(+)-Dehydroabietic Acid, an Abietane-Type Diterpene, Inhibits Staphylococcus aureus Biofilms in Vitro

Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of Three Anti-Biofilms Compounds
2.2. Antibacterial versus Anti-Biofilm Effects of Compounds 1–3
2.3. Non-Specific Cytotoxicity on Mammalian Cells
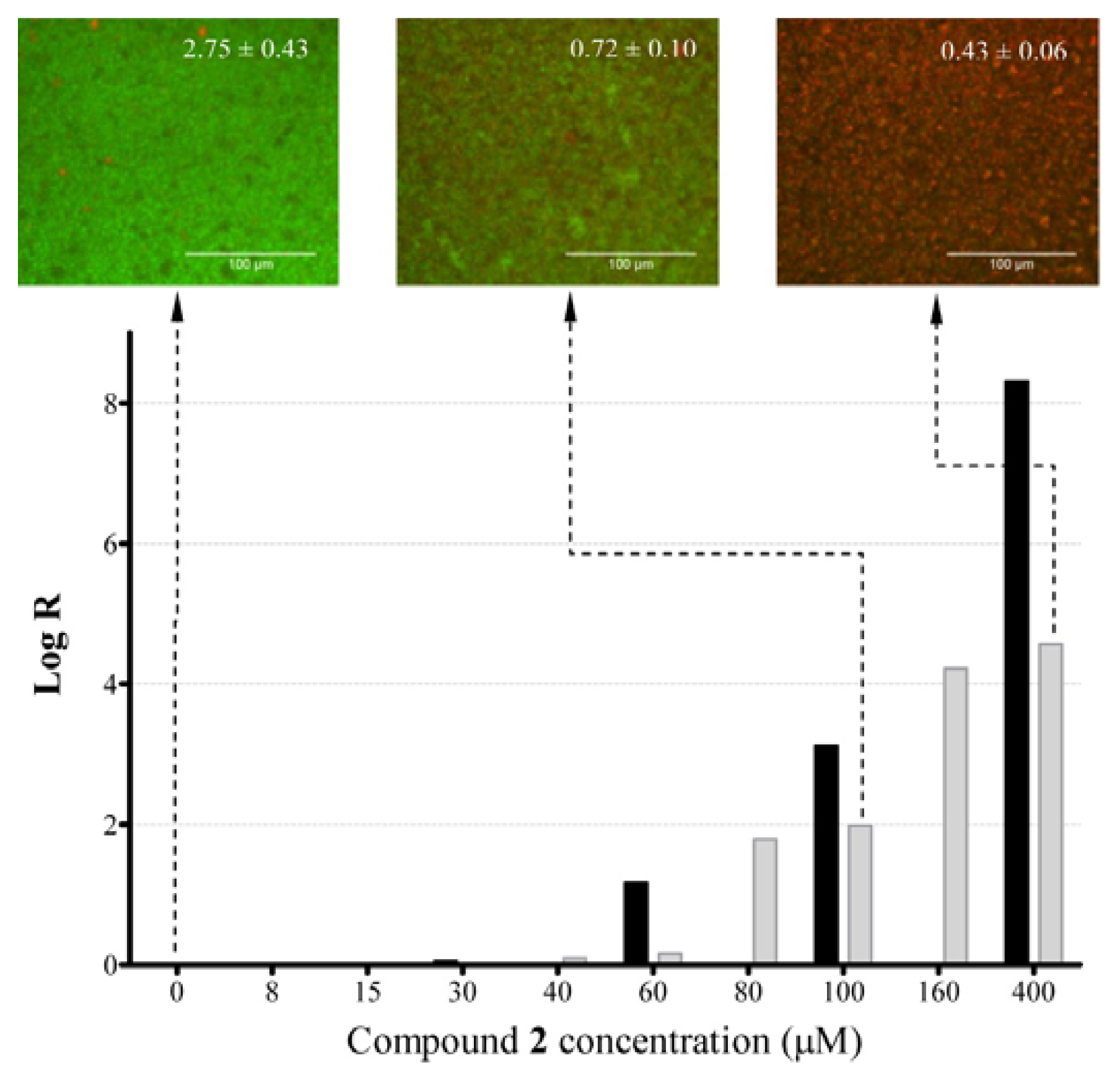
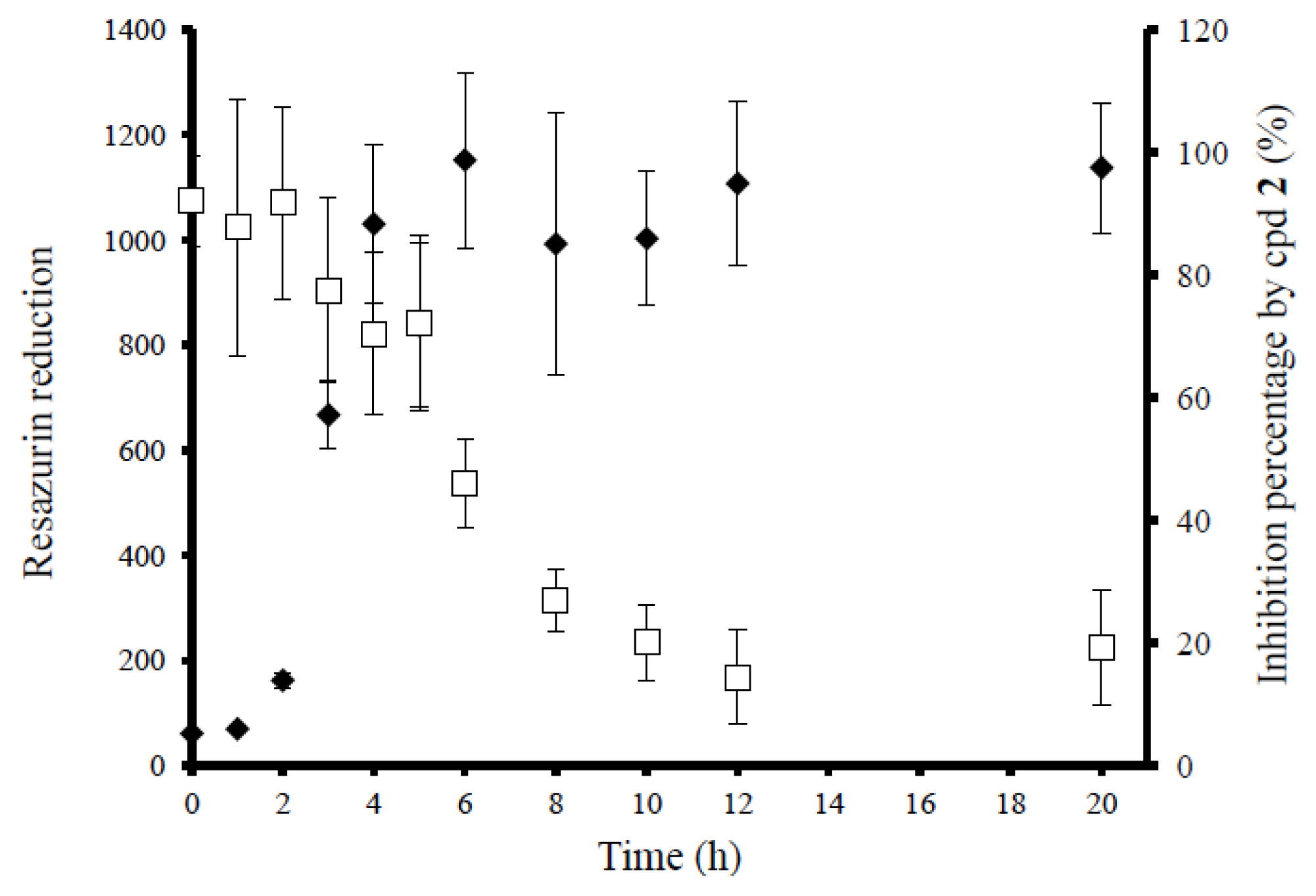
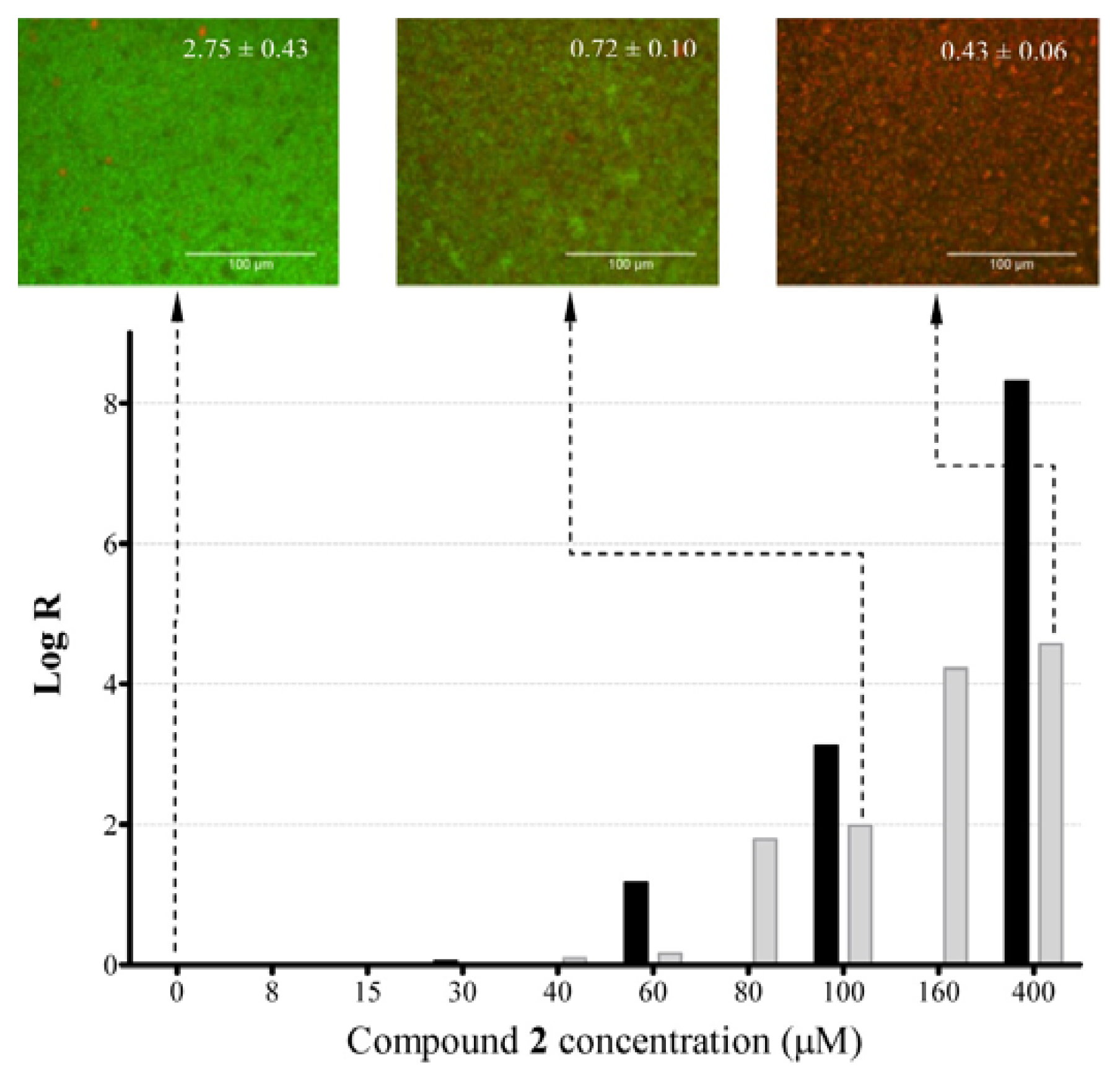
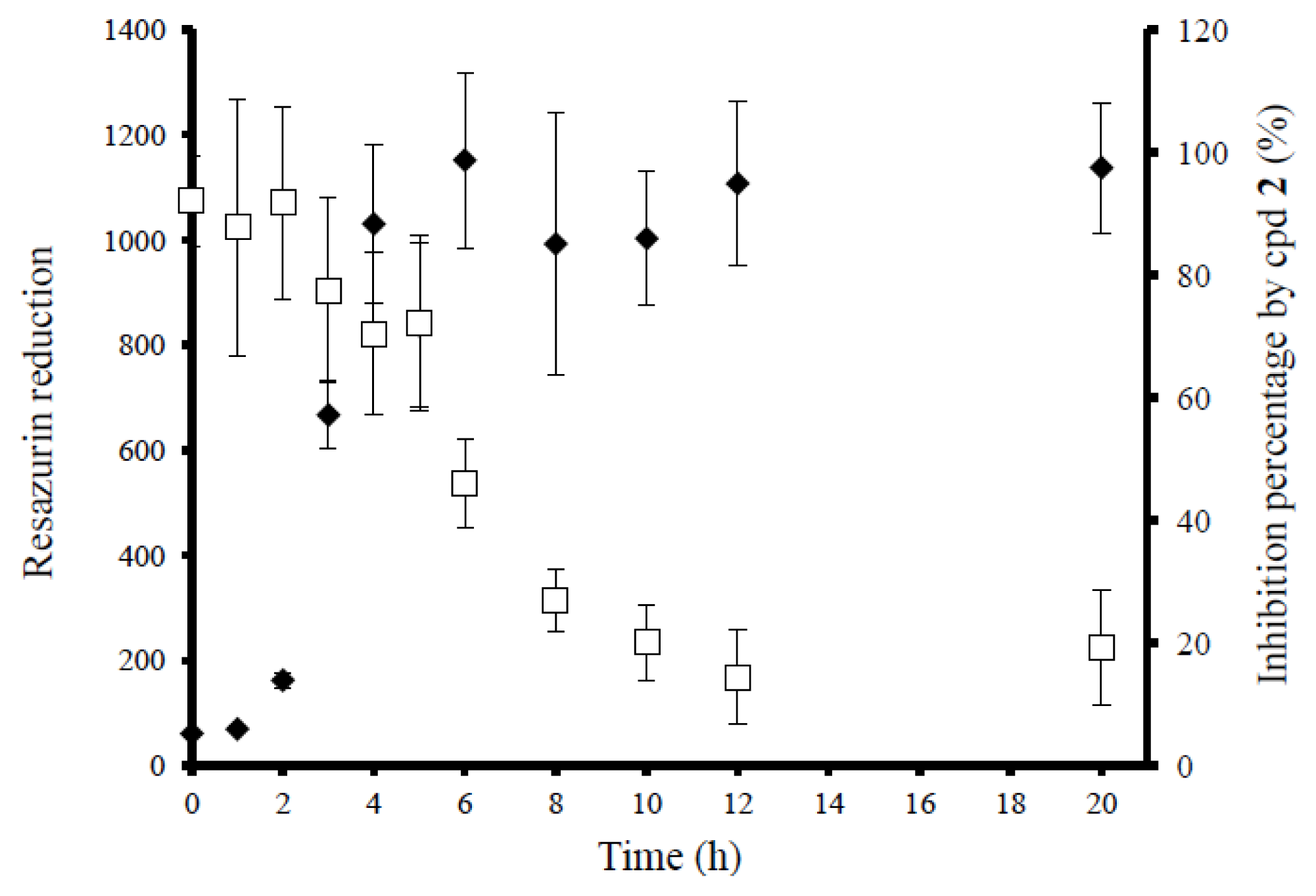
2.4. Anti-Biofilm Killing Efficacy of Compound 2 and Biocompatibility Index
2.5. Mechanistic Insights into Anti-Biofilm Activity of Compound 2
3. Experimental Section
3.1. Compounds Collection—Chemistry
3.2. Biofilm Formation Assay
3.3. Compounds Exposure
3.4. Quantification of Biofilms
3.5. Bacteriostatic Effect on Planktonic Cells
3.6. Cytotoxicity Assessment on Mammalian Cells
3.7. Studies on the Anti-Biofilm Activity of Compound 2
3.7.1. Determination of Log Reduction (Log R)
3.7.2. Imaging with Wide-Field Fluorescence Microscopy (FM)
3.7.3. Effect on Initial Bacterial Adhesion
3.7.4. Effects on Biofilm Proliferation or Maturation Phases
3.7.5. Bacterial Adhesion in the Presence of Host Proteins
3.8. Statistical Analysis and Data Processing
4. Conclusions
Acknowledgments
Appendix
Chemistry
Nordehydroabietylamine (1)
(+)-Dehydroabietic acid (2)
(+)-Dehydroabietylamine (3)
1-[4-(Benzyloxy)phenyl]-3-[[(1R,4aS)-7-isopropyl-1,4a-dimethyl-1,2,3,4,4a,9,10,10aoctahydrophenanthren- 1-yl]methyl]urea (4)
1-(3-Chlorophenyl)-3-[[(1R,4aS)-7-isopropyl-1,4a-dimethyl-1,2,3,4,4a,9,10,10aoctahydrophenanthren- 1-yl]methyl]urea (5)
1-[[(1R,4aS)-7-isopropyl-1,4a-dimethyl-1,2,3,4,4a,9,10,10a-octahydrophenanthren-1-yl]methyl]-3-(4- phenoxyphenyl)urea (6)
1-(4-Chloro-2-phenoxyphenyl)-3-[[(1R,4aS)-7-isopropyl-1,4a-dimethyl-1,2,3,4,4a,9,10,10aoctahydrophenanthren- 1-yl]methyl]urea (7)
1-(4-Chloro-2,5-dimethoxyphenyl)-3-[[(1R,4aS)-7-isopropyl-1,4a-dimethyl-1,2,3,4,4a,9,10,10aoctahydrophenanthren- 1-yl]methyl]urea (8)
1-(tert-Butyl)-3-[[(1R,4aS)-7-isopropyl-1,4a-dimethyl-1,2,3,4,4a,9,10,10a-octahydrophenanthren-1- yl]methyl]urea (9)
1-(Benzo[d][1,3]dioxol-5-yl)-3-[[(1R,4aS)-7-isopropyl-1,4a-dimethyl-1,2,3,4,4a,9,10,10aoctahydrophenanthren- 1-yl]methyl]urea (10)
Supplementary Video
Movie showing the interaction of compound 2 with S. aureus biofilms grown under flow conditions using Bioflux 200 (Fluxion Biosciences, CA, USA). Biofilms were grown in 48-well microplates (20 dyne/cm2) having microfluidic channels (370 μm × 70 μm) that were inoculated with S. aureus (106 CFU/mL) in 1:10 diluted TSB. Inoculation was performed by pumping at 2 dyne/cm2 for 2 s, from the “outlet” wells to the “inlet” wells to prevent contamination of the inlet wells. Following 1 h incubation for biofilm attachment (on static conditions) at 37 °C, flow was resumed at 0.4 dyne/cm2 for 18 h from inlet to outlet wells. After biofilm formation, compound 2 (100 μM, in 1:10 TSB) was added into the inlet wells and flow was restarted (0.2 dyne/cm2) for additional 24 h. The video (13 s) was automatically created from timelapse images taken every 1 s with a coupled Transmitted Light Evos ×l microscope (AMG, WA, USA) with 40× objective, 15 min after starting the exposure to 2.Conflict of Interest
References
- Toté, K.; Berghe, D.V.; Deschacht, M.; de Wit, K.; Maes, L.; Cos, P. Inhibitory efficacy of various antibiotics on matrix and viable mass of Staphylococcus aureus and Pseudomonas aeruginosa biofilms. Int. J. Antimicrob. Agents 2009, 33, 525–531. [Google Scholar]
- Melchior, M.B.; Fink-Gremmels, J.; Gaastra, W. Comparative assessment of the antimicrobial susceptibility of Staphylococcus aureus isolates from bovine mastitis in biofilm versus planktonic culture. J. Vet. Med. B 2006, 53, 326–332. [Google Scholar]
- Girard, L.P.; Ceri, H.; Gibb, A.P.; Olson, M.; Sepandj, F. MIC versus MBEC to determine the antibiotic sensitivity of Staphylococcus aureus in peritoneal dialysis peritonitis. Perit. Dial. Int 2010, 30, 652–656. [Google Scholar]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev 2002, 15, 167–193. [Google Scholar]
- Parsek, M.R.; Singh, P.K. Bacterial biofilms: An emerging link to disease pathogenesis. Annu. Rev. Microbiol 2003, 57, 677–701. [Google Scholar]
- Worthington, R.J.; Richards, J.J.; Melander, C. Small molecule control of bacterial biofilms. Org. Biomol. Chem 2012, 10, 7457–7474. [Google Scholar]
- Landini, P.; Antoniani, D.; Burgess, J.G.; Nijland, R. Molecular mechanisms of compounds affecting bacterial biofilm formation and dispersal. Appl. Microbiol. Biotechnol 2010, 86, 813–823. [Google Scholar]
- Skogman, M.E.; Vuorela, P.M.; Fallarero, A. Combining biofilm matrix measurements with biomass and viability assays in susceptibility assessments of antimicrobials against Staphylococcus aureus biofilms. J. Antibiot 2012, 65, 453–459. [Google Scholar]
- Phillips, M.A.; Croteau, R.B. Resin-based defenses in conifers. Trends Plant Sci 1999, 4, 184–190. [Google Scholar]
- Gaspar-Marques, C.; Rijo, P.; Simões, M.F.; Duarte, M.A.; Rodriguez, B. Abietanes from Plectranthus grandidentatus and P. hereroensis against methicillin- and vancomycin-resistant bacteria. Phytomedicine 2006, 13, 267–271. [Google Scholar]
- Oluwatuyi, M.; Kaatz, G.W.; Gibbons, S. Antibacterial and resistance modifying activity of Rosmarinus officinalis. Phytochemistry 2004, 65, 3249–3254. [Google Scholar]
- Yang, Z.; Kitano, Y.; Chiba, K.; Shibata, N.; Kurokawa, H.; Doi, Y.; Arakawa, Y.; Tada, M. Synthesis of variously oxidized abietane diterpenes and their antibacterial activities against MRSA and VRE. Bioorg. Med. Chem 2001, 9, 347–356. [Google Scholar]
- Ali, F.; Sangwan, P.L.; Koul, S.; Pandey, A.; Bani, S.; Abdullah, S.T.; Sharma, P.R.; Kitchlu, S.; Khan, I.A. 4-epi-pimaric acid: A phytomolecule as a potent antibacterial and anti-biofilm agent for oral cavity pathogens. Eur. J. Clin. Microbiol. Infect. Dis 2012, 31, 149–159. [Google Scholar]
- KuŸma, Ł.; Rózalski, M.; Walencka, E.; Rózalska, B.; Wysokińska, H. Antimicrobial activity of diterpenoids from hairy roots of Salvia sclarea l.: Salvipisone as a potential anti-biofilm agent active against antibiotic resistant Staphylococci. Phytomedicine 2007, 14, 31–35. [Google Scholar]
- Esteban, J.; Molina-Manso, D.; Spiliopoulou, I.; Cordero-Ampuero, J.; Fernández-Roblas, R.; Foka, A.; Gómez-Barrena, E. Biofilm development by clinical isolates of Staphylococcus spp. from retrieved orthopedic prostheses. Acta Orthop 2010, 81, 674–679. [Google Scholar]
- Lorenz, U.; Schäfer, T.; Ohlsen, K.; Tiurbe, G.C.; Bühler, C.; Germer, C.T.; Kellersmann, R. In vivo detection of Staphylococcus aureus in biofilm on vascular prostheses using non-invasive biophotonic imaging. Eur. J. Vasc. Endovasc. Surg 2011, 41, 68–75. [Google Scholar]
- Donlan, R.M. Biofilm elimination on intravascular catheters: Important considerations for the infectious disease practitioner. Clin. Infect. Dis 2011, 52, 1038–1045. [Google Scholar]
- Baveja, J.K.; Willcox, M.D.; Hume, E.B.; Kumar, N.; Odell, R.; Poole-Warren, L.A. Furanones as potential anti-bacterial coatings on biomaterials. Biomaterials 2004, 25, 5003–5012. [Google Scholar]
- Opperman, T.J.; Kwasny, S.M.; Williams, J.D.; Khan, A.R.; Peet, N.P.; Moir, D.T.; Bowlin, T.L. Aryl rhodanines specifically inhibit Staphylococcal and Enterococcal biofilm formation. Antimicrob. Agents Chemother 2009, 53, 4357–4367. [Google Scholar]
- Lin, M.H.; Chang, F.R.; Hua, M.Y.; Wu, Y.C.; Liu, S.T. Inhibitory effects of 1,2,3,4,6-penta-o-galloyl-beta-d-glucopyranose on biofilm formation by Staphylococcus aureus. Antimicrob. Agents Chemother 2011, 55, 1021–1027. [Google Scholar]
- Lönn-Stensrud, J.; Landin, M.; Benneche, T.; Petersen, F.; Scheie, A. Furanones, potential agents for preventing Staphylococcus epidermidis biofilm infections? J. Antimicrob. Chemother 2009, 63, 309–316. [Google Scholar]
- Sandberg, M.E.; Schellmann, D.; Brunhofer, G.; Erker, T.; Busygin, I.; Leino, R.; Vuorela, P.M.; Fallarero, A. Pros and cons of using resazurin staining for quantification of viable Staphylococcus aureus biofilms in a screening assay. J. Microbiol. Methods 2009, 78, 104–106. [Google Scholar]
- Saginur, R.; Stdenis, M.; Ferris, W.; Aaron, S.D.; Chan, F.; Lee, C.; Ramotar, K. Multiple combination bactericidal testing of Staphylococcal biofilms from implant-associated infections. Antimicrob. Agents Chemother 2006, 50, 55–61. [Google Scholar]
- Olson, M.E.; Ceri, H.; Morck, D.W.; Buret, A.G.; Read, R.R. Biofilm bacteria: Formation and comparative susceptibility to antibiotics. Can. J. Vet. Res 2002, 66, 86–92. [Google Scholar]
- Ayars, G.H.; Altman, L.C.; Frazier, C.E.; Chi, E.Y. The toxicity of constituents of cedar and pine woods to pulmonary epithelium. J. Allergy Clin. Immunol 1989, 83, 610–618. [Google Scholar]
- Sepúlveda, B.; Astudillo, L.; Rodríguez, J.A.; Yáñez, T.; Theoduloz, C.; Schmeda-Hirschmann, G. Gastroprotective and cytotoxic effect of dehydroabietic acid derivatives. Pharmacol. Res 2005, 52, 429–437. [Google Scholar]
- Pitts, B.; Hamilton, M.A.; Zelver, N.; Stewart, P.S. A microtiter-plate screening method for biofilm disinfection and removal. J. Microbiol. Methods 2003, 54, 269–276. [Google Scholar]
- Müller, G.; Kramer, A. Biocompatibility index of antiseptic agents by parallel assessment of antimicrobial activity and cellular cytotoxicity. J. Antimicrob. Chemother 2008, 61, 1281–1287. [Google Scholar]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol 2004, 2, 95–108. [Google Scholar]
- Haraguchi, H.; Oike, S.; Muroi, H.; Kubo, I. Mode of antibacterial action of totarol, a diterpene from Podocarpus nagi. Planta. Med 1996, 62, 122–125. [Google Scholar]
- Shinji, H.; Yosizawa, Y.; Tajima, A.; Iwase, T.; Sugimoto, S.; Seki, K.; Mizunoe, Y. Role of fibronectin-binding proteins a and b in in vitro cellular infections and in vivo septic infections by Staphylococcus aureus. Infect. Immun 2011, 79, 2215–2223. [Google Scholar]
- Vazquez, V.; Liang, X.; Horndahl, J.K.; Ganesh, V.K.; Smeds, E.; Foster, T.J.; Hook, M. Fibrinogen is a ligand for the S. aureus MSCRAMM bbp (bone sialoprotein-binding protein). J. Biol. Chem. 2011. [Google Scholar] [CrossRef]
- Ganesh, V.K.; Barbu, E.M.; Deivanayagam, C.C.; Le, B.; Anderson, A.S.; Matsuka, Y.V.; Lin, S.L.; Foster, T.J.; Narayana, S.V.; Hook, M. Structural and biochemical characterization of Staphylococcus aureus clumping factor b:Ligand interactions. J. Biol. Chem 2011, 286, 25963–25972. [Google Scholar]
- Sandberg, M.; Määttänen, A.; Peltonen, J.; Vuorela, P.M.; Fallarero, A. Automating a 96-well microtitre plate model for Staphylococcus aureus biofilms: An approach to screening of natural antimicrobial compounds. Int. J. Antimicrob. Agents 2008, 32, 233–240. [Google Scholar]
- Karlsson, D.; Fallarero, A.; Brunhofer, G.; Mayer, C.; Prakash, O.; Mohan, C.G.; Vuorela, P.; Erker, T. The exploration of thienothiazines as selective butyrylcholinesterase inhibitors. Eur. J. Pharm. Sci 2012, 47, 190–205. [Google Scholar]
- Ozaki, S. Recent advances in isocyanate chemistry. Chem. Rev 1972, 72, 457–496. [Google Scholar]

{kind=link}
{kind=link}
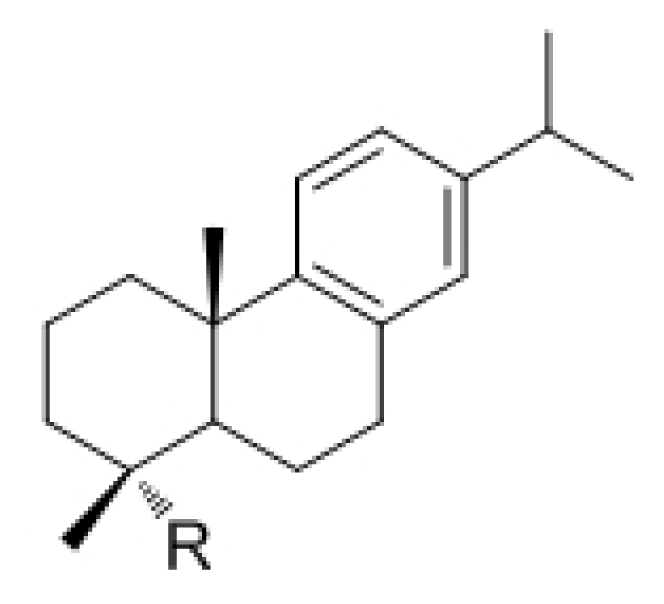{kind=link}
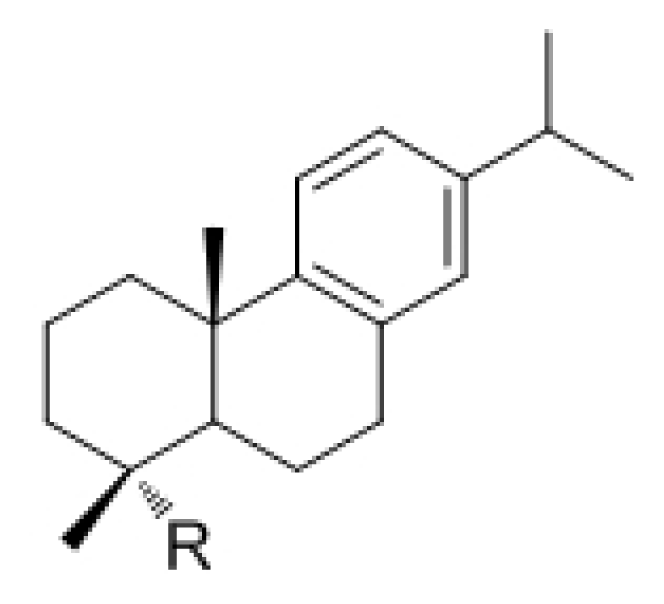 | |||
|---|---|---|---|
| Code | R | Code | R |
| 1 | NH2 | 6 | CH2NHCONH-p-(C6H4)OPh |
| 2 | CO2H | 7 | CH2NHCONH(C6H3)(3-OPh)(4-Cl) |
| 3 | CH2NH2 | 8 | CH2NHCONH(C6H2)(3,6-OMe)(4-Cl) |
| 4 | CH2NHCONH-p-(C6H4)OBn | 9 | CH2NHCONH(tert-Bu) |
| 5 | CH2NHCONH-m-(C6H4)Cl | 10 | CH2NHCONH-5-(benzo-1,3-dioxolyl) |
| Effects on biofilms | Effect on planktonic Bacteria3 | ||||
|---|---|---|---|---|---|
| IC50, μM [mg/L] (95% confidence intervals) | MIC, μM [mg/L] | ||||
| Cpd | Prior to biofilm formation 1 | Post-biofilm formation 2 | Turbidity | ||
| Viability | Biomass | Viability | Biomass | ||
| 1 | 124.0 [33.7] (114.6–134.1) | 115.6 [31.4] (105.7–126.5) | 288.4 [78.3] (266.2–312.5) | 293.7 [79.7] (183.1–471.0) | 160 [43.4] |
| 2 | 27.8 [8.35] (20.2–38.2) | 33.9 [10.2] (23.9–48.3) | 112.8 [33.9] (92.9–132.5) | 81.2 [24.4] (50.9–129.2) | 70 [21.0] |
| 3 | 28.2 [8.05] (26.3–30.2) | 17.4 [33.7] (15.5–19.5) | 83.4 [23.8] (73.6–94.5) | 74.4 [21.2] (70.8–78.2) | 40 [11.4] |
| Penicillin | 0.13 [0.048] (0.12–0.13) | 0.024 [0.0089] (0.02–0.03) | 45.2% a | 56.5% a | 0.12 [0.045] |
| Cell line | Cytotoxicity (LC50) μM [mg/L] | Biocompatibility Index * |
|---|---|---|
| GT1-7 | 176.3 [52.9] | 1.3 |
| HepG2 | 153.0 [45.9] | 1.1 |
| HL | 106.4 [31.9] | 0.76 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fallarero, A.; Skogman, M.; Kujala, J.; Rajaratnam, M.; Moreira, V.M.; Yli-Kauhaluoma, J.; Vuorela, P. (+)-Dehydroabietic Acid, an Abietane-Type Diterpene, Inhibits Staphylococcus aureus Biofilms in Vitro. Int. J. Mol. Sci. 2013, 14, 12054-12072. https://doi.org/10.3390/ijms140612054
Fallarero A, Skogman M, Kujala J, Rajaratnam M, Moreira VM, Yli-Kauhaluoma J, Vuorela P. (+)-Dehydroabietic Acid, an Abietane-Type Diterpene, Inhibits Staphylococcus aureus Biofilms in Vitro. International Journal of Molecular Sciences. 2013; 14(6):12054-12072. https://doi.org/10.3390/ijms140612054
Chicago/Turabian StyleFallarero, Adyary, Malena Skogman, Janni Kujala, Mohanathas Rajaratnam, Vânia M. Moreira, Jari Yli-Kauhaluoma, and Pia Vuorela. 2013. "(+)-Dehydroabietic Acid, an Abietane-Type Diterpene, Inhibits Staphylococcus aureus Biofilms in Vitro" International Journal of Molecular Sciences 14, no. 6: 12054-12072. https://doi.org/10.3390/ijms140612054
APA StyleFallarero, A., Skogman, M., Kujala, J., Rajaratnam, M., Moreira, V. M., Yli-Kauhaluoma, J., & Vuorela, P. (2013). (+)-Dehydroabietic Acid, an Abietane-Type Diterpene, Inhibits Staphylococcus aureus Biofilms in Vitro. International Journal of Molecular Sciences, 14(6), 12054-12072. https://doi.org/10.3390/ijms140612054