Wharton’s Jelly-Derived Mesenchymal Stem Cells: Phenotypic Characterization and Optimizing Their Therapeutic Potential for Clinical Applications


{kind=link}
Abstract
:1. Introduction
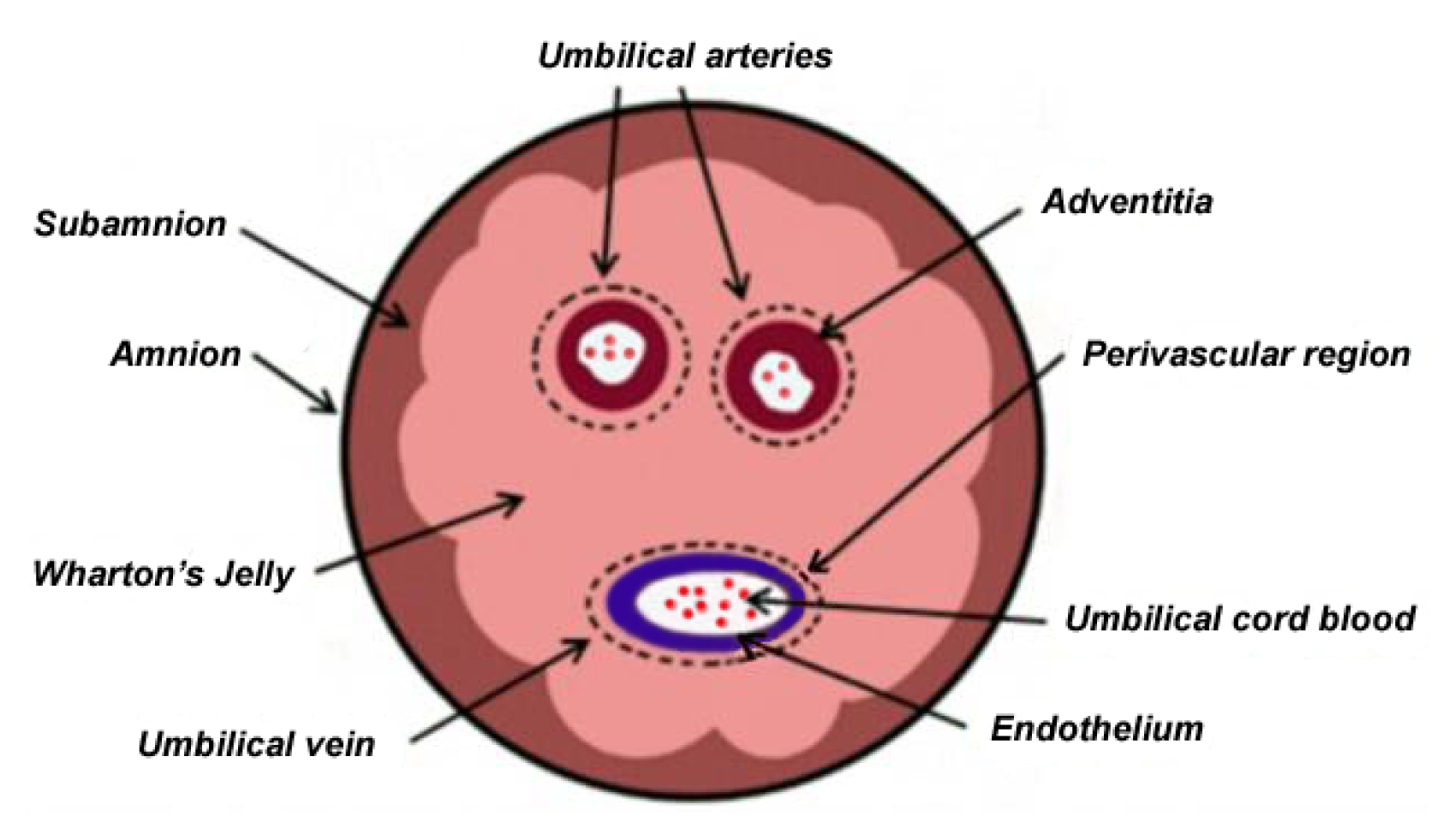
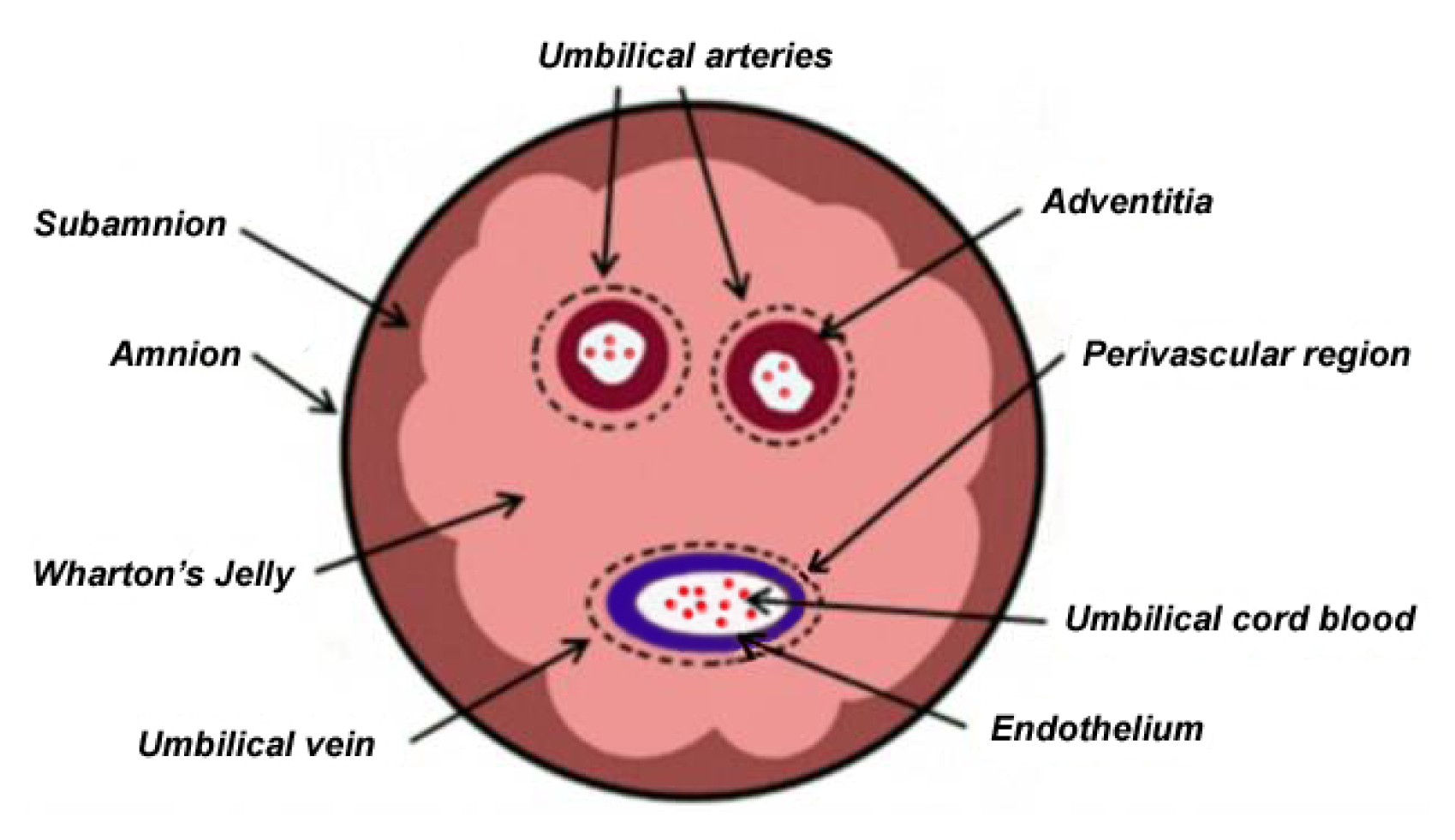
2. Anatomical Relationship of Various UC Structures and WJ as Sources of MSCs
3. Characteristic Features of WJ-MSCs for Cell Therapy
3.1. Sources of Stem Cells
3.2. Immunomodulatory Property of WJ
3.3. Phenotypic Characterization of WJ
4. Clinical Applications of WJ-Derived Stem Cells
4.1. Cancer Therapy
4.2. Liver Disease
4.3. Cardiovascular Diseases
4.4. Cartilage Regeneration
4.5. Peripheral Nerve Repair
4.6. Cardiac Differentiation of Human WJ-Derived Stem Cells
5. The New Research Frontiers in WJ Research
5.1. Clonal MSCs
5.2. Use of Magnetic Resonance Imaging in Contrast Labeled-UC Stem Cells
6. Conclusions
Acknowledgments
Conflict of Interest
References
- La Rocca, G. Connecting the dots: The promises of wharton’s jelly stem cells for tissue repair and regeneration. Open Tissue Eng. Regen. Med. J 2011, 4, 3–5. [Google Scholar]
- McElreavey, K.D.; Irvine, A.I.; Ennis, K.T.; McLean, W.H. Isolation, culture and characterisation of fibroblast-like cells derived from the wharton’s jelly portion of human umbilical cord. Biochem. Soc. Trans 1991, 19, 29S. [Google Scholar]
- Wharton, T. Adenographia; Freer, S., Translator; Oxford, UK; Oxford University Press, 1996. [Google Scholar]
- Chacko, A.W.; Reynolds, S.R.M. Architecture of distended and nondistended human umbilical cord tissues, with special reference to the arteries and veins. Contrib. Embryol 1954, 35, 135–150. [Google Scholar]
- Kadner, A.; Hoerstrup, S.P.; Tracy, J.; Breymann, C.; Maurus, C.F.; Melnitchouk, S.; Kadner, G.; Zund, G.; Turina, M. Human umbilical cord cells: A new cell source for cardiovascular tissue engineering. Ann. Thorac. Surg 2002, 74, S1422–S1428. [Google Scholar]
- Kadner, A.; Zund, G.; Maurus, C.; Breymann, C.; Yakarisik, S.; Kadner, G.; Turina, M.; Hoerstrup, S.P. Human umbilical cord cells for cardiovascular tissue engineering: A comparative study. Eur. J. Cardio-Thorac. Surg 2004, 25, 635–641. [Google Scholar]
- Mitchell, K.E.; Weiss, M.L.; Mitchell, B.M.; Martin, P.; Davis, D.; Morales, L.; Helwig, B.; Beerenstrauch, M.; Abou-Easa, K.; Hildreth, T.; et al. Matrix cells from wharton’s jelly form neurons and glia. Stem Cells 2003, 21, 50–60. [Google Scholar]
- Naughton, B.A.; San Roman, J.; Liu, K.; Purchio, A.; Pavelec, R.; Rekettye, L. Cells isolated from wharton’s jelly of the human umbilical cord develop a cartilage phenotype when treated with tgf-β in vitro. FASEB J 1997, 11, A19. [Google Scholar]
- Purchio, A.F.; Naughton, B.A.; Roman, J.S. Production of Cartilage Tissue Using Cells Isolated from Wharton’s Jelly. U.S. Patent 5,919,702, 1999. [Google Scholar]
- Romanov, Y.A.; Svintsitskaya, V.A.; Smirnov, V.N. Searching for alternative sources of postnatal human mesenchymal stem cells: Candidate msc-like cells from umbilical cord. Stem Cells 2003, 21, 105–110. [Google Scholar]
- Takechi, K.; Kuwabara, Y.; Mizuno, M. Ultrastructural and immunohistochemical studies of wharton’s jelly umbilical cord cells. Placenta 1993, 14, 235–245. [Google Scholar]
- Kobayashi, K.; Kubota, T.; Aso, T. Study on myofibroblast differentiation in the stromal cells of wharton’s jelly: Expression and localization of alpha-smooth muscle actin. Early Hum. Dev 1998, 51, 223–233. [Google Scholar]
- Markov, V.; Kusumi, K.; Tadesse, M.G.; William, D.A.; Hall, D.M.; Lounev, V.; Carlton, A.; Leonard, J.; Cohen, R.I.; Rappaport, E.F.; et al. Identification of cord blood-derived mesenchymal stem/stromal cell populations with distinct growth kinetics, differentiation potentials, and gene expression profiles. Stem Cells Dev 2007, 16, 53–73. [Google Scholar]
- Baudin, B.; Bruneel, A.; Bosselut, N.; Vaubourdolle, M. A protocol for isolation and culture of human umbilical vein endothelial cells. Nat. Protoc 2007, 2, 481–485. [Google Scholar]
- Wang, X.Y.; Lan, Y.; He, W.Y.; Zhang, L.; Yao, H.Y.; Hou, C.M.; Tong, Y.; Liu, Y.L.; Yang, G.; Liu, X.D.; et al. Identification of mesenchymal stem cells in aorta-gonad-mesonephros and yolk sac of human embryos. Blood 2008, 111, 2436–2443. [Google Scholar]
- Bongso, A.; Fong, C.Y. The therapeutic potential, challenges and future clinical directions of stem cells from the wharton’s jelly of the human umbilical cord. Stem Cell Rev 2013, 9, 226–240. [Google Scholar]
- Jeschke, M.G.; Gauglitz, G.G.; Phan, T.T.; Herndon, D.N.; Kita, K. Umbilical cord lining membrane and wharton’s jelly-derived mesenchymal stem cells: The similarities and differences. Open Tissue Eng. Regen. Med. J 2011, 4, 21–27. [Google Scholar]
- Prasanna, S.J.; Jahnavi, V.S. Wharton’s jelly mesenchymal stem cells as off-the -shelf cellular therapeutics: A closer look into their regenerative and immunomodulatory properties. Open Tissue Eng. Regen. Med. J 2011, 4, 28–38. [Google Scholar]
- Troyer, D.L.; Weiss, M.L. Wharton’s jelly-derived cells are a primitive stromal cell population. Stem Cells 2008, 26, 591–599. [Google Scholar]
- Karahuseyinoglu, S.; Cinar, O.; Kilic, E.; Kara, F.; Akay, G.G.; Demiralp, D.O.; Tukun, A.; Uckan, D.; Can, A. Biology of stem cells in human umbilical cord stroma: In situ and in vitro surveys. Stem Cells 2007, 25, 319–331. [Google Scholar]
- Nanaev, A.K.; Kohnen, G.; Milovanov, A.P.; Domogatsky, S.P.; Kaufmann, P. Stromal differentiation and architecture of the human umbilical cord. Placenta 1997, 18, 53–64. [Google Scholar]
- Can, A.; Karahuseyinoglu, S. Concise review: Human umbilical cord stroma with regard to the source of fetus-derived stem cells. Stem Cells 2007, 25, 2886–2895. [Google Scholar]
- Baksh, D.; Yao, R.; Tuan, R.S. Comparison of proliferative and multilineage differentiation potential of human mesenchymal stem cells derived from umbilical cord and bone marrow. Stem Cells 2007, 25, 1384–1392. [Google Scholar]
- Sarugaser, R.; Lickorish, D.; Baksh, D.; Hosseini, M.M.; Davies, J.E. Human umbilical cord perivascular (hucpv) cells: A source of mesenchymal progenitors. Stem Cells 2005, 23, 220–229. [Google Scholar]
- Kita, K.; Gauglitz, G.G.; Phan, T.T.; Herndon, D.N.; Jeschke, M.G. Isolation and characterization of mesenchymal stem cells from the sub-amniotic human umbilical cord lining membrane. Stem Cells Dev 2010, 19, 491–502. [Google Scholar]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar]
- Baldwin, T. Morality and human embryo research. Introduction to the talking point on morality and human embryo research. EMBO Rep 2009, 10, 299–300. [Google Scholar]
- Laurent, L.C.; Ulitsky, I.; Slavin, I.; Tran, H.; Schork, A.; Morey, R.; Lynch, C.; Harness, J.V.; Lee, S.; Barrero, M.J.; et al. Dynamic changes in the copy number of pluripotency and cell proliferation genes in human escs and ipscs during reprogramming and time in culture. Cell Stem Cell 2011, 8, 106–118. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar]
- Gutierrez-Aranda, I.; Ramos-Mejia, V.; Bueno, C.; Munoz-Lopez, M.; Real, P.J.; Macia, A.; Sanchez, L.; Ligero, G.; Garcia-Parez, J.L.; Menendez, P. Human induced pluripotent stem cells develop teratoma more efficiently and faster than human embryonic stem cells regardless the site of injection. Stem Cells 2010, 28, 1568–1570. [Google Scholar]
- Ogawa, M.; Larue, A.C.; Mehrotra, M. Hematopoietic stem cells are pluripotent and not just “hematopoietic”. Blood Cells Mol. Dis 2013, 51, 3–8. [Google Scholar]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar]
- Semenov, O.V.; Koestenbauer, S.; Riegel, M.; Zech, N.; Zimmermann, R.; Zisch, A.H.; Malek, A. Multipotent mesenchymal stem cells from human placenta: Critical parameters for isolation and maintenance of stemness after isolation. Am. J. Obstet. Gynecol 2010, 202, e191–e193. [Google Scholar]
- Ilancheran, S.; Moodley, Y.; Manuelpillai, U. Human fetal membranes: A source of stem cells for tissue regeneration and repair? Placenta 2009, 30, 2–10. [Google Scholar]
- Strakova, Z.; Livak, M.; Krezalek, M.; Ihnatovych, I. Multipotent properties of myofibroblast cells derived from human placenta. Cell Tissue Res 2008, 332, 479–488. [Google Scholar]
- Pappa, K.I.; Anagnou, N.P. Novel sources of fetal stem cells: Where do they fit on the developmental continuum? Regen. Med 2009, 4, 423–433. [Google Scholar]
- Marcus, A.J.; Woodbury, D. Fetal stem cells from extra-embryonic tissues: Do not discard. J. Cell. Mol. Med 2008, 12, 730–742. [Google Scholar]
- Prasanna, S.J.; Gopalakrishnan, D.; Shankar, S.R.; Vasandan, A.B. Pro-inflammatory cytokines, ifngamma and tnfalpha, influence immune properties of human bone marrow and wharton jelly mesenchymal stem cells differentially. PLoS One 2010, 5, e9016. [Google Scholar]
- La Rocca, G.; Anzalone, R.; Corrao, S.; Magno, F.; Loria, T.; Lo Iacono, M.; Di Stefano, A.; Giannuzzi, P.; Marasa, L.; Cappello, F.; et al. Isolation and characterization of oct-4+/hla-g+ mesenchymal stem cells from human umbilical cord matrix: Differentiation potential and detection of new markers. Histochem. Cell Biol 2009, 131, 267–282. [Google Scholar]
- Rachakatla, R.S.; Pyle, M.M.; Ayuzawa, R.; Edwards, S.M.; Marini, F.C.; Weiss, M.L.; Tamura, M.; Troyer, D. Combination treatment of human umbilical cord matrix stem cell-based interferon-beta gene therapy and 5-fluorouracil significantly reduces growth of metastatic human breast cancer in scid mouse lungs. Cancer Investig 2008, 26, 662–670. [Google Scholar]
- Weiss, M.L.; Anderson, C.; Medicetty, S.; Seshareddy, K.B.; Weiss, R.J.; VanderWerff, I.; Troyer, D.; McIntosh, K.R. Immune properties of human umbilical cord wharton’s jelly-derived cells. Stem Cells 2008, 26, 2865–2874. [Google Scholar]
- Deuse, T.; Stubbendorff, M.; Tang-Quan, K.; Phillips, N.; Kay, M.A.; Eiermann, T.; Phan, T.T.; Volk, H.D.; Reichenspurner, H.; Robbins, R.C.; et al. Immunogenicity and immunomodulatory properties of umbilical cord lining mesenchymal stem cells. Cell Transplant 2011, 20, 655–667. [Google Scholar]
- Selmani, Z.; Naji, A.; Zidi, I.; Favier, B.; Gaiffe, E.; Obert, L.; Borg, C.; Saas, P.; Tiberghien, P.; Rouas-Freiss, N.; et al. Human leukocyte antigen-g5 secretion by human mesenchymal stem cells is required to suppress t lymphocyte and natural killer function and to induce cd4+cd25highfoxp3+ regulatory t cells. Stem Cells 2008, 26, 212–222. [Google Scholar]
- Griffin, M.D.; Ritter, T.; Mahon, B.P. Immunological aspects of allogeneic mesenchymal stem cell therapies. Hum. Gene Ther 2010, 21, 1641–1655. [Google Scholar]
- Zarkhin, V.; Talisetti, A.; Li, L.; Wozniak, L.J.; McDiarmid, S.V.; Cox, K.; Esquivel, C.; Sarwal, M.M. Expression of soluble hla-g identifies favorable outcomes in liver transplant recipients. Transplantation 2010, 90, 1000–1005. [Google Scholar]
- Djouad, F.; Charbonnier, L.M.; Bouffi, C.; Louis-Plence, P.; Bony, C.; Apparailly, F.; Cantos, C.; Jorgensen, C.; Noel, D. Mesenchymal stem cells inhibit the differentiation of dendritic cells through an interleukin-6-dependent mechanism. Stem Cells 2007, 25, 2025–2032. [Google Scholar]
- Cho, P.S.; Messina, D.J.; Hirsh, E.L.; Chi, N.; Goldman, S.N.; Lo, D.P.; Harris, I.R.; Popma, S.H.; Sachs, D.H.; Huang, C.A. Immunogenicity of umbilical cord tissue derived cells. Blood 2008, 111, 430–438. [Google Scholar]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress t-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar]
- Najar, M.; Rouas, R.; Raicevic, G.; Boufker, H.I.; Lewalle, P.; Meuleman, N.; Bron, D.; Toungouz, M.; Martiat, P.; Lagneaux, L. Mesenchymal stromal cells promote or suppress the proliferation of t lymphocytes from cord blood and peripheral blood: The importance of low cell ratio and role of interleukin-6. Cytotherapy 2009, 11, 570–583. [Google Scholar]
- Gotherstrom, C.; Ringden, O.; Westgren, M.; Tammik, C.; Le Blanc, K. Immunomodulatory effects of human foetal liver-derived mesenchymal stem cells. Bone Marrow Transplant 2003, 32, 265–272. [Google Scholar]
- Tipnis, S.; Viswanathan, C.; Majumdar, A.S. Immunosuppressive properties of human umbilical cord-derived mesenchymal stem cells: Role of b7-h1 and ido. Immunol. Cell Biol 2010, 88, 795–806. [Google Scholar]
- Conconi, M.T.; Di Liddo, R.; Tommasini, M.; Calore, C.; Parnigotto, P.P. Phenotype and differentiation potential of stromal populations obtained from various zones of human umbilical cord: An overview. Open Tissue Eng. Regen. Med. J 2011, 4, 6–20. [Google Scholar]
- Rey Nores, J.E.; Bensussan, A.; Vita, N.; Stelter, F.; Arias, M.A.; Jones, M.; Lefort, S.; Borysiewicz, L.K.; Ferrara, P.; Labeta, M.O. Soluble cd14 acts as a negative regulator of human t cell activation and function. Eur. J. Immunol 1999, 29, 265–276. [Google Scholar]
- Conconi, M.T.; Burra, P.; Di Liddo, R.; Calore, C.; Turetta, M.; Bellini, S.; Bo, P.; Nussdorfer, G.G.; Parnigotto, P.P. Cd105(+) cells from wharton’s jelly show in vitro and in vivo myogenic differentiative potential. Int. J. Mol. Med 2006, 18, 1089–1096. [Google Scholar]
- Dabelea, D.; Pettitt, D.J. Intrauterine diabetic environment confers risks for type 2 diabetes mellitus and obesity in the offspring, in addition to genetic susceptibility. J. Pediatr. Endocrinol. Metab 2001, 14, 1085–1091. [Google Scholar]
- Gillman, M.W.; Rifas-Shiman, S.; Berkey, C.S.; Field, A.E.; Colditz, G.A. Maternal gestational diabetes, birth weight, and adolescent obesity. Pediatrics 2003, 111, e221–e226. [Google Scholar]
- Clausen, T.D.; Mathiesen, E.R.; Hansen, T.; Pedersen, O.; Jensen, D.M.; Lauenborg, J.; Damm, P. High prevalence of type 2 diabetes and pre-diabetes in adult offspring of women with gestational diabetes mellitus or type 1 diabetes: The role of intrauterine hyperglycemia. Diabetes Care 2008, 31, 340–346. [Google Scholar]
- Tang, W.; Zeve, D.; Suh, J.M.; Bosnakovski, D.; Kyba, M.; Hammer, R.E.; Tallquist, M.D.; Graff, J.M. White fat progenitor cells reside in the adipose vasculature. Science 2008, 322, 583–586. [Google Scholar]
- Tchoukalova, Y.; Koutsari, C.; Jensen, M. Committed subcutaneous preadipocytes are reduced in human obesity. Diabetologia 2007, 50, 151–157. [Google Scholar]
- Pierdomenico, L.; Lanuti, P.; Lachmann, R.; Grifone, G.; Cianci, E.; Gialò, L.; Pacella, S.; Romano, M.; Vitacolonna, E.; Miscia, S. Diabetes mellitus during pregnancy interferes with the biological characteristics of wharton’s jelly mesenchymal stem cells. Open Tissue Eng. Regen. Med. J 2011, 4, 103–111. [Google Scholar]
- Ayuzawa, R.; Doi, C.; Rachakatla, R.S.; Pyle, M.M.; Maurya, D.K.; Troyer, D.; Tamura, M. Naive human umbilical cord matrix derived stem cells significantly attenuate growth of human breast cancer cells in vitro and in vivo. Cancer Lett 2009, 280, 31–37. [Google Scholar]
- Ganta, C.; Chiyo, D.; Ayuzawa, R.; Rachakatla, R.; Pyle, M.; Andrews, G.; Weiss, M.; Tamura, M.; Troyer, D. Rat umbilical cord stem cells completely abolish rat mammary carcinomas with no evidence of metastasis or recurrence 100 days post-tumor cell inoculation. Cancer Res 2009, 69, 1815–1820. [Google Scholar]
- Tamura, M.; Kawabata, A.; Ohta, N.; Uppalapati, L.; Becker, K.G.; Troyer, D. Wharton’s jelly stem cells as agents for cancer therapy. Open Tissue Eng. Regen. Med. J 2011, 4, 39–47. [Google Scholar]
- Nakamizo, A.; Marini, F.; Amano, T.; Khan, A.; Studeny, M.; Gumin, J.; Chen, J.; Hentschel, S.; Vecil, G.; Dembinski, J.; et al. Human bone marrow-derived mesenchymal stem cells in the treatment of gliomas. Cancer Res 2005, 65, 3307–3318. [Google Scholar]
- Rachakatla, R.S.; Marini, F.; Weiss, M.L.; Tamura, M.; Troyer, D. Development of human umbilical cord matrix stem cell-based gene therapy for experimental lung tumors. Cancer Gene Ther 2007, 14, 828–835. [Google Scholar]
- Matsuzuka, T.; Rachakatla, R.S.; Doi, C.; Maurya, D.K.; Ohta, N.; Kawabata, A.; Pyle, M.M.; Pickel, L.; Reischman, J.; Marini, F.; et al. Human umbilical cord matrix-derived stem cells expressing interferon-beta gene significantly attenuate bronchioloalveolar carcinoma xenografts in scid mice. Lung Cancer 2010, 70, 28–36. [Google Scholar]
- Scheers, I.; Lombard, C.; Najimi, M.; Sokal, E. Cell therapy for the treatment of metabolic liver disease: An update on the umbilical cord derived stem cells candidates. Open Tissue Eng. Regen. Med. J 2011, 4, 48–53. [Google Scholar]
- Campard, D.; Lysy, P.A.; Najimi, M.; Sokal, E.M. Native umbilical cord matrix stem cells express hepatic markers and differentiate into hepatocyte-like cells. Gastroenterology 2008, 134, 833–848. [Google Scholar]
- Anzalone, R.; Lo Iacono, M.; Corrao, S.; Magno, F.; Loria, T.; Cappello, F.; Zummo, G.; Farina, F.; La Rocca, G. New emerging potentials for human wharton’s jelly mesenchymal stem cells: Immunological features and hepatocyte-like differentiative capacity. Stem Cells Dev 2010, 19, 423–438. [Google Scholar]
- Semenov, O.; Breymann, C. Mesenchymal stem cells derived from wharton’s jelly and their potential for cardio-vascular tissue engineering. Open Tissue Eng. Regen. Med. J 2011, 4, 64–71. [Google Scholar]
- Mayer, J.E., Jr. Uses of homograft conduits for right ventricle to pulmonary artery connections in the neonatal period. Semin. Thorac. Cardiovasc. Surg. 1995, 7, 130–132. [Google Scholar]
- Schoen, F.J.; Levy, R.J. Tissue heart valves: Current challenges and future research perspectives. J. Biomed. Mater. Res 1999, 47, 439–465. [Google Scholar]
- Shinoka, T.; Ma, P.X.; Shum-Tim, D.; Breuer, C.K.; Cusick, R.A.; Zund, G.; Langer, R.; Vacanti, J.P.; Mayer, J.E., Jr. Tissue-engineered heart valves. Autologous valve leaflet replacement study in a lamb model. Circulation 1996, 94, II164–II168. [Google Scholar]
- Kenar, H.; Kose, G.T.; Toner, M.; Kaplan, D.L.; Hasirci, V. A 3d aligned microfibrous myocardial tissue construct cultured under transient perfusion. Biomaterials 2011, 32, 5320–5329. [Google Scholar]
- Lo Iacono, M.; Anzalone, R.; Corrao, S.; Giuffrè, M.; Di Stefano, A.; Giannuzzi, P.; Cappello, F.; Farina, F.; La Rocca, G. Perinatal and wharton’s jelly-derived mesenchymal stem cells in cartilage regenerative medicine and tissue engineering strategies. Open Tissue Eng. Regen. Med. J 2011, 4, 72–81. [Google Scholar]
- Arufe, M.C.; De la Fuente, A.; Mateos, J.; Fuentes, I.; de Toro, F.J.; Blanco, F.J. Analysis of the chondrogenic potential and secretome of mesenchymal stem cells derived from human umbilical cord stroma. Stem Cells Dev 2011, 20, 1199–1212. [Google Scholar]
- Wang, L.; Seshareddy, K.; Weiss, M.L.; Detamore, M.S. Effect of initial seeding density on human umbilical cord mesenchymal stromal cells for fibrocartilage tissue engineering. Tissue Eng. A 2009, 15, 1009–1017. [Google Scholar]
- Fong, C.Y.; Subramanian, A.; Gauthaman, K.; Venugopal, J.; Biswas, A.; Ramakrishna, S.; Bongso, A. Human umbilical cord wharton’s jelly stem cells undergo enhanced chondrogenic differentiation when grown on nanofibrous scaffolds and in a sequential two-stage culture medium environment. Stem Cell Rev 2012, 8, 195–209. [Google Scholar]
- Wang, L.; Zhao, L.; Detamore, M.S. Human umbilical cord mesenchymal stromal cells in a sandwich approach for osteochondral tissue engineering. J. Tissue Eng. Regen. Med 2011, 5, 712–721. [Google Scholar]
- Ishikawa, N.; Suzuki, Y.; Ohta, M.; Cho, H.; Suzuki, S.; Dezawa, M.; Ide, C. Peripheral nerve regeneration through the space formed by a chitosan gel sponge. J. Biomed. Mater. Res. Part A 2007, 83, 33–40. [Google Scholar]
- Ohta, M.; Suzuki, Y.; Chou, H.; Ishikawa, N.; Suzuki, S.; Tanihara, M.; Suzuki, Y.; Mizushima, Y.; Dezawa, M.; Ide, C. Novel heparin/alginate gel combined with basic fibroblast growth factor promotes nerve regeneration in rat sciatic nerve. J. Biomed. Mater. Res. Part A 2004, 71, 661–668. [Google Scholar]
- Hall, S. Nerve repair: A neurobiologist’s view. J. Hand Surg. 2001, 26, 129–136. [Google Scholar]
- Dezawa, M.; Takahashi, I.; Esaki, M.; Takano, M.; Sawada, H. Sciatic nerve regeneration in rats induced by transplantation of in vitro differentiated bone-marrow stromal cells. Eur. J. Neurosci 2001, 14, 1771–1776. [Google Scholar]
- Matsuse, D.; Kitada, M.; Kohama, M.; Nishikawa, K.; Makinoshima, H.; Wakao, S.; Fujiyoshi, Y.; Heike, T.; Nakahata, T.; Akutsu, H.; et al. Human umbilical cord-derived mesenchymal stromal cells differentiate into functional schwann cells that sustain peripheral nerve regeneration. J. Neuropathol. Exp. Neurol 2010, 69, 973–985. [Google Scholar]
- Kuroda, Y.; Kitada, M.; Wakao, S.; Dezawa, M. Mesenchymal stem cells and umbilical cord as sources for schwann cell differentiation: Their potential in peripheral nerve repair. Open Tissue Eng. Regen. Med. J 2011, 4, 54–63. [Google Scholar]
- Peng, J.; Wang, Y.; Zhang, L.; Zhao, B.; Zhao, Z.; Chen, J.; Guo, Q.; Liu, S.; Sui, X.; Xu, W.; et al. Human umbilical cord wharton’s jelly-derived mesenchymal stem cells differentiate into a schwann-cell phenotype and promote neurite outgrowth in vitro. Brain res. Bull 2011, 84, 235–243. [Google Scholar]
- Xu, Q.; Zhang, H.T.; Liu, K.; Rao, J.H.; Liu, X.M.; Wu, L.; Xu, B.N. In vitro and in vivo magnetic resonance tracking of sinerem-labeled human umbilical mesenchymal stromal cell-derived schwann cells. Cell. Mol. Neurobiol 2011, 31, 365–375. [Google Scholar]
- López, Y.; Seshareddy, K.; Trevino, E.; Cox, J.; Weiss, M.L. Evaluating the impact of oxygen concentration and plating density on human wharton’s jelly-derived mesenchymal stromal cells. Open Tissue Eng. Regen. Med. J 2011, 4, 82–94. [Google Scholar]
- Breitbach, M.; Bostani, T.; Roell, W.; Xia, Y.; Dewald, O.; Nygren, J.M.; Fries, J.W.; Tiemann, K.; Bohlen, H.; Hescheler, J.; et al. Potential risks of bone marrow cell transplantation into infarcted hearts. Blood 2007, 110, 1362–1369. [Google Scholar]
- Bittira, B.; Kuang, J.Q.; Al-Khaldi, A.; Shum-Tim, D.; Chiu, R.C. In vitro preprogramming of marrow stromal cells for myocardial regeneration. Ann. Thorac. Surg 2002, 74, 1154–1159, ; discussion 1159–1160.. [Google Scholar]
- Tomita, S.; Mickle, D.A.; Weisel, R.D.; Jia, Z.Q.; Tumiati, L.C.; Allidina, Y.; Liu, P.; Li, R.K. Improved heart function with myogenesis and angiogenesis after autologous porcine bone marrow stromal cell transplantation. J. Thorac. Cardiovasc. Surg 2002, 123, 1132–1140. [Google Scholar]
- Wang, H.S.; Hung, S.C.; Peng, S.T.; Huang, C.C.; Wei, H.M.; Guo, Y.J.; Fu, Y.S.; Lai, M.C.; Chen, C.C. Mesenchymal stem cells in the wharton’s jelly of the human umbilical cord. Stem Cells 2004, 22, 1330–1337. [Google Scholar]
- Matsuura, K.; Nagai, T.; Nishigaki, N.; Oyama, T.; Nishi, J.; Wada, H.; Sano, M.; Toko, H.; Akazawa, H.; Sato, T.; et al. Adult cardiac sca-1-positive cells differentiate into beating cardiomyocytes. J. Biol. Chem 2004, 279, 11384–11391. [Google Scholar]
- Maltsev, V.A.; Rohwedel, J.; Hescheler, J.; Wobus, A.M. Embryonic stem cells differentiate in vitro into cardiomyocytes representing sinusnodal, atrial and ventricular cell types. Mechan. Dev 1993, 44, 41–50. [Google Scholar]
- Hollweck, T.; Hartmann, I.; Eblenkamp, M.; Wintermantel, E.; Reichart, B.; Überfuhr, P.; Eissner, G. Cardiac differentiation of human wharton’s jelly stem cells—Experimental comparison of protocols. Open Tissue Eng. Regen. Med. J 2011, 4, 95–102. [Google Scholar]
- Fathi, F.; Murasawa, S.; Hasegawa, S.; Asahara, T.; Kermani, A.J.; Mowla, S.J. Cardiac differentiation of p19cl6 cells by oxytocin. Int. J. Cardiol 2009, 134, 75–81. [Google Scholar]
- Lopez, Y.; Lutjemeier, B.; Seshareddy, K.; Trevino, E.M.; Hageman, K.S.; Musch, T.I.; Borgarelli, M.; Weiss, M.L. Wharton’s jelly or bone marrow mesenchymal stromal cells improve cardiac function following myocardial infarction for more than 32 weeks in a rat model: A preliminary report. Curr. Stem Cell Res. Ther 2013, 8, 46–59. [Google Scholar]
- Schugar, R.C.; Chirieleison, S.M.; Wescoe, K.E.; Schmidt, B.T.; Askew, Y.; Nance, J.J.; Evron, J.M.; Peault, B.; Deasy, B.M. High harvest yield, high expansion, and phenotype stability of cd146 mesenchymal stromal cells from whole primitive human umbilical cord tissue. J. Biomed. Biotechnol 2009, 2009, 789526. [Google Scholar]
- Farias, V.A.; Linares-Fernandez, J.L.; Penalver, J.L.; Paya Colmenero, J.A.; Ferron, G.O.; Duran, E.L.; Fernandez, R.M.; Olivares, E.G.; O’Valle, F.; Puertas, A.; et al. Human umbilical cord stromal stem cell express cd10 and exert contractile properties. Placenta 2011, 32, 86–95. [Google Scholar]
- Sarugaser, R.; Hanoun, L.; Keating, A.; Stanford, W.L.; Davies, J.E. Human mesenchymal stem cells self-renew and differentiate according to a deterministic hierarchy. PLoS One 2009, 4, e6498. [Google Scholar]
- Sarugaser, R.; Ennis, J.; Stanford, W.L.; Davies, J.E. Isolation, propagation, and characterization of human umbilical cord perivascular cells (hucpvcs). Methods Mol. Biol 2009, 482, 269–279. [Google Scholar]
- Lange-Consiglio, A.; Corradetti, B.; Rutigliano, L.; Cremonesi, F.; Bizzarro, D. In vitro studies of horse umbilical cord matrix-derived cells: From characterization to labeling for magnetic resonance imaging. Open Tissue Eng. Regen. Med. J 2011, 4, 120–133. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, D.-W.; Staples, M.; Shinozuka, K.; Pantcheva, P.; Kang, S.-D.; Borlongan, C.V. Wharton’s Jelly-Derived Mesenchymal Stem Cells: Phenotypic Characterization and Optimizing Their Therapeutic Potential for Clinical Applications. Int. J. Mol. Sci. 2013, 14, 11692-11712. https://doi.org/10.3390/ijms140611692
Kim D-W, Staples M, Shinozuka K, Pantcheva P, Kang S-D, Borlongan CV. Wharton’s Jelly-Derived Mesenchymal Stem Cells: Phenotypic Characterization and Optimizing Their Therapeutic Potential for Clinical Applications. International Journal of Molecular Sciences. 2013; 14(6):11692-11712. https://doi.org/10.3390/ijms140611692
Chicago/Turabian StyleKim, Dae-Won, Meaghan Staples, Kazutaka Shinozuka, Paolina Pantcheva, Sung-Don Kang, and Cesar V. Borlongan. 2013. "Wharton’s Jelly-Derived Mesenchymal Stem Cells: Phenotypic Characterization and Optimizing Their Therapeutic Potential for Clinical Applications" International Journal of Molecular Sciences 14, no. 6: 11692-11712. https://doi.org/10.3390/ijms140611692
APA StyleKim, D.-W., Staples, M., Shinozuka, K., Pantcheva, P., Kang, S.-D., & Borlongan, C. V. (2013). Wharton’s Jelly-Derived Mesenchymal Stem Cells: Phenotypic Characterization and Optimizing Their Therapeutic Potential for Clinical Applications. International Journal of Molecular Sciences, 14(6), 11692-11712. https://doi.org/10.3390/ijms140611692