (receptor for advanced glycation end products) immunoglobulin domains. J. Biol. Chem 2007, 282, 31317–31331. [Google Scholar]
- Yamada, J.; Jinno, S. Upregulation of calcium binding protein, S100A6, in activated astrocytes is linked to glutamate toxicity. Neuroscience 2012, 226, 119–129. [Google Scholar]
- Filipek, A.; Wojda, U. p30, a novel protein target of mouse calcyclin (S100A6). Biochem. J 1996, 320, 585–587. [Google Scholar]
- Filipek, A.; Jastrzebska, B.; Nowotny, M.; Kwiatkowska, K.; Hetman, M.; Surmacz, L.; Wyroba, E.; Kuznicki, J. Ca2+-dependent translocation of the calcyclin-binding protein in neurons and neuroblastoma NB-2a cells. J. Biol. Chem 2002, 277, 21103–21109. [Google Scholar]
- Wu, J.; Tan, X.; Peng, X.; Yuan, J.; Qiang, B. Translocation and phosphorylation of calcyclin binding protein during retinoic acid-induced neuronal differentiation of neuroblastoma SH-SY5Y cells. J. Biochem. Mol. Biol 2003, 36, 354–358. [Google Scholar]
- Jastrzebska, B.; Filipek, A.; Nowicka, D.; Kaczmarek, L.; Kúznicki, J. Calcyclin (S100A6) binding protein (CacyBP) is highly expressed in brain neurons. J. Histochem. Cytochem 2000, 48, 1195–1202. [Google Scholar]
- Schneider, G.; Filipek, A. S100A6 binding protein and Siah-1 interacting protein (CacyBP/SIP): Spotlight on properties and cellular function. Amino Acids 2011, 41, 773–780. [Google Scholar]
- Douma, B.R.K.; Korte, S.M.; Buwalda, B.; la Fleur, S.E.; Bohus, B.; Luiten, P.G.M. Repeated blockade of mineralocorticoid receptors, but not of glucocorticoid receptors impairs food rewarded spatial learning. Psychoneuroendocrinology 1998, 23, 33–44. [Google Scholar]
- Wei, G.; Hongyu, X.; Ming, L.; Jason, H.H.; Xiaosheng, H. Association between reduced expression of hippocampal glucocorticoid receptors and cognitive dysfunction in a rat model of traumatic brain injury due to lateral head acceleration. Neurosci. Lett 2013, 533, 50–54. [Google Scholar]
- Filipek, A.; Kuznicki, J. Molecular cloning and expression of a mouse brain cDNA encoding a novel protei target of calcyclin. J. Neurochem 1998, 70, 1793–1798. [Google Scholar]
- Shapira, M.; Licht, A.; Milman, A.; Pick, C.G.; Shohami, E.; Eldar-Finkelman, H. Role of glycogen synthase kinase-3 beta in early depressive behavior induced by mild traumatic brain injury. Mol. Cell. Neurosci 2007, 34, 571–577. [Google Scholar]
- Li, H.L.; Wang, H.H.; Liu, S.J.; Deng, Y.Q.; Zhang, Y.J.; Tian, Q.; Wang, X.C.; Chen, X.Q.; Yang, Y.; Zhang, J.Y.; et al. Phosphorylation of tau antagonizes apoptosis by stabilizing β-catenin, a mechanism involved in Alzheimer’s neurodegeneration. Proc. Natl. Acad. Sci. USA 2007, 104, 3591–3596. [Google Scholar]
- Fukushima, T.; Zapata, J.M.; Singha, N.C.; Thomas, M.; Kress, C.L.; Krajewska, M.; Krejewiski, S.; Ronai, Z.; Reed, J.C.; Matsuzawa, S. Critical function for SIP, a ubiquitin E3 ligase component of the β-catenin degradation pathway for thymocyte development and G1 check point. Immunity 2006, 24, 29–39. [Google Scholar]
- Schneider, G.; Nieznanski, K.; Kilanczyk, E.; Bieganowski, P.; Kuznicki, J.; Filipek, A. CacyBP/SIP interacts with tubulin in neuroblastoma NB2a cells and induces formation of globular tubulin assemblies. Biochim. Biophys. Acta 2007, 1773, 1628–1636. [Google Scholar]
- Filipek, A.; Schneider, G.; Mietelska, A.; Figiel, L.; Niewiadomska, G. Age-dependent changes in neuronal distribution of CacyBP/SIP: Comparison to tubulin and the tau protein. J. Neural Transm 2008, 115, 1257–1264. [Google Scholar]
- Xiao-Sheng, H.; Sheng-Yu, Y.; Xiang, Z.; Zhou, F.; Jian-ning, Z. Diffuse axonal injury due to lateral head rotation in a rat model. J. Neurosurg 2000, 93, 626–633. [Google Scholar]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar]

© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fang, B.; Liang, M.; Yang, G.; Ye, Y.; Xu, H.; He, X.; Huang, J.H. Expression of S100A6 in Rat Hippocampus after Traumatic Brain Injury Due to Lateral Head Acceleration. Int. J. Mol. Sci. 2014, 15, 6378-6390. https://doi.org/10.3390/ijms15046378
Fang B, Liang M, Yang G, Ye Y, Xu H, He X, Huang JH. Expression of S100A6 in Rat Hippocampus after Traumatic Brain Injury Due to Lateral Head Acceleration. International Journal of Molecular Sciences. 2014; 15(4):6378-6390. https://doi.org/10.3390/ijms15046378
Chicago/Turabian StyleFang, Bo, Ming Liang, Guitao Yang, Yuqin Ye, Hongyu Xu, Xiaosheng He, and Jason H. Huang. 2014. "Expression of S100A6 in Rat Hippocampus after Traumatic Brain Injury Due to Lateral Head Acceleration" International Journal of Molecular Sciences 15, no. 4: 6378-6390. https://doi.org/10.3390/ijms15046378
APA StyleFang, B., Liang, M., Yang, G., Ye, Y., Xu, H., He, X., & Huang, J. H. (2014). Expression of S100A6 in Rat Hippocampus after Traumatic Brain Injury Due to Lateral Head Acceleration. International Journal of Molecular Sciences, 15(4), 6378-6390. https://doi.org/10.3390/ijms15046378