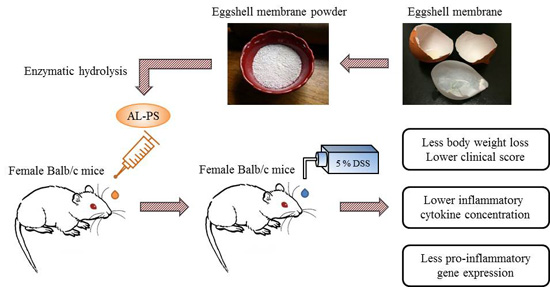
Hydrolysate from Eggshell Membrane Ameliorates Intestinal Inflammation in Mice

Abstract
:
1. Introduction
2. Results and Discussion
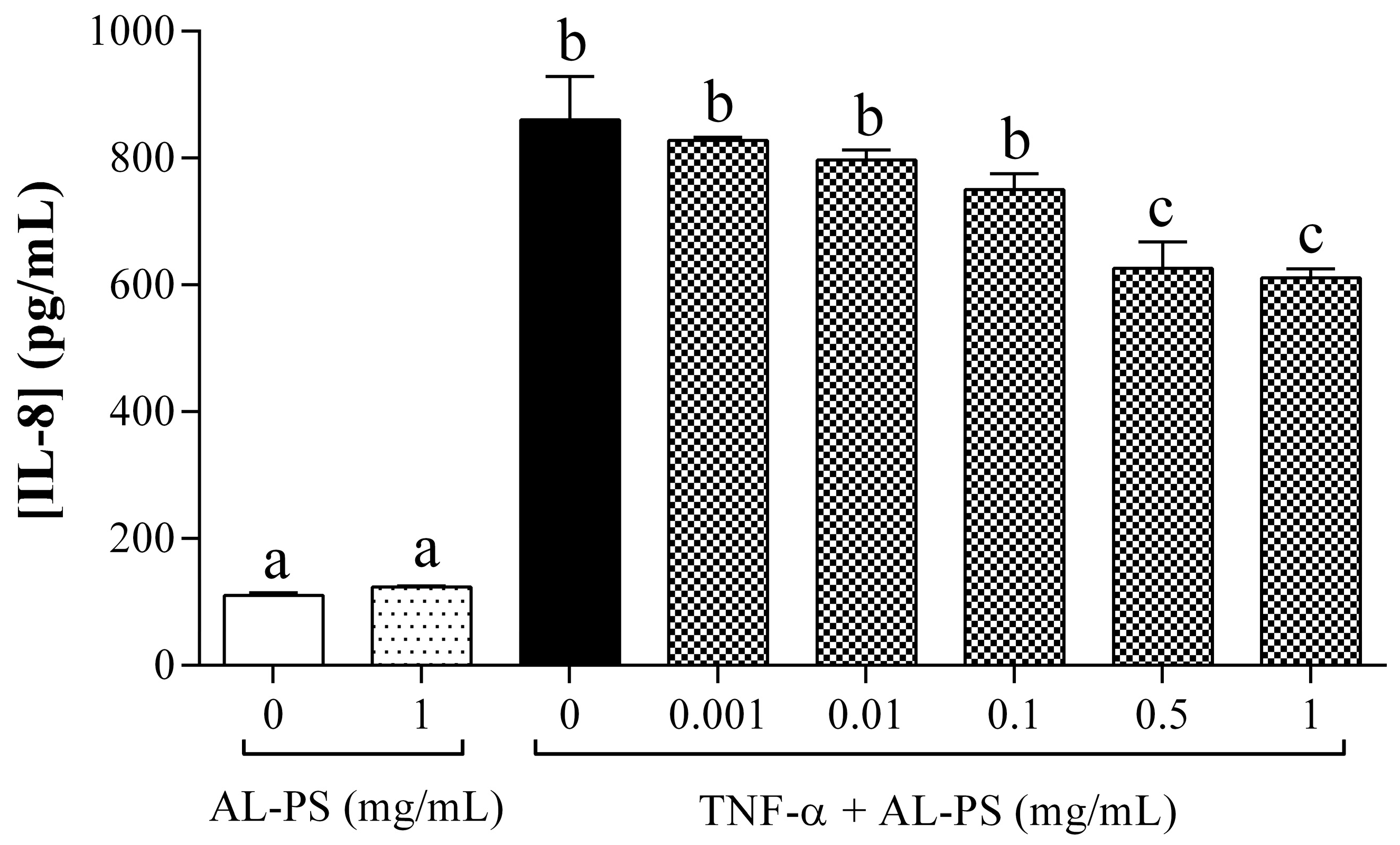
2.1. Anti-Inflammatory Activity of AL-PS in Vitro
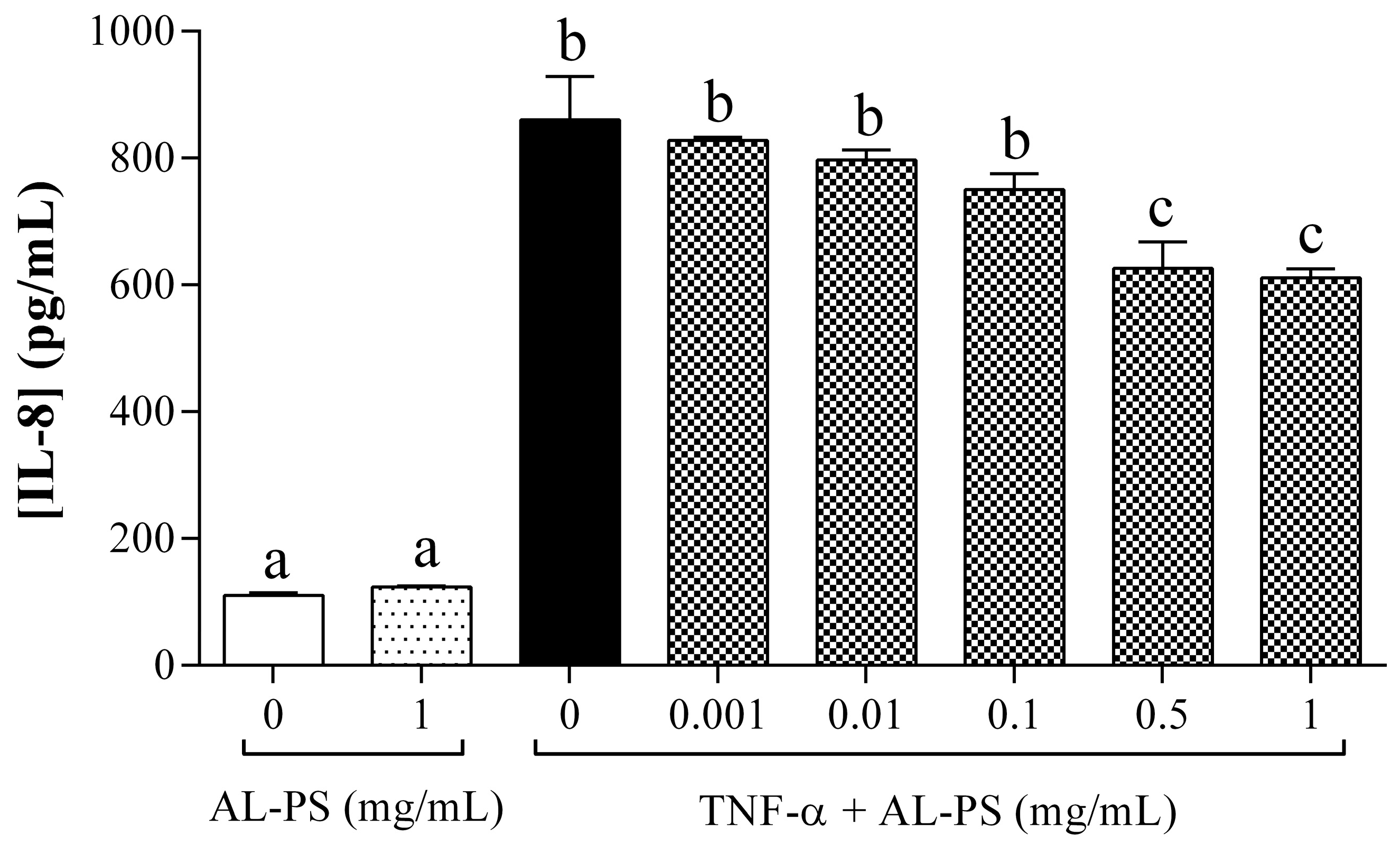
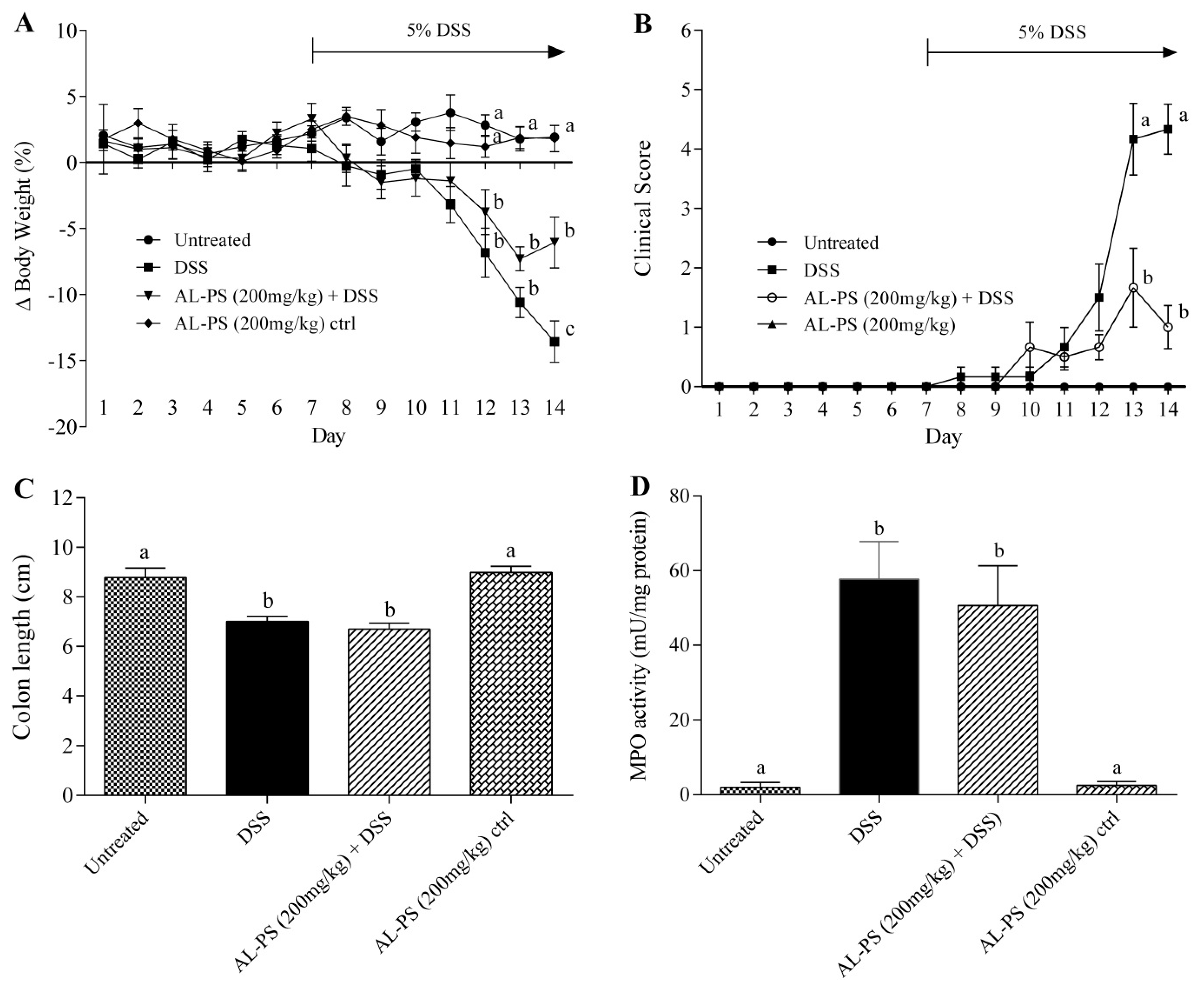
2.2. Attenuation of Inflammation Signs in Vivo
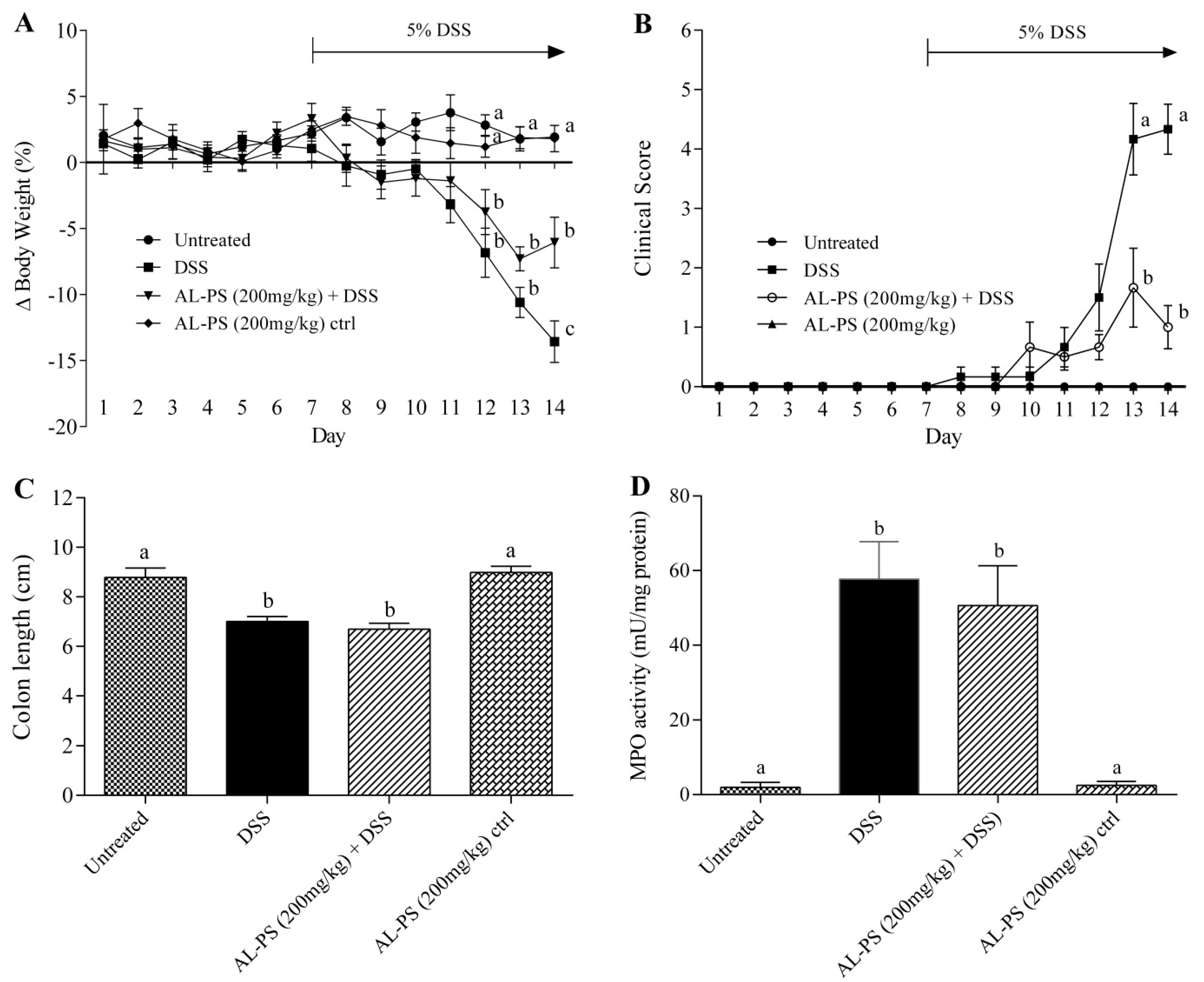
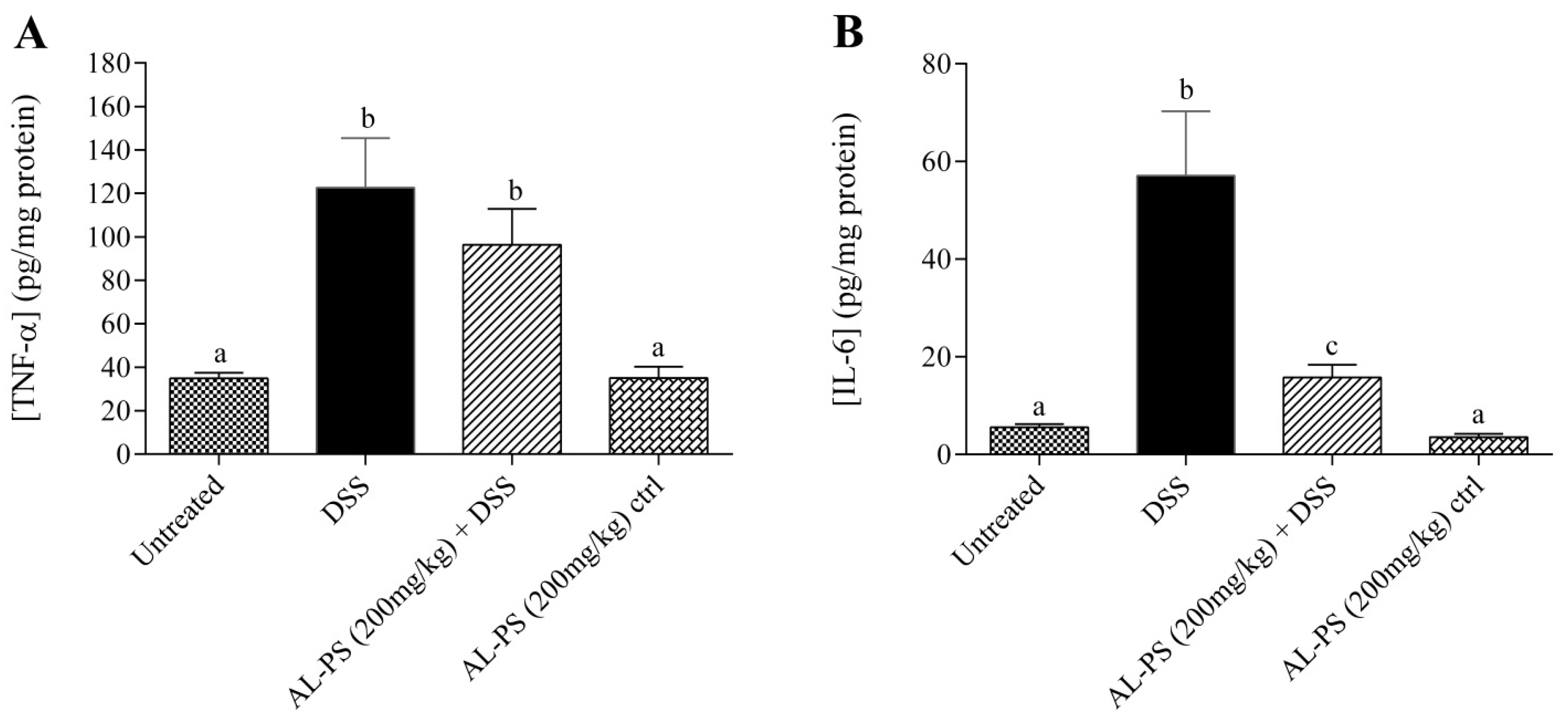
2.3. Cytokine Concentration
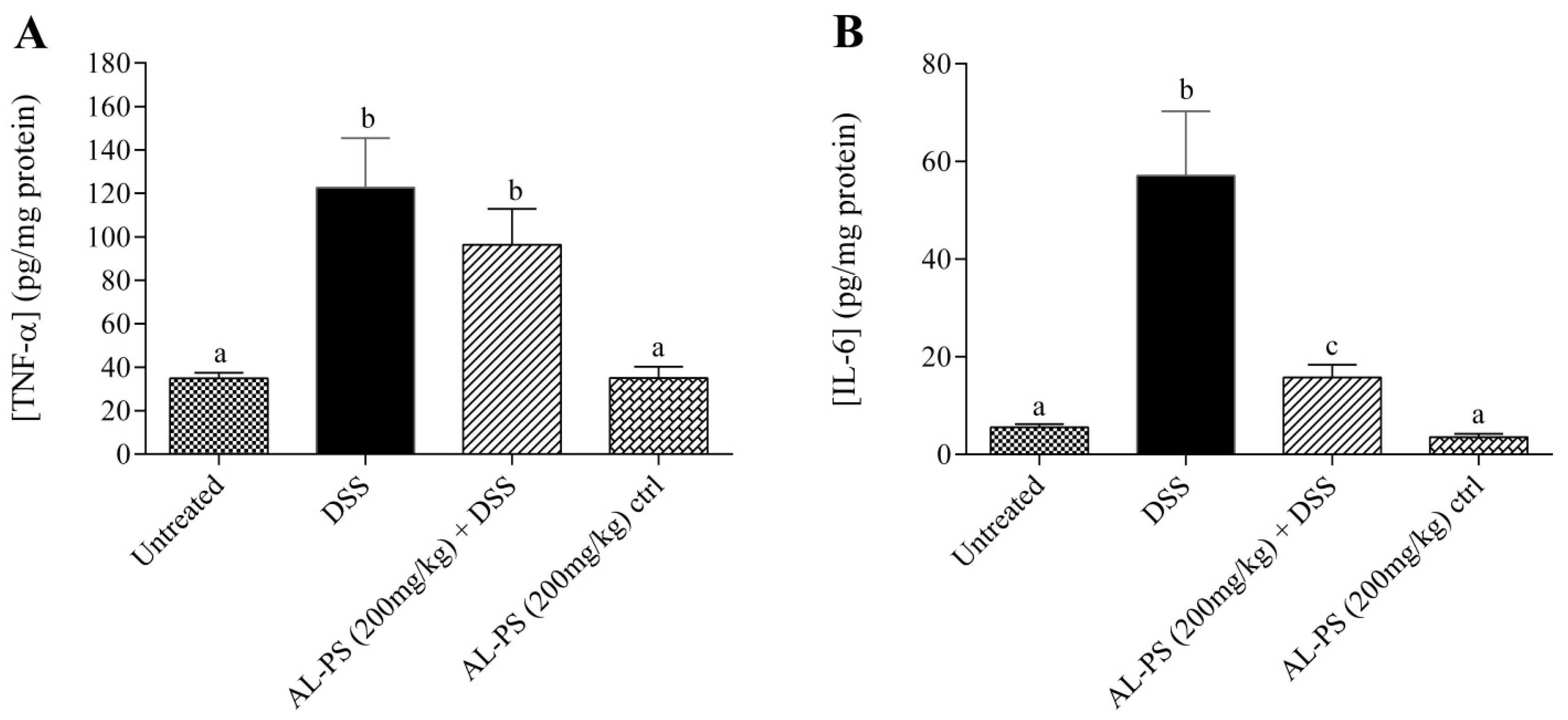
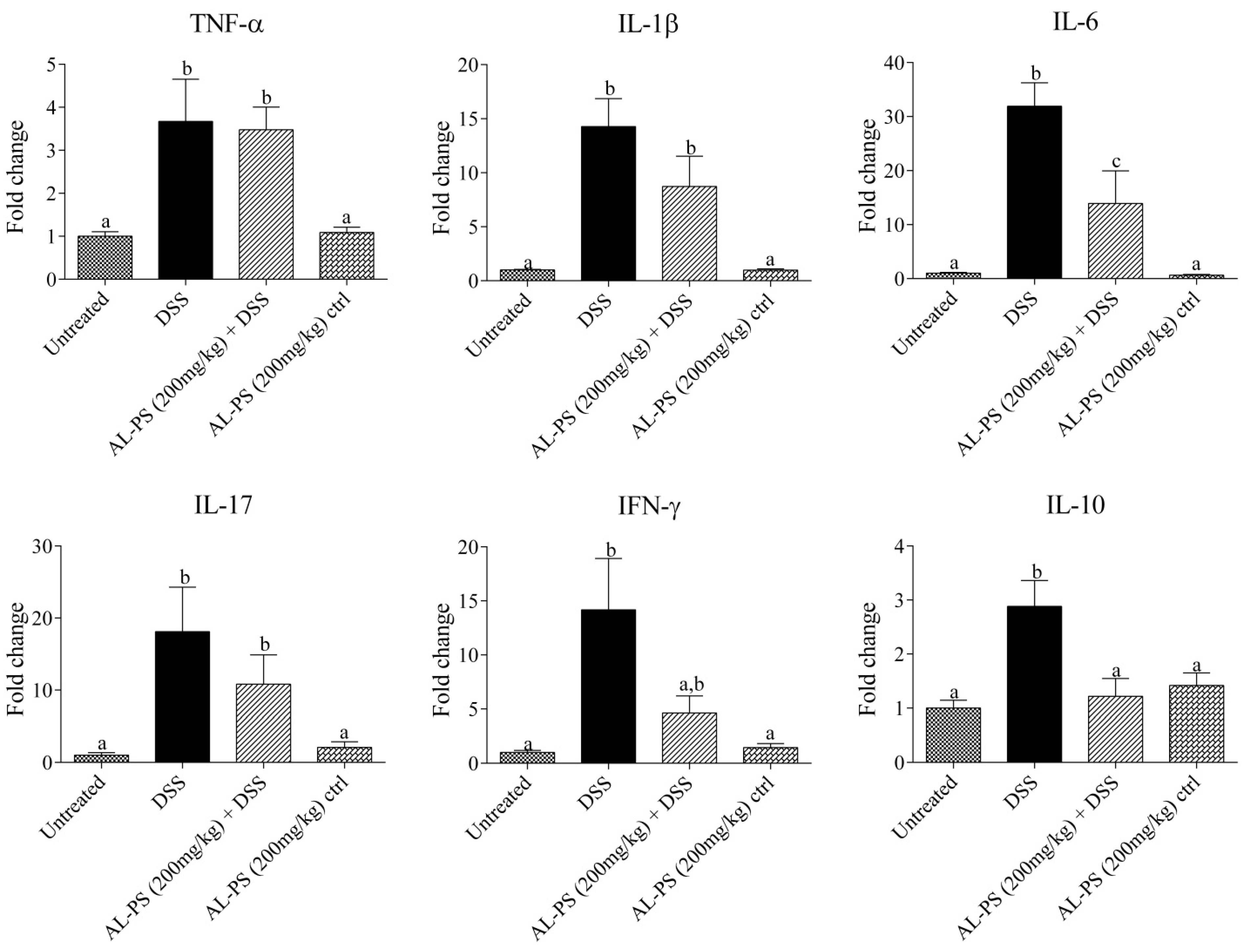
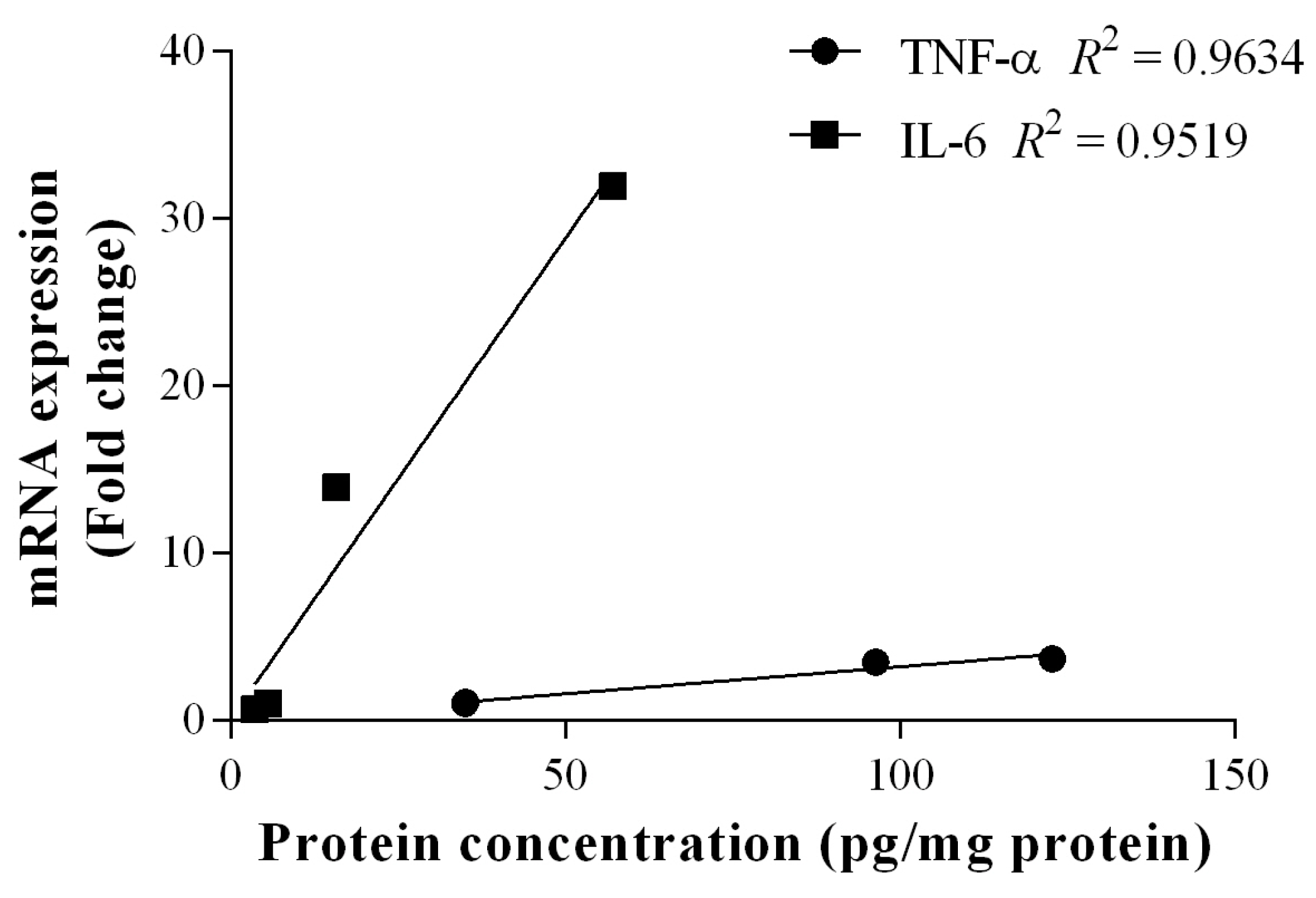
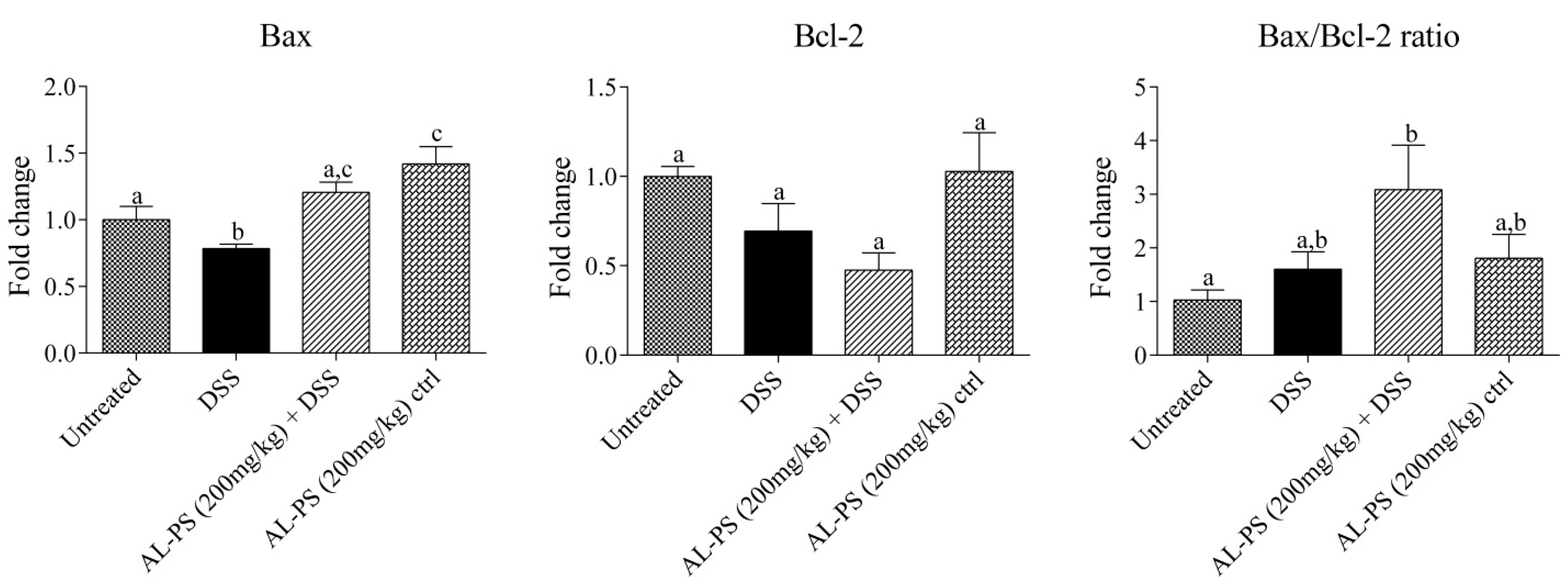
2.4. Gene Expression
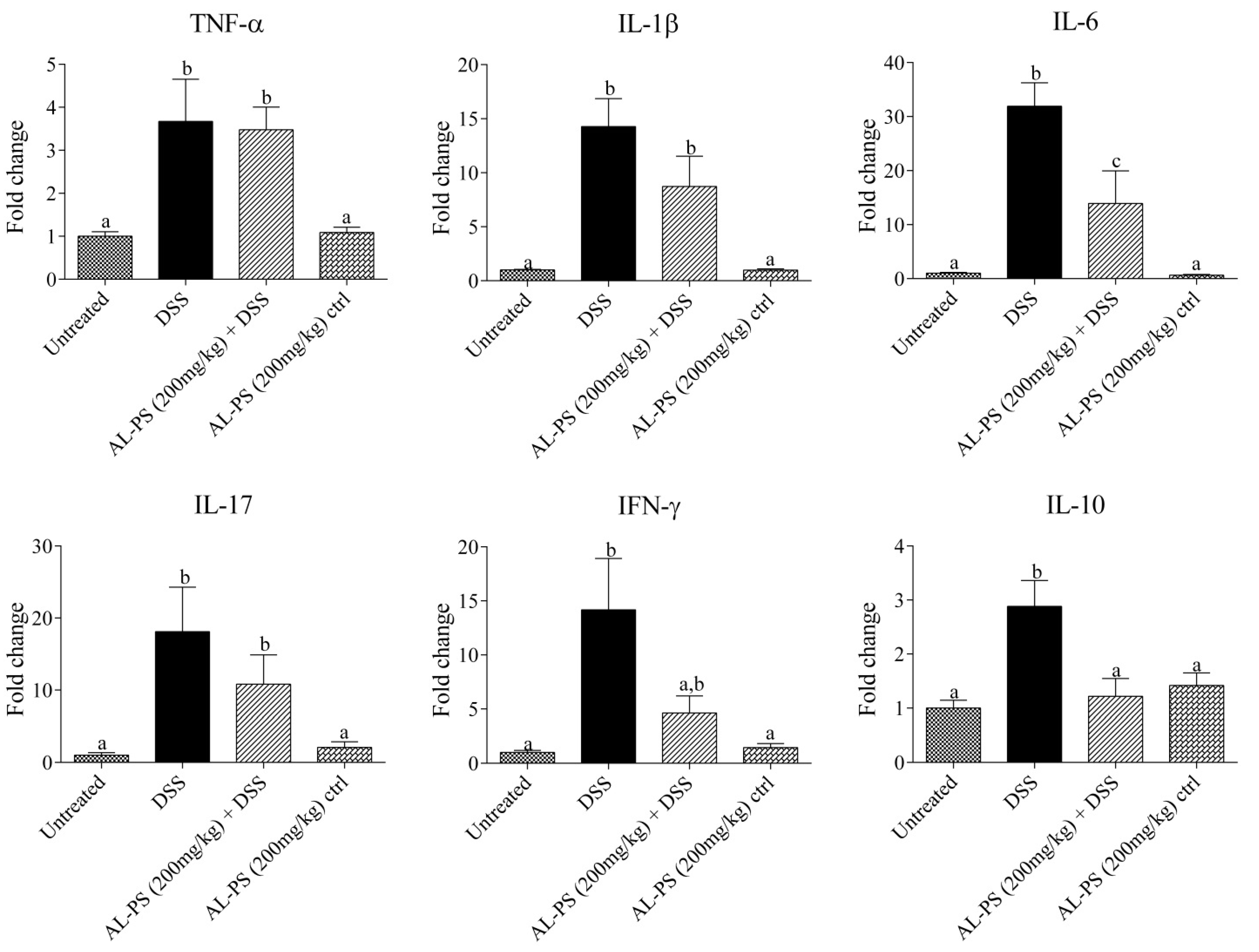
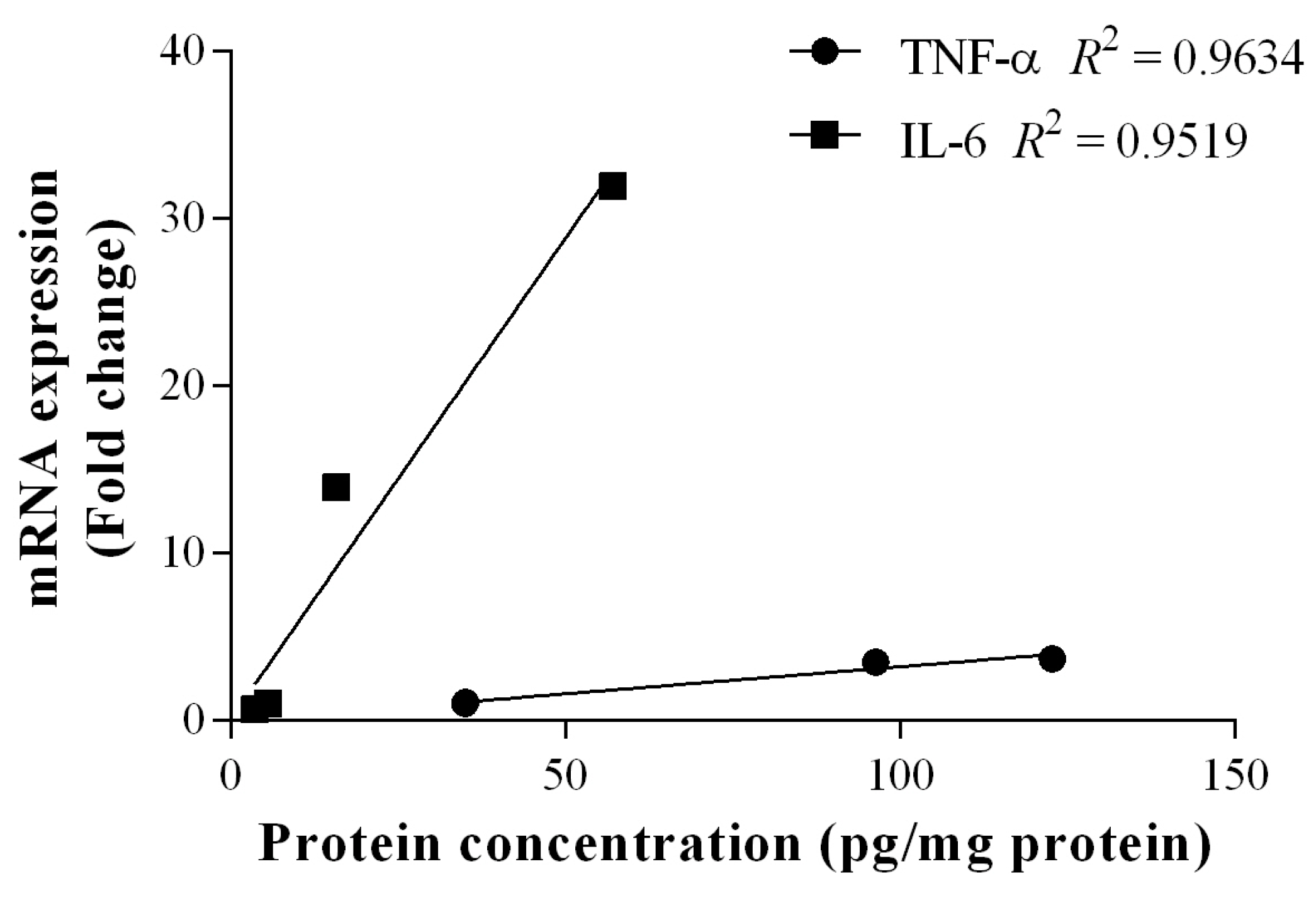
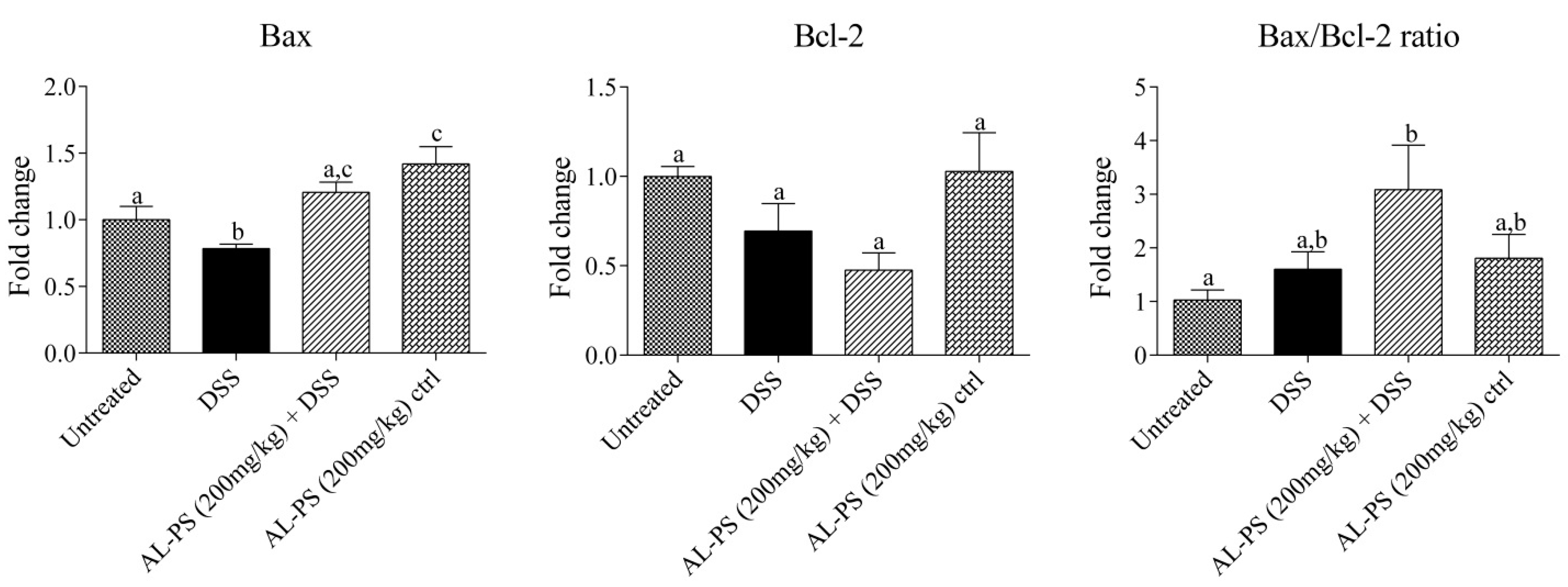
2.5. Discussion
3. Experimental Section
3.1. Materials
3.2. Preparation of ESM Hydro Lysate
3.3. Cell Culture
3.4. In Vitro Induction of Inflammation
3.5. Determination of IL-8 Secretion
3.6. Animals and Experimental Design
3.7. Clinical Evaluation of Colitis
3.8. Determination of TNF-α and IL-6 Concentration in Tissue
3.9. Determination of MPO Activity
3.10. RNA Isolation and Determination of Gene Expression in the Colon

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Gene | Forward Primer (5'-3') | Reverse Primer (5'-3') |
|---|---|---|---|
| NM_008084 | GAPDH | AACTTTGGCATTGTGGAAGG | GGATGCAGGGATGATGTTCT |
| NM_013693 | TNF-α | CCCCAAAGGGATGAGAAGTT | CACTTGGTGGTTTGCTACGA |
| NM_008361 | IL-1β | GGATGAGGACATGAGCACCT | AGCTCATATGGGTCCGACAG |
| NM_031168 | IL-6 | CCGGAGAGGAGACTTCACAG | CAGAATTGCCATTGCACAAC |
| NM_010548 | IL-10 | GCCTTATCGGAAATGATCCA | AGGGGAGAAATCGATGACAG |
| NM_010552 | IL-17A | CCAGGGAGAGCTTCATCTGT | AGGAAGTCCTTGGCCTCAGT |
| NM_008337 | IFN-γ | GCTCTTCCTCATGGCTGTTT | GTCACCATCCTTTTGCCAGT |
| NM_007527 | Bax | GTGAGCGGCTGCTTGTCT | GGTCCCGAAGTAGGAGAGGA |
| NM_009741 | Bcl-2 | TGGATCCAGGATAACGGAAG | CAAACAGAGGTCGCATGCTG |
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhu, H.; Li, Y.R. Oxidative stress and redox signaling mechanisms of inflammatory bowel disease: Updated experimental and clinical evidence. Exp. Biol. Med. 2012, 237, 474–480. [Google Scholar] [CrossRef]
- Fiocchi, C. Inflammatory bowel disease: Etiology and pathogenesis. Gastroenterology 1998, 115, 182–205. [Google Scholar] [CrossRef] [PubMed]
- Piechota-Polanczyk, A.; Fichna, J. Review article: The role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Mayer, L. The immune response in inflammatory bowel disease. Am. J. Gastroenterol. 2007, 102, 2058–2069. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Muñoz, F.; Dominguez-Lopez, A.; Yamamoto-Furusho, J.K. Role of cytokines in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 4280–4288. [Google Scholar] [CrossRef] [PubMed]
- Mudter, J.; Neurath, M.F. IL-6 signaling in inflammatory bowel disease: Pathophysiological role and clinical relevance. Inflamm. Bowel Dis. 2007, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Rios, A. Current view of the immunopathogenesis in inflammatory bowel disease and its implications for therapy. World J. Gastroenterol. 2008, 14, 1972–1980. [Google Scholar] [CrossRef] [PubMed]
- Guidi, L.; Pugliese, D.; Armuzzi, A. Update on the management of inflammatory bowel disease: Specific role of adalimumab. Clin. Exp. Gastroenterol. 2011, 4, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, T.T.; Cominelli, F. Cytokine therapy for crohn’s disease: Advances in translational research. Ann. Rev. Med. 2007, 58, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Tillack, C.; Ehmann, L.M.; Friedrich, M.; Laubender, R.P.; Papay, P.; Vogelsang, H.; Stallhofer, J.; Beigel, F.; Bedynek, A.; Wetzke, M.; et al. Anti-TNF antibody-induced psoriasiform skin lesions in patients with inflammatory bowel disease are characterised by interferon-γ-expressing Th1 cells and IL-17a/IL-22-expressing Th17 cells and respond to anti-IL-12/IL-23 antibody treatment. Gut 2014, 63, 567–577. [Google Scholar]
- Wlodarczyk, M.; Sobolewska, A.; Wojcik, B.; Loga, K.; Fichna, J.; Wisniewska-Jarosinska, M. Correlations between skin lesions induced by anti-tumor necrosis factor-α and selected cytokines in crohn’s disease patients. World J. Gastroenterol. 2014, 20, 7019–7026. [Google Scholar] [CrossRef] [PubMed]
- Maebuchi, M.; Samoto, M.; Kohno, M.; Ito, R.; Koikeda, T.; Hirotsuka, M.; Nakabou, Y. Improvement in the intestinal absorption of soy protein by enzymatic digestion to oligopeptide in healthy adult men. Food Sci. Technol. Res. 2007, 13, 45–53. [Google Scholar] [CrossRef]
- Young, D.; Ibuki, M.; Nakamori, T.; Fan, M.; Mine, Y. Soy-derived di-and tripeptides alleviate colon and ileum inflammation in pigs with dextran sodium sulfate-induced colitis. J. Nutr. 2012, 142, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kovacs-Nolan, J.; Archbold, T.; Fan, M.Z.; Juneja, L.R.; Okubo, T.; Mine, Y. Therapeutic potential of hen egg white peptides for the treatment of intestinal inflammation. J. Funct. Foods 2009, 1, 161–169. [Google Scholar] [CrossRef]
- Mochizuki, M.; Shigemura, H.; Hasegawa, N. Anti-inflammatory effect of enzymatic hydrolysate of corn gluten in an experimental model of colitis. J. Pharm. Pharmacol. 2010, 62, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Håversen, L.; Baltzer, L.; Dolphin, G.; Hanson, L.; Mattsby-Baltzer, I. Anti-inflammatory activities of human lactoferrin in acute dextran sulphate-induced colitis in mice. Scand. J. Immunol. 2003, 57, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Yi, F.; Guo, Z.-X.; Zhang, L.-X.; Yu, J.; Li, Q. Soluble eggshell membrane protein: Preparation, characterization and biocompatibility. Biomaterials 2004, 25, 4591–4599. [Google Scholar] [CrossRef] [PubMed]
- Hincke, M.; Gautron, J.; Panheleux, M.; Garcia-Ruiz, J.; McKee, M.; Nys, Y. Identification and localization of lysozyme as a component of eggshell membranes and eggshell matrix. Matrix Biol. 2000, 19, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Ruff, K.J.; DeVore, D.P.; Leu, M.D.; Robinson, M.A. Eggshell membrane: A possible new natural therapeutic for joint and connective tissue disorders. Results from two open-label human clinical studies. Clin. Interv. Aging 2009, 4, 235–240. [Google Scholar]
- Ruff, K.J.; Winkler, A.; Jackson, R.W.; DeVore, D.P.; Ritz, B.W. Eggshell membrane in the treatment of pain and stiffness from osteoarthritis of the knee: A randomized, multicenter, double-blind, placebo-controlled clinical study. Clin. Rheumatol. 2009, 28, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Benson, K.F.; Ruff, K.J.; Jensen, G.S. Effects of natural eggshell membrane (NEM) on cytokine production in cultures of peripheral blood mononuclear cells: Increased suppression of tumor necrosis factor-α levels after in vitro digestion. J. Med. Food 2012, 15, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Ruff, K.J.; DeVore, D.P. Reduction of pro-inflammatory cytokines in rats following 7-day oral supplementation with a proprietary eggshell membrane-derived product. Mod. Res. Inflamm. 2014, 3, 19–25. [Google Scholar] [CrossRef]
- Shi, Y.; Kovacs-Nolan, J.; Jiang, B.; Tsao, R.; Mine, Y. Antioxidant activity of enzymatic hydrolysates from eggshell membrane proteins and its protective capacity in human intestinal epithelial Caco-2 cells. J. Funct. Foods 2014, 10, 35–45. [Google Scholar] [CrossRef]
- Kim, J.-A.; Kim, D.-K.; Kang, O.-H.; Choi, Y.-A.; Park, H.-J.; Choi, S.-C.; Kim, T.-H.; Yun, K.-J.; Nah, Y.-H.; Lee, Y.-M. Inhibitory effect of luteolin on TNF-α-induced IL-8 production in human colon epithelial cells. Int. Immunopharmacol. 2005, 5, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Shah, Y.M.; Ma, X.; Morimura, K.; Kim, I.; Gonzalez, F.J. Pregnane X receptor activation ameliorates DSS-induced inflammatory bowel disease via inhibition of NF-κB target gene expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1114–G1122. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Fuss, I.J.; Blumberg, R.S. The immunology of mucosal models of inflammation 1. Ann. Rev. Immunol. 2002, 20, 495–549. [Google Scholar] [CrossRef]
- Kovacs-Nolan, J.; Zhang, H.; Ibuki, M.; Nakamori, T.; Yoshiura, K.; Turner, P.V.; Matsui, T.; Mine, Y. The pept1-transportable soy tripeptide vpy reduces intestinal inflammation. Biochim. Biophys. Acta 2012, 1820, 1753–1763. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F.; Finotto, S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine Growth Factor Rev. 2011, 22, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Playford, R.; Ghosh, S. Cytokines and growth factor modulators in intestinal inflammation and repair. J. Pathol. 2005, 205, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 cells. Ann. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Miossec, P. IL-17 and Th17 cells in human inflammatory diseases. Microbes Infect. 2009, 11, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Herfarth, H.; Brand, K.; Rath, H.; Rogler, G.; Schölmerich, J.; Falk, W. Nuclear factor-κB activity and intestinal inflammation in dextran sulphate sodium (DSS)-induced colitis in mice is suppressed by gliotoxin. Clin. Exp. Immunol. 2000, 120, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Bruewer, M.; Luegering, A.; Kucharzik, T.; Parkos, C.A.; Madara, J.L.; Hopkins, A.M.; Nusrat, A. Proinflammatory cytokines disrupt epithelial barrier function by apoptosis-independent mechanisms. J. Immunol. 2003, 171, 6164–6172. [Google Scholar] [CrossRef] [PubMed]
- Obermeier, F.; Kojouharoff, G.; Hans, W.; Scholmerich, J.; Gross, V.; Falk, W. Interferon-gamma (IFN-γ)- and tumour necrosis factor (TNF)-induced nitric oxide as toxic effector molecule in chronic dextran sulphate sodium (DSS)-induced colitis in mice. Clin. Exp. Immunol. 1999, 116, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Kovacs-Nolan, J.; Yang, C.; Archbold, T.; Fan, M.; Mine, Y. L-cysteine supplementation attenuates local inflammation and restores gut homeostasis in a porcine model of colitis. Biochim. Biophys. Acta 2009, 1790, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Kucharzik, T.; Stoll, R.; Lügering, N.; Domschke, W. Circulating antiinflammatory cytokine IL-10 in patients with inflammatory bowel disease (IBD). Clin. Exp. Immunol. 1995, 100, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Tomoyose, M.; Mitsuyama, K.; Ishida, H.; Toyonaga, A.; Tanikawa, K. Role of interleukin-10 in a murine model of dextran sulfate sodium-induced colitis. Scand. J. Gastroenterol. 1998, 33, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Ardizzone, S.; Porro, G.B. Biologic therapy for inflammatory bowel disease. Drugs 2005, 65, 2253–2286. [Google Scholar] [CrossRef] [PubMed]
- Ina, K.; Itoh, J.; Fukushima, K.; Kusugami, K.; Yamaguchi, T.; Kyokane, K.; Imada, A.; Binion, D.G.; Musso, A.; West, G.A. Resistance of crohn’s disease T cells to multiple apoptotic signals is associated with a Bcl-2/Bax mucosal imbalance. J. Immunol. 1999, 163, 1081–1090. [Google Scholar] [PubMed]
- Maxwell, J.R.; Brown, W.A.; Smith, C.L.; Byrne, F.R.; Viney, J.L. Methods of inducing inflammatory bowel disease in mice. Curr. Protoc. Pharmacol. 2009. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics Methods and Protocols; Springer: Cambridge, MA, USA, 1999; pp. 365–386. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Rupa, P.; Jiang, B.; Mine, Y. Hydrolysate from Eggshell Membrane Ameliorates Intestinal Inflammation in Mice. Int. J. Mol. Sci. 2014, 15, 22728-22742. https://doi.org/10.3390/ijms151222728
Shi Y, Rupa P, Jiang B, Mine Y. Hydrolysate from Eggshell Membrane Ameliorates Intestinal Inflammation in Mice. International Journal of Molecular Sciences. 2014; 15(12):22728-22742. https://doi.org/10.3390/ijms151222728
Chicago/Turabian StyleShi, Yaning, Prithy Rupa, Bo Jiang, and Yoshinori Mine. 2014. "Hydrolysate from Eggshell Membrane Ameliorates Intestinal Inflammation in Mice" International Journal of Molecular Sciences 15, no. 12: 22728-22742. https://doi.org/10.3390/ijms151222728
APA StyleShi, Y., Rupa, P., Jiang, B., & Mine, Y. (2014). Hydrolysate from Eggshell Membrane Ameliorates Intestinal Inflammation in Mice. International Journal of Molecular Sciences, 15(12), 22728-22742. https://doi.org/10.3390/ijms151222728