Curcumin Enhances Cytotoxic Effects of Bortezomib in Human Multiple Myeloma H929 Cells: Potential Roles of NF-κB/JNK

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
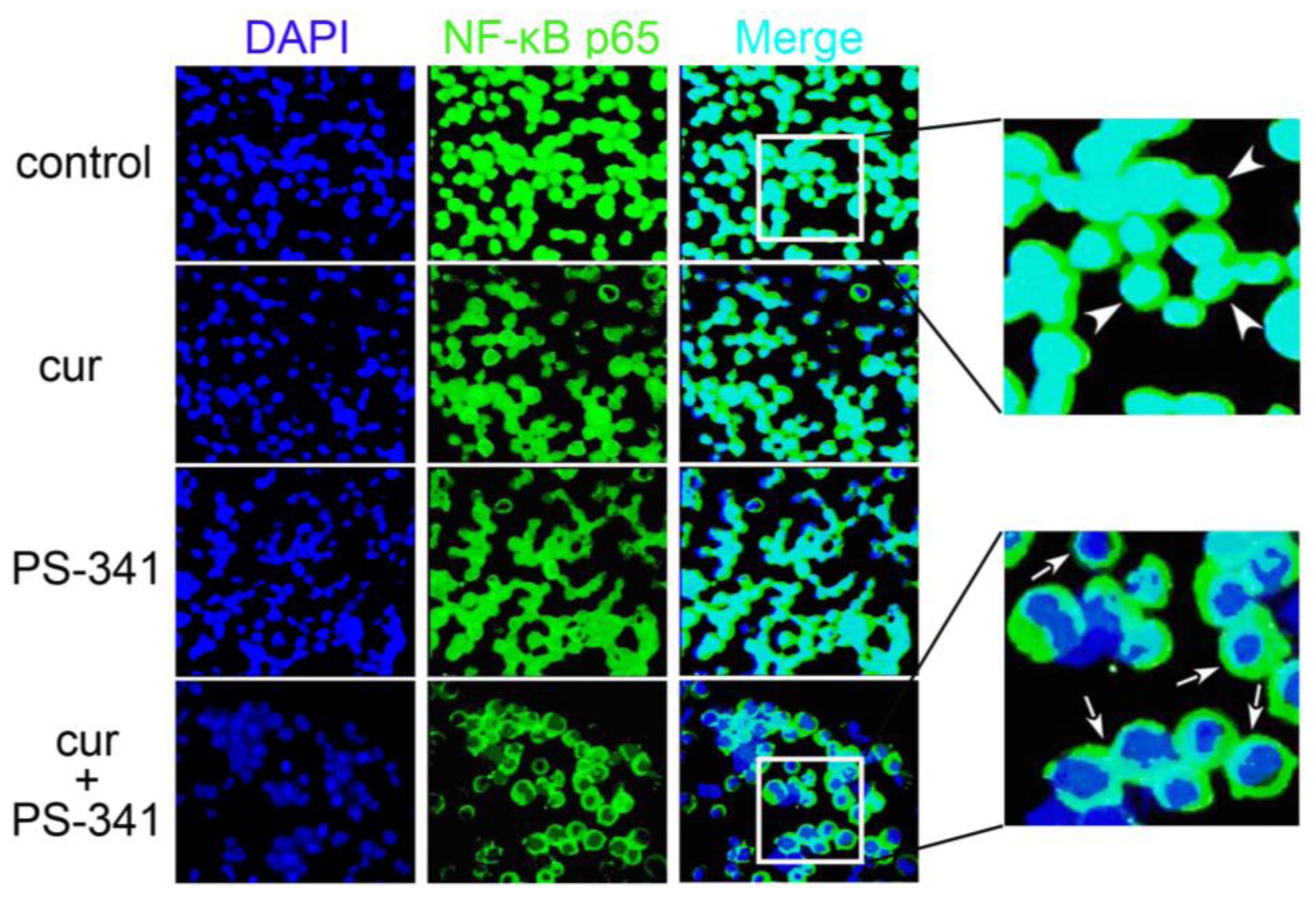
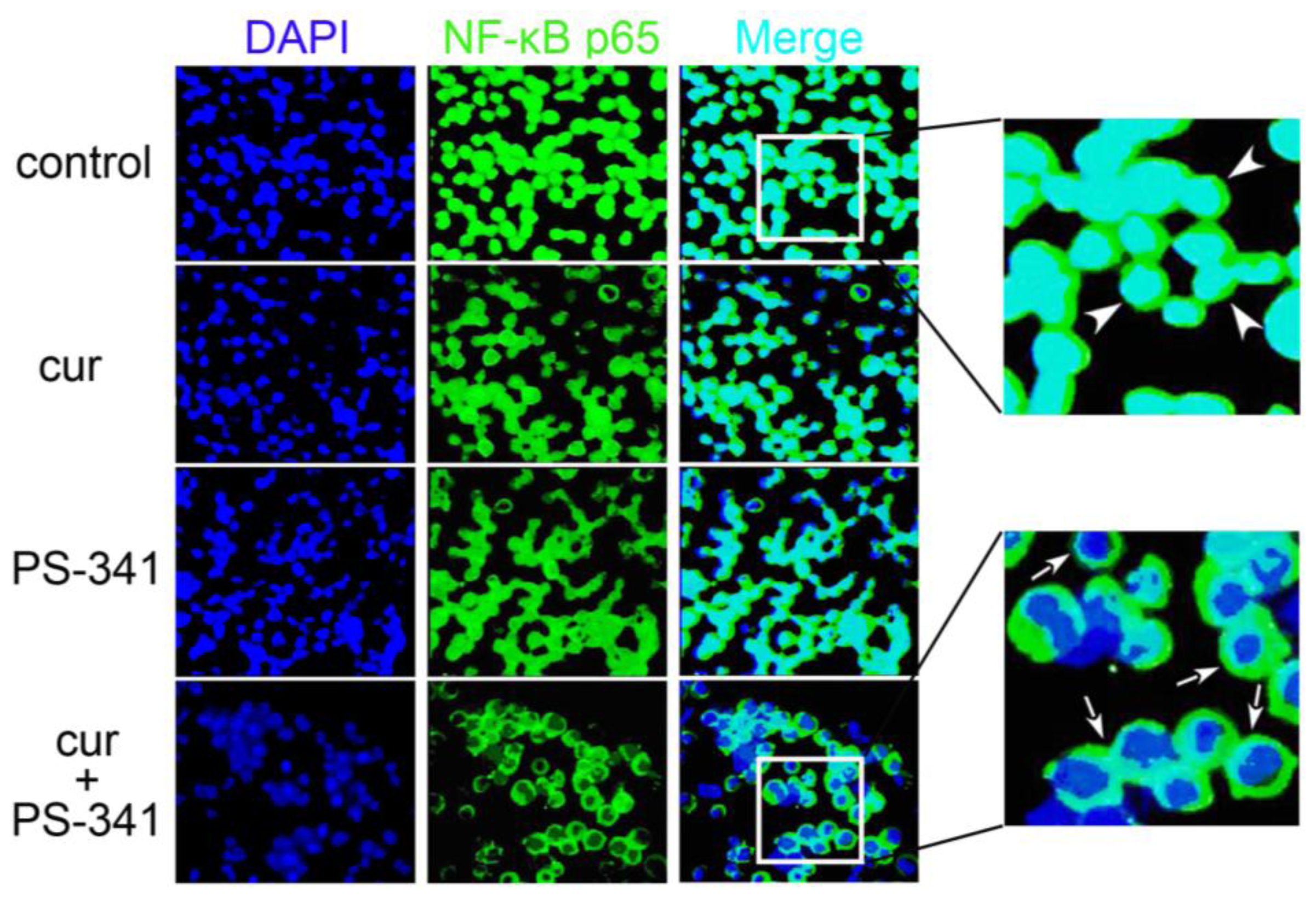
2.1. Curcumin and PS-341 Altered Expression and Distribution Profiles of NF-κB p65
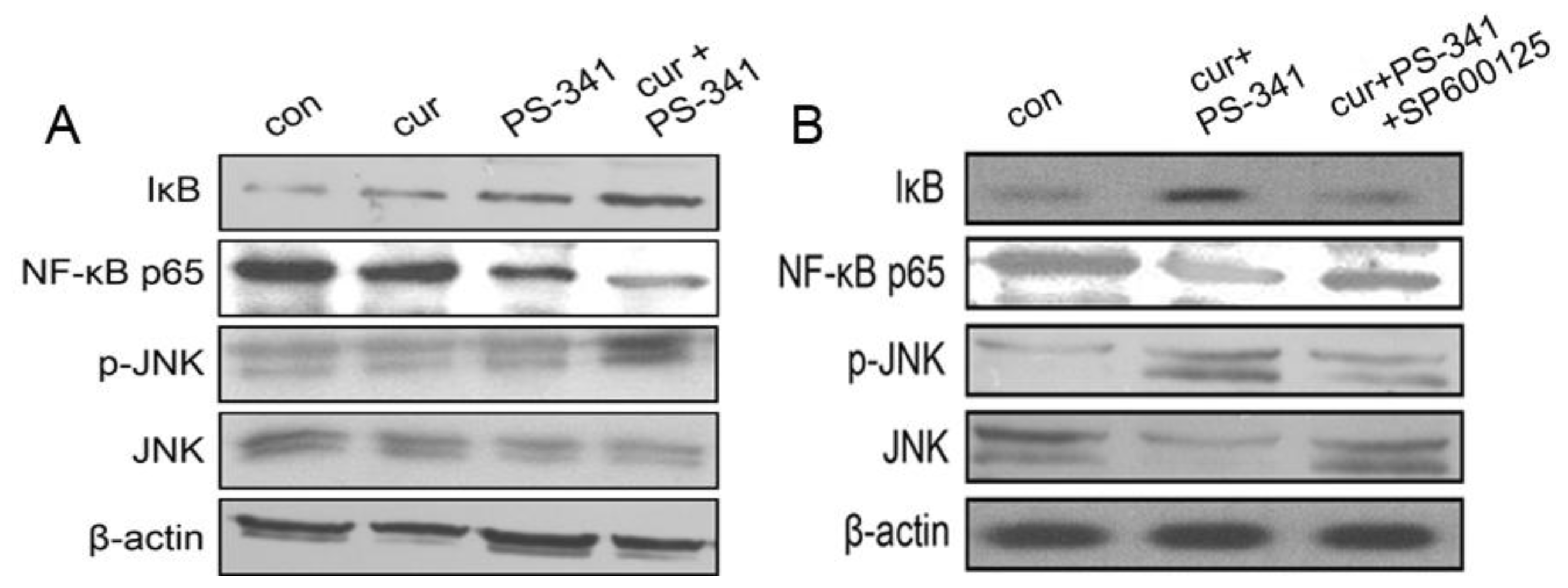
2.2. Curcumin and PS-341 Stabilized IκB through JNK Mechanism
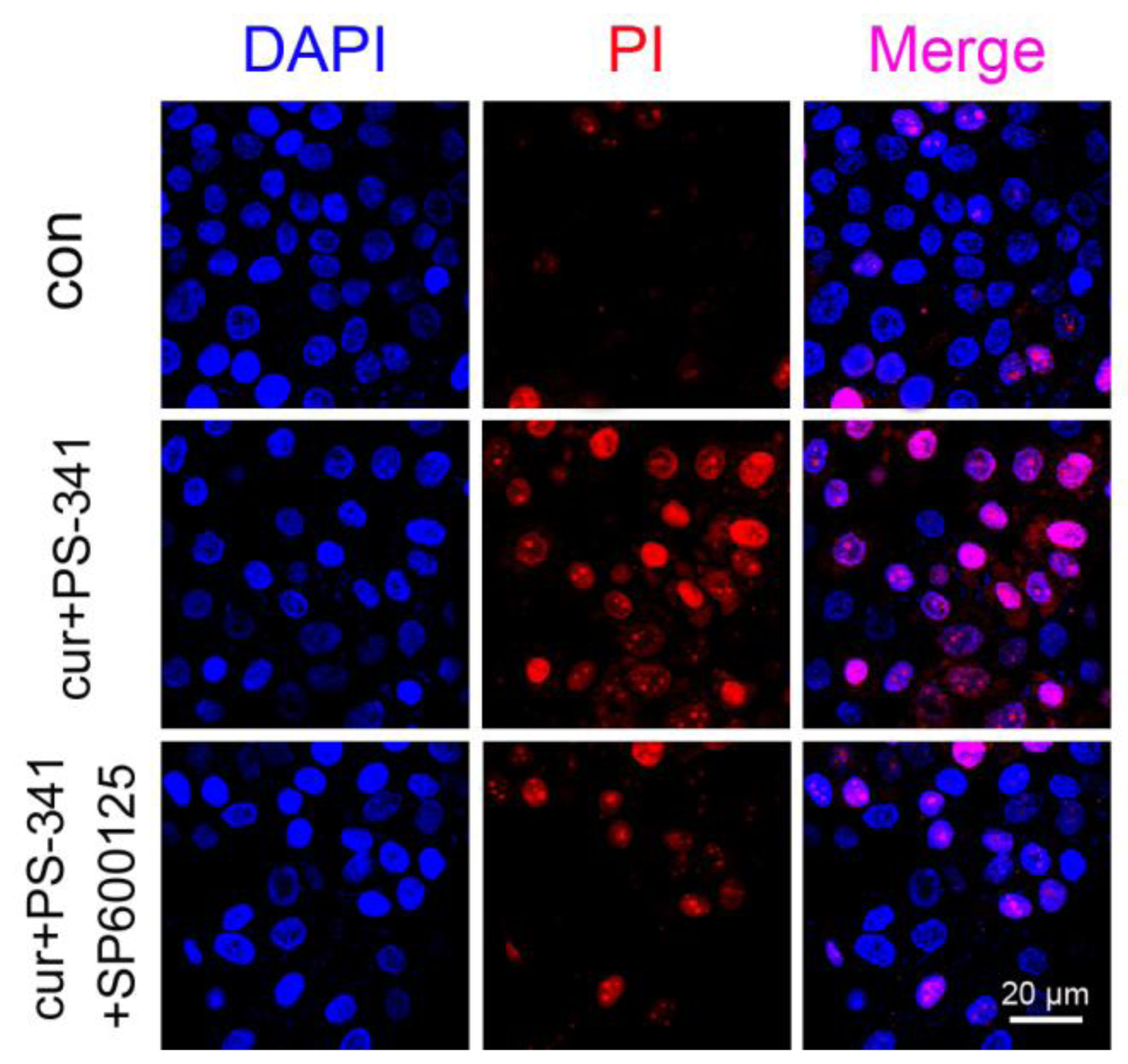
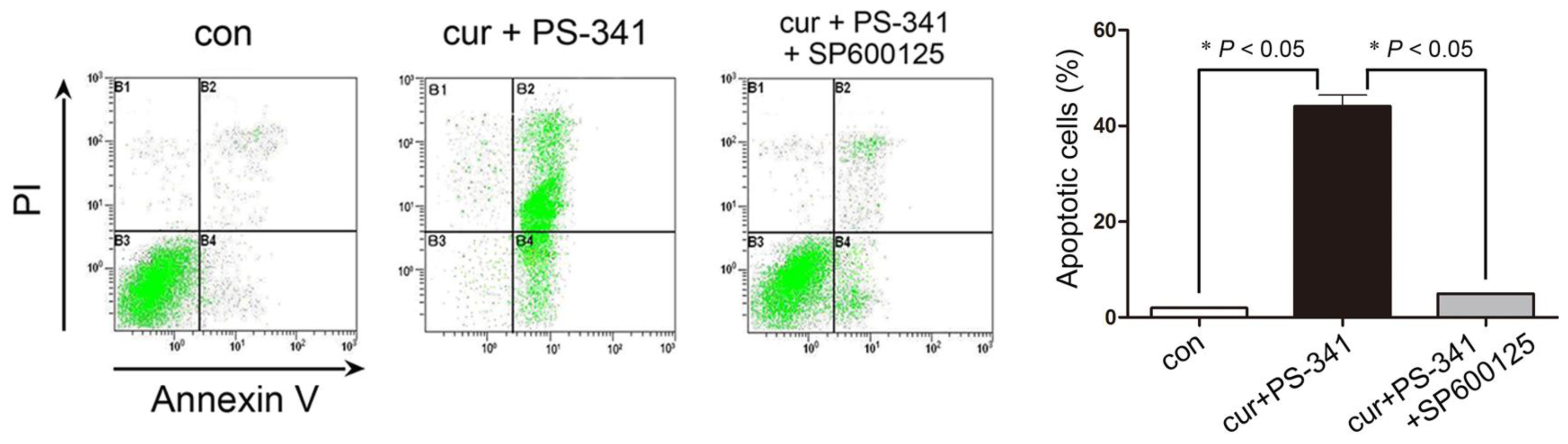
2.3. Blockage of JNK Restored H929 Cells Survival
3. Experimental Section
3.1. Reagents
3.2. Cell Culture
3.3. Fluorescent Immunocytochemistry of NF-κB p65
3.4. Assay of IκB, NF-κB p65 and JNK
3.5. Analysis of JNK in H929 Cell Apoptosis
3.6. Statistical Analysis
4. Conclusion
Acknowledgments
References
- Kyle, R.A.; Rajkumar, S.V. Multiple myeloma. N. Engl. J. Med 2004, 351, 1860–1873. [Google Scholar]
- Adams, J.; Kauffman, M. Development of the proteasome inhibitor Velcade (Bortezomib). Cancer Invest 2004, 22, 304–311. [Google Scholar]
- Almond, J.B.; Cohen, G.M. The proteasome: A novel target for cancer chemotherapy. Leukemia 2002, 16, 433–443. [Google Scholar]
- Ling, Y.H.; Liebes, L.; Ng, B.; Buckley, M.; Elliott, P.J.; Adams, J.; Jiang, J.D.; Muggia, F.M.; Perez-Soler, R. PS-341, a novel proteasome inhibitor, induces Bcl-2 phosphorylation and cleavage in association with G2-M phase arrest and apoptosis. Mol. Cancer Ther 2002, 1, 841–849. [Google Scholar]
- Dai, Y.; Rahmani, M.; Grant, S. Proteasome inhibitors potentiate leukemic cell apoptosis induced by the cyclin-dependent kinase inhibitor flavopiridol through a SAPK/JNK- and NF-κB-dependent process. Oncogene 2003, 22, 7108–7122. [Google Scholar]
- Shah, J.J.; Orlowski, R.Z. Proteasome inhibitors in the treatment of multiple myeloma. Leukemia 2009, 23, 1964–1979. [Google Scholar]
- Kouroukis, C.T.; Belch, A.; Crump, M.; Eisenhauer, E.; Gascoyne, R.D.; Meyer, R.; Lohmann, R.; Lopez, P.; Powers, J.; Turner, R.; et al. Flavopiridol in untreated or relapsed mantle-cell lymphoma: Results of a phase II study of the National Cancer Institute of Canada Clinical Trials Group. J. Clin. Oncol 2003, 21, 1740–1745. [Google Scholar]
- Holkova, B.; Grant, S. Combining proteasome with cell cycle inhibitors: A dual attack potentially applicable to multiple hematopoietic malignancies. Expert Rev. Hematol 2011, 4, 483–486. [Google Scholar]
- Singh, S.; Aggarwal, B.B. Activation of transcription factor NF-kappa B is suppressed by curcumin (diferuloylmethane). J. Biol. Chem 1995, 270, 24995–25000. [Google Scholar]
- Hussain, A.R.; Ahmed, M.; Al-Jomah, N.A.; Khan, A.S.; Manogaran, P.; Sultana, M.; Abubaker, J.; Platanias, L.C.; Al-Kuraya, K.S.; Uddin, S. Curcumin suppresses constitutive activation of nuclear factor-kappa B and requires functional Bax to induce apoptosis in Burkitt’s lymphoma cell lines. Mol. Cancer Ther 2008, 7, 3318–3329. [Google Scholar]
- Lee, K.H.; Chow, Y.L.; Sharmili, V.; Abas, F.; Alitheen, N.B.M.; Shaari, K.; Israf, D.A.; Lajis, N.H.; Syahida, A. BDMC33, a curcumin derivative suppresses inflammatory responses in macrophage-like cellular system: Role of inhibition in NF-κB and MAPK signaling pathways. Int. J. Mol. Sci 2012, 13, 2985–3008. [Google Scholar]
- Patel, B.B.; Sengupta, R.; Qazi, S.; Vachhani, H.; Yu, Y.; Rishi, A.K.; Majumdar, A.P. Curcumin enhances the effects of 5-fluorouracil and oxaliplatin in mediating growth inhibition of colon cancer cells by modulating EGFR and IGF-1R. Int. J. Cancer 2008, 122, 267–273. [Google Scholar]
- Aggarwal, B.B.; Shishodia, S.; Takada, Y.; Banerjee, S.; Newman, R.A.; Bueso-Ramos, C.E.; Price, J.E. Curcumin suppresses the paclitaxel-induced nuclear factor-kappaB pathway in breast cancer cells and inhibits lung metastasis of human breast cancer in nude mice. Clin. Cancer Res 2005, 11, 7490–7498. [Google Scholar]
- Sung, B.; Kunnumakkara, A.B.; Sethi, G.; Anand, P.; Guha, S.; Aggarwal, B.B. Curcumin circumvents chemoresistance in vitro and potentiates the effect of thalidomide and bortezomib against human multiple myeloma in nude mice model. Mol. Cancer Ther 2009, 8, 959–970. [Google Scholar]
- Park, J.; Ayyappan, V.; Bae, E.K.; Lee, C.; Kim, B.S.; Kim, B.K.; Lee, Y.Y.; Ahn, K.S.; Yoon, S.S. Curcumin in combination with bortezomib synergistically induced apoptosis in human multiple myeloma U266 cells. Mol. Oncol 2008, 2, 317–326. [Google Scholar]
- Zhang, X.Y.; Bai, Q.X.; Huang, G.S.; Zhao, H.; Chen, J.J.; Yang, L.J. Effect of curcumin in combination with bortezomib on proliferation and apoptosis of human multiple myeloma cell line H929 and its mechanism. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2011, 19, 684–688. [Google Scholar]
- Mujtaba, T.; Kanwar, J.; Wan, S.B.; Chan, T.H.; Dou, Q.P. Sensitizing human multiple myeloma cells to the proteasome inhibitor bortezomib by novel curcumin analogs. Int. J. Mol. Med 2012, 29, 102–106. [Google Scholar]
- Liang, J.; Saad, Y.; Lei, T.; Wang, J.; Qi, D.; Yang, Q.; Kolattukudy, P.E.; Fu, M. MCP-induced protein 1 deubiquitinates TRAF proteins and negatively regulates JNK and NF-κB signaling. J. Exp. Med 2010, 207, 2959–2973. [Google Scholar]
- Hsieh, I.N.; Chang, A.S.; Teng, C.M.; Chen, C.C.; Yang, C.R. Aciculatin inhibits lipopolysaccharide-mediated inducible nitric oxide synthase and cyclooxygenase-2 expression via suppressing NF-κB and JNK/p38 MAPK activation pathways. J. Biomed. Sci 2011, 18. [Google Scholar] [CrossRef]

© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bai, Q.-X.; Zhang, X.-Y. Curcumin Enhances Cytotoxic Effects of Bortezomib in Human Multiple Myeloma H929 Cells: Potential Roles of NF-κB/JNK. Int. J. Mol. Sci. 2012, 13, 4831-4838. https://doi.org/10.3390/ijms13044831
Bai Q-X, Zhang X-Y. Curcumin Enhances Cytotoxic Effects of Bortezomib in Human Multiple Myeloma H929 Cells: Potential Roles of NF-κB/JNK. International Journal of Molecular Sciences. 2012; 13(4):4831-4838. https://doi.org/10.3390/ijms13044831
Chicago/Turabian StyleBai, Qing-Xian, and Xiao-Yan Zhang. 2012. "Curcumin Enhances Cytotoxic Effects of Bortezomib in Human Multiple Myeloma H929 Cells: Potential Roles of NF-κB/JNK" International Journal of Molecular Sciences 13, no. 4: 4831-4838. https://doi.org/10.3390/ijms13044831
APA StyleBai, Q.-X., & Zhang, X.-Y. (2012). Curcumin Enhances Cytotoxic Effects of Bortezomib in Human Multiple Myeloma H929 Cells: Potential Roles of NF-κB/JNK. International Journal of Molecular Sciences, 13(4), 4831-4838. https://doi.org/10.3390/ijms13044831