Evolution of Microsatellite Loci of Tropical and Temperate Anguilla Eels

Abstract
:1. Introduction
2. Results and Discussion
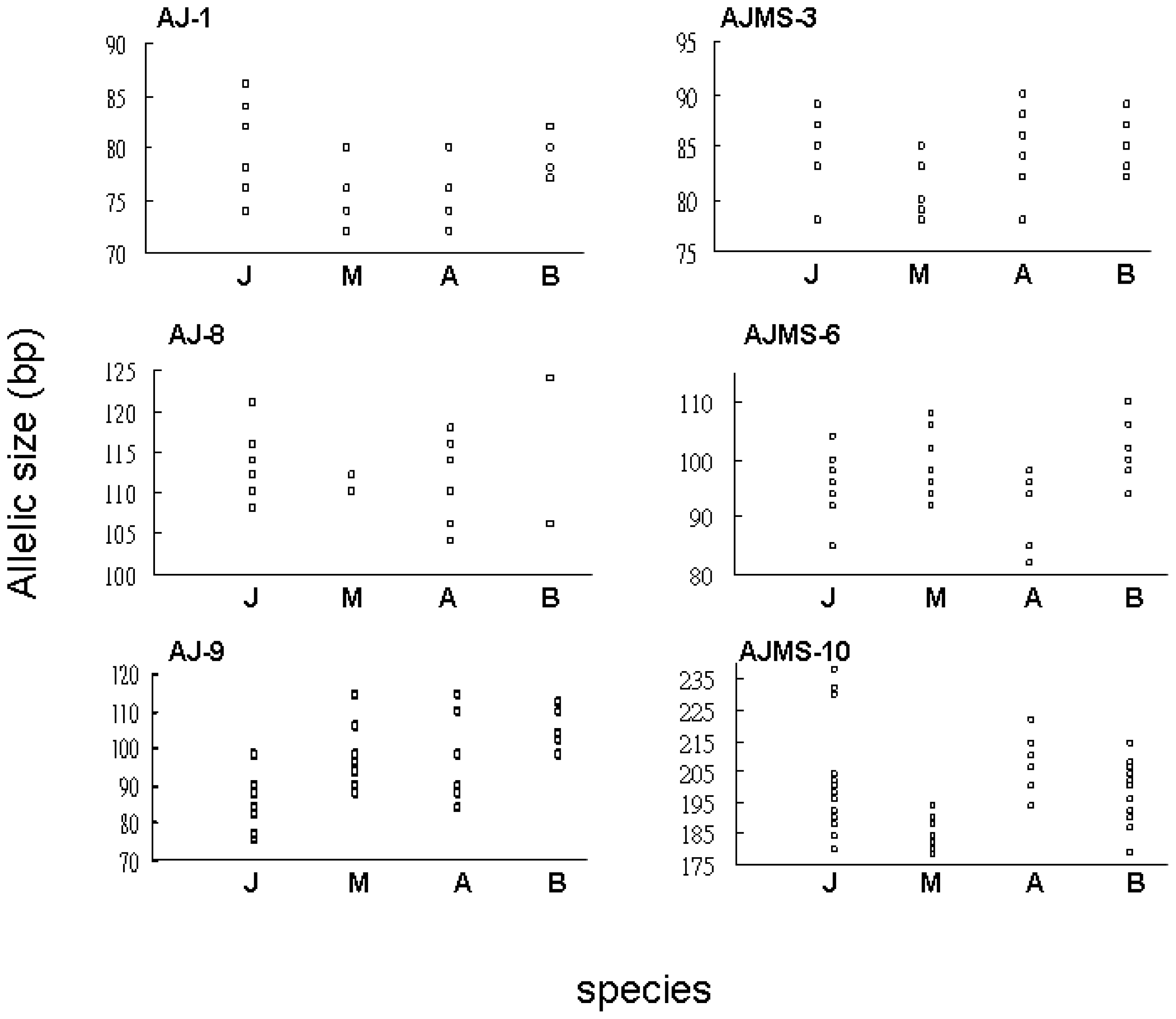
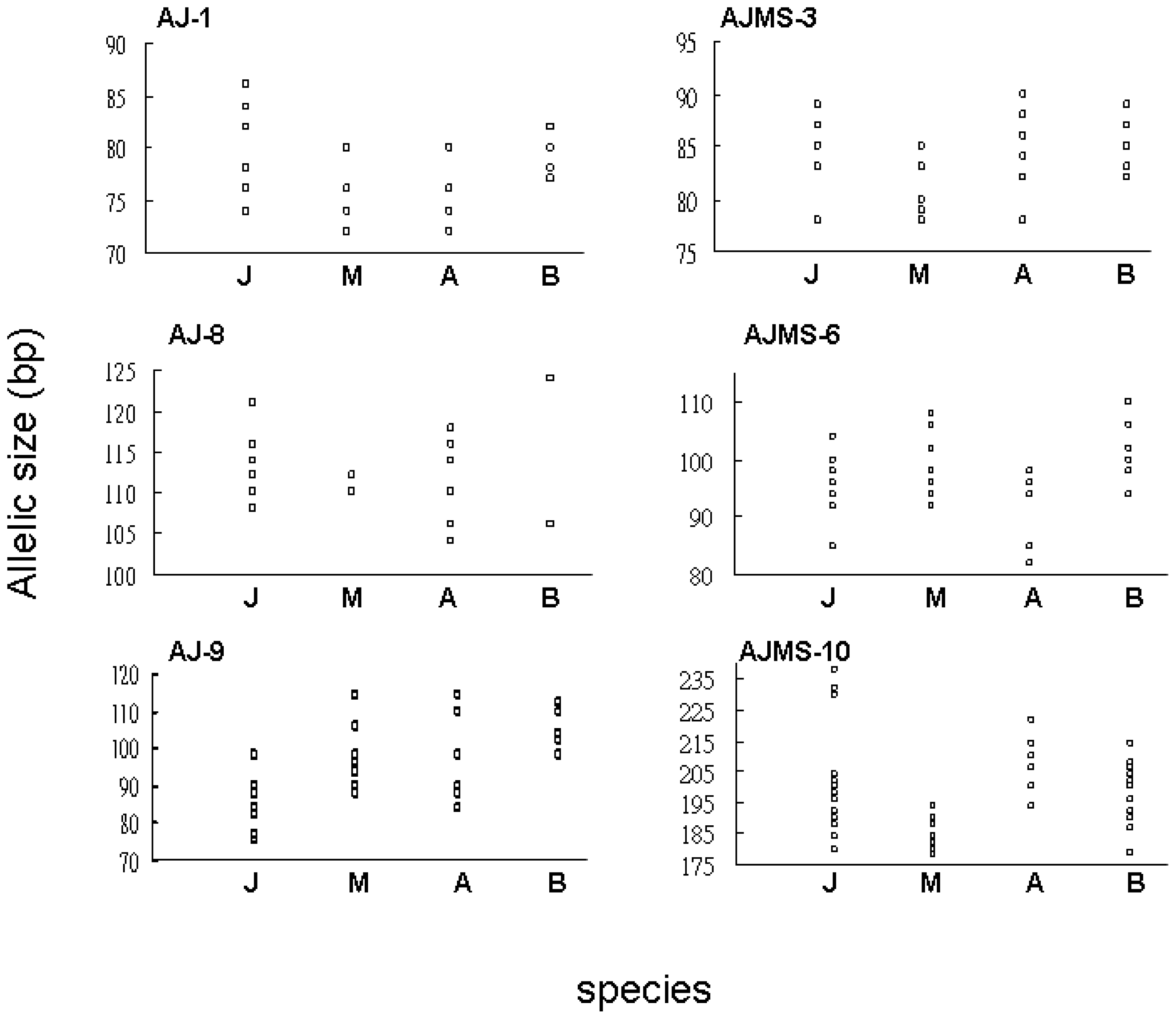
2.1. Microsatellite Variation
2.2. Flanking Region Analysis
2.3. Evolutionary Mechanism of Microsatellite Loci
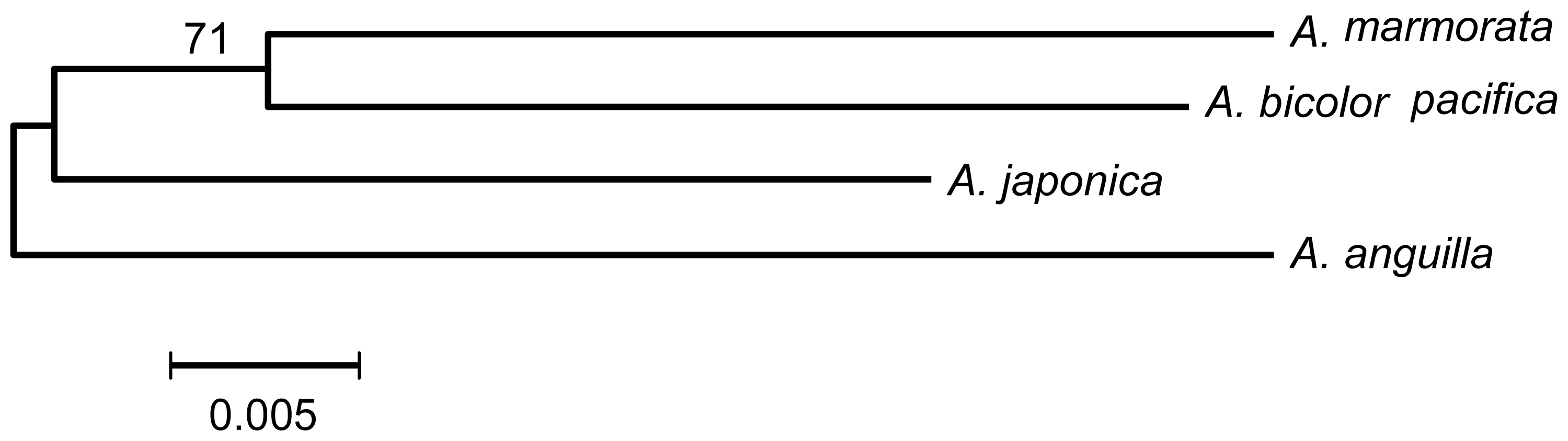
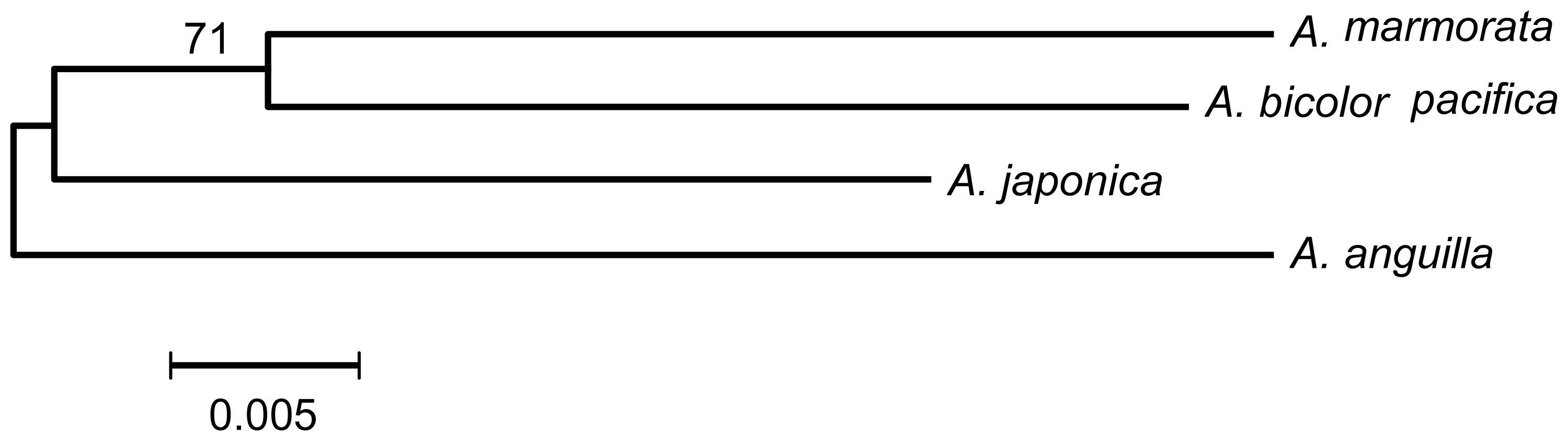
2.4. Are A. japonica and A. anguilla a Sister Group?
3. Experimental Section
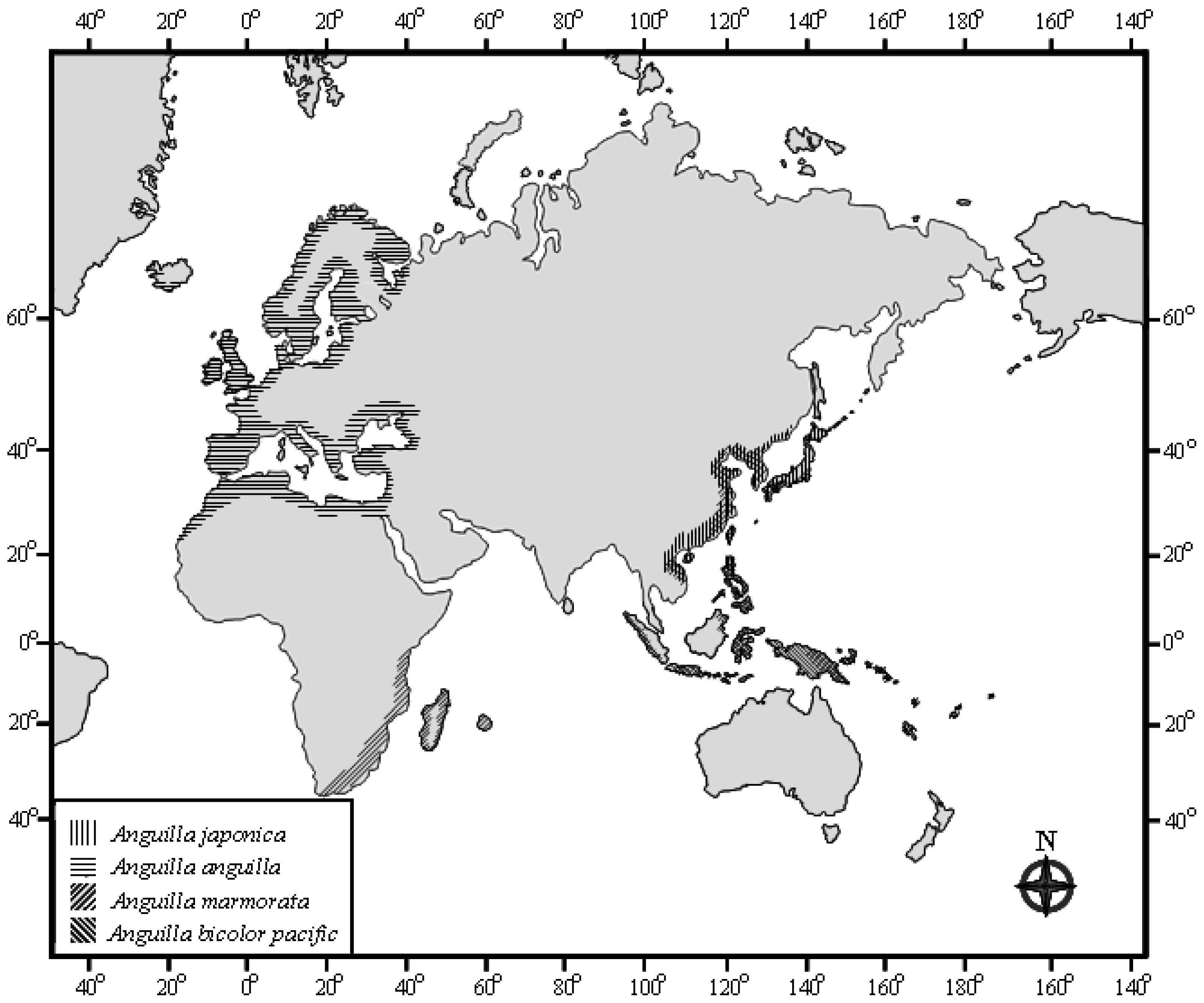
3.1. Sampling
3.2. Microsatellite Cloning and Sequencing
3.3. Data Analysis
4. Conclusions
Acknowledgments
References
- Teng, H.Y.; Lin, T.S.; Tzeng, C.S. A new Anguilla species and a reanalysis of the phylogeny of freshwater eels. Zool. Stud 2009, 48, 808–822. [Google Scholar]
- Vladykov, V.D.; March, H. Distribution of leptocephali of two species of Anguilla in the western North Atlantic based on collections made between 1933 and 1968. Syllogeus 1975, 6, 1–38. [Google Scholar]
- Ege, V. A revision of the genus Anguilla Shaw a systematic phylogenetic and geographical study. Dana Rep 1939, 16, 1–256. [Google Scholar]
- Jespersen, P. Indo-Pacific leptocephali of the genus Anguilla. Dana Rep 1942, 22, 1–128. [Google Scholar]
- Arai, T.; Marui, M.; Otake, T.; Tsukamoto, K. Inshore migration of a tropical eel Anguilla marmorata from Taiwanese and Japanese coasts. Fish. Sci 2002, 68, 152–157. [Google Scholar]
- Aoyama, J.; Mochioka, N.; Otake, T.; Ishikawa, S.; Kawakami, Y.; Castle, P.; Nishida, M.; Tsukamoto, K. Distribution and dispersal of anguillid leptocephali in the western Pacific Ocean revealed by molecular analysis. Mar. Ecol. Prog. Ser 1999, 188, 193–200. [Google Scholar]
- Bastrop, R.; Strehlow, B.; Jürse, K.; Sturmbauer, C. A new molecular phylogenetic hypothesis for the evolution of freshwater eels. Mol. Phylogenet. Evol 2000, 14, 250–258. [Google Scholar]
- Minegishi, Y.; Aoyama, J.; Inoue, J.G.; Miya, M.; Nishida, M.; Tsukamoto, K. Molecular phylogeny and evolution of the freshwater eels genus Anguilla based on the whole mitochondrial genome sequences. Mol. Phylogenet. Evol 2005, 34, 134–146. [Google Scholar]
- Lin, Y.S.; Poh, Y.P.; Tzeng, C.S. Phylogeny of freshwater eels inferred from mitochondrial genes. Mol. Phylogenet. Evol 2001, 20, 252–261. [Google Scholar]
- Aoyama, J.; Nishida, M.; Tsukamoto, K. Molecular phylogeny and evolution of the freshwater eels genus Anguilla. Mol. Phylogenet. Evol 2001, 20, 450–459. [Google Scholar]
- Baranski, M.; Moen, T.; Våge, D.I. Mapping of quantitative trait loci for flesh colour and growth traits in Atlantic salmon (Salmo salar). Genet. Sel. Evol 2010, 42. [Google Scholar] [CrossRef]
- Malausa, T.; Dalecky, A.; Ponsard, S.; Audiot, P.; Streiff, R.; Chaval, Y.; Bourguet, D. Genetic structure and gene flow in French populations of two Ostrinia taxa: Host races or sibling species? Mol. Ecol 2007, 6, 4210–4222. [Google Scholar]
- Pilot, M.; Dabrowski, M.J.; Jancewicz, E.; Schtickzelle, N.; Gliwicz, J. Temporally stable genetic variability and dynamic kinship structure in a fluctuating population of the root vole Microtus oeconomus. Mol. Ecol 2010, 19, 2800–2812. [Google Scholar]
- Gottelli, D.; Sillero-Zubiri, C.; Applebaum, G.D.; Roy, M.S.; Girman, D.J.; Garcia-Moreno, J.; Ostrander, E.A.; Wayne, R.K. Molecular genetics of the most endangered canid: The Ethiopian wolf Canis simensis. Mol. Ecol. 1994, 3, 301–312. [Google Scholar]
- Viard, F.; Bremond, P.; Labbo, R.; Justy, F.; Delay, B.; Jarne, P. Microsatellites and the genetics of highly selfing populations in the freshwater snail Bulinus truncates. Genetics 1996, 142, 1237–1247. [Google Scholar]
- Pinheiro, M.; Ahlquist, T.; Danielsen, S.A.; Lind, G.E.; Veiga, I.; Pinto, C.; Costa, V.; Afonso, L.; Sousa, O.; Fragoso, M. Colorectal carcinomas with microsatellite instability display a different pattern of target gene mutations according to large bowel site of origin. BMC Cancer 2010, 10. [Google Scholar] [CrossRef]
- Asahida, T.; Gray, A.K.; Gharrett, A.J. Use of microsatellite locus flanking regions for phylogenetic analysis? A preliminary study of Sebastes subgenera. Environ. Biol. Fish 2004, 69, 461–470. [Google Scholar]
- Zardoya, R.; Vollmer, D.M.; Craddock, C.; Streelman, J.T.; Karl, S.; Meyer, A. Evolutionary conservation of microsatellite flanking regions and their use in resolving the phylogeny of Cichlid fishes (Pisces: Perciformes). Proc. Biol. Sci 1996, 263, 1589–1598. [Google Scholar]
- Goldstein, D.B.; Schlötterer, C. Microsatellites Evolution and Applications; Oxford University Press, Inc: New York, NY, USA, 1999. [Google Scholar]
- Weber, J.L.; Wong, C. Mutation of human short tandem repeats. Hum. Mol. Genet 1993, 2, 1123–1128. [Google Scholar]
- Primmer, C.R.; Ellegren, H.; Saino, N.; Moller, A.P. Directional evolution in germline microsatellite mutations. Nat. Genet 1996, 13, 391–393. [Google Scholar]
- Kimura, M.; Crow, J.F. The number of alleles that can be maintained in a finite population. Genetics 1964, 49, 725–738. [Google Scholar]
- Crow, J.F.; Kimura, M. An Introduction to Population Genetic Theory; Harper & Row, Press: New York, NY, USA, 1970. [Google Scholar]
- Kimura, M.; Ohta, T. Stepwise mutation model and distribution of allelic frequencies in a finite population. Proc. Natl. Acad. Sci. USA 1978, 75, 2868–2872. [Google Scholar]
- Di Rienzo, A.; Peterson, A.C.; Garza, J.C.; Valders, A.M.; Slatkin, M.; Freimer, N.B. Mutational processes of simple-sequence repeat loci in human populations. Proc. Natl. Acad. Sci. USA 1994, 91, 3166–3170. [Google Scholar]
- Schlötterer, C.; Tautz, D. Slippage synthesis of simple sequence DNA. Nucleic Acids Res 1992, 20, 211–215. [Google Scholar]
- Strand, M.; Prolla, T.A.; Liskay, R.M.; Petes, T.D. Destabilization of tracts of simple repetitive DNA in yeast by mutations affecting DNA mismatch repair. Nature 1993, 365, 274–276. [Google Scholar]
- Levinson, G.; Gutman, G.A. High frequencies of short frameshifts in poly-CA/TG tandem repeats borne by bacteriophage M13 in Escherichia coli K-12. Nucleic Acids Res 1987, 15, 5289–5304. [Google Scholar]
- Weber, J.L. Informativeness of human (dC-dA)n (dG-dT)n polymorphisms. Genomics 1990, 7, 524–530. [Google Scholar]
- Wierdl, M.; Dominska, M.; Petes, T.D. Microsatellite instability in yeast: Dependence on the length of the microsatellite. Genetics 1997, 146, 769–779. [Google Scholar]
- Eisen, J.A. Mechanistic Basis for Microsatellite Instability. In Microsatellites; Goldstein, D.B., Schlötterer, C., Eds.; Oxford University Press, Inc.: New York, NY, USA, 1999; pp. 34–48. [Google Scholar]
- Stephan, W. Tandem-repetitive noncoding DNA: Forms and forces. Mol. Biol. Evol 1989, 6, 198–212. [Google Scholar]
- Kuittinen, H.; Salguero, D.; Aguadé, M. Parallel patterns of sequence variation within and between populations at three loci of Arabidopsis thaliana. Mol. Biol. Evol 2002, 19, 2030–2034. [Google Scholar]
- Blankenship, S.M.; May, B.; Hedgecock, D. Evolution of a perfect simple sequence repeat locus in the context of its flanking sequence. Mol. Biol. Evol 2002, 19, 1943–1951. [Google Scholar]
- Colson, I.; Goldstein, D.B. Evidence for complex mutations at microsatellite loci in Drosophila. Genetics 1999, 152, 617–627. [Google Scholar]
- Patterson, C. Osteichthyes: Teleostei. In The Fossil Record; Benton, M.J., Ed.; Chapman & Hall: Landon, UK, 1993; Volume 2, pp. 621–656. [Google Scholar]
- Haq, B.U. Paleoceanography: A Synoptic Overview of 200 Million Years of Ocean History. In Marine Geography and Oceanography of Arabian Sea and Coastal Pakistan; Haq, B.U., Milliman, D.J., Eds.; Van Nostrand-Reinhold: New York, NY, USA, 1984; pp. 201–234. [Google Scholar]
- Aoyama, J.; Tsukamoto, K. Evolution of the freshwater eels. Naturwissenschaften 1997, 84, 17–21. [Google Scholar]
- Tsukamoto, K.; Aoyama, J. Evolution of the freshwater eels of the genus Anguilla: A probable scenario. Environ. Biol. Fish 1998, 52, 139–148. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatic, T. Molecular Cloning a Laboratory Manual, 2nd ed; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Tseng, M.C.; Chen, C.A.; Kao, H.W.; Tzeng, W.N.; Lee, S.C. Polymorphisms of GA/GT microsatellite loci from Anguilla japonica. Mar. Biotech 2001, 3, 275–280. [Google Scholar]
- Tseng, M.C.; Tzeng, W.N.; Lee, S.C. Population genetic structure of the Japanese eel Anguilla japonica in the northwest Pacific Ocean: Evidence of non-panmictic populations. Mar. Ecol. Prog. Ser 2006, 308, 221–230. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98 NT. Nucleic Acids Symp. Ser 1999, 41, 95–98. [Google Scholar]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res 2004, 14, 1188–1190. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 40. Mol. Biol. Evol 2007, 24, 1596–1599. [Google Scholar]
- Xia, X.; Xie, Z. DAMBE: Data analysis in molecular biology and evolution. J. Hered 2001, 92, 371–373. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rate of base substitution through comparative studies of nucleotide sequences. J. Mol. Evol 1980, 16, 111–120. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor method: A new method for reconstructing phylogenetic tree. Mol. Biol. Evol 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus Species | Ta | No. of Sequences | Repeat Regions |
|---|---|---|---|
| AJ-1 | |||
| A. japonica | 58 | 11 | (TG)7~13 |
| A. marmorata | 54 | 17 | (TG)6, 7, 8, 10; TC(TG)6 |
| A. anguilla | 54 | 10 | (TG)5~10 |
| A. bicolor pacifica | 54 | 11 | (TG)7~9 |
| AJ-8 | |||
| A. japonica | 56 | 12 | (TG)14~18, 20 |
| A. marmorata | 52 | 2 | (TG)14, 15 |
| A. anguilla | 52 | 11 | (TG)12, 13, 15, 17, 19; (TG)6(TA)(TG)11 |
| A. bicolor pacifica | 52 | 3 | (TG)13, 22 |
| AJ-9 | |||
| A. japonica | 54 | 11 | (TG)8, 9, 11, 12, 14, 15; (TG)8AG(TG)10 |
| A. marmorata | 56 | 9 | (TG)14, 15, 17, 18, 19, 23, 27 |
| A. anguilla | 54 | 12 | (TG)12; (TG)10, 19, 21AG(TG)4 |
| (TG)3CG(GT)10,11 | |||
| (TG)7CG(TG)4AG(TG)6 | |||
| A. bicolor pacifica | 54 | 8 | (TG)9, 10, 13, 14(AG)1, 2(TG)9~12 |
| (TG)5TA(TG)3AG(TG)11 | |||
| AJMS-3 | |||
| A. japonica | 56 | 12 | (TG)7~10 |
| A. marmorata | 56 | 9 | (TG)7, 9, 10 |
| A. anguilla | 54 | 8 | (TG)7, 10, 11; (TG)3, 8CG(TG)1, 4 |
| A. bicolor pacifica | 54 | 8 | (TG)7, 8, 10 |
| AJMS-6 | |||
| A. japonica | 56 | 18 | (TG)9, 12~16, 18 |
| A. marmorata | 54 | 12 | (TG)12, 13, 14, 16, 18, 19 |
| A. anguilla | 54 | 14 | (TG)7, 10, 13, 14, 15 |
| A. bicolor pacifica | 54 | 16 | (TG)12~18TA(TG)0, 2 |
| AJMS-10 | |||
| A. japonica | 58 | 12 | (GA)10, 12, 14~16, 18~22, 35, 36, 39 |
| A. marmorata | 54 | 14 | (GA)8~12, 14, 15, 17 |
| A. anguilla | 54 | 15 | (GA)22 |
| (GA)9, 12, 13, 15, 16, 17GG(GA)6, 9, 10, 11,12 | |||
| A. bicolor pacifica | 54 | 19 | (GA)13~15,17,19, 20, 22, 23, 26 |
| (GA)6, 7GG(GA)5, 12 | |||
| (GA)7CA(GA)12, 13; (GA)9AT(GA)9 | |||
| A. japonica | A. marmorata | A. bicolor pacifica | A. anguilla | |
|---|---|---|---|---|
| A. japonica | --- | 0.050 ± 0.11 | 0.055 ± 0.011 | 0.053 ± 0.011 |
| A. marmorata | 22 | --- | 0.044 ± 0.010 | 0.061 ± 0.012 |
| A. bicolor pacifica | 24 | 20 | --- | 0.057 ± 0.011 |
| A. anguilla | 23 | 27 | 25 | --- |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tseng, M.-C. Evolution of Microsatellite Loci of Tropical and Temperate Anguilla Eels. Int. J. Mol. Sci. 2012, 13, 4281-4294. https://doi.org/10.3390/ijms13044281
Tseng M-C. Evolution of Microsatellite Loci of Tropical and Temperate Anguilla Eels. International Journal of Molecular Sciences. 2012; 13(4):4281-4294. https://doi.org/10.3390/ijms13044281
Chicago/Turabian StyleTseng, Mei-Chen. 2012. "Evolution of Microsatellite Loci of Tropical and Temperate Anguilla Eels" International Journal of Molecular Sciences 13, no. 4: 4281-4294. https://doi.org/10.3390/ijms13044281
APA StyleTseng, M.-C. (2012). Evolution of Microsatellite Loci of Tropical and Temperate Anguilla Eels. International Journal of Molecular Sciences, 13(4), 4281-4294. https://doi.org/10.3390/ijms13044281