Secretory Phospholipases A2 in Durum Wheat (Triticum durum Desf.): Gene Expression, Enzymatic Activity, and Relation to Drought Stress Adaptation


Abstract
:
1. Introduction
2. Results
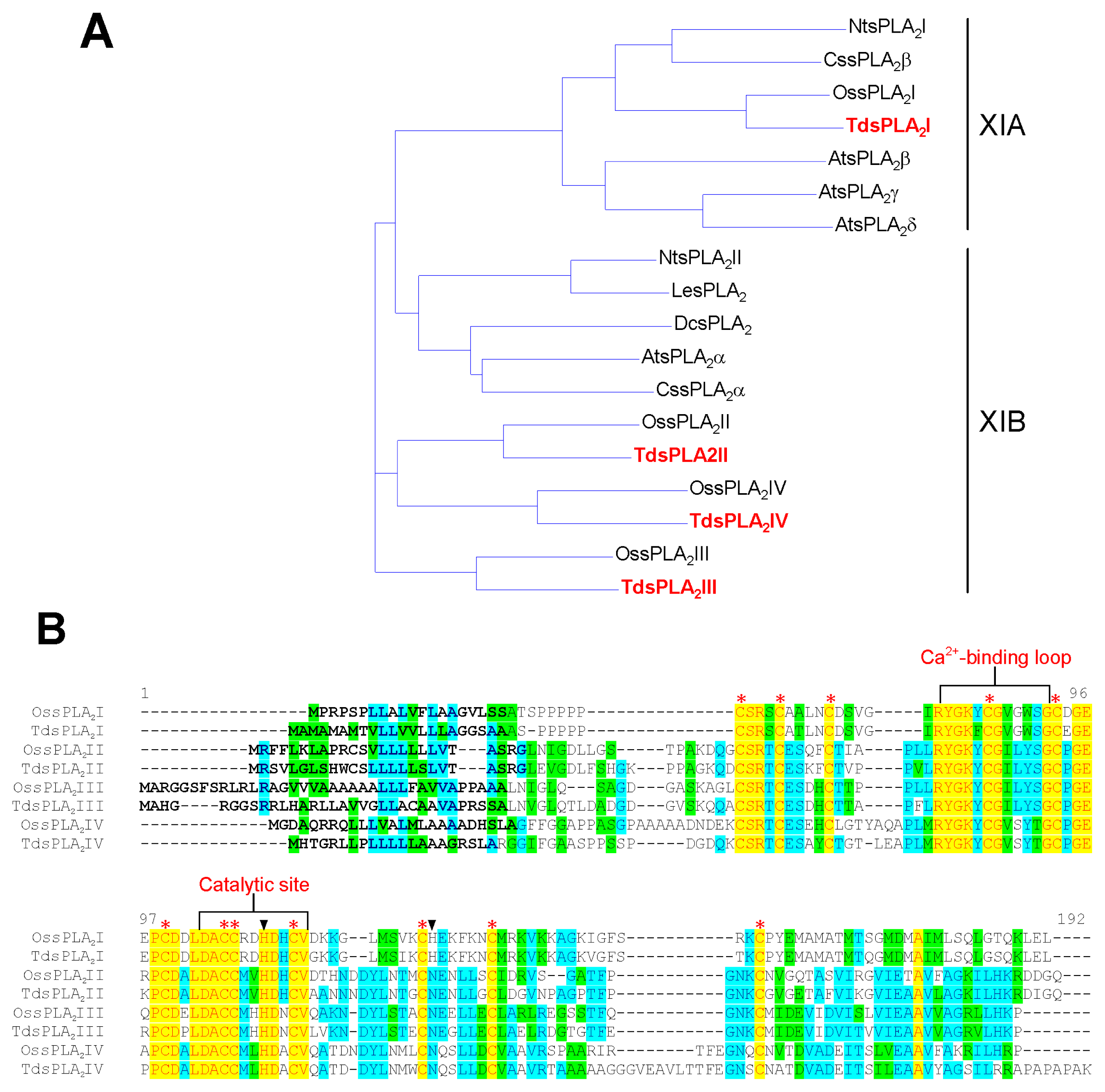
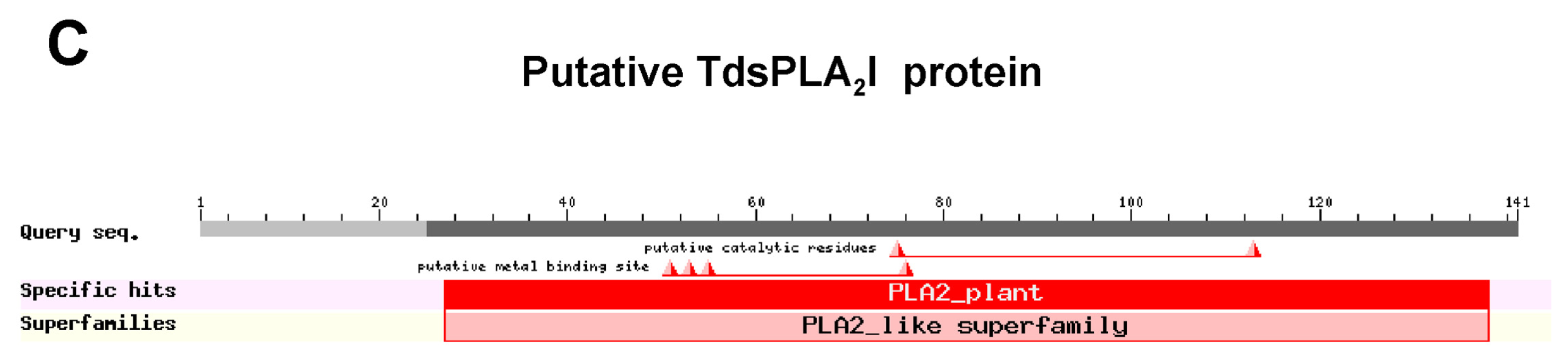
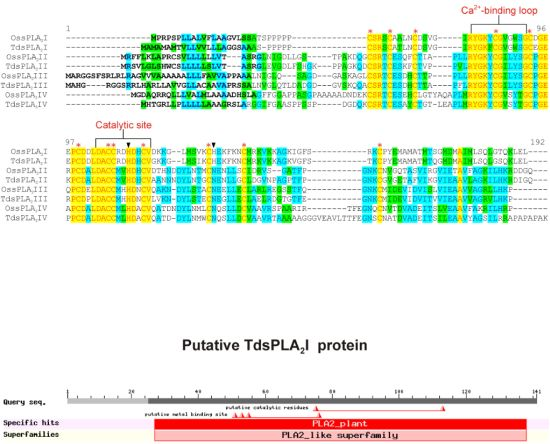
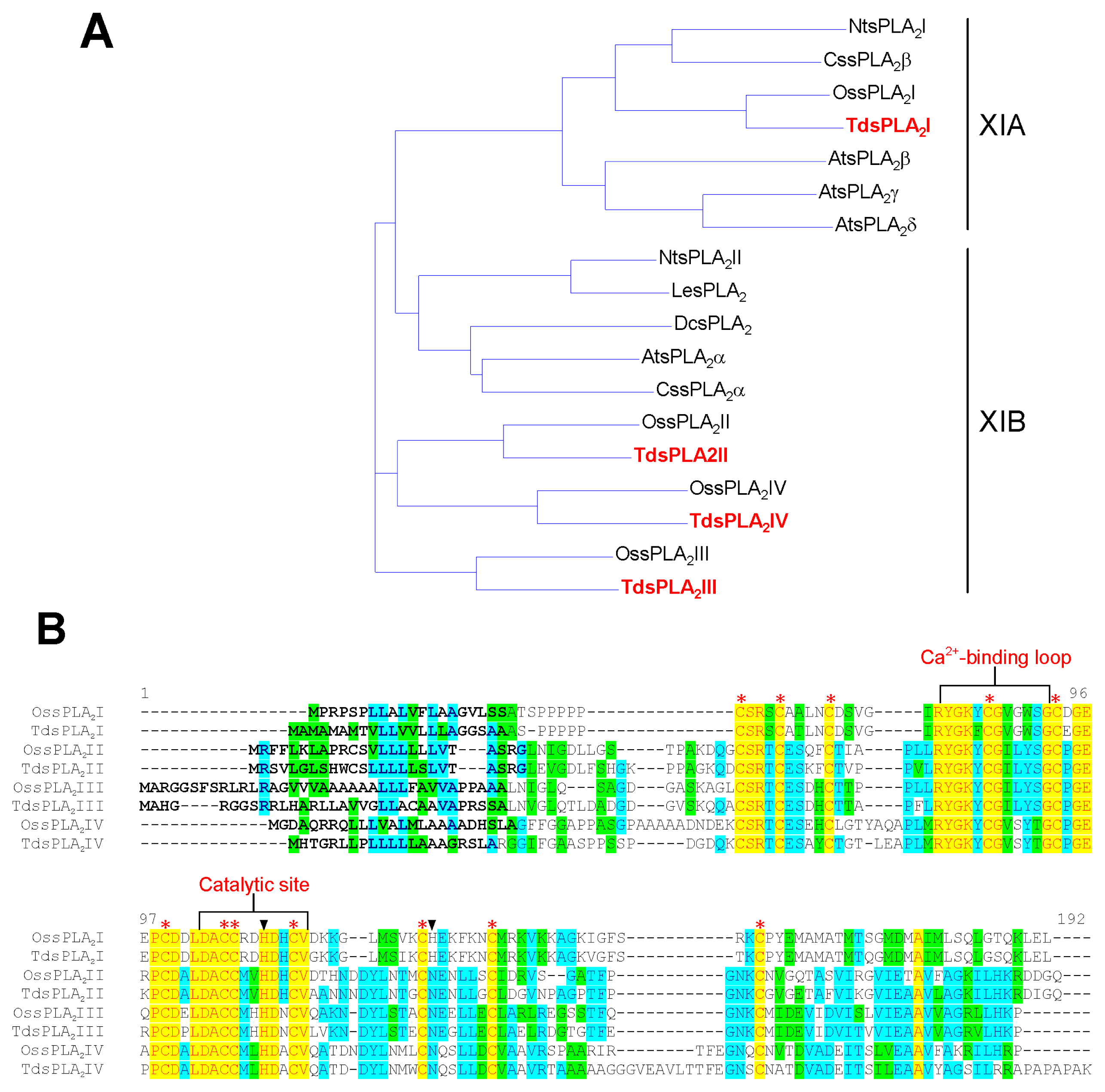
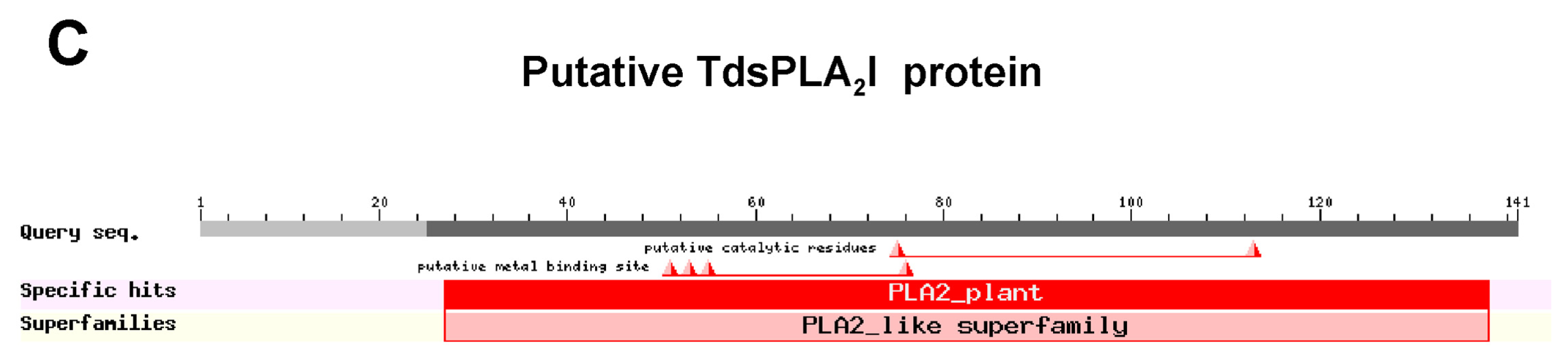
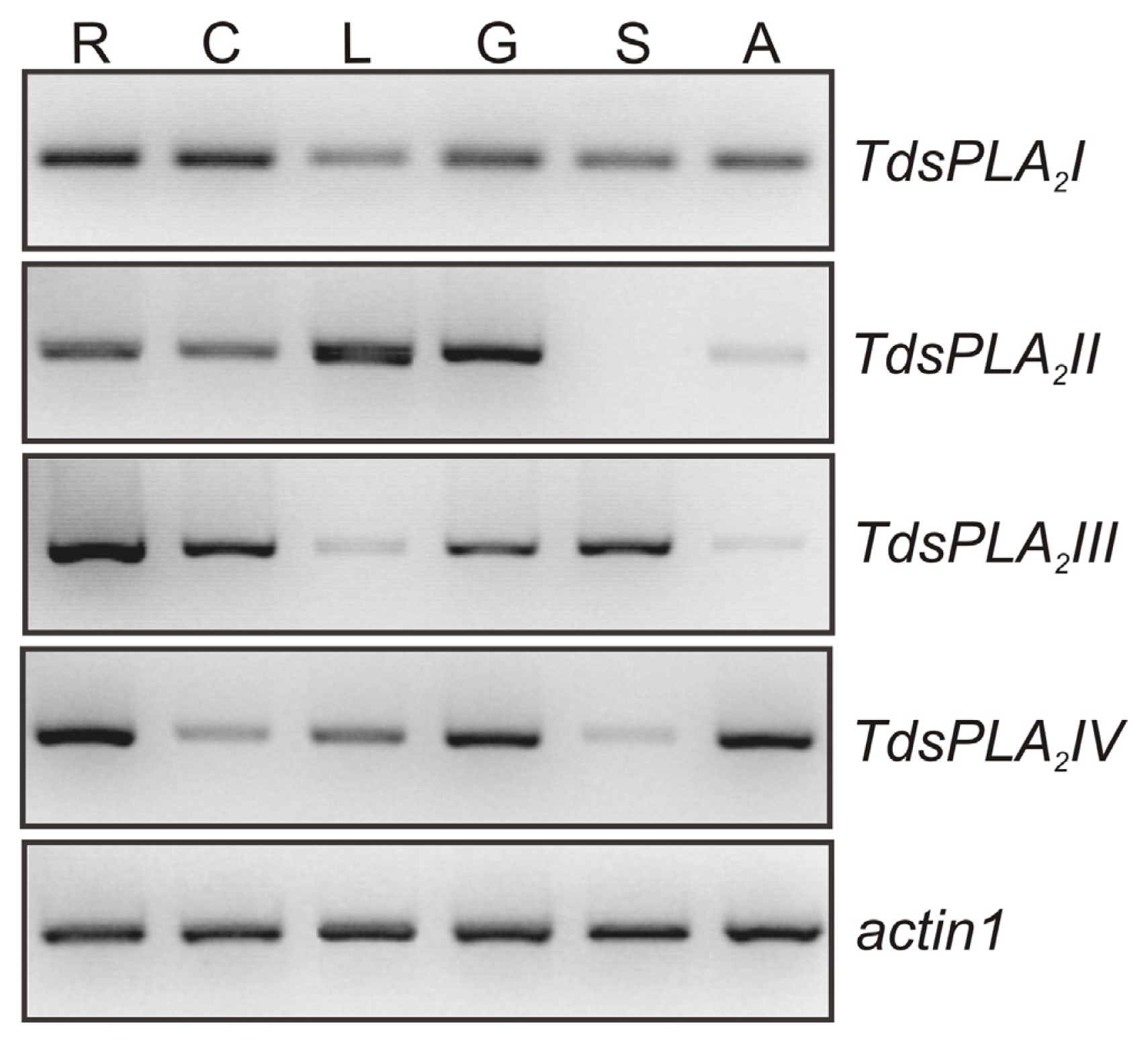
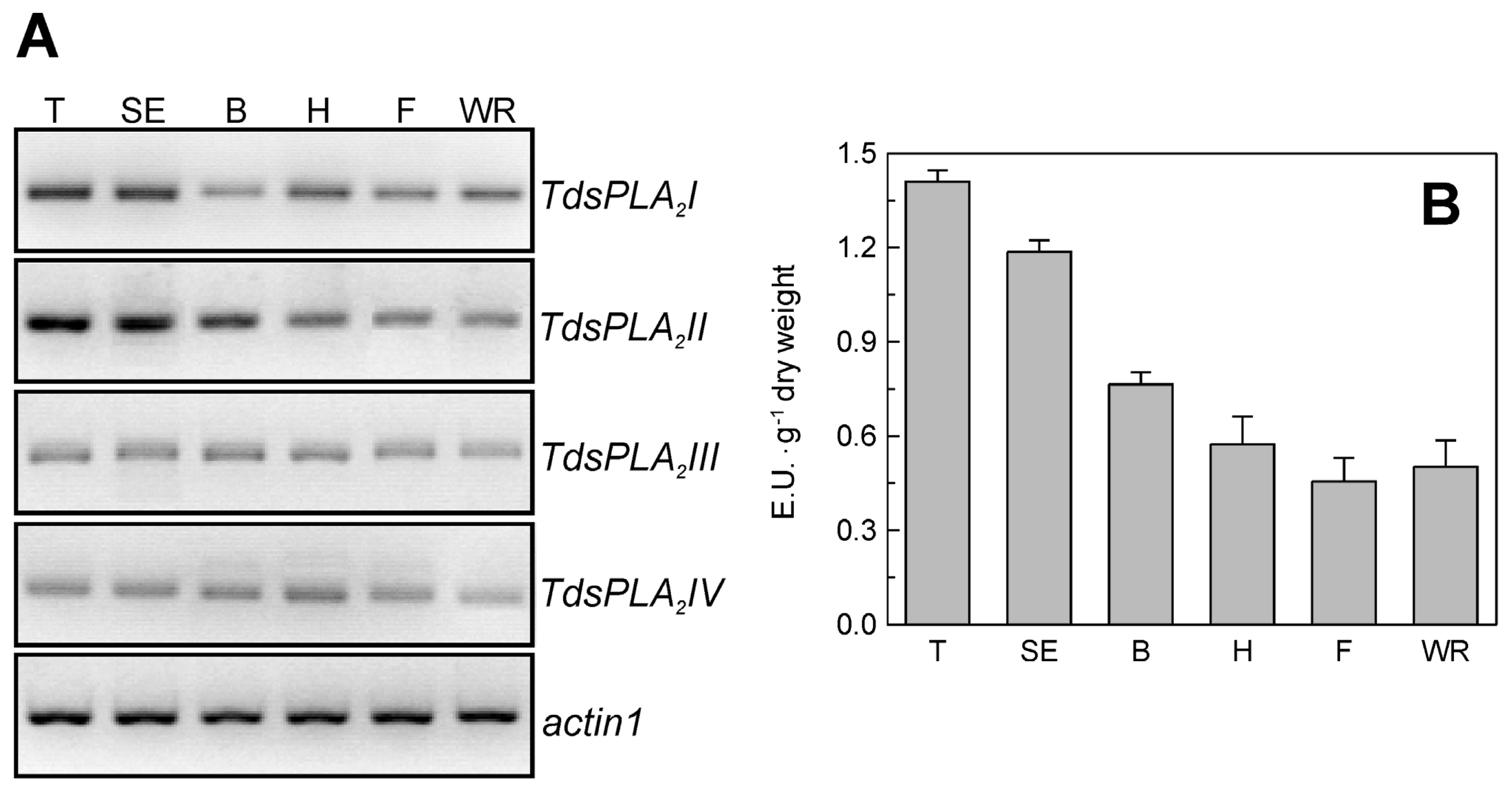
2.1. Isolation and Characterization of Durum Wheat Full-Length sPLA2 cDNAs and Gene Expression Analysis in Different Tissues
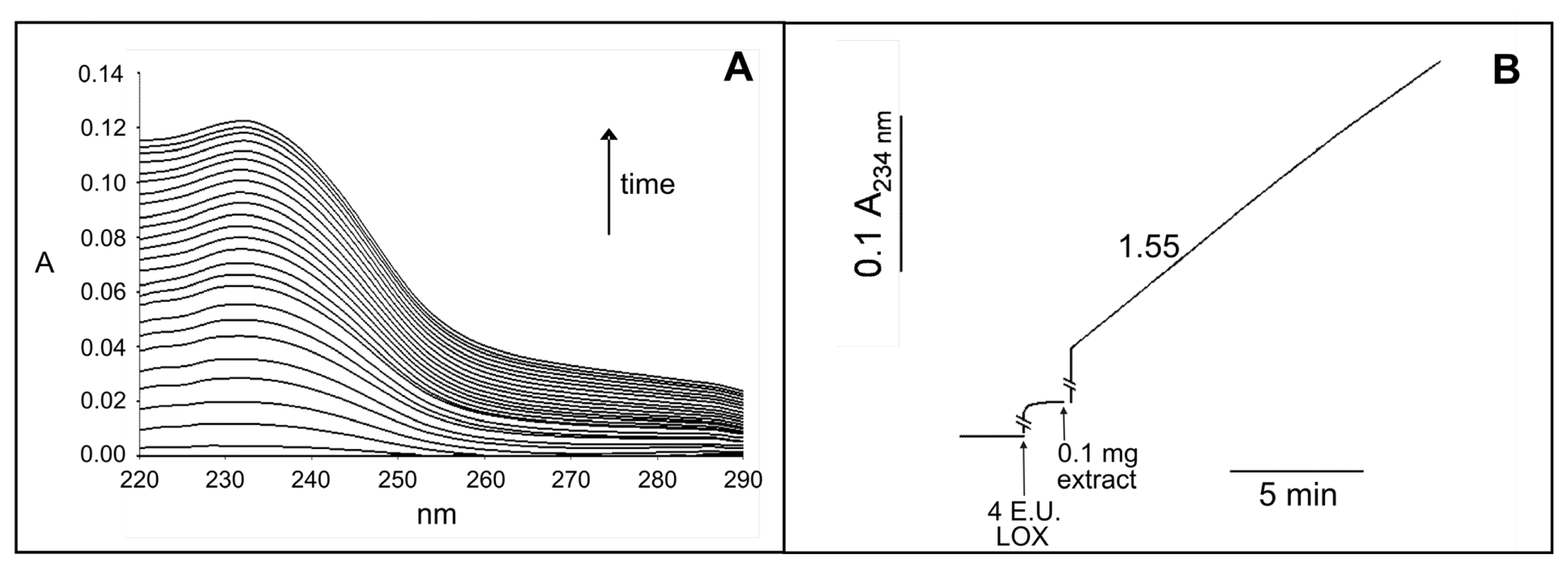
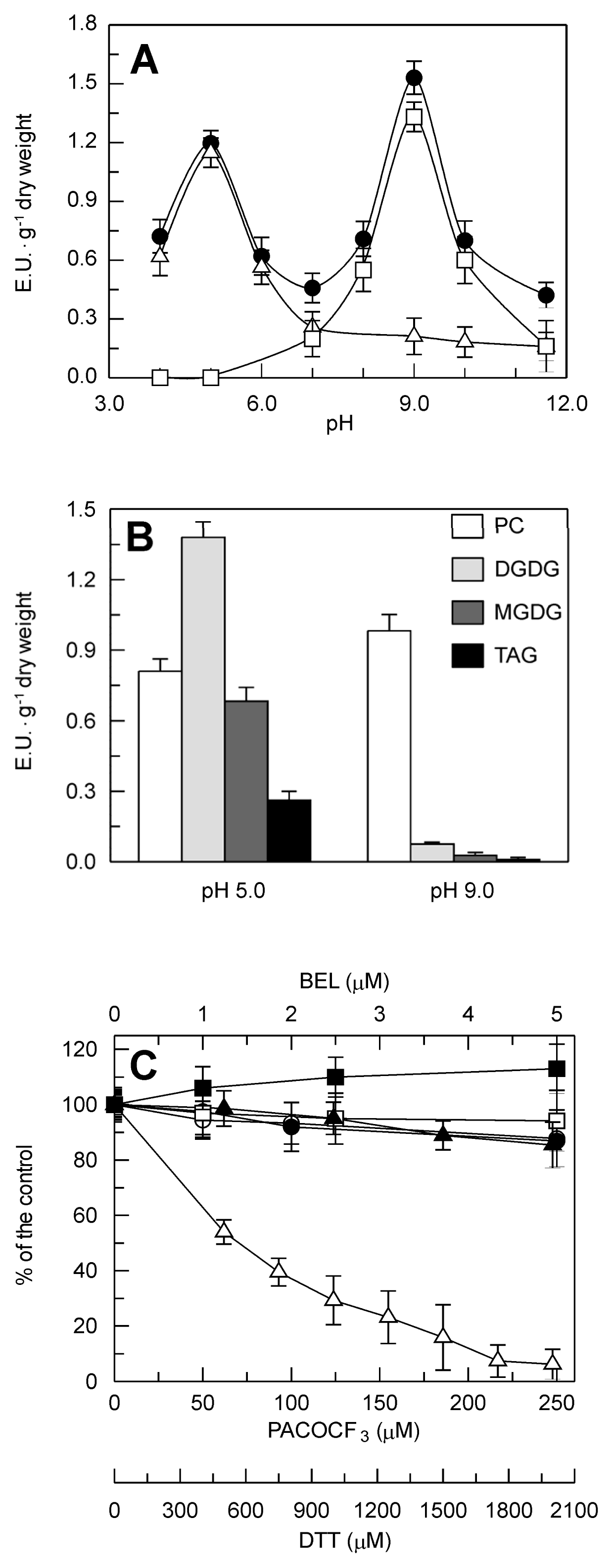
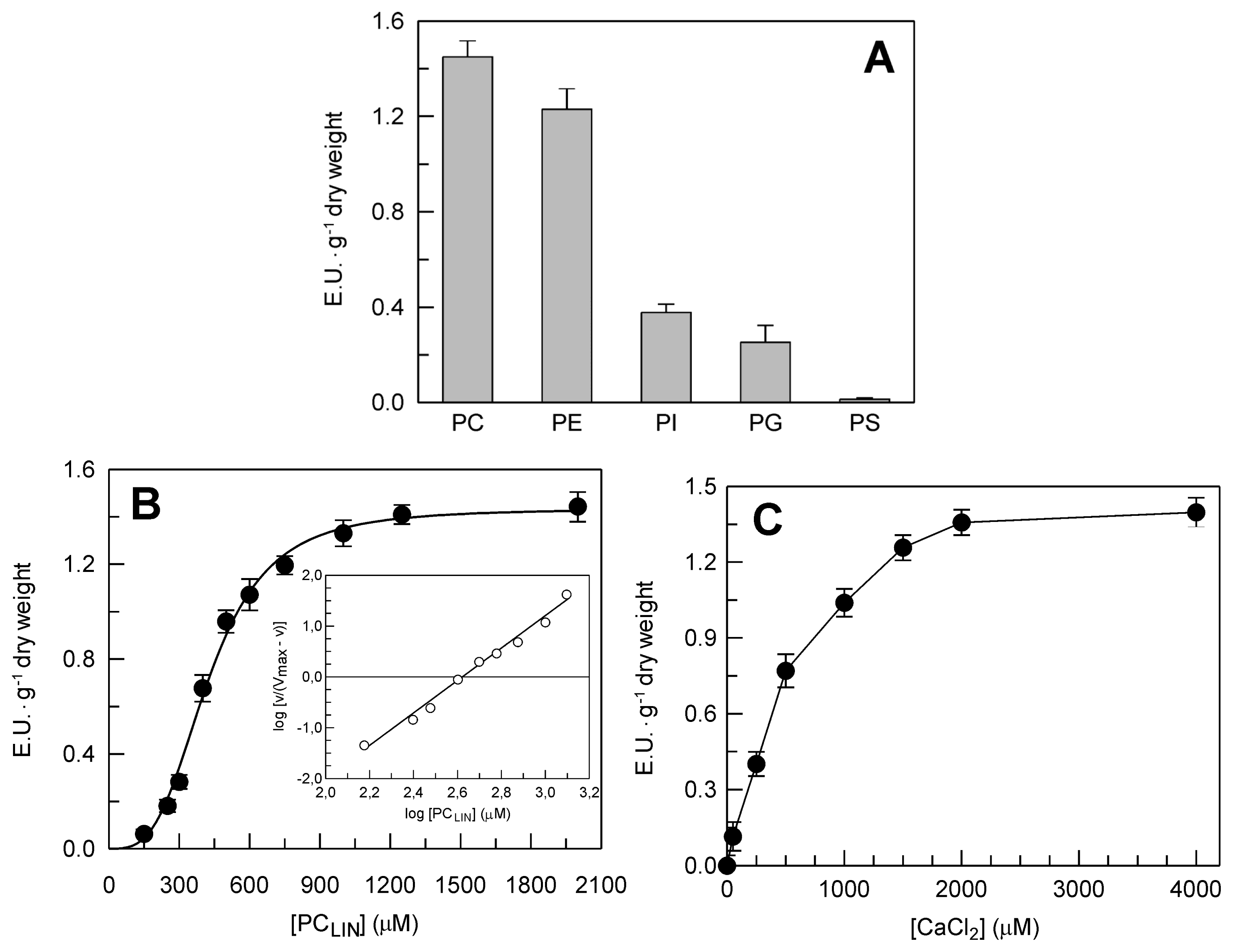
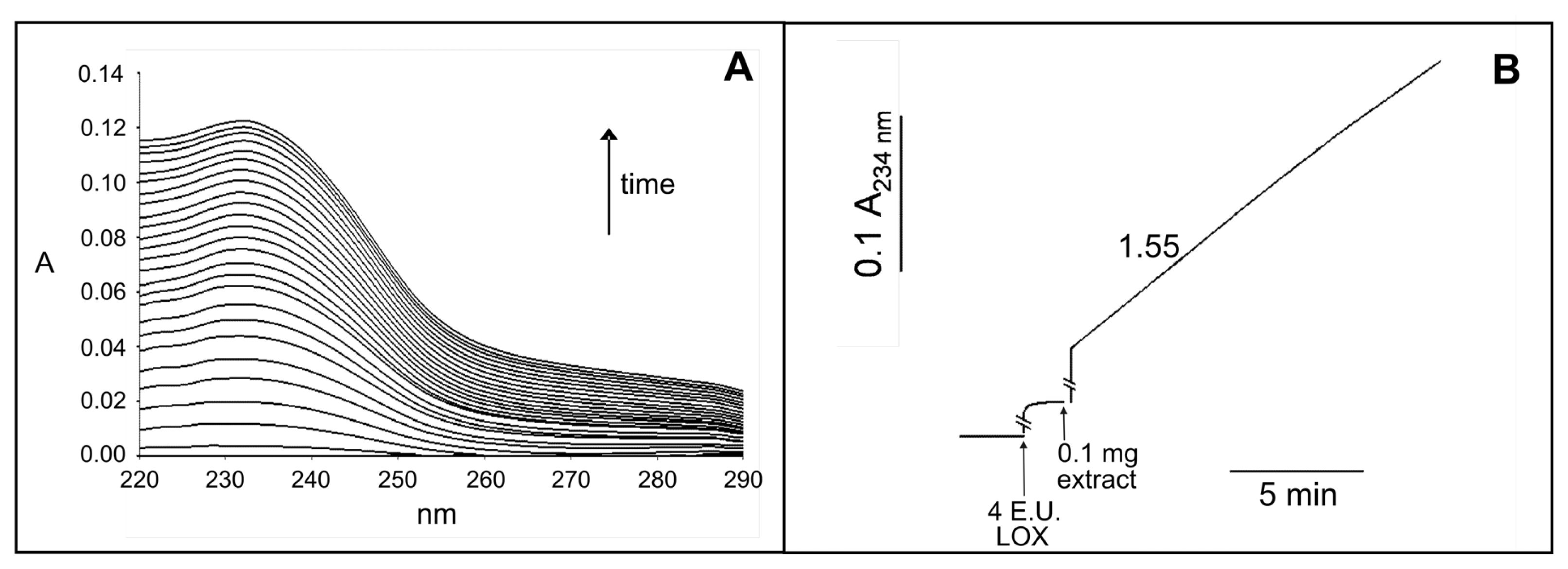
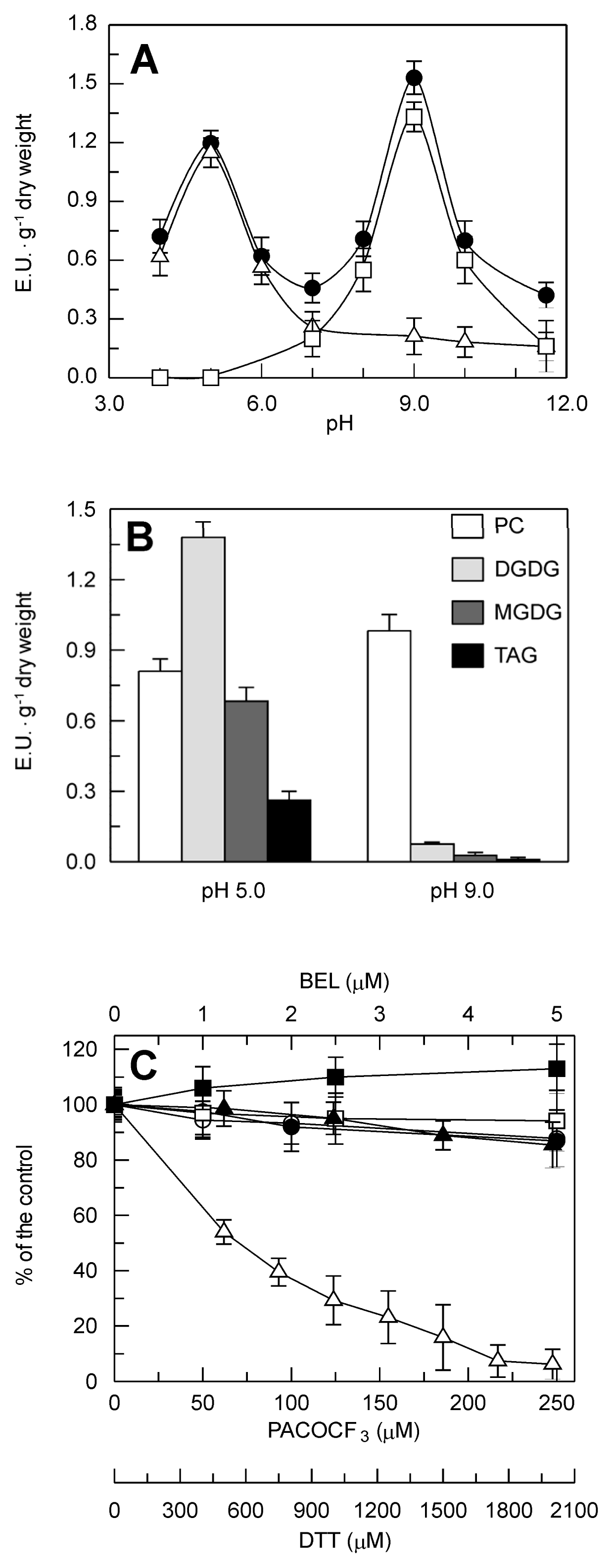
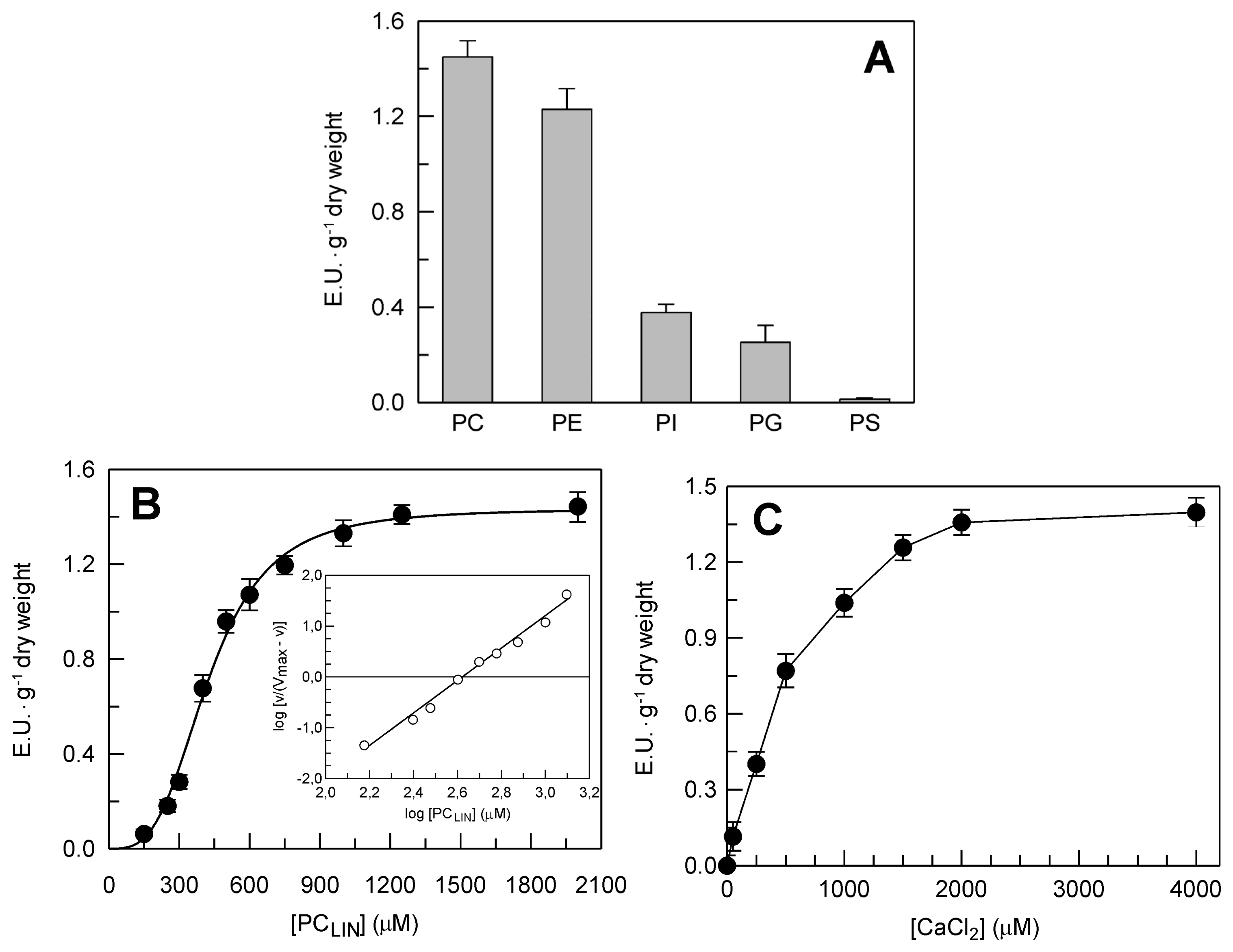
2.2. Identification and Biochemical Characterization of a Ca2+-Dependent PLA2 Activity in Durum Wheat Leaves
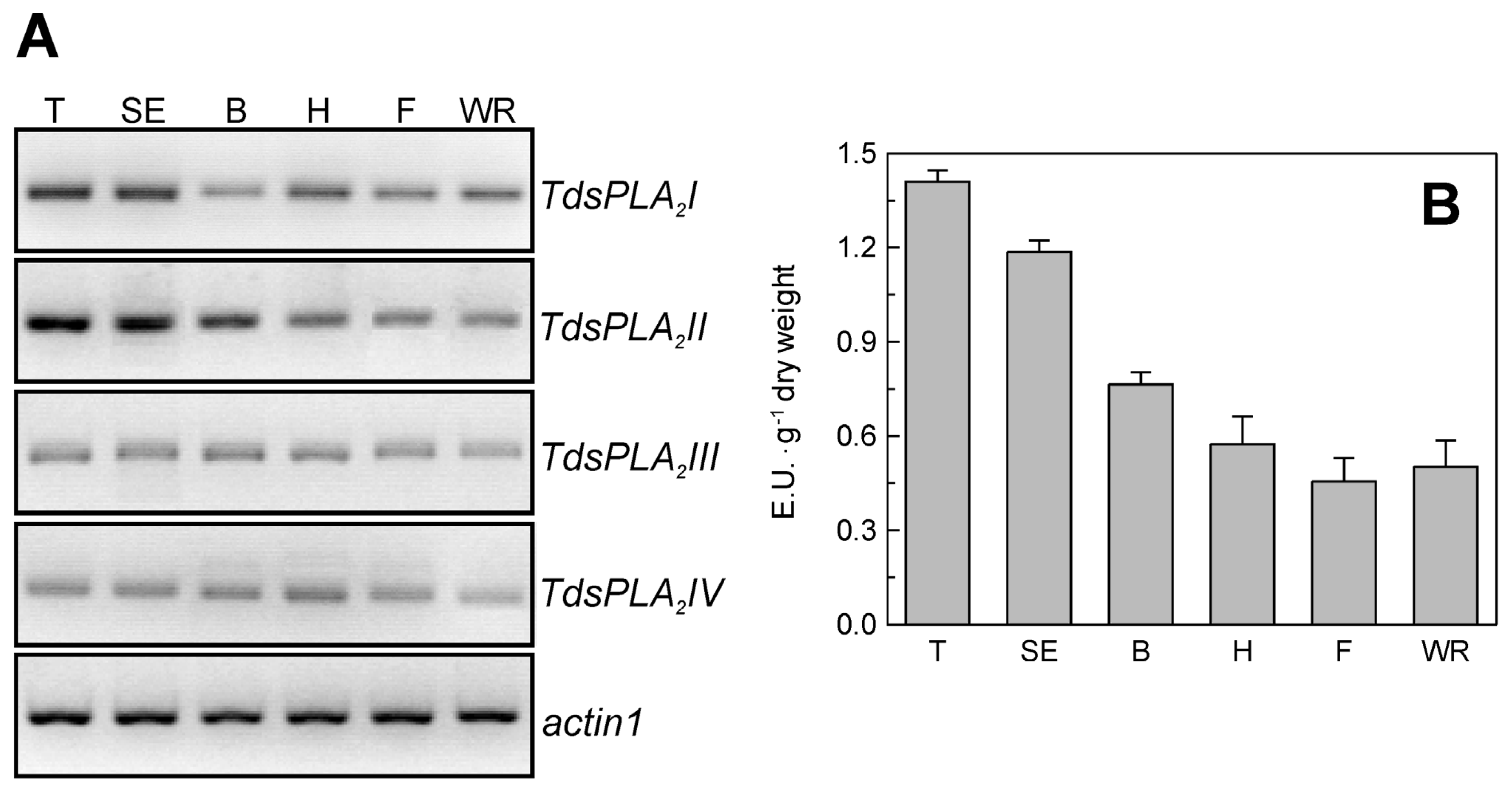
2.3. Evaluation of TdsPLA2 Gene Expression and DWL-PLA2 Activity in Durum Wheat Leaves at Different Developmental Stages
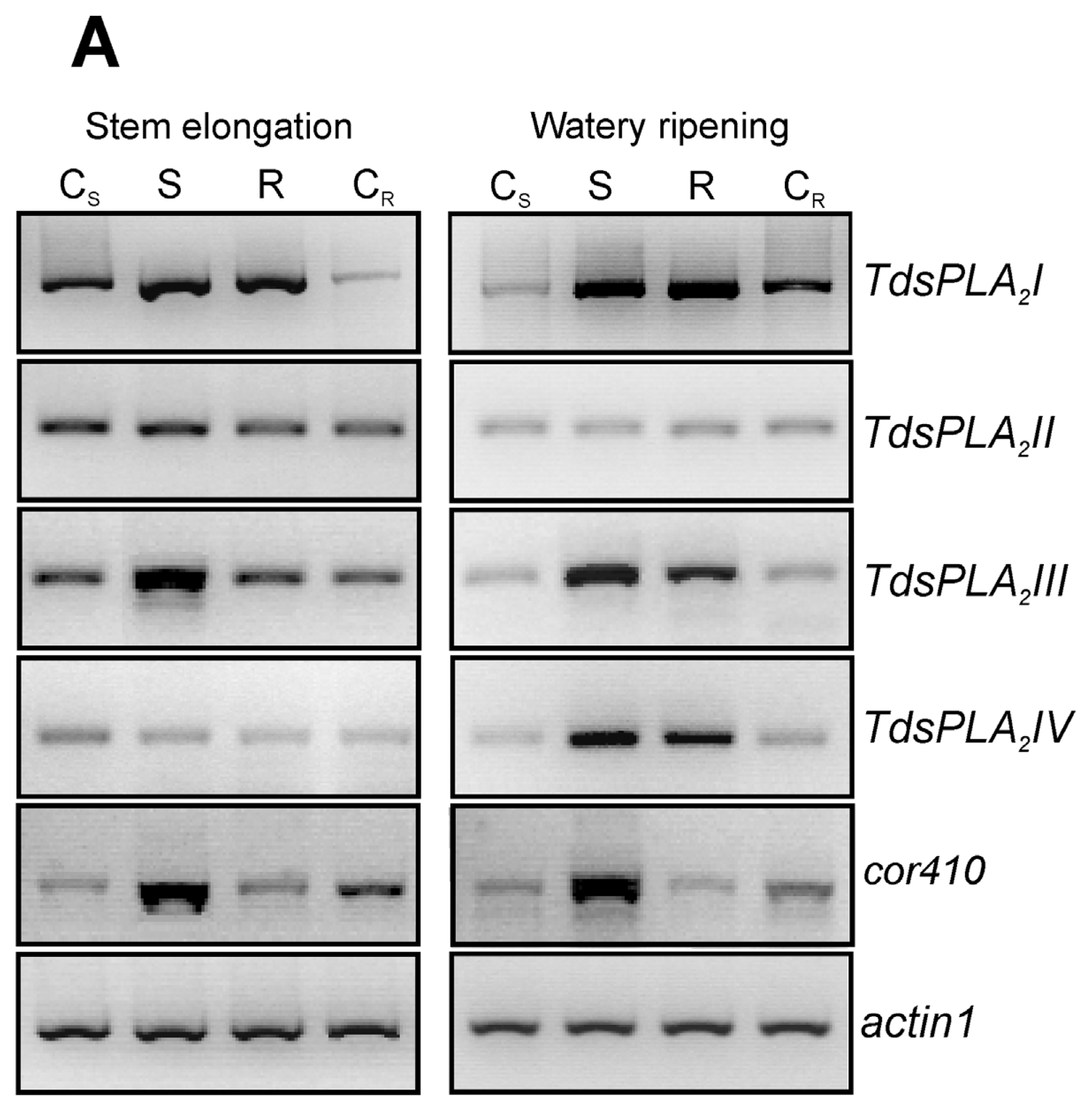
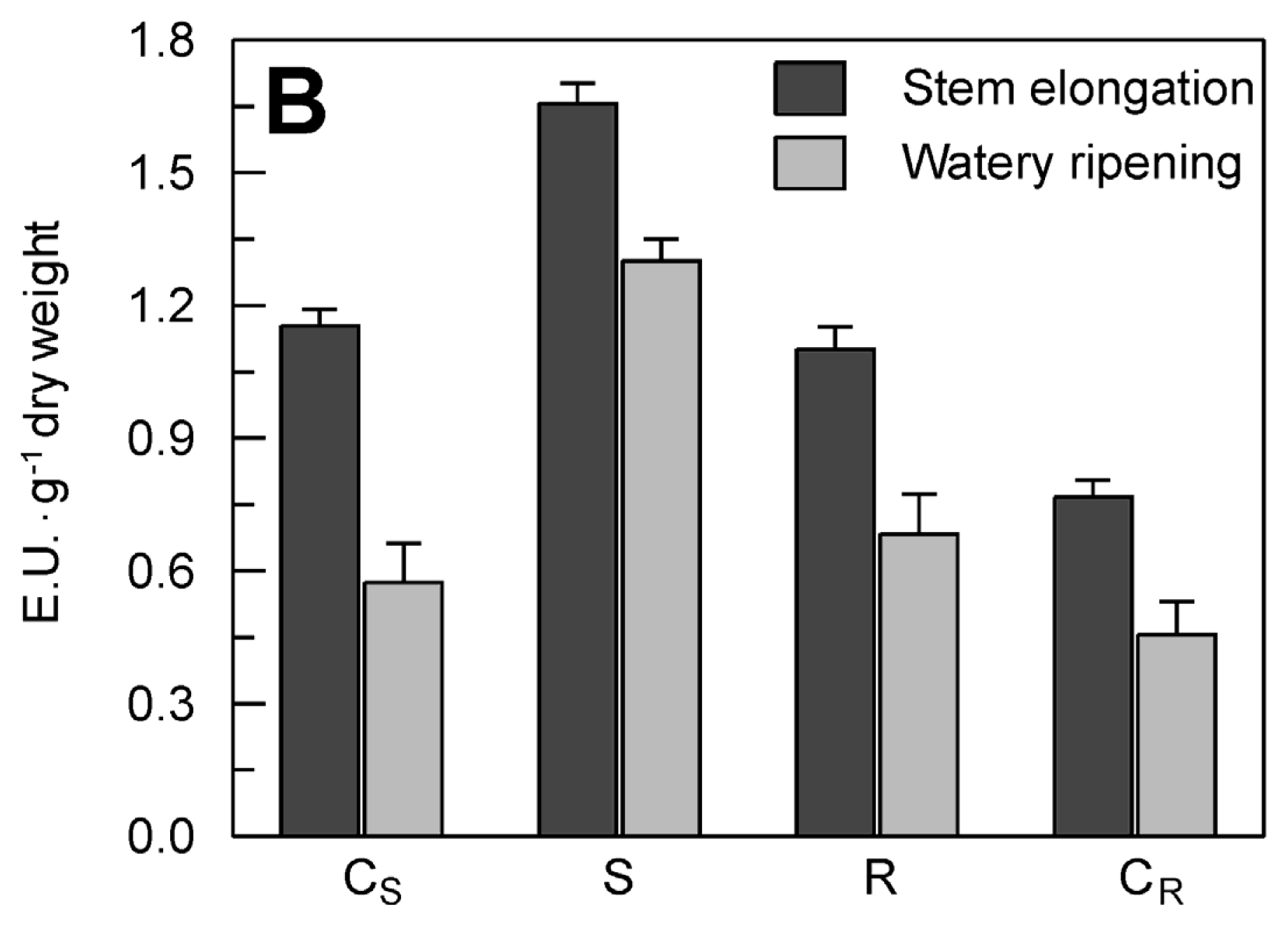
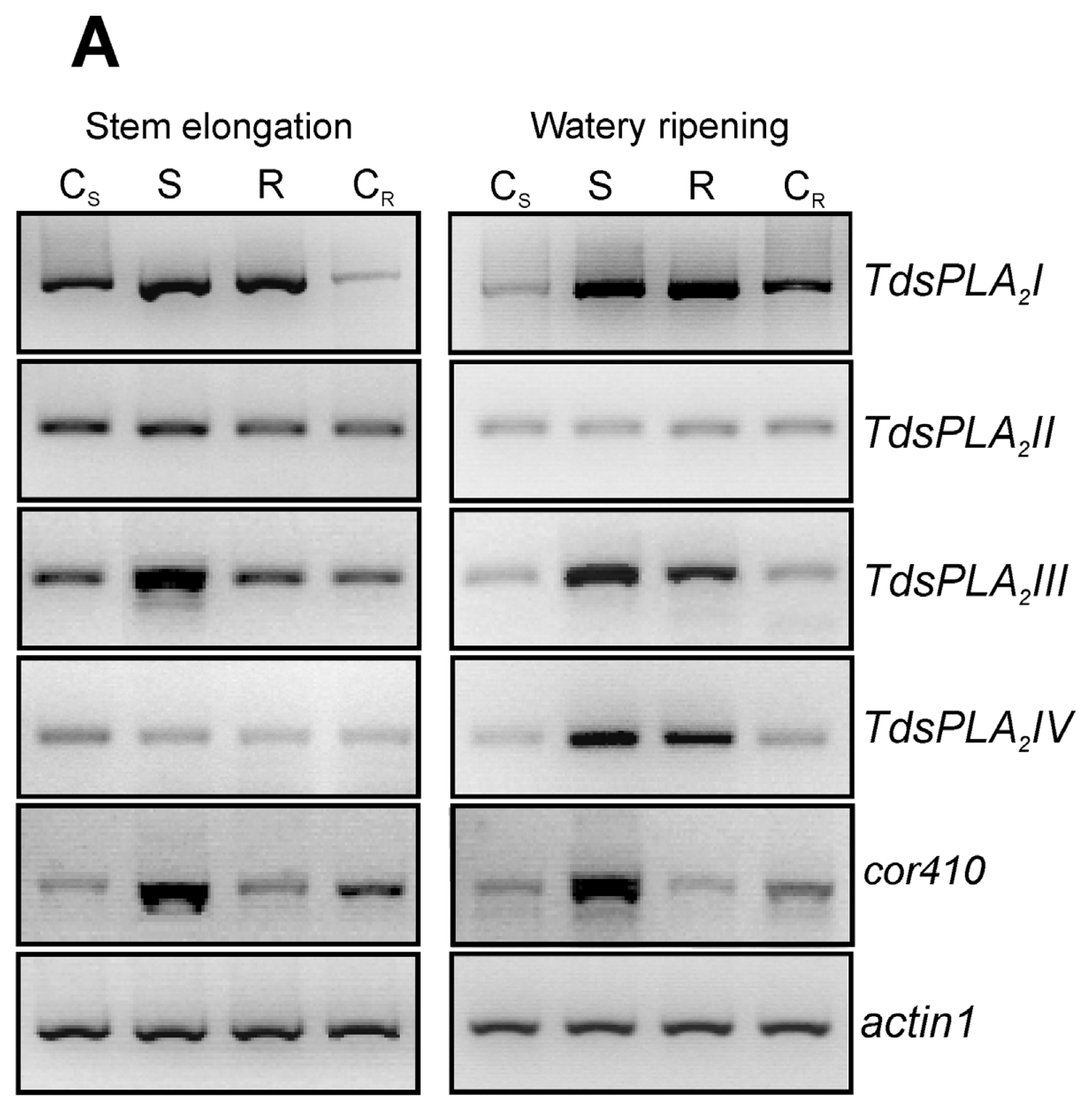
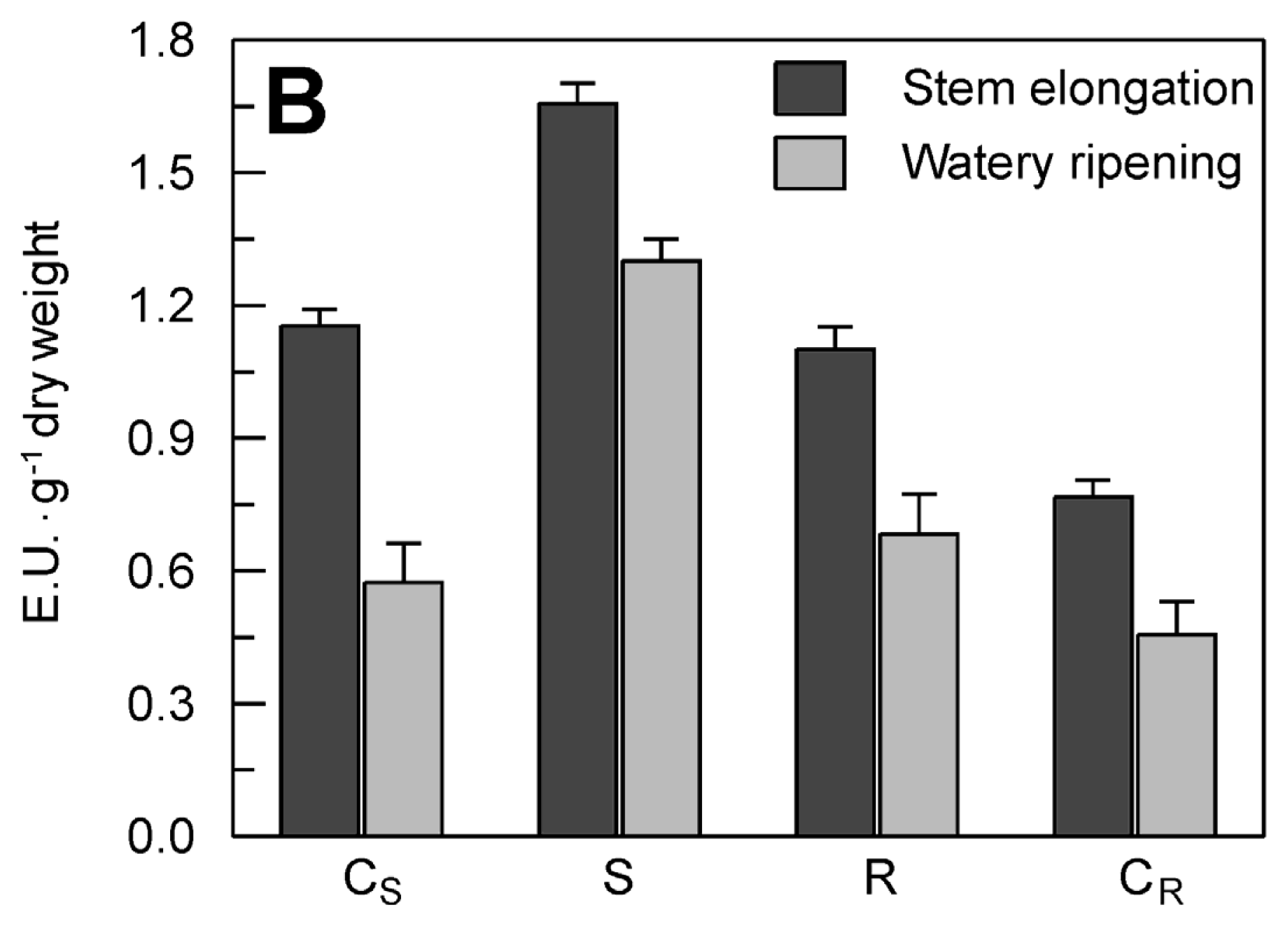
2.4. Evaluation of the Effect of the Drought Stress on TdsPLA2 Gene Expression and DWL-PLA2 Activity in Durum Wheat Leaves
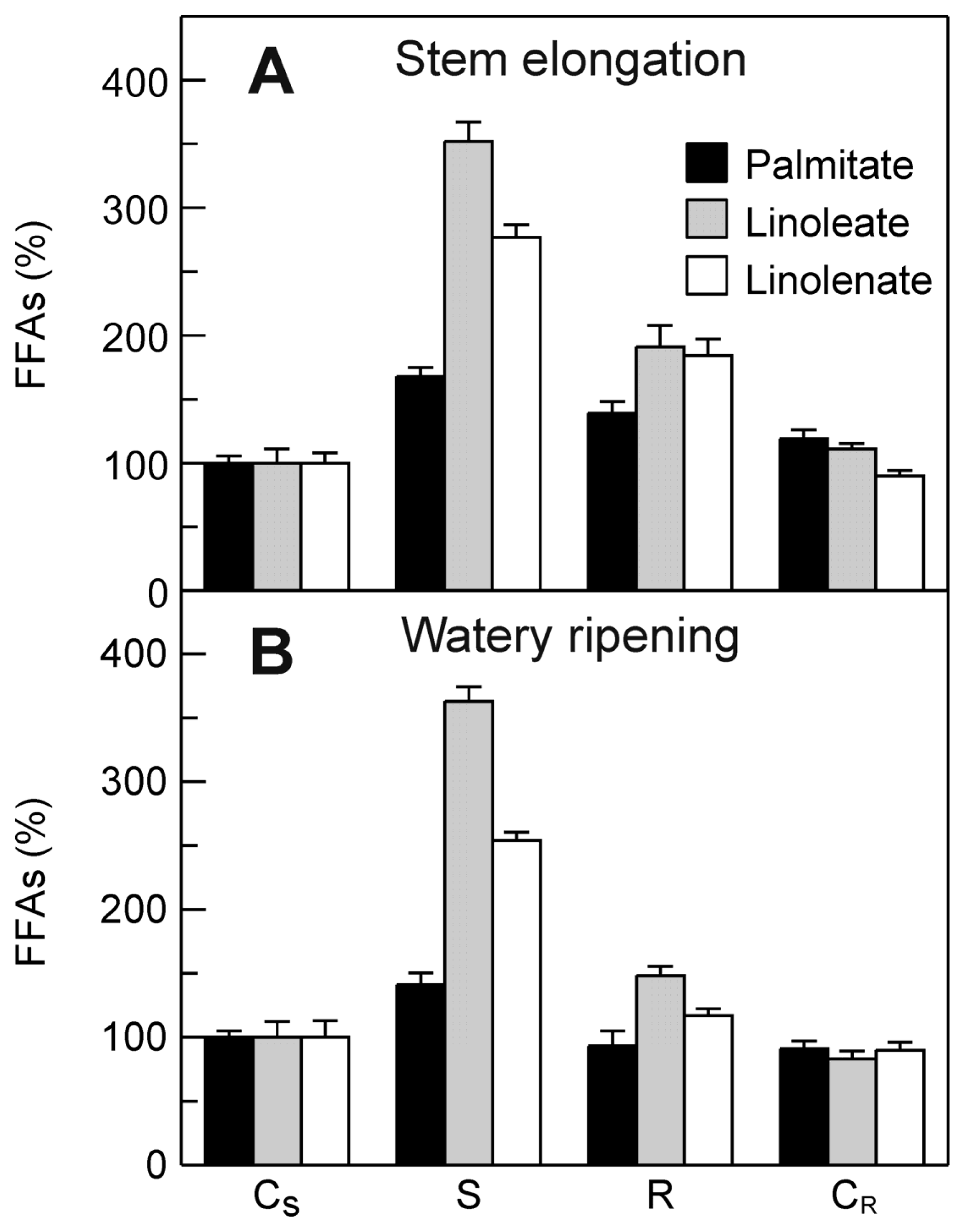
2.5. Evaluation of the Effect of the Drought Stress on FFA Content in Durum Wheat Leaves
3. Discussion
3.1. Identification and Characterization of the sPLA2 Genes and the DWL-PLA2 Activity in Durum Wheat
3.2. Effect of Drought Stress on TdsPLA2 Gene Expression, DWL-PLA2 Activity and FFA Release
4. Experimental Section
4.1. Plant Material and Growing Conditions
4.2. Isolation and Sequence Analysis of the Full-Length sPLA2 Transcripts
4.3. Semi-Quantitative RT-PCR Analysis
4.4. Assay of PLA2 Activity in the Crude Leaf Extract
4.5. Determination of FFA Content in Durum Wheat Leaves
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Munnik, T.; Testerink, C. Plant phospholipid signaling: “In a nutshell”. J. Lipid Res 2009, 50, S260–S265. [Google Scholar]
- Wang, G.; Ryu, S.; Wang, X. Plant phospholipases: An overview. Methods Mol. Biol 2012, 861, 123–137. [Google Scholar]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta 2006, 1761, 1246–1259. [Google Scholar]
- Scherer, G.F.E.; Ryu, S.B.; Wang, X.; Matos, A.R.; Heitz, T. Patatin-related phospholipase A: Nomenclature, subfamilies and functions in plants. Trends Plant Sci 2010, 15, 693–700. [Google Scholar]
- Ståhl, U.; Ek, B.; Stymne, S. Purification and characterization of a low-molecular-weight phospholipase A2 from developing seeds of elm. Plant Physiol 1998, 117, 197–205. [Google Scholar]
- Ståhl, U.; Lee, M.; Sjodahl, S.; Archer, D.; Cellini, F.; Ek, B.; Iannacone, N.; MacKenzie, D.; Semeraro, L.; Tramontano, E.; et al. Plant low-molecular-weight phospholipase A2s (PLA2s) are structurally related to the animal secretory PLA2s and are present as a family of isoforms in rice (Oryza sativa). Plant Mol. Biol 1999, 41, 481–490. [Google Scholar]
- Domingues, S.J.S.; Souza, T.F.; Soares, A.M.S.; Jacinto, T.; Machado, O.L.T. Activation of phospholipase PLA2 activity in Ricinus communis leaves in response to mechanical wounding. Braz. J. Plant Physiol 2007, 19, 35–42. [Google Scholar]
- Kim, J.Y.; Chung, Y.S.; Ok, S.H.; Lee, S.G.; Chung, W.I.; Kim, I.Y.; Shin, J.S. Characterization of the full-length sequences of phospholipase A2 induced during flower development. Biochim. Biophys. Acta 1999, 1489, 389–392. [Google Scholar]
- Lee, H.Y.; Bahn, S.C.; Shin, J.S.; Hwang, I.; Back, K.; Doelling, J.H.; Ryu, S.B. Multiple forms of secretory phospholipase A2 in plants. Prog. Lipid Res 2005, 44, 52–67. [Google Scholar]
- Mariani, M.E.; Villarreal, M.A.; Cheung, F.; Leiva, E.P.; Madoery, R.R.; Fidelio, G.D. In silico and in vitro characterization of phospholipase A2 isoforms from soybean (Glycine max). Biochimie 2012, 94, 2608–2619. [Google Scholar]
- Matos, A.R.; Pham-Thi, A.-T. Lipid deacylating enzymes in plants: Old activities, new genes. Plant Physiol. Biochem 2009, 47, 491–503. [Google Scholar]
- Reina-Pinto, J.J.; Voisin, D.; Kurdyukov, S.; Faust, A.; Haslam, R.P.; Michaelson, L.V.; Efremova, N.; Franke, B.; Schreiber, L.; Napier, J.A.; et al. Misexpression of FATTY ACID ELONGATION1 in the Arabidopsis epidermis induces cell death and suggests a critical role for phospholipase A2 in this process. Plant Cell 2009, 21, 1252–1272. [Google Scholar]
- Gustavsson, M.H.; Sommarin, M. Characterization of a plasma membrane-associated phospholipase A2 activity increased in response to cold acclimation. J. Plant Physiol 2002, 159, 1219–1227. [Google Scholar]
- Holk, A.; Rietz, S.; Zahn, M.; Quader, H.; Scherer, G.F.E. Molecular identification of cytosolic, patatin-related phospholipases A from Arabidopsis with potential functions in plant signal transduction. Plant Physiol 2002, 130, 90–101. [Google Scholar]
- Rietz, S.; Dermendjiev, G.; Oppermann, E.; Tafesse, F.G.; Effendi, Y.; Holk, A.; Parkera, J.E.; Teige, M.; Scherer, G.F.E. Roles of Arabidopsis patatin-related phospholipases A in root development are related to auxin responses and phosphate deficiency. Mol. Plant 2010, 3, 525–538. [Google Scholar]
- Li, M.; Bahn, S.C.; Guo, L.; Musgrave, W.; Berg, H.; Welti, R.; Wang, X. Patatin-related phospholipase pPLAIIIβ-induced changes in lipid metabolism alter cellulose content and cell elongation in Arabidopsis. Plant Cell 2011, 23, 1107–1123. [Google Scholar]
- Lee, H.Y.; Bahn, S.C.; Kang, Y.-M.; Lee, K.H.; Kim, H.J.; Noh, E.K.; Palta, J.P.; Shin, J.S.; Ryu, S.B. Secretory low molecular weight phospholipase A2 plays important roles in cell elongation and shoot gravitropism in Arabidopsis. Plant Cell 2003, 15, 1990–2002. [Google Scholar]
- Seo, J.; Lee, H.Y.; Choi, H.; Choi, Y.; Lee, Y.; Kim, Y.-W.; Ryu, S.B.; Lee, Y. Phospholipase A2β mediates light-induced stomatal opening in Arabidopsis. J. Exp. Bot 2008, 59, 3587–3594. [Google Scholar]
- Lee, O.R.; Kim, S.J.; Kim, H.J.; Hong, J.K.; Ryu, S.B.; Lee, S.H.; Ganguli, A.; Cho, H.-T. Phospholipase A2 is required for PIN-FORMED protein trafficking to the plasma membrane in the Arabidopsis root. Plant Cell 2010, 22, 1812–1825. [Google Scholar]
- Kim, H.J.; Ok, S.H.; Bahn, S.C.; Jang, J.; Oh, S.A.; Park, S.K.; Twell, D.; Ryu, S.B.; Shin, J.S. Endoplasmic reticulum- and Golgi-localized phospholipase A2 plays critical roles in Arabidopsis pollen development and germination. Plant Cell 2011, 23, 94–110. [Google Scholar]
- Jung, J.; Kumar, K.; Lee, H.Y.; Park, Y., II; Cho, H.-T.; Ryu, S.B. Translocation of phospholipase A2α to apoplasts is modulated by developmental stages and bacterial infection in. Arabidopsis. Front. Plant Sci 2012. [Google Scholar] [CrossRef]
- Heinze, M.; Herre, M.; Massalski, C.; Hermann, I.; Conrad, U.; Roos, W. Signal transfer in the plant plasma membrane: Phospholipase A2 is regulated via an inhibitory Gα protein and a cyclophilin. Biochem. J. 2012. [Google Scholar] [CrossRef]
- Ryu, S.B.; Lee, H.Y.; Hwang, I.W.; Jiwan, P.P. Transgenic plants with increased resistance to biotic and abiotic stresses and accelerated flowering time due to overexpression of a secretory phospholipase A2 (sPLA2). PCT Int. Appl. Patent US2010/0100984A1, 22 April 2010. [Google Scholar]
- Singh, A.; Baranwal, V.; Shankar, A.; Kanwar, P.; Ranjan, R.; Yadav, S.; Pandey, A.; Kapoor, S.; Pandey, G.K. Rice phospholipase A superfamily: Organization, phylogenetic and expression analysis during abiotic stresses and development. PLoS One 2012, 7, e30947. [Google Scholar]
- Elias, E.M.; Manthey, F.A. End products: present and future uses. In Durum Wheat Breeding: Current Approaches and Future Strategies; Royo, C., Nachit, M.M., di Fonzo, N., Araus, J.L., Pfeiffer, W.H., Slafer, G.A., Eds.; Food Products Press: New York, NY, USA, 2005; Volume 1, pp. 63–86. [Google Scholar]
- Mansfeld, J. Plant phospholipases A2: Perspectives on biotechnological applications. Biotechnol. Lett 2009, 31, 1373–1380. [Google Scholar]
- NCBI Conserved Domain Database. Available online: http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml accessed on 12 March 2012.
- Yang, H.-C.; Mosior, M.; Johnson, C.A.; Chen, Y.; Dennis, E.A. Group-specific assays that distinguish between the four major types of mammalian phospholipase A2. Anal. Biochem 1999, 269, 278–288. [Google Scholar]
- Winget, J.M.; Pan, Y.H.; Bahnson, B.J. The interfacial binding surface of phospholipase A2s. Biochim. Biophys. Acta 2006, 1761, 1260–1269. [Google Scholar]
- URGI Wheat Sequence Repository. Available online: http://urgi.versailles.inra.fr/Species/Wheat/Sequence-Repository accessed on 10 September 2012.
- Ježek, J.; Jabůrek, M.; Zelenka, J.; Ježek, P. Mitochondrial phospholipase A2 activated by reactive oxygen species in heart mitochondria induces mild uncoupling. Physiol. Res 2010, 59, 737–747. [Google Scholar]
- Trono, D.; Soccio, M.; Laus, M.N.; Pastore, D. The existence of phospholipase A2 activity in plant mitochondria and its activation by hyperosmotic stress in durum wheat (Triticum durum Desf.). Plant Sci. 2013, 199–200, 91–102. [Google Scholar]
- Fujikawa, Y.; Fujikawa, R.; Iijima, N.; Esaka, M. Characterization of secretory phospholipase A2 with phospholipase A1 activity in tobacco, Nicotiana tabacum (L.). Lipids 2012, 47, 303–312. [Google Scholar]
- Mansfeld, J.; Ulbrich-Hofmann, R. Secretory phospholipase A2α from Arabidopsis thaliana: Functional parameters and substrate preference. Chem. Phys. Lipids 2007, 150, 156–166. [Google Scholar]
- Fujikawa, R.; Fujikawa, Y.; Iijima, N.; Esaka, M. Molecular cloning, expression, and characterization of secretory phospholipase A2 in tobacco. Lipids 2005, 40, 901–908. [Google Scholar]
- AOCS Lipid Library. Available online: http://lipidlibrary.aocs.org accessed on 25 October 2012.
- Bahn, S.C.; Lee, H.Y.; Kim, H.J.; Ryu, S.B.; Shin, J.S. Characterization of Arabidopsis secretory phospholipase A2-γ cDNA and its enzymatic properties. FEBS Lett 2003, 553, 113–118. [Google Scholar]
- Jung, K.M.; Kim, D.K. Purification and characterization of a membrane-associated 48-kilodalton phospholipase A2 in leaves of broad bean. Plant Physiol 2000, 123, 1057–1068. [Google Scholar]
- Rietz, S.; Holk, A.; Scherer, G.F.E. Expression of the patatin related phospholipase A gene AtPLA IIA in Arabidopsis thaliana is up-regulated by salicylic acid, wounding, ethylene, and iron and phosphate deficiency. Planta 2004, 219, 743–753. [Google Scholar]
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar]
- Passioura, J.B.; Condon, A.G.; Richards, R.A. Water deficits, the development of leaf area and crop productivity. In Water Deficits Plant Responses from Cell to Community; Smith, J.A.C., Griffiths, H., Eds.; BIOS Scientific Publishers limited: Oxford, UK, 1993; pp. 253–264. [Google Scholar]
- Kader, M.A.; Lindberg, S. Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal Behav 2010, 5, 233–238. [Google Scholar]
- Reddy, A.S.N. Calcium: Silver bullet in signaling. Plant Sci 2001, 160, 381–404. [Google Scholar]
- Schachtman, D.P.; Goodger, J.Q.D. Chemical root to shoot signalling under drought. Trends Plant Sci 2008, 13, 281–287. [Google Scholar]
- Felle, H.H.; Herrmann, A.; Hückelhoven, R.; Kogel, K.-H. Root-to-shoot signalling: apoplastic alkalinization, a general stress response and defence factor in barley (Hordeum vulgare). Protoplasma 2005, 227, 17–24. [Google Scholar]
- Sahsah, Y.; Campos, P.; Gareil, M.; Zuily-Fodil, Y.; Pham-Thi, A.T. Enzymatic degradation of polar lipids in Vigna unguiculata leaves and influence of drought stress. Physiol. Plant 1998, 104, 577–586. [Google Scholar]
- Matos, A.R.; d’Arcy-Lameta, A.; França, M.; Pêtres, S.; Edelman, L.; Kader, J.; Zuily-Fodil, Y.; Pham-Thi, A.T. A novel patatin-like gene stimulated by drought stress encodes a galactolipid acyl hydrolase. FEBS Lett 2001, 491, 188–192. [Google Scholar]
- Matos, A.R.; Gigon, A.; Laffray, D.; Petrês, S.; Zuily-Fodil, Y.; Pham-Thi, A.T. Effects of progressive drought stress on the expression of patatin-like lipid acyl hydrolase genes in Arabidopsis leaves. Physiol. Plant 2008, 134, 110–120. [Google Scholar]
- Alferez, F.; Lluch, Y.; Burns, J.K. Phospholipase A2 and postharvest peel pitting in citrus fruit. Postharv. Biol. Technol 2008, 49, 69–76. [Google Scholar]
- Liao, H.-L.; Burns, J.K. Light controls phospholipase A2α and β gene expression in Citrus sinensis. J. Exp. Bot 2010, 61, 2469–2478. [Google Scholar]
- Navari-Izzo, F.; Milone, M.T.; Quartacci, M.F.; Pinzini, C. Metabolic changes in wheat plants subjected to a water-deficit stress programme. Plant Sci 1993, 92, 151–157. [Google Scholar]
- Quartacci, M.F.; Pinzino, C.; Sgherri, C.L.M.; Navari-Izzo, F. Lipid composition and protein dynamics in thylakoids of two wheat cultivars differently sensitive to drought. Plant Physiol 1995, 108, 191–197. [Google Scholar]
- Weber, H. Fatty acid-derived signals in plants. Trends Plant Sci 2002, 7, 217–224. [Google Scholar]
- Lehmann, J.; Atzorn, R.; Brückner, C.; Reinbothe, S.; Leopold, J.; Wasternack, C. Accumulation of jasmonate, abscisic acid, specific transcripts and proteins in osmotically stressed barley leaf segments. Planta 1995, 1, 156–162. [Google Scholar]
- Walia, H.; Wilson, C.; Condamine, P.; Liu, X.; Ismail, A.M.; Closem, T.J. Large-scale expression profiling and physiological characterization of jasmonic acid mediated adaptation of barley to salinity stress. Plant Cell Environ 2007, 30, 410–421. [Google Scholar]
- Rice Annotation Project Database. Available online: http://rapdb.dna.affrc.go.jp accessed on 10 May 2011.
- ExPASy ProtParam tool. Available online: http://web.expasy.org/protparam/ accessed on 21 February 2012.
- iPSORT. Available online: http://ipsort.hgc.jp accessed on 20 March 2012.
- TargetP. Available online: http://www.cbs.dtu.dk/services/TargetP accessed on 20 March 2012.
- Predotar. Available online: http://urgi.versailles.inra.fr/predotar/predotar.html accessed on 20 March 2012.
- Harris, D.A. Spectrophotometric assays. In Spectrophotometry and Spectrofluorimetry: A Practical Approach; Bashford, C.L., Harris, D.A., Eds.; IRL Press: Oxford, UK, 1987; pp. 59–61. [Google Scholar]
- Pastore, D.; Trono, D.; Padalino, L.; Simone, S.; Valenti, D.; Di Fonzo, N.; Passarella, S. Inhibition by α-tocopherol and L-ascorbate of linoleate hydroperoxidation and β-carotene bleaching activities in durum wheat semolina. J. Cereal Sci 2000, 31, 41–54. [Google Scholar]
- Gambacorta, G.; Sinigaglia, M.; Schena, A.; Baiano, A.; Lamacchia, C.; Pati, S.; La Notte, E. Changes in free fatty acid and diacylglycerol compounds in short-ripening dry-cured sausage. J. Food Lipids 2009, 16, 1–18. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amplicon | Forward primer (5′→3′) | Reverse primer (5′→3′) | Annealing temperature (°C) | Product size (bp) |
|---|---|---|---|---|
| Full-length | ||||
| TdsPLA2I | ATGGCGATGGCGATGGCGATG | CTACAGTTCTAACTTCTGGCTGCCC | 60 | 426 |
| TdsPLA2II | ATGAGATCGGTGCTCGGTC | CTACTGCCCGATGTCGCG | 58 | 474 |
| TdsPLA2III | ATGGCGCATGGCAGAGGC | CTAGGGCTTGTGCAGGACCCG | 60 | 489 |
| TdsPLA2IV | ATGCACACCGGCCGCCTCCTCCC | CTACTTCGCCGGGGCCGGCGCC | 62 | 513 |
| Fragment | ||||
| TdsPLA2I | GTCCTCCTCCTGGCCGGGGGC | CAGGCATCGAGGTCGTCGCAGGG | 67 | 170 |
| TdsPLA2II | TGCTTCTCCTCTCGCTGGTGACG | TCGCCGGGGCAGCCGCTGTAG | 62 | 184 |
| TdsPLA2III | ACGTCGGCCTCCAGACCCTCG | TCCAGCAGGCCCTCGTTGCAC | 63 | 253 |
| TdsPLA2IV | GCGGTACGGCAAGTACTGCGGCG | CCACGCAGTCCAGCAGGCTCTGG | 66 | 157 |
| actin1 | CTTCGGACCCAAGAAAGAAAGCC | CACCGCCCGTATTTCTCTAGTAGCC | 62 | 280 |
| cor410 | AGAAGAAGGAGGAGGAGGACAAGAAGAAGG | AGAAGCCTTTCTTCTCCTCCTCGGG | 58 | 432 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Verlotta, A.; Liberatore, M.T.; Cattivelli, L.; Trono, D. Secretory Phospholipases A2 in Durum Wheat (Triticum durum Desf.): Gene Expression, Enzymatic Activity, and Relation to Drought Stress Adaptation. Int. J. Mol. Sci. 2013, 14, 5146-5169. https://doi.org/10.3390/ijms14035146
Verlotta A, Liberatore MT, Cattivelli L, Trono D. Secretory Phospholipases A2 in Durum Wheat (Triticum durum Desf.): Gene Expression, Enzymatic Activity, and Relation to Drought Stress Adaptation. International Journal of Molecular Sciences. 2013; 14(3):5146-5169. https://doi.org/10.3390/ijms14035146
Chicago/Turabian StyleVerlotta, Angelo, Maria T. Liberatore, Luigi Cattivelli, and Daniela Trono. 2013. "Secretory Phospholipases A2 in Durum Wheat (Triticum durum Desf.): Gene Expression, Enzymatic Activity, and Relation to Drought Stress Adaptation" International Journal of Molecular Sciences 14, no. 3: 5146-5169. https://doi.org/10.3390/ijms14035146
APA StyleVerlotta, A., Liberatore, M. T., Cattivelli, L., & Trono, D. (2013). Secretory Phospholipases A2 in Durum Wheat (Triticum durum Desf.): Gene Expression, Enzymatic Activity, and Relation to Drought Stress Adaptation. International Journal of Molecular Sciences, 14(3), 5146-5169. https://doi.org/10.3390/ijms14035146