Asymmetric Dimethylarginine in Chronic Obstructive Pulmonary Disease (ADMA in COPD)

Abstract
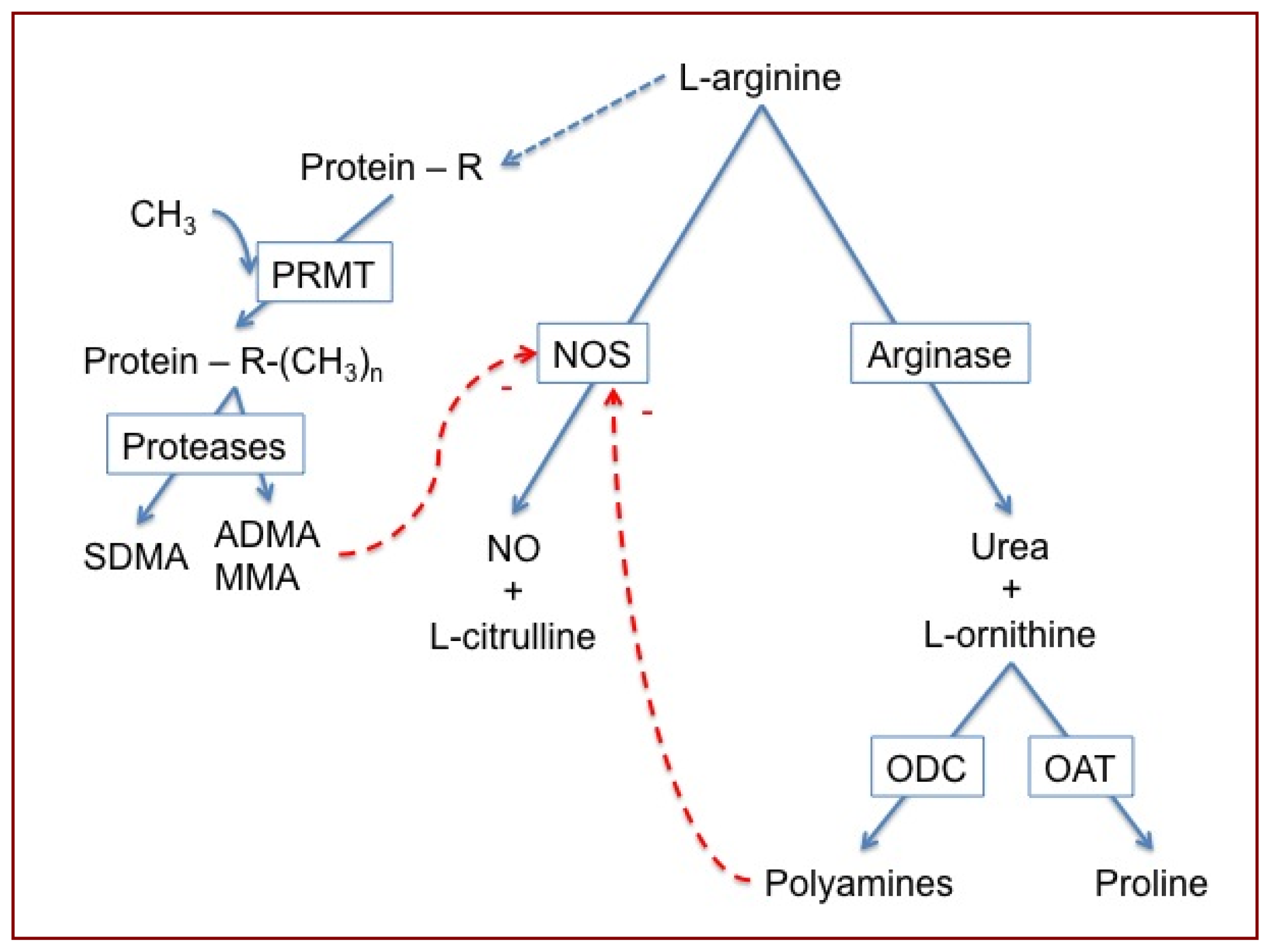
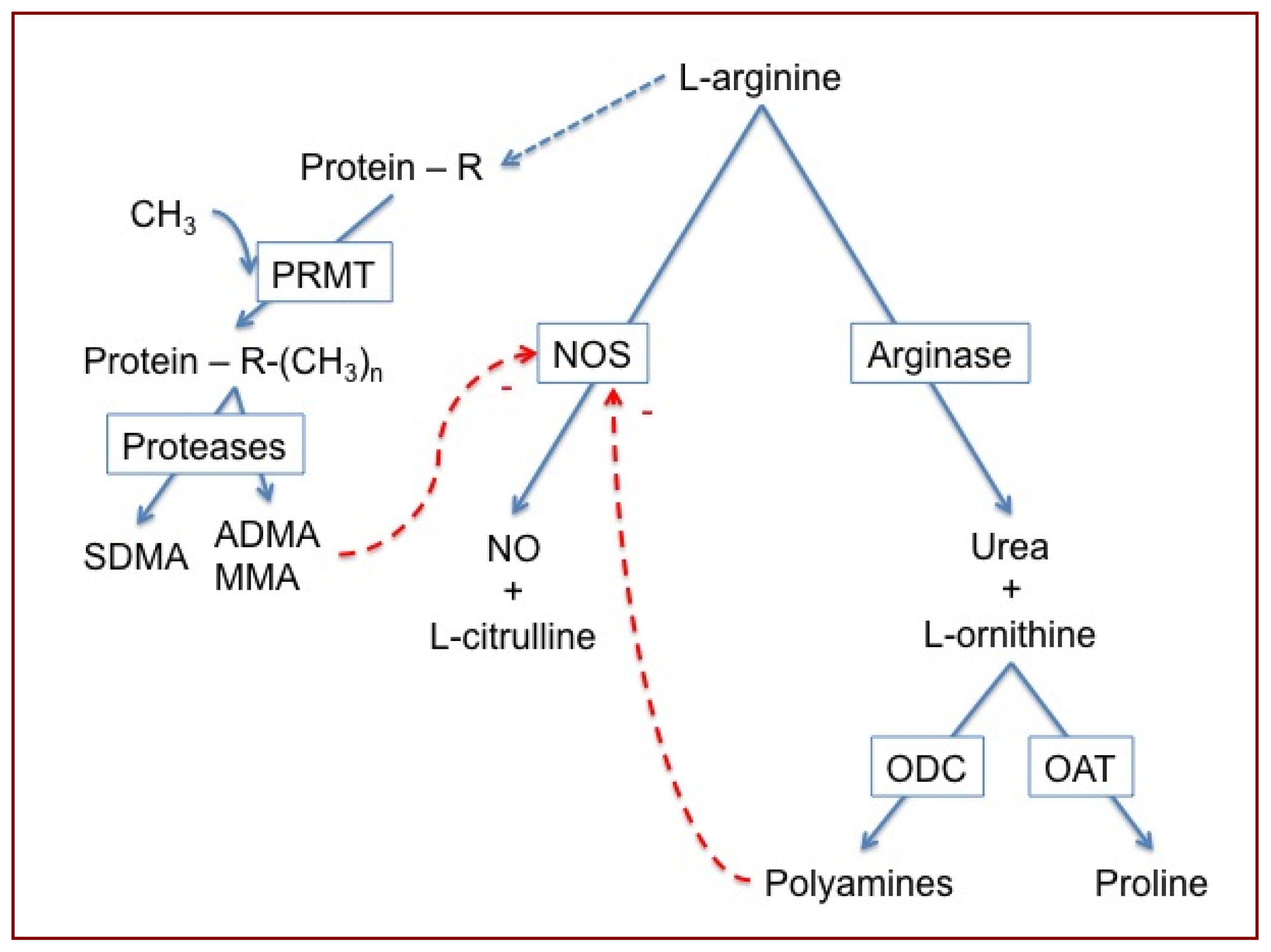
:1. Introduction
2. Results and Discussion
2.1. Patient Demographics and Lung Function
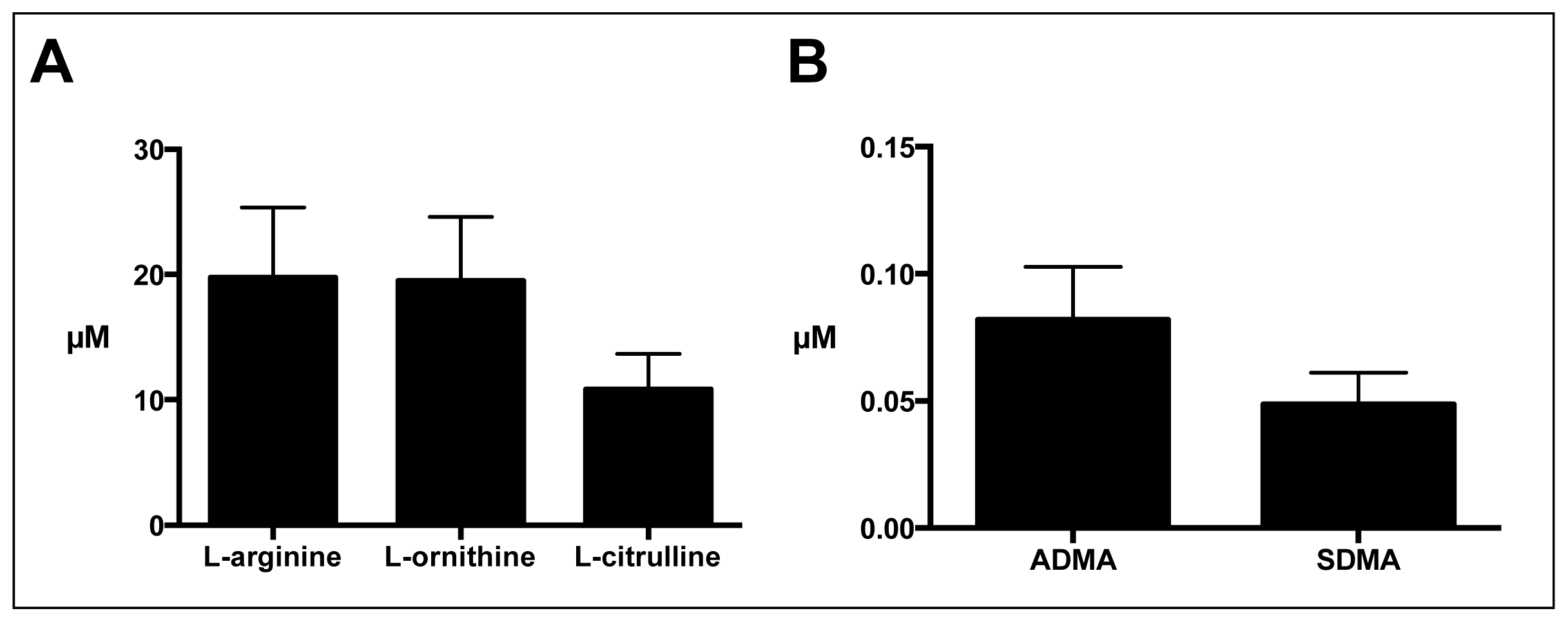
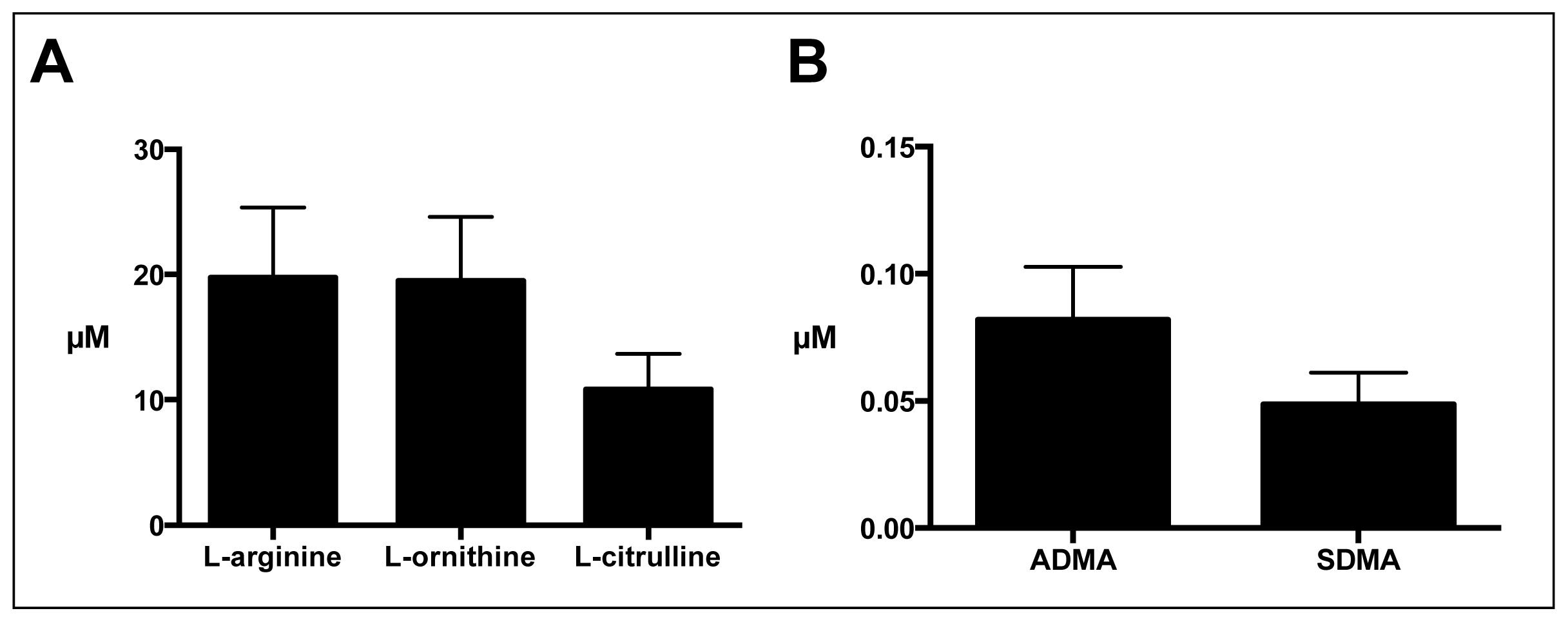
2.2. Sputum l-Arginine Metabolites and Indices of Arginase and NO Synthase Activity
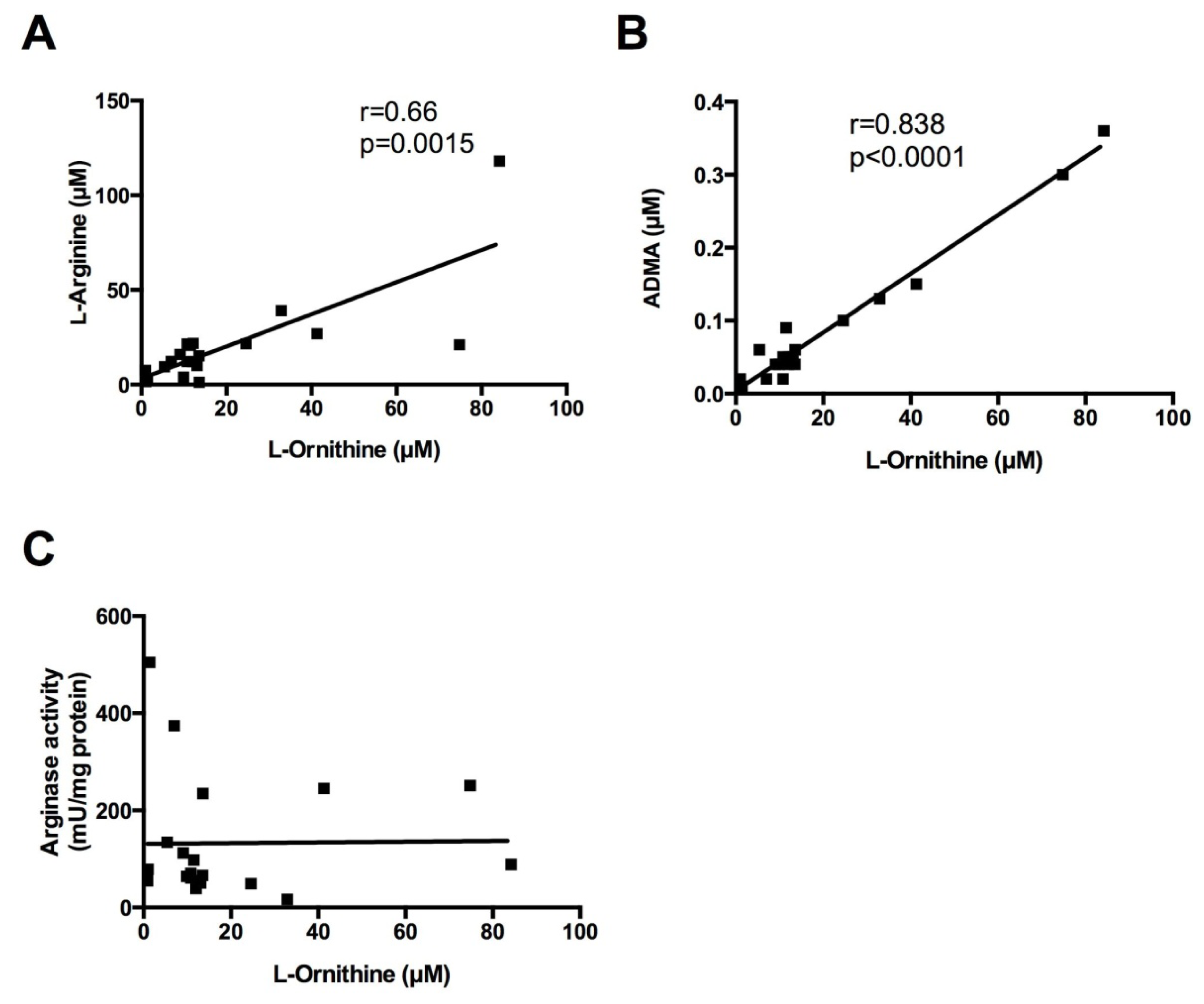
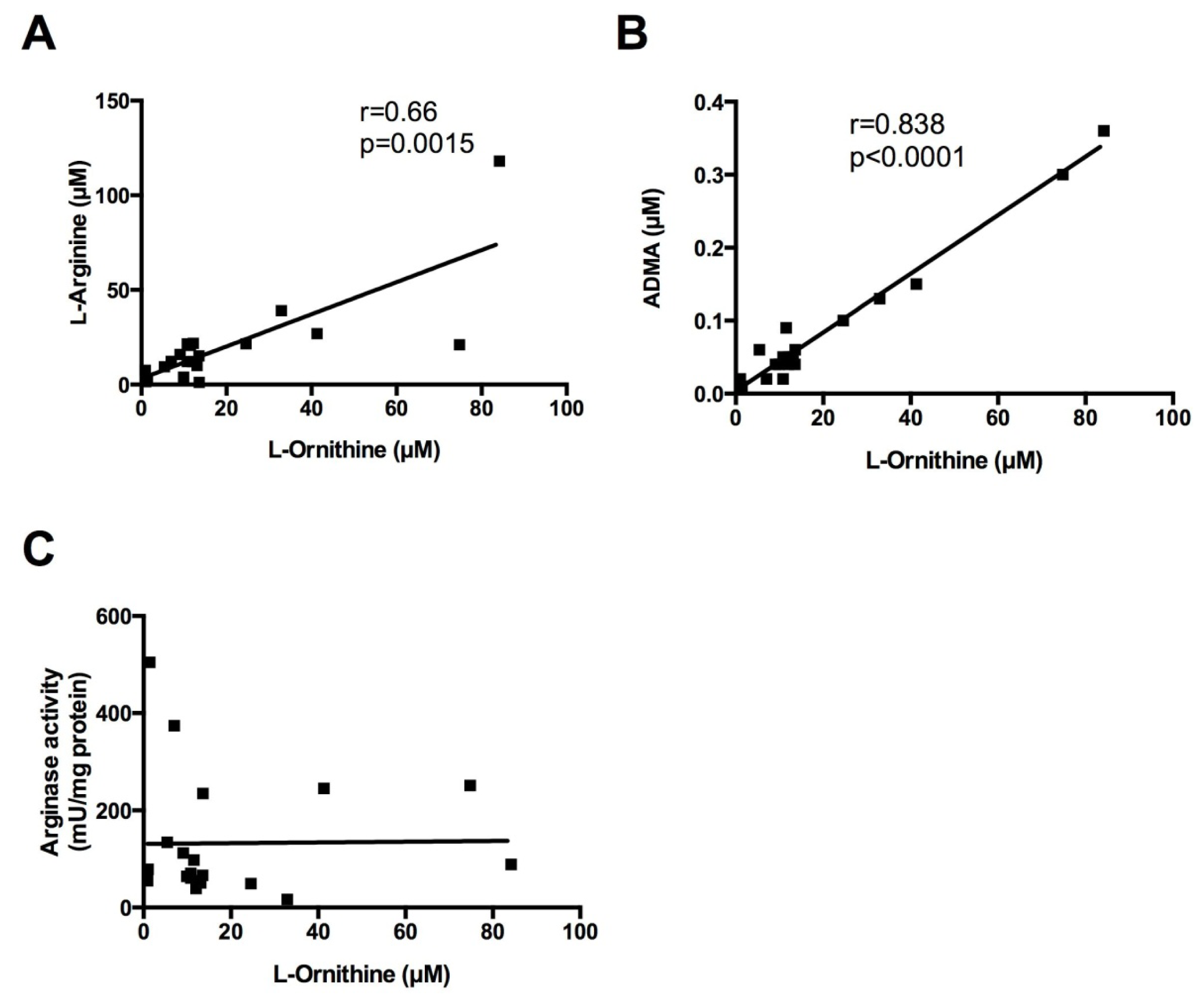
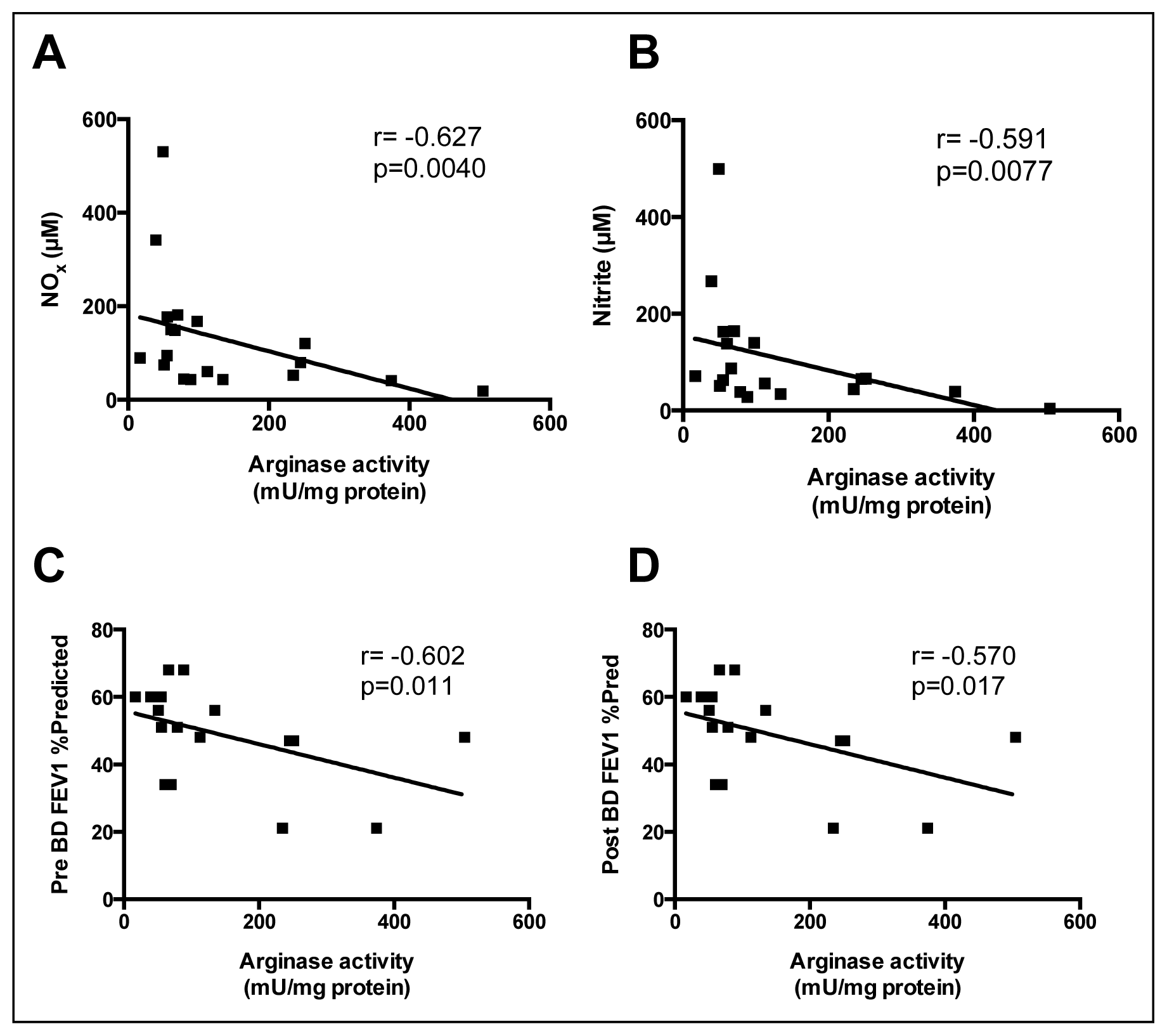
2.3. Correlations between Sputum Measures of l-Arginine Metabolites and Arginase Activity
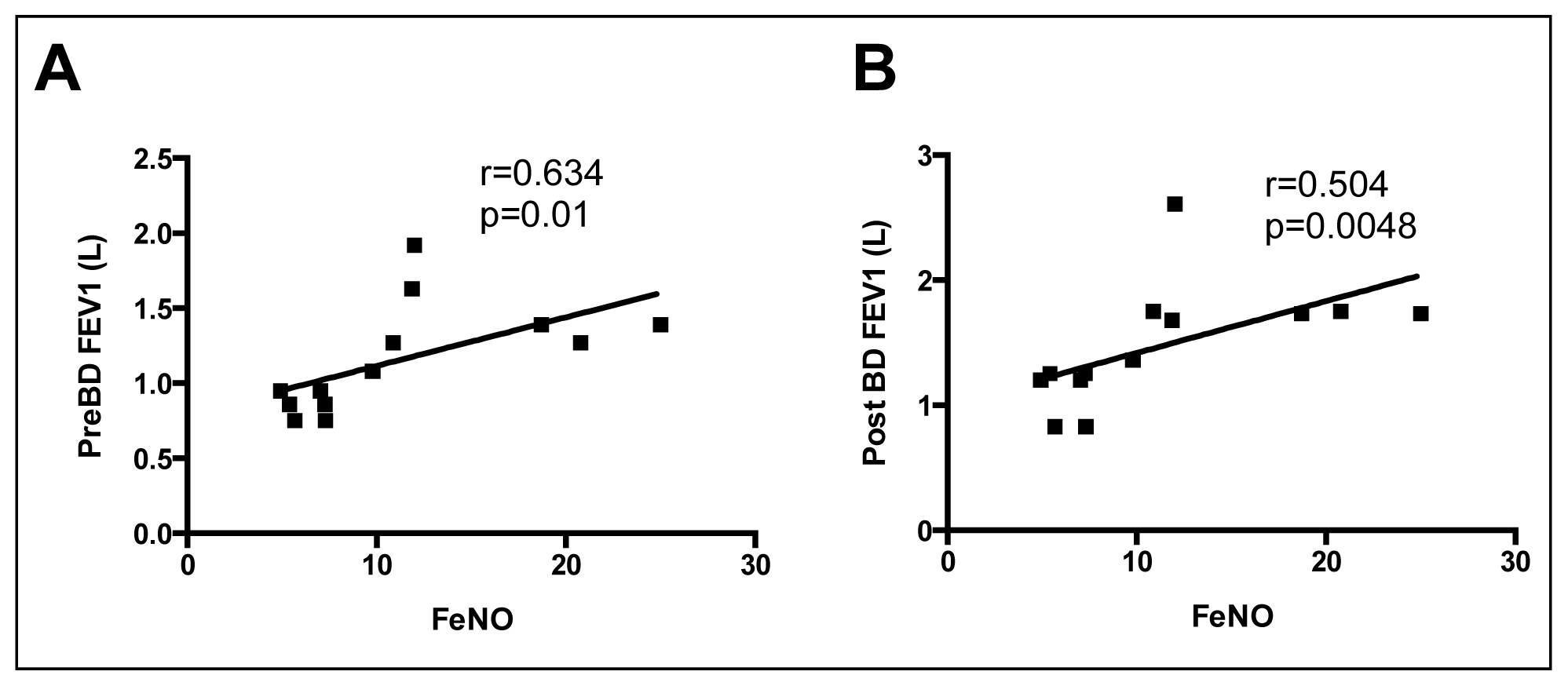
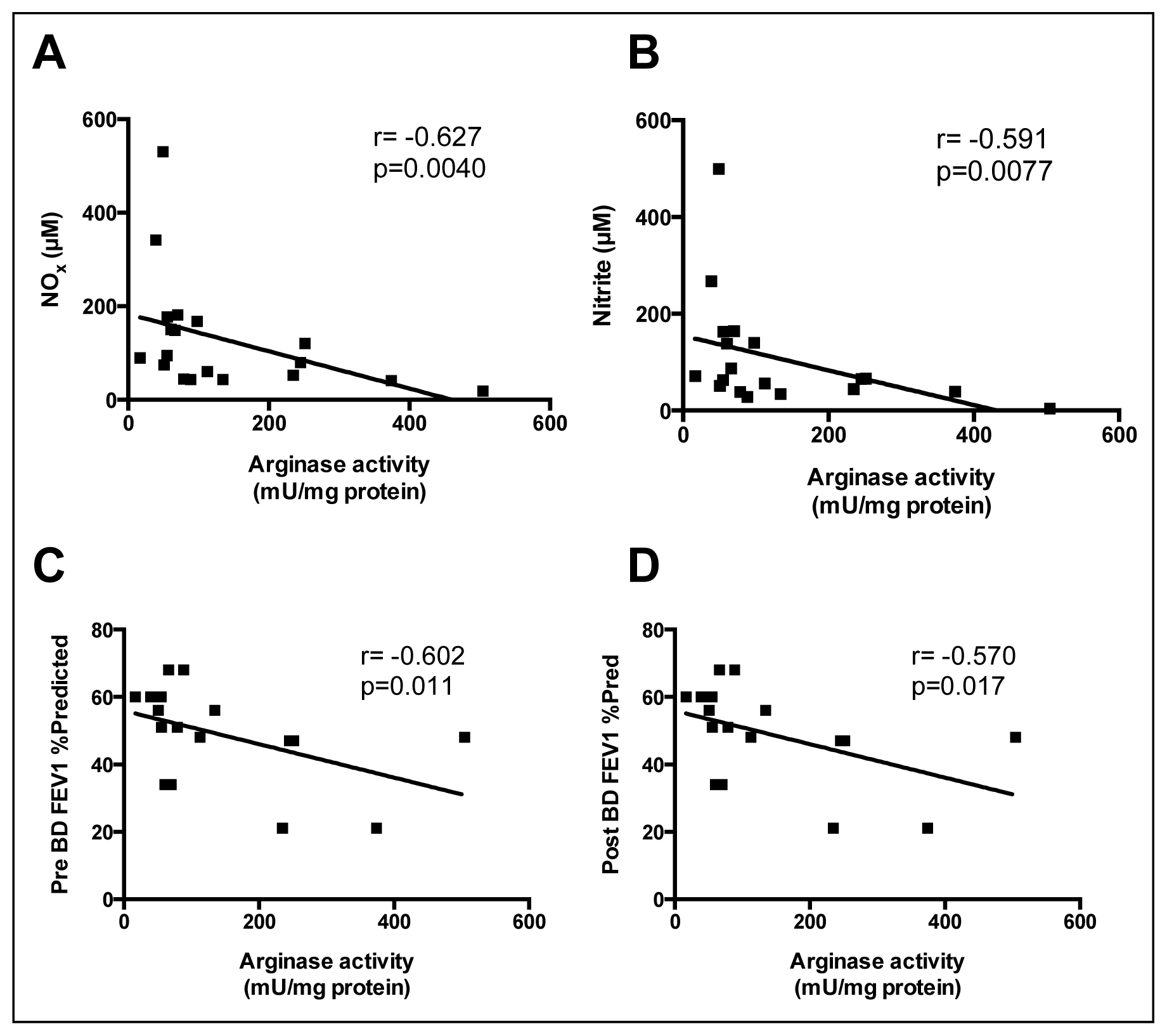
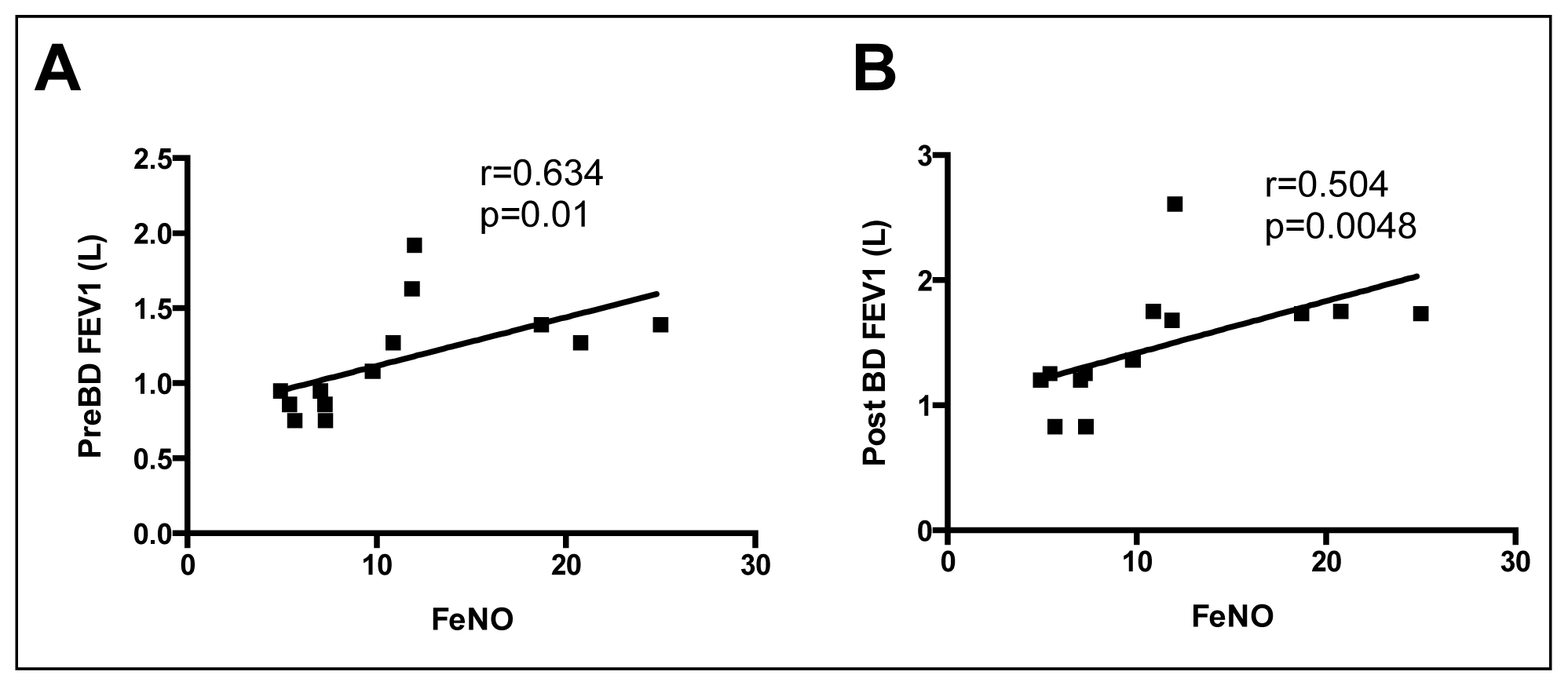
2.4. Correlations between Sputum l-Arginine Metabolites, Expired NO, Arginase Activity and Lung Function
3. Experimental Section
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsJAS: Analysis and interpretation of data, drafting and revision of the manuscript; MLD: Patient recruitment, acquisition and analysis of data, and revising of the manuscript; AWY: Patient recruitment, acquisition and analysis of data, and revision of the manuscript; PS: Drafting and revision of the manuscript; GMG: Study design, patient recruitment, acquisition and analysis of data, drafting and revision of the manuscript; HG: Study design, analysis and interpretation of data, drafting and revision of the manuscript.
References
- North, M.L.; Scott, J.A. l-Arginine metabolism in the lung: Reciprocal regulation of the NOS and arginase pathways. Open Nitric Oxide J 2011, 3 Suppl 1-M7, 48–54. [Google Scholar]
- Maarsingh, H.; Pera, T.; Meurs, H. Arginase and pulmonary diseases. Naunyn Schmiedebergs Arch. Pharmacol 2008, 378, 171–184. [Google Scholar]
- Grasemann, H.; Al-Saleh, S.; Scott, J.A.; Shehnaz, D.; Mehl, A.; Amin, R.; Rafii, M.; Pencharz, P.; Belik, J.; Ratjen, F. Asymmetric dimethylarginine contributes to airway nitric oxide deficiency in patients with cystic fibrosis. Am. J. Respir. Crit. Care Med 2011, 183, 1363–1368. [Google Scholar]
- Scott, J.A.; North, M.L.; Rafii, M.; Huang, H.; Pencharz, P.; Subbarao, P.; Belik, J.; Grasemann, H. Asymmetric dimethylarginine is increased in asthma. Am. J. Respir. Crit. Care Med 2011, 184, 779–785. [Google Scholar]
- Scott, J.A.; Grasemann, H. Asymmetric dimethylarginine: A disease marker for asthma? Chest 2013, 144, 367–368. [Google Scholar]
- Grasemann, H.; Schwiertz, R.; Matthiesen, S.; Racke, K.; Ratjen, F. Increased arginase activity in cystic fibrosis airways. Am. J. Respir. Crit. Care Med 2005, 172, 1523–1528. [Google Scholar]
- North, M.L.; Khanna, N.; Marsden, P.A.; Grasemann, H.; Scott, J.A. Functionally important role for arginase 1 in the airway hyperresponsiveness of asthma. Am. J. Physiol. Lung Cell. Mol. Physiol 2009, 296, L911–L920. [Google Scholar]
- Zimmermann, N.; King, N.E.; Laporte, J.; Yang, M.; Mishra, A.; Pope, S.M.; Muntel, E.E.; Witte, D.P.; Pegg, A.A.; Foster, P.S.; et al. Dissection of experimental asthma with DNA microarray analysis identifies arginase in asthma pathogenesis. J. Clin. Investig 2003, 111, 1863–1874. [Google Scholar]
- Zimmermann, N.; Rothenberg, M.E. The arginine-arginase balance in asthma and lung inflammation. Eur. J. Pharmacol 2006, 533, 253–262. [Google Scholar]
- Gaston, B.; Drazen, J.M.; Loscalzo, J.; Stamler, J.S. The biology of nitrogen oxides in the airways. Am. J. Respir. Crit. Care Med 1994, 149, 538–551. [Google Scholar]
- Morris, C.R. Arginine and asthma. Nestle Nutr. Inst. Workshop Ser 2013, 77, 1–15. [Google Scholar]
- Meurs, H.; McKay, S.; Maarsingh, H.; Hamer, M.A.; Macic, L.; Molendijk, N.; Zaagsma, J. Increased arginase activity underlies allergen-induced deficiency of cNOS-derived nitric oxide and airway hyperresponsiveness. Br. J. Pharmacol 2002, 136, 391–398. [Google Scholar]
- Meurs, H.; Maarsingh, H.; Zaagsma, J. Arginase and asthma: Novel insights into nitric oxide homeostasis and airway hyperresponsiveness. Trends Pharmacol. Sci 2003, 24, 450–455. [Google Scholar]
- North, M.L.; Meurs, H.; Zaagsma, J.; Scott, J.A.; Maarsingh, H. Arginase in asthma—Recent developments in animal and human studies. Open Nitric Oxide J 2010, 2, 20–36. [Google Scholar]
- Grasemann, H.; Shehnaz, D.; Enomoto, M.; Leadley, M.; Belik, J.; Ratjen, F. l-ornithine derived polyamines in cystic fibrosis airways. PLoS One 2012, 7, e46618. [Google Scholar]
- North, M.L.; Grasemann, H.; Khanna, N.; Inman, M.D.; Gauvreau, G.M.; Scott, J.A. Increased ornithine-derived polyamines cause airway hyperresponsiveness in a mouse model of asthma. Am. J. Respir. Cell. Mol. Biol 2013, 48, 694–702. [Google Scholar]
- Tanaka, H.; Masuda, T.; Tokuoka, S.; Komai, M.; Nagao, K.; Takahashi, Y.; Nagai, H. The effect of allergen-induced airway inflammation on airway remodeling in a murine model of allergic asthma. Inflamm. Res 2001, 50, 616–624. [Google Scholar]
- Holguin, F.; Comhair, S.A.; Hazen, S.L.; Powers, R.W.; Khatri, S.S.; Bleecker, E.R.; Busse, W.W.; Calhoun, W.J.; Castro, M.; Fitzpatrick, A.M.; et al. An association between l-arginine/asymmetric dimethyl arginine balance, obesity, and the age of asthma onset phenotype. Am. J. Respir. Crit. Care Med 2013, 187, 153–159. [Google Scholar]
- Lara, A.; Khatri, S.B.; Wang, Z.; Comhair, S.A.; Xu, W.; Dweik, R.A.; Bodine, M.; Levison, B.S.; Hammel, J.; Bleecker, E.; et al. Alterations of the arginine metabolome in asthma. Am. J. Respir. Crit. Care Med 2008, 178, 673–681. [Google Scholar]
- Vallance, P.; Leone, A.; Calver, A.; Collier, J.; Moncada, S. Accumulation of an endogenous inhibitor of nitric oxide synthesis in chronic renal failure. Lancet 1992, 339, 572–575. [Google Scholar]
- Schwedhelm, E.; Boger, R.H. The role of asymmetric and symmetric dimethylarginines in renal disease. Nat. Rev. Nephrol 2011, 7, 275–285. [Google Scholar]
- Kurz, K.; Teerlink, T.; Sarcletti, M.; Weiss, G.; Zangerle, R.; Fuchs, D. Plasma concentrations of the cardiovascular risk factor asymmetric dimethylarginine (ADMA) are increased in patients with HIV-1 infection and correlate with immune activation markers. Pharmacol. Res 2009, 60, 508–514. [Google Scholar]
- Boger, R.H.; Maas, R.; Schulze, F.; Schwedhelm, E. Asymmetric dimethylarginine (ADMA) as a prospective marker of cardiovascular disease and mortality—An update on patient populations with a wide range of cardiovascular risk. Pharmacol. Res 2009, 60, 481–487. [Google Scholar]
- Vestbo, J.; Hurd, S.S.; Agusti, A.G.; Jones, P.W.; Vogelmeier, C.; Anzueto, A.; Barnes, P.J.; Fabbri, L.M.; Martinez, F.J.; Nishimura, M.; et al. Global strategy for the diagnosis, management, and prevention of chronic obstructive pulmonary disease: GOLD executive summary. Am. J. Respir. Crit. Care Med 2013, 187, 347–365. [Google Scholar]
- Bergeron, C.; Boulet, L.P.; Page, N.; Laviolette, M.; Zimmermann, N.; Rothenberg, M.E.; Hamid, Q. Influence of cigarette smoke on the arginine pathway in asthmatic airways: Increased expression of arginase I. J. Allergy Clin. Immunol 2007, 119, 391–397. [Google Scholar]
- Tadie, J.M.; Henno, P.; Leroy, I.; Danel, C.; Naline, E.; Faisy, C.; Riquet, M.; Levy, M.; Israel-Biet, D.; Delclaux, C. Role of nitric oxide synthase/arginase balance in bronchial reactivity in patients with chronic obstructive pulmonary disease. Am. J. Physiol. Lung Cell. Mol. Physiol 2008, 294, L489–L497. [Google Scholar]
- Scrimini, S.; Pons, J.; Agusti, A.; Soriano, J.B.; Cosio, B.G.; Torrecilla, J.A.; Nunez, B.; Cordova, R.; Iglesias, A.; Jahn, A.; et al. Differential effects of smoking and COPD upon circulating myeloid derived suppressor cells. Respir Med 2013, 107, 1895–1903. [Google Scholar]
- Guzman-Grenfell, A.; Nieto-Velazquez, N.; Torres-Ramos, Y.; Montoya-Estrada, A.; Ramirez-Venegas, A.; Ochoa-Cautino, L.; Flores-Trujillo, F.; Hicks, J.J. Increased platelet and erythrocyte arginase activity in chronic obstructive pulmonary disease associated with tobacco or wood smoke exposure. J. Investig. Med 2011, 59, 587–592. [Google Scholar]
- Hodge, S.; Matthews, G.; Mukaro, V.; Ahern, J.; Shivam, A.; Hodge, G.; Holmes, M.; Jersmann, H.; Reynolds, P.N. Cigarette smoke-induced changes to alveolar macrophage phenotype and function are improved by treatment with procysteine. Am. J. Respir. Cell. Mol. Biol 2011, 44, 673–681. [Google Scholar]
- Pera, T.; Zuidhof, A.B.; Smit, M.; Menzen, M.H.; Klein, T.; Flik, G.; Zaagsma, J.; Meurs, H.; Maarsingh, H. Arginase inhibition prevents inflammation and remodeling in a guinea pig model of COPD. J. Pharmacol. Exp. Ther 2014. [Google Scholar] [CrossRef]
- Grasemann, H.; Tullis, E.; Ratjen, F. A randomized controlled trial of inhaled l-arginine in patients with cystic fibrosis. J. Cyst. Fibros 2013, 12, 468–474. [Google Scholar]
- Zakrzewicz, D.; Eickelberg, O. From arginine methylation to ADMA: A novel mechanism with therapeutic potential in chronic lung diseases. BMC Pulm. Med 2009. [Google Scholar] [CrossRef]
- Gaston, B. The biochemistry of asthma. Biochim. Biophys. Acta 2011, 1810, 1017–1024. [Google Scholar]
- Maarsingh, H.; Zaagsma, J.; Meurs, H. Arginine homeostasis in allergic asthma. Eur. J. Pharmacol 2008, 585, 375–384. [Google Scholar]
- See, K.C.; Christiani, D.C. Normal values and thresholds for the clinical interpretation of exhaled nitric oxide levels in the US general population: Results from the National Health and Nutrition Examination Survey 2007–2010. Chest 2013, 143, 107–116. [Google Scholar]
- Dweik, R.A.; Boggs, P.B.; Erzurum, S.C.; Irvin, C.G.; Leigh, M.W.; Lundberg, J.O.; Olin, A.C.; Plummer, A.L.; Taylor, D.R. An official ATS clinical practice guideline: Interpretation of exhaled nitric oxide levels (FENO) for clinical applications. Am. J. Respir. Crit. Care Med 2011, 184, 602–615. [Google Scholar]
- ATS/ERS recommendations for standardized procedures for the online and offline measurement of exhaled lower respiratory nitric oxide and nasal nitric oxide, 2005. Am. J. Respir. Crit. Care Med 2005, 171, 912–930.
- Brusasco, V.; Crapo, R.; Viegi, G. Coming together: The ATS/ERS consensus on clinical pulmonary function testing. Eur. Respir. J 2005, 26, 1–2. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measure | Mean ± SEM n |
|---|---|
| Age (years) (mean ± SEM) | 62.3 ± 2.6 |
| Height (cm) (mean ± SEM) | 174.0 ± 1.8 |
| Weight (kg) (mean ± SEM) | 76.4 ± 3.0 |
| Gender (m/f) | 8/2 |
| Current/Ex smoker | 4/6 |
| Pack years | 37 ± 5 |
| COPD Diagnosis (years) (mean ± SEM) | 7 ± 2.6 |
| Inhaled Glucocorticosteroids (y/n) | 6/4 |
| Body Surface Area (m2) (mean ± SEM) | 1.90 ± 0.04 |
| Measure | Mean ± SEM |
|---|---|
| Arginase activity (mU/mg protein) | 132.5 ± 28.4 |
| FeNO (ppb) | 17 ± 4 |
| Sputum NOx (μM) | 194 ± 70 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Scott, J.A.; Duongh, M.; Young, A.W.; Subbarao, P.; Gauvreau, G.M.; Grasemann, H. Asymmetric Dimethylarginine in Chronic Obstructive Pulmonary Disease (ADMA in COPD). Int. J. Mol. Sci. 2014, 15, 6062-6071. https://doi.org/10.3390/ijms15046062
Scott JA, Duongh M, Young AW, Subbarao P, Gauvreau GM, Grasemann H. Asymmetric Dimethylarginine in Chronic Obstructive Pulmonary Disease (ADMA in COPD). International Journal of Molecular Sciences. 2014; 15(4):6062-6071. https://doi.org/10.3390/ijms15046062
Chicago/Turabian StyleScott, Jeremy A., MyLinh Duongh, Aaron W. Young, Padmaja Subbarao, Gail M. Gauvreau, and Hartmut Grasemann. 2014. "Asymmetric Dimethylarginine in Chronic Obstructive Pulmonary Disease (ADMA in COPD)" International Journal of Molecular Sciences 15, no. 4: 6062-6071. https://doi.org/10.3390/ijms15046062
APA StyleScott, J. A., Duongh, M., Young, A. W., Subbarao, P., Gauvreau, G. M., & Grasemann, H. (2014). Asymmetric Dimethylarginine in Chronic Obstructive Pulmonary Disease (ADMA in COPD). International Journal of Molecular Sciences, 15(4), 6062-6071. https://doi.org/10.3390/ijms15046062