Choosing an Appropriate Infection Model to Study Quorum Sensing Inhibition in Pseudomonas Infections


Abstract
:
1. Introduction
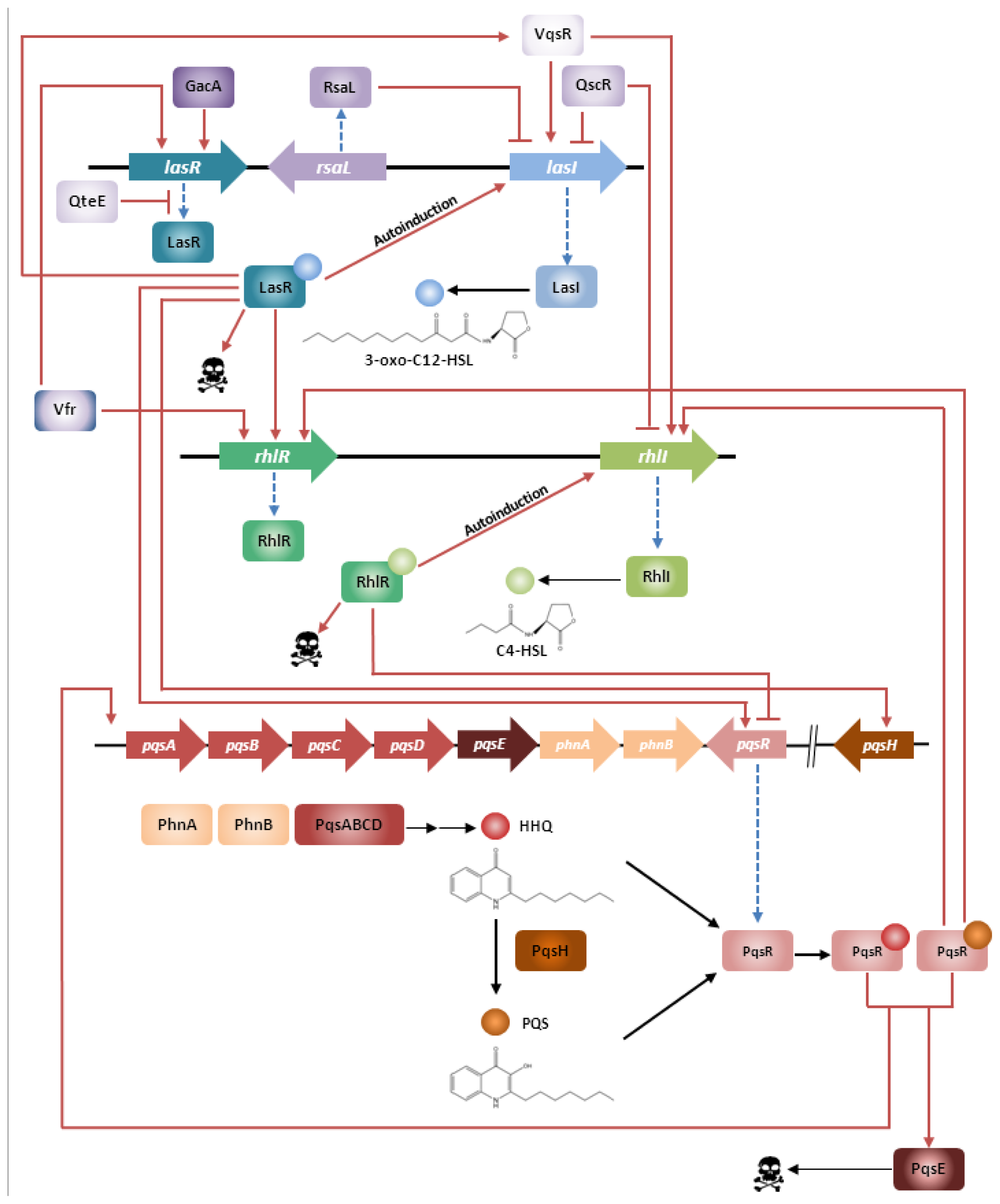
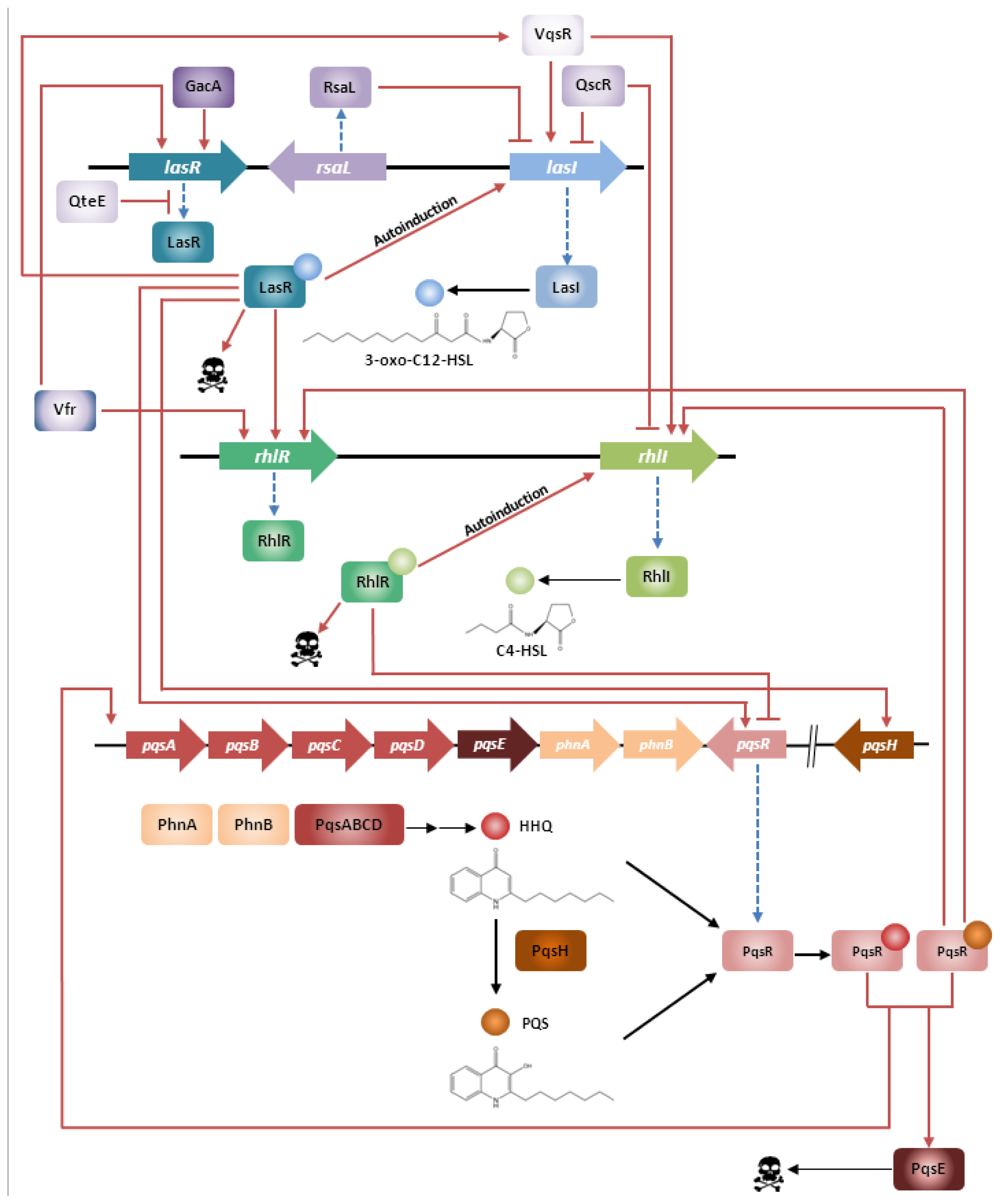
2. Quorum Sensing in Pseudomonas aeruginosa
3. P. aeruginosa-Related Human Infections
4. Models Available for Studying the Pathogenicity of P. aeruginosa
4.1. Plants
4.2. Cell Lines
4.3. Dictyostelium discoideum
4.4. Caenorhabditis elegans
4.5. Drosophila melanogaster (Fruit Flies)
4.6. Galleria mellonella (Wax Moth)
4.7. Bombyx mori (Silkworm)
4.8. Danio rerio (Zebrafish)
4.9. Rodents
4.9.1. Cystic Fibrosis Model
4.9.2. Burn Wound Model
4.9.3. Foreign Body Implants Model
4.9.4. Urinary Tract Infections (UTIs) Model
5. Quorum Quenching
5.1. Natural QSIs
5.2. Synthetic QSIs
6. Discussion
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Atkinson, S.; Williams, P. Quorum sensing and social networking in the microbial world. J. R. Soc. Interface 2009, 6, 959–978. [Google Scholar]
- Bassler, B.L. How bacteria talk to each other: Regulation of gene expression by quorum sensing. Curr. Opin. Microbiol 1999, 2, 582–587. [Google Scholar]
- Cámara, M.; Williams, P.; Hardman, A. Controlling infection by tuning in and turning down the volume of bacterial small-talk. Lancet Infect. Dis 2002, 2, 667–676. [Google Scholar]
- Winzer, K.; Hardie, K.R.; Williams, P. Bacterial cell-to-cell communication: Sorry, can’t talk now—Gone to lunch! Curr. Opin. Microbiol 2002, 5, 216–222. [Google Scholar]
- Williams, P.; Cámara, M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol 2009, 12, 182–191. [Google Scholar]
- Bielecki, P.; Glik, J.; Kawecki, M.; dos Santos, V.A.M. Towards understanding Pseudomonas aeruginosa burn wound infections by profiling gene expression. Biotechnol. Lett 2008, 30, 777–790. [Google Scholar]
- De Kievit, T.R.; Iglewski, B.H. Bacterial quorum sensing in pathogenic relationships. Infect. Immun 2000, 68, 4839–4849. [Google Scholar]
- Gambello, M.J.; Iglewski, B.H. Cloning and characterization of the Pseudomonas aeruginosa lasR gene, a transcriptional activator of elastase expression. J. Bacteriol 1991, 173, 3000–3009. [Google Scholar]
- Smith, R.; Iglewski, B.H. P. aeruginosa quorum-sensing systems and virulence. Curr. Opin. Microbiol 2003, 6, 56–60. [Google Scholar]
- Rampioni, G.; Schuster, M.; Greenberg, E.P.; Bertani, I.; Grasso, M.; Venturi, V.; Zennaro, E.; Leoni, L. RsaL provides quorum sensing homeostasis and functions as a global regulator of gene expression in Pseudomonas aeruginosa. Mol. Microbiol 2007, 66, 1557–1565. [Google Scholar]
- Albus, A.M.; Pesci, E.C.; Runyen-janecky, L.J.; West, S.E.H.; Iglewski, B.H. Vfr controls quorum sensing in Pseudomonas. J. Bacteriol 1997, 179, 3928–3935. [Google Scholar]
- Reimmann, C.; Beyeler, M.; Latifi, A.; Winteler, H.; Foglino, M.; Lazdunski, A.; Haas, D. The global activator GacA of Pseudomonas aeruginosa PAO positively controls the production of the autoinducer N-butyryl-homoserine lactone and the formation of the virulence factors pyocyanin, cyanide, and lipase. Mol. Microbiol 1997, 24, 309–319. [Google Scholar]
- Siehnel, R.; Traxler, B.; An, D.D.; Parsek, M.R.; Schaefer, A.L.; Singh, P.K. A unique regulator controls the activation threshold of quorum-regulated genes in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2010, 107, 7916–7921. [Google Scholar]
- Boontham, P.; Robins, A.; Chandran, P.; Pritchard, D.; Cámara, M.; Williams, P.; Chuthapisith, S.; McKechnie, A.; Rowlands, B.J.; Eremin, O. Significant immunomodulatory effects of Pseudomonas aeruginosa quorum-sensing signal molecules: Possible link in human sepsis. Clin. Sci. (Lond. ) 2008, 115, 343–351. [Google Scholar]
- Tateda, K.; Ishii, Y.; Horikawa, M.; Matsumoto, T.; Miyairi, S.; Pechere, J.C.; Standiford, T.J.; Ishiguro, M.; Yamaguchi, K. The Pseudomonas aeruginosa autoinducer N-3-oxododecanoyl homoserine lactone accelerates apoptosis in macrophages and neutrophils. Infect. Immun 2003, 71, 5785–5793. [Google Scholar]
- Shiner, E.K.; Terentyev, D.; Bryan, A.; Sennoune, S.; Martinez-Zaguilan, R.; Li, G.; Gyorke, S.; Williams, S.C.; Rumbaugh, K.P. Pseudomonas aeruginosa autoinducer modulates host cell responses through calcium signalling. Cell. Microbiol 2006, 8, 1601–1610. [Google Scholar]
- Mayer, M.L.; Sheridan, J.A.; Blohmke, C.J.; Turvey, S.E.; Hancock, R.E.W. The Pseudomonas aeruginosa autoinducer 3O-C12 homoserine lactone provokes hyperinflammatory responses from cystic fibrosis airway epithelial cells. PLoS One 2011, 6, e16246. [Google Scholar]
- Latifi, A.; Foglino, M.; Tanaka, K.; Williams, P.; Lazdunski, A. A hierarchical quorum-sensing cascade in Pseudomonas aeruginosa links the transcriptional activators LasR and RhIR (VsmR) to expression of the stationary-phase sigma factor RpoS. Mol. Microbiol 1996, 21, 1137–1146. [Google Scholar]
- Venturi, V.; Friscina, A.; Bertani, I.; Devescovi, G.; Aguilar, C. Quorum sensing in the Burkholderia cepacia complex. Res. Microbiol 2004, 155, 238–244. [Google Scholar]
- Winzer, K.; Falconer, C.; Garber, N.C.; Diggle, P.; Camara, M.; Williams, P.; Diggle, S.P. The Pseudomonas aeruginosa lectins PA-IL and PA-IIL are controlled by quorum sensing and by RpoS. J. Bacteriol 2000, 182, 478–483. [Google Scholar]
- Dekimpe, V.; Déziel, E. Revisiting the quorum-sensing hierarchy in Pseudomonas aeruginosa: The transcriptional regulator RhlR regulates LasR-specific factors. Microbiology 2009, 155, 712–723. [Google Scholar]
- Schuster, M.; Lostroh, C.P.; Ogi, T.; Greenberg, E.P. Identification, timing, and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: A transcriptome analysis. J. Bacteriol 2003, 185, 2066–2079. [Google Scholar]
- Wagner, V.E.; Bushnell, D.; Passador, L.; Brooks, A.I.; Iglewski, B.H. Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: Effects of growth phase and environment. J. Bacteriol 2003, 185, 2080–2095. [Google Scholar]
- Diggle, S.P.; Winzer, K.; Chhabra, S.R.; Worrall, K.E.; Cámara, M.; Williams, P. The Pseudomonas aeruginosa quinolone signal molecule overcomes the cell density-dependency of the quorum sensing hierarchy, regulates rhl-dependent genes at the onset of stationary phase and can be produced in the absence of LasR. Mol. Microbiol 2003, 50, 29–43. [Google Scholar]
- Wade, D.S.; Calfee, M.W.; Rocha, E.R.; Ling, A.; Engstrom, E.; Coleman, J.P.; Everett, C.; Ling, E.A.; Pesci, E.C. Regulation of Pseudomonas quinolone signal synthesis in Pseudomonas aeruginosa. J. Bacteriol 2005, 187, 4372–4380. [Google Scholar]
- Aendekerk, S.; Diggle, S.P.; Song, Z.; Høiby, N.; Cornelis, P.; Williams, P.; Cámara, M. The MexGHI-OpmD multidrug efflux pump controls growth, antibiotic susceptibility and virulence in Pseudomonas aeruginosa via 4-quinolone-dependent cell-to-cell communication. Microbiology 2005, 151, 1113–1125. [Google Scholar]
- Pesci, E.C.; Milbank, J.B.; Pearson, J.P.; McKnight, S.; Kende, A.S.; Greenberg, E.P.; Iglewski, B.H. Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 11229–11234. [Google Scholar]
- Lee, J.; Wu, J.; Deng, Y.; Wang, J.; Wang, C.; Wang, J.; Chang, C.; Dong, Y.; Williams, P.; Zhang, L.-H. A cell-cell communication signal integrates quorum sensing and stress response. Nat. Chem. Biol 2013, 9, 339–343. [Google Scholar]
- Heurlier, K.; Dénervaud, V.; Haas, D. Impact of quorum sensing on fitness of Pseudomonas aeruginosa. Int. J. Med. Microbiol 2006, 296, 93–102. [Google Scholar]
- Bjarnsholt, T.; Jensen, P.Ø.; Jakobsen, T.H.; Phipps, R.; Nielsen, A.K.; Rybtke, M.T.; Tolker-Nielsen, T.; Givskov, M.; Høiby, N.; Ciofu, O. Quorum sensing and virulence of Pseudomonas aeruginosa during lung infection of cystic fibrosis patients. PLoS One 2010, 5, e10115. [Google Scholar]
- Govan, J.R.; Deretic, V. Microbial pathogenesis in cystic fibrosis: Mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev 1996, 60, 539–574. [Google Scholar]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of Pseudomonas aeruginosa infection: Lessons from a versatile opportunist. Microbes Infect 2000, 2, 1051–1060. [Google Scholar]
- Willcox, M.D.P.; Zhu, H.; Conibear, T.C.R.; Hume, E.B.H.; Givskov, M.; Kjelleberg, S.; Rice, S.A. Role of quorum sensing by Pseudomonas aeruginosa in microbial keratitis and cystic fibrosis. Microbiology 2008, 154, 2184–2194. [Google Scholar]
- Bjarnsholt, T.; Givskov, M. Quorum-sensing blockade as a strategy for enhancing host defences against bacterial pathogens. Philos. Trans. R. Soc. Lond. B Biol. Sci 2007, 362, 1213–1222. [Google Scholar]
- Ichikawa, J.K.; Norris, A.; Bangera, M.G.; Geiss, G.K.; van’t Wout, A.B.; Bumgarner, R.E.; Lory, S. Interaction of Pseudomonas aeruginosa with epithelial cells: Identification of differentially regulated genes by expression microarray analysis of human cDNAs. Proc. Natl. Acad. Sci. USA 2000, 97, 9659–9664. [Google Scholar]
- Cosson, P.; Zulianello, L.; Join-lambert, O.; Faurisson, F.; Gebbie, L.; Benghezal, M.; van Delden, C.; Curty, L.K.; Köhler, T. Pseudomonas aeruginosa virulence analyzed in a Dictyostelium discoideum host system. J. Bacteriol 2002, 184, 3027–3033. [Google Scholar]
- Mahajan-Miklos, S.; Tan, M.W.; Rahme, L.G.; Ausubel, F.M. Molecular mechanisms of bacterial virulence elucidated using a Pseudomonas aeruginosa-Caenorhabditis elegans pathogenesis model. Cell 1999, 96, 47–56. [Google Scholar]
- Apidianakis, Y.; Rahme, L.G. Drosophila melanogaster as a model host for studying Pseudomonas aeruginosa infection. Nat. Protoc 2009, 4, 1285–1294. [Google Scholar]
- Zaidi, T.; Bajmoczi, M.; Zaidi, T.; Golan, D.E.; Pier, G.B. Disruption of CFTR-dependent lipid rafts reduces bacterial levels and corneal disease in a murine model of Pseudomonas aeruginosa keratitis. Investig. Ophthalmol. Vis. Sci 2008, 49, 1000–1009. [Google Scholar]
- Prithiviraj, B.; Bais, H.P.; Weir, T.; Suresh, B.; Najarro, E.H.; Dayakar, V.; Schweizer, H.P.; Vivanco, J.M.; Dayakar, B. Down regulation of virulence factors of Pseudomonas aeruginosa by salicylic acid attenuates its virulence on Arabidopsis thaliana and Caenorhabditis elegans. Infect. Immun 2005, 73, 5319–5328. [Google Scholar]
- Zhang, Y.; Hu, Y.; Yang, B.; Ma, F.; Lu, P.; Li, L.; Wan, C.; Rayner, S.; Chen, S. Duckweed (Lemna minor) as a model plant system for the study of human microbial pathogenesis. PLoS One 2010, 5, e13527. [Google Scholar]
- Wieland, C.W.; Siegmund, B.; Senaldi, G.; Vasil, M.L.; Dinarello, C.A.; Fantuzzi, G. Pulmonary inflammation induced by Pseudomonas aeruginosa lipopolysaccharide, phospholipase C, and exotoxin A: Role of interferon regulatory factor 1. Infect. Immun 2002, 70, 1352–1358. [Google Scholar]
- Ye, L.; Li, G.; Li, H.; Wang, L. Pseudomonas aeruginosa pvdQ gene prevents Caco-2 cells from obstruction of quorum-sensing signal. Curr. Microbiol 2011, 62, 32–37. [Google Scholar]
- Kukavica-Ibrulj, I.; Levesque, R.C. Animal models of chronic lung infection with Pseudomonas aeruginosa: Useful tools for cystic fibrosis studies. Lab. Anim 2008, 42, 389–412. [Google Scholar]
- Bals, R.; Hiemstra, P.S. Innate immunity in the lung: How epithelial cells fight against respiratory pathogens. Eur. Respir. J 2004, 23, 327–333. [Google Scholar]
- Davidson, D.J.; Gray, M.A.; Kilanowski, F.M.; Tarran, R.; Randell, S.H.; Sheppard, D.N.; Argent, B.E.; Dorin, J.R. Murine epithelial cells: Isolation and culture. J. Cyst. Fibros 2004, 3, 59–62. [Google Scholar]
- Ulrich, M.; Döring, G. Three-dimensional human airway epithelial cell cultures. J. Cyst. Fibros 2004, 3, 55–57. [Google Scholar]
- Willems, T.; Jorissen, M. Sequential monolayer-suspension culture of human airway epithelial cells. J. Cyst. Fibros 2004, 3, 53–54. [Google Scholar]
- Cosson, P. A non-mammalian system to study bacterial infections. Altex 2007, 24, 78–79. [Google Scholar]
- Lima, W.C.; Lelong, E.; Cosson, P. What can Dictyostelium bring to the study of Pseudomonas infections? Semin. Cell Dev. Biol 2011, 22, 77–81. [Google Scholar]
- Kurz, C.L.; Ewbank, J.J. Infection in a dish: High-throughput analyses of bacterial pathogenesis. Curr. Opin. Microbiol 2007, 10, 10–16. [Google Scholar]
- Pukatzki, S.; Kessin, R.H.; Mekalanos, J.J. The human pathogen Pseudomonas aeruginosa utilizes conserved virulence pathways to infect the social amoeba Dictyostelium discoideum. Proc. Natl. Acad. Sci. USA 2002, 99, 3159–3164. [Google Scholar]
- Steinert, M.; Heuner, K. Dictyostelium as host model for pathogenesis. Cell. Microbiol 2005, 7, 307–314. [Google Scholar]
- Irazoqui, J.E.; Urbach, J.M.; Ausubel, F.M. Evolution of host innate defence: Insights from Caenorhabditis elegans and primitive invertebrates. Nat. Rev. Immunol 2010, 10, 47–58. [Google Scholar]
- Aballay, A.; Ausubel, F.M. Caenorhabditis elegans as a host for the study of host-pathogen interactions. Curr. Opin. Microbiol 2002, 5, 97–101. [Google Scholar]
- Kurz, C.L.; Ewbank, J.J. Caenorhabditis elegans for the study of host-pathogen interactions. Trends Microbiol 2000, 8, 142–144. [Google Scholar]
- Mylonakis, E.; Ausubel, F.M.; Tang, R.J.; Calderwood, S.B. The art of serendipity: Killing of Caenorhabditis elegans by human pathogens as a model of bacterial and fungal pathogenesis. Expert Rev. Anti Infect. Ther 2003, 1, 167–173. [Google Scholar]
- Cezairliyan, B.; Vinayavekhin, N.; Grenfell-Lee, D.; Yuen, G.J.; Saghatelian, A.; Ausubel, F.M. Identification of Pseudomonas aeruginosa phenazines that kill Caenorhabditis elegans. PLoS Pathog 2013, 9, e1003101. [Google Scholar]
- Tan, M.W.; Mahajan-Miklos, S.; Ausubel, F.M. Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to model mammalian bacterial pathogenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 715–720. [Google Scholar]
- Tan, M.W.; Rahme, L.G.; Sternberg, J.A.; Tompkins, R.G.; Ausubel, F.M. Pseudomonas aeruginosa killing of Caenorhabditis elegans used to identify P. aeruginosa virulence factors. Proc. Natl. Acad. Sci. USA 1999, 96, 2408–2413. [Google Scholar]
- Darby, C.; Cosma, C.L.; Thomas, J.H.; Manoil, C. Lethal paralysis of Caenorhabditis elegans by Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 15202–15207. [Google Scholar]
- Zaborin, A.; Romanowski, K.; Gerdes, S.; Holbrook, C.; Lepine, F.; Long, J.; Poroyko, V.; Diggle, S.P.; Wilke, A.; Righetti, K.; et al. Red death in Caenorhabditis elegans caused by Pseudomonas aeruginosa PAO1. Proc. Natl. Acad. Sci. USA 2009, 106, 6327–6332. [Google Scholar]
- Kirienko, N.V.; Kirienko, D.R.; Larkins-Ford, J.; Wahlby, C.; Ruvkun, G.; Ausubel, F.M. Pseudomonas aeruginosa disrupts Caenorhabditis elegans iron homeostasis, causing a hypoxic response and death. Cell Host Microbe 2013, 13, 406–416. [Google Scholar]
- Rubin, G.M.; Lewis, E.B. A brief history of Drosophila’s contributions to genome research. Science 2000, 287, 2216–2218. [Google Scholar]
- Adams, M.D.; Ceniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar]
- Tzou, P.; de Gregorio, E.; Lemaitre, B. How Drosophila combats microbial infection: A model to study innate immunity and host-pathogen interactions. Curr. Opin. Microbiol 2002, 5, 102–110. [Google Scholar]
- Apidianakis, Y.; Rahme, L.G.; Heitman, J.; Ausubel, F.M.; Calderwood, S.B.; Mylonakis, E. Challenge of Drosophila melanogaster with Cryptococcus neoformans and role of the innate immune response. Eukaryot. Cell 2004, 3, 413–419. [Google Scholar]
- Lutter, E.I.; Purighalla, S.; Duong, J.; Storey, D.G. Lethality and cooperation of Pseudomonas aeruginosa quorum-sensing mutants in Drosophila melanogaster infection models. Microbiology 2012, 158, 2125–2132. [Google Scholar]
- Limmer, S.; Haller, S.; Drenkard, E.; Lee, J.; Yu, S.; Kocks, C.; Ausubel, F.M.; Ferrandon, D. Pseudomonas aeruginosa RhlR is required to neutralize the cellular immune response in a Drosophila melanogaster oral infection model. Proc. Natl. Acad. Sci. USA 2011, 108, 17378–17383. [Google Scholar]
- D’Argenio, D.A.; Gallagher, L.A.; Berg, C.A.; Manoil, C. Drosophila as a model host for Pseudomonas aeruginosa infection. J. Bacteriol 2001, 183, 1466–1471. [Google Scholar]
- Chugani, S.A.; Whiteley, M.; Lee, K.M.; D’Argenio, D.; Manoil, C.; Greenberg, E.P. QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2001, 98, 2752–2757. [Google Scholar]
- Peleg, A.Y.; Jara, S.; Monga, D.; Eliopoulos, G.M.; Moellering, R.C.; Mylonakis, E. Galleria mellonella as a model system to study Acinetobacter baumannii pathogenesis and therapeutics. Antimicrob. Agents Chemother 2009, 53, 2605–2609. [Google Scholar]
- Seed, K.D.; Dennis, J.J. Development of Galleria mellonella as an alternative infection model for the Burkholderia cepacia complex. Infect. Immun 2008, 76, 1267–1275. [Google Scholar]
- Lebreton, F.; Le Bras, F.; Reffuveille, F.; Ladjouzi, R.; Giard, J.-C.; Leclercq, R.; Cattoir, V. Galleria mellonella as a model for studying Enterococcus faecium host persistence. J. Mol. Microbiol. Biotechnol 2011, 21, 191–196. [Google Scholar]
- Leuko, S.; Raivio, T.L. Mutations that impact the enteropathogenic Escherichia coli Cpx envelope stress response attenuate virulence in Galleria mellonella. Infect. Immun 2012, 80, 3077–3085. [Google Scholar]
- Harding, C.R.; Schroeder, G.N.; Reynolds, S.; Kosta, A.; Collins, J.W.; Mousnier, A.; Frankel, G. Legionella pneumophila pathogenesis in the Galleria mellonella infection model. Infect. Immun 2012, 80, 2780–2790. [Google Scholar]
- Joyce, S.A.; Gahan, C.G.M. Molecular pathogenesis of Listeria monocytogenes in the alternative model host Galleria mellonella. Microbiology 2010, 156, 3456–3468. [Google Scholar]
- Mukherjee, K.; Altincicek, B.; Hain, T.; Domann, E.; Vilcinskas, A.; Chakraborty, T. Galleria mellonella as a model system for studying Listeria pathogenesis. Appl. Environ. Microbiol 2010, 76, 310–317. [Google Scholar]
- Ziprin, R.; Hartman, P.A. Toxicity of Pseudomonas aeruginosa bacterins and cell walls to the greater wax moth, Galleria mellonella. J. Invertebr. Pathol 1971, 17, 265–269. [Google Scholar]
- Dunphy, G.B.; Morton, D.B.; Kropinski, A.; Chadwick, J.M. Pathogenicity of lipopolysaccharide mutants of Pseudomonas aeruginosa for larvae of Galleria mellonella: Bacterial properties associated with virulence. J. Invertebr. Pathol 1986, 55, 48–55. [Google Scholar]
- Miyata, S.; Casey, M.; Frank, D.W.; Ausubel, F.M.; Drenkard, E. Use of the Galleria mellonella caterpillar as a model host to study the role of the type III secretion system in Pseudomonas aeruginosa pathogenesis. Infect. Immun 2003, 71, 2404–2413. [Google Scholar]
- Harrison, F.; Browning, L.E.; Vos, M.; Buckling, A. Cooperation and virulence in acute Pseudomonas aeruginosa infections. BMC Biol. 2006, 4. [Google Scholar] [CrossRef]
- Andrejko, M.; Mizerska-Dudka, M.; Jakubowicz, T. Antibacterial activity in vivo and in vitro in the hemolymph of Galleria mellonella infected with Pseudomonas aeruginosa. Comp. Biochem. Physiol. B. Biochem. Mol. Biol 2009, 152, 118–123. [Google Scholar]
- Desbois, A.P.; Coote, P.J. Wax moth larva (Galleria mellonella): An in vivo model for assessing the efficacy of antistaphylococcal agents. J. Antimicrob. Chemother 2011, 66, 1785–1790. [Google Scholar]
- Evans, B.A.; Rozen, D.E. A Streptococcus pneumoniae infection model in larvae of the wax moth Galleria mellonella. Eur. J. Clin. Microbiol. Infect. Dis 2012, 31, 2653–2660. [Google Scholar]
- Scully, L.R.; Bidochka, M.J. An alternative insect pathogenic strategy in an Aspergillus flavus auxotroph. Mycol. Res 2009, 113, 230–239. [Google Scholar]
- Fallon, J.; Troy, N.; Kavanagh, K. Pre-exposure of Galleria mellonella larvae to different doses of Aspergillus fumigatus conidia causes differential activation of cellular and humoral immune responses. Virulence 2011, 2, 413–421. [Google Scholar]
- Mesa-Arango, A.C.; Forastiero, A.; Bernal-Martínez, L.; Cuenca-Estrella, M.; Mellado, E.; Zaragoza, O. The non-mammalian host Galleria mellonella can be used to study the virulence of the fungal pathogen Candida tropicalis and the efficacy of antifungal drugs during infection by this pathogenic yeast. Med. Mycol 2012, 51, 461–472. [Google Scholar]
- Brennan, M.; Thomas, D.Y.; Whiteway, M.; Kavanagh, K. Correlation between virulence of Candida albicans mutants in mice and Galleria mellonella larvae. FEMS. Immunol. Med. Microbiol 2002, 34, 153–157. [Google Scholar]
- Mylonakis, E.; Moreno, R.; El Khoury, J.B.; Idnurm, A.; Heitman, J.; Calderwood, S.B.; Ausubel, F.M.; Diener, A. Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect. Immun 2005, 73, 3842–2850. [Google Scholar]
- Coleman, J.J.; Muhammed, M.; Kasperkovitz, P.V.; Vyas, J.M.; Mylonakis, E. Fusarium pathogenesis investigated using Galleria mellonella as a heterologous host. Fungal Biol 2011, 115, 1279–1289. [Google Scholar]
- Koch, G.; Nadal Jimenez, P.; Cool, R.H.; Quax, W.J. Assessing Pseudomonas Virulence with Non-Mammalian Host: Galleria mellonella. In Pseudomonas aeruginosa: Methods and Protocols; Ramos, J.-L., Filloux, A., Eds.; Springer: New York, NY, USA, 2013; Volume 31. [Google Scholar]
- Jander, G.; Rahme, L.G.; Ausubel, F.M. Positive correlation between virulence of Pseudomonas aeruginosa mutants in mice and insects. J. Bacteriol 2000, 182, 3843–3845. [Google Scholar]
- Seitz, V.; Clermont, A.; Wedde, M.; Hummel, M.; Vilcinskas, A.; Schlatterer, K.; Podsiadlowski, L. Identification of immunorelevant genes from greater wax moth (Galleria mellonella) by a subtractive hybridization approach. Dev. Comp. Immunol 2003, 27, 207–215. [Google Scholar]
- Andrejko, M.; Mizerska-Dudka, M.; Jakubowicz, T. Changes in Galleria mellonella apolipophorin III level during Pseudomonas aeruginosa infection. J. Invertebr. Pathol 2008, 97, 14–19. [Google Scholar]
- Imperi, F.; Massai, F.; Ramachandran Pillai, C.; Longo, F.; Zennaro, E.; Rampioni, G.; Visca, P.; Leoni, L. New life for an old drug: The anthelmintic drug niclosamide inhibits Pseudomonas aeruginosa quorum sensing. Antimicrob. Agents Chemother 2013, 57, 996–1005. [Google Scholar]
- Kaito, C.; Sekimizu, K. A silkworm model of pathogenic bacterial infection. Drug Discov. Ther 2007, 1, 89–93. [Google Scholar]
- Chieda, Y.; Iiyama, K.; Yasunaga-Aoki, C.; Lee, J.M.; Kusakabe, T.; Shimizu, S. Pathogenicity of gacA mutant of Pseudomonas aeruginosa PA01 in the silkworm, Bombyx mori. FEMS Microbiol. Lett 2005, 244, 181–186. [Google Scholar]
- Davis, J.M.; Clay, H.; Lewis, J.L.; Ghori, N.; Herbomel, P.; Ramakrishnan, L. Real-time visualization of Mycobacterium-macrophage interactions leading to initiation of granuloma formation in zebrafish embryos. Immunity 2002, 17, 693–702. [Google Scholar]
- Van der Sar, A.M.; Musters, R.J.P.; van Eeden, F.J.M.; Appelmelk, B.J.; Vandenbroucke-Grauls, C.M.J.E.; Bitter, W. Zebrafish embryos as a model host for the real time analysis of Salmonella typhimurium infections. Cell. Microbiol 2003, 5, 601–611. [Google Scholar]
- Pressley, M.E.; Phelan, P.E.; Witten, P.E.; Mellon, M.T.; Kim, C.H. Pathogenesis and inflammatory response to Edwardsiella tarda infection in the zebrafish. Dev. Comp. Immunol 2005, 29, 501–513. [Google Scholar]
- Prajsnar, T.K.; Cunliffe, V.T.; Foster, S.J.; Renshaw, S.A. A novel vertebrate model of Staphylococcus aureus infection reveals phagocyte-dependent resistance of zebrafish to non-host specialized pathogens. Cell. Microbiol 2008, 10, 2312–2325. [Google Scholar]
- Neely, M.N.; Pfeifer, J.D.; Caparon, M. Streptococcus-zebrafish model of bacterial pathogenesis. Infect. Immun 2002, 70, 3904–3914. [Google Scholar]
- Clatworthy, A.E.; Lee, J.S.-W.; Leibman, M.; Kostun, Z.; Davidson, A.J.; Hung, D.T. Pseudomonas aeruginosa infection of zebrafish involves both host and pathogen determinants. Infect. Immun 2009, 77, 1293–1303. [Google Scholar]
- Snouwaert, J.N.; Brigman, K.K.; Latour, A.M.; Malouf, N.N.; Boucher, C.; Smithies, O.; Koller, B.H. An animal model for cystic fibrosis made by gene targeting. Science 1992, 257, 1083–1088. [Google Scholar]
- O’Neal, W.K.; Hasty, P.; McCray, P.B.; Casey, B.; Rivera-Pérez, J.; Welsh, M.J.; Beaudet, A.L.; Bradley, A. A severe phenotype in mice with a duplication of exon 3 in the cystic fibrosis locus. Hum. Mol. Genet 1993, 2, 1561–1569. [Google Scholar]
- Davidson, D.J.; Dorin, J.R. The CF mouse: An important tool for studying cystic fibrosis. Expert Rev. Mol. Med 2001, 3, 1–27. [Google Scholar]
- Guilbault, C.; Saeed, Z.; Downey, G.P.; Radzioch, D. Cystic fibrosis mouse models. Am. J. Respir. Cell Mol. Biol 2007, 36, 1–7. [Google Scholar]
- Keiser, N.W.; Engelhardt, J.F. New animal models of cystic fibrosis: What are they teaching us? Curr. Opin. Pulm. Med 2011, 17, 478–483. [Google Scholar]
- Cash, H.A.; Woods, D.E.; McCullough, B.; Johanson, W.G.; Bass, J.A. A rat model of chronic respiratory infection with Pseudomonas aeruginosa. Am. Rev. Respir. Dis 1979, 119, 453–459. [Google Scholar]
- Starke, J.R.; Edwards, M.S.; Langston, C.; Baker, C.J. A mouse model of chronic pulmonary infection with Pseudomonas aeruginosa and Pseudomonas cepacia. Pediatr. Res 1987, 22, 698–702. [Google Scholar]
- Pennington, J.E.; Hickey, W.F.; Blackwood, L.L.; Arnaut, M.A. Active immunization with lipopolysaccharide Pseudomonas antigen for chronic Pseudomonas bronchopneumonia in guinea pigs. J. Clin. Investig 1981, 68, 1140–1148. [Google Scholar]
- Pedersen, S.S.; Shand, G.H.; Hansen, B.L.; Hansen, G.N. Induction of experimental chronic Pseudomonas aeruginosa lung infection with P. aeruginosa entrapped in alginate microspheres. APMIS 1990, 98, 203–211. [Google Scholar]
- Hoffmann, N. Animal models of chronic Pseudomonas aeruginosa lung infection in cystic fibrosis. Drug Discov. Today Dis. Models 2007, 4, 99–104. [Google Scholar]
- Hoffmann, N.; Rasmussen, T.B.; Jensen, P.; Stub, C.; Hentzer, M.; Ciofu, O.; Givskov, M.; Johansen, H.K.; Høiby, N.; Jensen, P.Ø.; et al. Novel mouse model of chronic Pseudomonas aeruginosa lung infection mimicking cystic fibrosis. Infect. Immun. 2005, 73, 2504–2514. [Google Scholar]
- Hoffmann, N.; Lee, B.; Hentzer, M.; Rasmussen, T.B.; Song, Z.; Johansen, H.K.; Givskov, M.; Høiby, N. Azithromycin blocks quorum sensing and alginate polymer formation and increases the sensitivity to serum and stationary-growth-phase killing of Pseudomonas aeruginosa and attenuates chronic P. aeruginosa lung infection in Cftr(−/−) mi. Antimicrob. Agents Chemother 2007, 51, 3677–3687. [Google Scholar]
- Mizukane, R.; Hirakata, Y.; Kaku, M.; Ishii, Y.; Furuya, N.; Ishida, K.; Koga, H.; Kohno, S.; Yamaguchi, K. Comparative in vitro exoenzyme-suppressing activities of azithromycin and other macrolide antibiotics against Pseudomonas aeruginosa. Antimicrob. Agents Chemother 1994, 38, 528–533. [Google Scholar]
- Rumbaugh, K.P.; Griswold, J.A.; Iglewski, B.H.; Hamood, A.N.; Barbara, H. Contribution of quorum sensing to the virulence of Pseudomonas aeruginosa in burn wound infections. Infect. Immun 1999, 67, 5854–5862. [Google Scholar]
- Rumbaugh, K.P.; Colmer, J.A.; Griswold, J.A.; Hamood, A.N. The effects of infection of thermal injury by Pseudomonas aeruginosa PAO1 on the murine cytokine response. Cytokine 2001, 16, 160–168. [Google Scholar]
- Barnea, Y.; Carmeli, Y.; Kuzmenko, B.; Gur, E.; Hammer-Munz, O.; Navon-Venezia, S. The establishment of a Pseudomonas aeruginosa-infected burn-wound sepsis model and the effect of imipenem treatment. Ann. Plast. Surg 2006, 56, 674–679. [Google Scholar]
- Rahme, L.G.; Stevens, E.J.; Wolfort, S.F.; Shao, J.; Tompkins, R.G.; Ausubel, F.M. Common virulence factors for bacterial pathogenicity in plants and animals. Science 1995, 268, 1899–1902. [Google Scholar]
- Li, N.; Hu, X.; Liu, Y.; Wang, Y.; Wang, Y.; Liu, J.; Cai, W.; Bai, X.; Zhu, X.; Han, J.; et al. Systemic inflammatory responses and multiple organ dysfunction syndrome following skin burn wound and Pseudomonas aeruginosa infection in mice. Shock 2013, 40, 152–159. [Google Scholar]
- Nidadavolu, P.; Amor, W.; Tran, P.L.; Dertien, J.; Colmer-Hamood, J.A.; Hamood, A.N. Garlic ointment inhibits biofilm formation by bacterial pathogens from burn wounds. J. Med. Microbiol 2012, 61, 662–671. [Google Scholar]
- Dickinson, G.M.; Bisno, A.L. Infections associated with indwelling devices: Infections related to extravascular devices. Antimicrob. Agents Chemother 1989, 33, 602–607. [Google Scholar]
- Christensen, L.D.; Moser, C.; Jensen, P.Ø.; Rasmussen, T.B.; Christophersen, L.; Kjelleberg, S.; Kumar, N.; Høiby, N.; Givskov, M.; Bjarnsholt, T. Impact of Pseudomonas aeruginosa quorum sensing on biofilm persistence in an in vivo intraperitoneal foreign-body infection model. Microbiology 2007, 153, 2312–2320. [Google Scholar]
- Christensen, L.D.; van Gennip, M.; Jakobsen, T.H.; Alhede, M.; Hougen, H.P.; Høiby, N.; Bjarnsholt, T.; Givskov, M. Synergistic antibacterial efficacy of early combination treatment with tobramycin and quorum-sensing inhibitors against Pseudomonas aeruginosa in an intraperitoneal foreign-body infection mouse model. J. Antimicrob. Chemother 2012, 67, 1198–1206. [Google Scholar]
- Bjarnsholt, T.; van Gennip, M.; Jakobsen, T.H.; Christensen, L.D.; Jensen, P.Ø.; Givskov, M. In vitro screens for quorum sensing inhibitors and in vivo confirmation of their effect. Nat. Protoc. 2010, 5, 282–293. [Google Scholar]
- Bjarnsholt, T.; Jensen, P.Ø.; Burmølle, M.; Hentzer, M.; Haagensen, J.A.J.; Hougen, H.P.; Calum, H.; Madsen, K.G.; Moser, C.; Molin, S.; et al. Pseudomonas aeruginosa tolerance to tobramycin, hydrogen peroxide and polymorphonuclear leukocytes is quorum-sensing dependent. Microbiology 2005, 151, 373–383. [Google Scholar]
- Harjai, K.; Kumar, R.; Singh, S. Garlic blocks quorum sensing and attenuates the virulence of Pseudomonas aeruginosa. FEMS Immunol. Med. Microbiol 2010, 58, 161–168. [Google Scholar]
- Goldsworthy, M.J.H. Gene expression of Pseudomonas aeruginosa and MRSA within a catheter-associated urinary tract infection biofilm model. Biosci. Horiz 2008, 1, 28–37. [Google Scholar]
- Bala, A.; Kumar, R.; Harjai, K. Inhibition of quorum sensing in Pseudomonas aeruginosa by azithromycin and its effectiveness in urinary tract infections. J. Med. Microbiol 2011, 60, 300–306. [Google Scholar]
- LaSarre, B.; Federle, M.J. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev 2013, 77, 73–111. [Google Scholar]
- Galloway, W.R.J.D.; Hodgkinson, J.T.; Bowden, S.D.; Welch, M.; Spring, D.R. Quorum sensing in Gram-negative bacteria: Small-molecule modulation of AHL and AI-2 quorum sensing pathways. Chem. Rev 2011, 111, 28–67. [Google Scholar]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv 2013, 31, 224–245. [Google Scholar]
- Hoang, T.T.; Schweizer, H.P. Characterization of Pseudomonas aeruginosa enoyl-acyl carrier protein reductase (FabI): A target for the antimicrobial triclosan and its role in acylated homoserine lactone synthesis. J. Bacteriol 1999, 181, 5489–5497. [Google Scholar]
- Calfee, M.W.; Coleman, J.P.; Pesci, E.C. Interference with Pseudomonas quinolone signal synthesis inhibits virulence factor expression by Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2001, 98, 11633–11637. [Google Scholar]
- Lesic, B.; Lépine, F.; Déziel, E.; Zhang, J.; Zhang, Q.; Padfield, K.; Castonguay, M.-H.; Milot, S.; Stachel, S.; Tzika, A.A.; et al. Inhibitors of pathogen intercellular signals as selective anti-infective compounds. PLoS Pathog 2007, 3, 1229–1239. [Google Scholar]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar]
- Wang, Y.; Dai, Y.; Zhang, Y.; Hu, Y.; Yang, B.; Chen, S. Effects of quorum sensing autoinducer degradation gene on virulence and biofilm formation of Pseudomonas aeruginosa. Sci. China C Life Sci 2007, 50, 385–391. [Google Scholar]
- Cao, Y.; He, S.; Zhou, Z.; Zhang, M.; Mao, W.; Zhang, H.; Yao, B. Orally administered thermostable N-acyl homoserine lactonase from Bacillus sp. strain AI96 attenuates Aeromonas hydrophila infection in zebrafish. Appl. Environ. Microbiol 2012, 78, 1899–1908. [Google Scholar]
- Rasmussen, T.B.; Givskov, M. Quorum sensing inhibitors: A bargain of effects. Microbiology 2006, 152, 895–904. [Google Scholar]
- Leadbetter, J.R.; Greenberg, E.P. Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus. J. Bacteriol 2000, 182, 6921–6926. [Google Scholar]
- Lin, Y.-H.; Xu, J.-L.; Hu, J.; Wang, L.-H.; Ong, S.L.; Leadbetter, J.R.; Zhang, L.-H. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol. Microbiol 2003, 47, 849–860. [Google Scholar]
- Park, S.; Kang, H.; Jang, H.; Lee, J.-K.; Koo, B.; Yum, D. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol 2005, 71, 2632–2641. [Google Scholar]
- Dong, Y.; Wang, L.; Zhang, L.-H. Quorum-quenching microbial infections: Mechanisms and implications. Philos. Trans. R. Soc. Lond. B Biol. Sci 2007, 362, 1201–1211. [Google Scholar]
- Wahjudi, M.; Papaioannou, E.; Hendrawati, O.; van Assen, A.H.G.; van Merkerk, R.; Cool, R.H.; Poelarends, G.J.; Quax, W.J. PA0305 of Pseudomonas aeruginosa is a quorum quenching acylhomoserine lactone acylase belonging to the Ntn hydrolase superfamily. Microbiology 2011, 157, 2042–2055. [Google Scholar]
- Sio, C.F.; Otten, L.G.; Cool, R.H.; Diggle, S.P.; Braun, P.G.; Bos, R.; Daykin, M.; Camara, M.; Williams, P.; Quax, W.J. Quorum quenching by an N-acyl-homoserine lactone acylase from Pseudomonas aeruginosa PAO1. Infect. Immun 2006, 74, 1673–1682. [Google Scholar]
- Sio, C.F.; Riemens, A.M.; van der Laan, J.-M.; Verhaert, R.M.D.; Quax, W.J. Directed evolution of a glutaryl acylase into an adipyl acylase. FEBS J 2002, 269, 4495–4504. [Google Scholar]
- Papaioannou, E.; Wahjudi, M.; Nadal-Jimenez, P.; Koch, G.; Setroikromo, R.; Quax, W.J. Quorum-quenching acylase reduces the virulence of Pseudomonas aeruginosa in a Caenorhabditis elegans infection model. Antimicrob. Agents Chemother 2009, 53, 4891–4897. [Google Scholar]
- Bijtenhoorn, P.; Mayerhofer, H.; Müller-Dieckmann, J.; Utpatel, C.; Schipper, C.; Hornung, C.; Szesny, M.; Grond, S.; Thürmer, A.; Brzuszkiewicz, E.; et al. A novel metagenomic short-chain dehydrogenase/reductase attenuates Pseudomonas aeruginosa biofilm formation and virulence on Caenorhabditis elegans. PLoS One 2011, 6, e26278. [Google Scholar]
- Pustelny, C.; Albers, A.; Büldt-Karentzopoulos, K.; Parschat, K.; Chhabra, S.R.; Cámara, M.; Williams, P.; Fetzner, S. Dioxygenase-mediated quenching of quinolone-dependent quorum sensing in Pseudomonas aeruginosa. Chem. Biol 2009, 16, 1259–1267. [Google Scholar]
- Hentzer, M.; Wu, H.; Andersen, J.B.; Riedel, K.; Rasmussen, T.B.; Bagge, N.; Kumar, N.; Schembri, M.A.; Song, Z.; Kristoffersen, P.; et al. Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J 2003, 22, 3803–3815. [Google Scholar]
- Wu, H.; Song, Z.; Hentzer, M.; Andersen, J.B.; Molin, S.; Givskov, M.; Høiby, N. Synthetic furanones inhibit quorum-sensing and enhance bacterial clearance in Pseudomonas aeruginosa lung infection in mice. J. Antimicrob. Chemother 2004, 53, 1054–1061. [Google Scholar]
- Maeda, T.; García-Contreras, R.; Pu, M.; Sheng, L.; Garcia, L.R.; Tomás, M.; Wood, T.K. Quorum quenching quandary: Resistance to antivirulence compounds. ISME J 2012, 6, 493–501. [Google Scholar]
- Rasmussen, T.B.; Bjarnsholt, T.; Skindersoe, M.E.; Hentzer, M.; Kristoffersen, P.; Köte, M.; Nielsen, J.; Eberl, L.; Givskov, M.; Ko, M. Screening for quorum-sensing inhibitors (QSI) by use of a novel genetic system, the QSI selector. J. Bacteriol 2005, 187, 1799–1814. [Google Scholar]
- Jakobsen, T.H.; van Gennip, M.; Phipps, R.K.; Shanmugham, M.S.; Christensen, L.D.; Alhede, M.; Skindersoe, M.E.; Rasmussen, T.B.; Friedrich, K.; Uthe, F.; et al. Ajoene, a sulfur-rich molecule from garlic, inhibits genes controlled by quorum sensing. Antimicrob. Agents Chemother 2012, 56, 2314–2325. [Google Scholar]
- Bjarnsholt, T.; Jensen, P.Ø.; Rasmussen, T.B.; Christophersen, L.; Calum, H.; Hentzer, M.; Hougen, H.-P.; Rygaard, J.; Moser, C.; Eberl, L.; et al. Garlic blocks quorum sensing and promotes rapid clearing of pulmonary Pseudomonas aeruginosa infections. Microbiology 2005, 151, 3873–3880. [Google Scholar]
- Kobayashi, T.; Tateda, K.; Matsumoto, T.; Miyazaki, S.; Watanabe, A.; Nukiwa, T.; Yamaguchi, K. Macrolide-treated Pseudomonas aeruginosa induces paradoxical host responses in the lungs of mice and a high mortality rate. J. Antimicrob. Chemother 2002, 50, 59–66. [Google Scholar]
- Rasmussen, T.B.; Skindersoe, M.E.; Bjarnsholt, T.; Phipps, R.K.; Christensen, K.B.; Jensen, P.O.; Andersen, J.B.; Koch, B.; Larsen, T.O.; Hentzer, M.; et al. Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology 2005, 151, 1325–1340. [Google Scholar]
- Teplitski, M.; Robinson, J.B.; Bauer, W.D. Plants secrete substances that mimic bacterial N-acyl homoserine lactone signal activities and affect population density-dependent behaviors in associated bacteria. Mol. Plant Microbe Interact 2000, 13, 637–648. [Google Scholar]
- Jakobsen, T.H.; Bragason, S.K.; Phipps, R.K.; Christensen, L.D.; van Gennip, M.; Alhede, M.; Skindersoe, M.; Larsen, T.O.; Høiby, N.; Bjarnsholt, T.; et al. Food as a source for quorum sensing inhibitors: Iberin from horseradish revealed as a quorum sensing inhibitor of Pseudomonas aeruginosa. Appl. Environ. Microbiol 2012, 78, 2410–2421. [Google Scholar]
- Kandasamy, S.; Khan, W.; Evans, F.; Critchley, A.T.; Prithiviraj, B. Tasco®: A product of Ascophyllum nodosum enhances immune response of Caenorhabditis elegans against Pseudomonas aeruginosa infection. Mar. Drugs 2012, 10, 84–105. [Google Scholar]
- Krishnan, T.; Yin, W.-F.; Chan, K.-G. Inhibition of quorum sensing-controlled virulence factor production in Pseudomonas aeruginosa PAO1 by Ayurveda spice clove (Syzygium aromaticum) bud extract. Sensors 2012, 12, 4016–4030. [Google Scholar]
- Rasamiravaka, T.; Jedrzejowski, A.; Kiendrebeogo, M.; Rajaonson, S.; Randriamampionona, D.; Rabemanantsoa, C.; Andriantsimahavandy, A.; Rasamindrakotroka, A.; Duez, P.; El Jaziri, M.; et al. Endemic malagasy Dalbergia species inhibit quorum sensing in Pseudomonas aeruginosa PAO1. Microbiology 2013, 159, 924–938. [Google Scholar]
- Zhou, L.; Zheng, H.; Tang, Y.; Yu, W.; Gong, Q. Eugenol inhibits quorum sensing at sub-inhibitory concentrations. Biotechnol. Lett 2013, 35, 631–637. [Google Scholar]
- Sarabhai, S.; Sharma, P.; Capalash, N. Ellagic acid derivatives from Terminalia chebula Retz. downregulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa PAO1 virulence. PLoS One 2013, 8, e53441. [Google Scholar]
- Smyth, A.R.; Cifelli, P.M.; Ortori, C.A.; Righetti, K.; Lewis, S.; Erskine, P.; Holland, E.D.; Givskov, M.; Williams, P.; Cámara, M.; et al. Garlic as an inhibitor of Pseudomonas aeruginosa quorum sensing in cystic fibrosis-a pilot randomized controlled trial. Pediatr. Pulmonol 2010, 45, 356–362. [Google Scholar]
- Persson, T.; Hansen, T.H.; Rasmussen, T.B.; Skindersø, M.E.; Givskov, M.; Nielsen, J. Rational design and synthesis of new quorum-sensing inhibitors derived from acylated homoserine lactones and natural products from garlic. Org. Biomol. Chem 2005, 3, 253–262. [Google Scholar]
- Kim, C.; Kim, J.; Park, H.-Y.; Lee, J.-H.; Park, H.-J.; Kim, C.K.; Yoon, J. Structural understanding of quorum-sensing inhibitors by molecular modeling study in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol 2009, 83, 1095–1103. [Google Scholar]
- Ko, D.-H.; Kim, D.J.; Lyu, C.S.; Min, I.K.; Moon, H. New cleavage approaches to combinatorial synthesis of homoserine lactones. Tetrahedron Lett 1998, 39, 297–300. [Google Scholar]
- Yang, Y.-X.; Xu, Z.-H.; Zhang, Y.-Q.; Tian, J.; Weng, L.-X.; Wang, L.-H. A new quorum-sensing inhibitor attenuates virulence and decreases antibiotic resistance in Pseudomonas aeruginosa. J. Microbiol 2012, 50, 987–993. [Google Scholar]
- Müh, U.; Schuster, M.; Heim, R.; Singh, A.; Olson, E.R.; Greenberg, E.P. Novel Pseudomonas aeruginosa quorum-sensing inhibitors identified in an ultra-high-throughput screen. Antimicrob. Agents Chemother 2006, 50, 3674–3679. [Google Scholar]
- Ishida, T.; Ikeda, T.; Takiguchi, N.; Kuroda, A.; Ohtake, H.; Kato, J. Inhibition of quorum sensing in Pseudomonas aeruginosa by N-acyl cyclopentylamides. Appl. Environ. Microbiol 2007, 73, 3183–3188. [Google Scholar]
- Cady, N.C.; McKean, K.A.; Behnke, J.; Kubec, R.; Mosier, A.P.; Kasper, S.H.; Burz, D.S.; Musah, R.A. Inhibition of biofilm formation, quorum sensing and infection in Pseudomonas aeruginosa by natural products-inspired organosulfur compounds. PLoS One 2012, 7, e38492. [Google Scholar]
- Majik, M.S.; Naik, D.; Bhat, C.; Tilve, S.; Tilvi, S.; D’Souza, L. Synthesis of (R)-norbgugaine and its potential as quorum sensing inhibitor against Pseudomonas aeruginosa. Bioorg. Med. Chem. Lett 2013, 23, 2353–2356. [Google Scholar]
- Nalca, Y.; Ja, L.; Bredenbruch, F.; Geffers, R.; Buer, J.; Ha, S.; Jänsch, L.; Häussler, S. Quorum-sensing antagonistic activities of azithromycin in Pseudomonas aeruginosa PAO1: A global approach. Antimicrob. Agents Chemother 2006, 50, 1680–1688. [Google Scholar]
- Hansen, C.R.; Pressler, T.; Koch, C.; Høiby, N. Long-term azithromycin treatment of cystic fibrosis patients with chronic Pseudomonas aeruginosa infection; an observational cohort study. J. Cyst. Fibros 2005, 4, 35–40. [Google Scholar]
- Van Delden, C.; Köhler, T.; Brunner-Ferber, F.; François, B.; Carlet, J.; Pechère, J.-C. Azithromycin to prevent Pseudomonas aeruginosa ventilator-associated pneumonia by inhibition of quorum sensing: A randomized controlled trial. Intensive Care Med 2012, 38, 1118–1125. [Google Scholar]
- Mahajan-Miklos, S.; Rahme, L.G.; Ausubel, F.M. Elucidating the molecular mechanisms of bacterial virulence using non-mammalian hosts. Mol. Microbiol 2000, 37, 981–988. [Google Scholar]
- Utari, P.D.; Quax, W.J. Caenorhabditis elegans reveals novel Pseudomonas aeruginosa virulence mechanism. Trends Microbiol 2013, 21, 315–316. [Google Scholar]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol 2009, 7, 654–665. [Google Scholar]
- Collier, D.N.; Anderson, L.; McKnight, S.L.; Noah, T.L.; Knowles, M.; Boucher, R.; Schwab, U.; Gilligan, P.; Pesci, E.C. A bacterial cell to cell signal in the lungs of cystic fibrosis patients. FEMS Microbiol. Lett 2002, 215, 41–46. [Google Scholar]
- Erickson, D.L.; Endersby, R.; Kirkham, A.; Vollman, D.D.; Rabin, H.R.; Mitchell, I.; Storey, D.G.; Stuber, K. Pseudomonas aeruginosa quorum-sensing systems may control virulence factor expression in the lungs of patients with cystic fibrosis. Infect. Immun 2002, 70, 1783–1790. [Google Scholar]
- Singh, G.; Wu, B.; Baek, M.S.; Camargo, A.; Nguyen, A.; Slusher, N.A.; Srinivasan, R.; Wiener-Kronish, J.P.; Lynch, S.V. Secretion of Pseudomonas aeruginosa type III cytotoxins is dependent on pseudomonas quinolone signal concentration. Microb. Pathog 2010, 49, 196–203. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
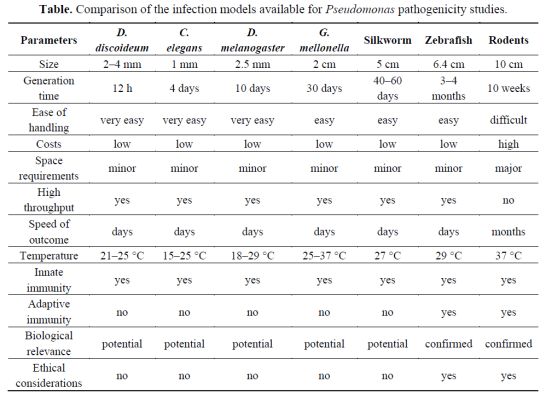
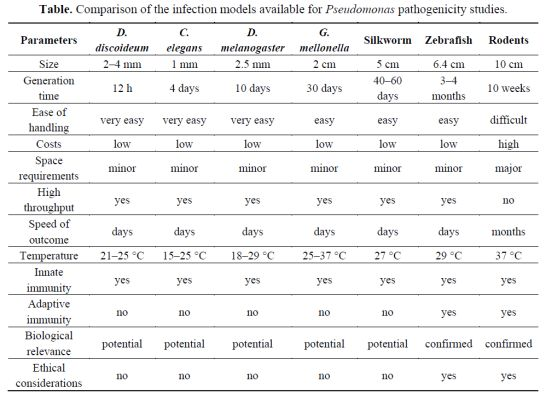
| Parameters | D. discoideum | C. elegans | D. melanogaster | G. mellonella | Silkworm | Zebrafish | Rodents |
|---|---|---|---|---|---|---|---|
| Size | 2–4 mm | 1 mm | 2.5 mm | 2 cm | 5 cm | 6.4 cm | 10 cm |
| Generation time | 12 h | 4 days | 10 days | 30 days | 40–60 days | 3–4 months | 10 weeks |
| Ease of handling | very easy | very easy | very easy | easy | easy | easy | difficult |
| Costs | low | low | low | low | low | low | high |
| Space requirements | minor | minor | minor | minor | minor | minor | major |
| High throughput | yes | yes | yes | yes | yes | yes | no |
| Speed of outcome | days | days | days | days | days | days | months |
| Temperature | 21–25 °C | 15–25 °C | 18–29 °C | 25–37 °C | 27 °C | 29 °C | 37 °C |
| Innate immunity | yes | yes | yes | yes | yes | yes | yes |
| Adaptive immunity | no | no | no | no | no | yes | yes |
| Biological relevance | potential | potential | potential | potential | potential | confirmed | confirmed |
| Ethical considerations | no | no | no | no | no | yes | yes |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Papaioannou, E.; Utari, P.D.; Quax, W.J. Choosing an Appropriate Infection Model to Study Quorum Sensing Inhibition in Pseudomonas Infections. Int. J. Mol. Sci. 2013, 14, 19309-19340. https://doi.org/10.3390/ijms140919309
Papaioannou E, Utari PD, Quax WJ. Choosing an Appropriate Infection Model to Study Quorum Sensing Inhibition in Pseudomonas Infections. International Journal of Molecular Sciences. 2013; 14(9):19309-19340. https://doi.org/10.3390/ijms140919309
Chicago/Turabian StylePapaioannou, Evelina, Putri Dwi Utari, and Wim J. Quax. 2013. "Choosing an Appropriate Infection Model to Study Quorum Sensing Inhibition in Pseudomonas Infections" International Journal of Molecular Sciences 14, no. 9: 19309-19340. https://doi.org/10.3390/ijms140919309
APA StylePapaioannou, E., Utari, P. D., & Quax, W. J. (2013). Choosing an Appropriate Infection Model to Study Quorum Sensing Inhibition in Pseudomonas Infections. International Journal of Molecular Sciences, 14(9), 19309-19340. https://doi.org/10.3390/ijms140919309