Arsenic and Antimony Transporters in Eukaryotes

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
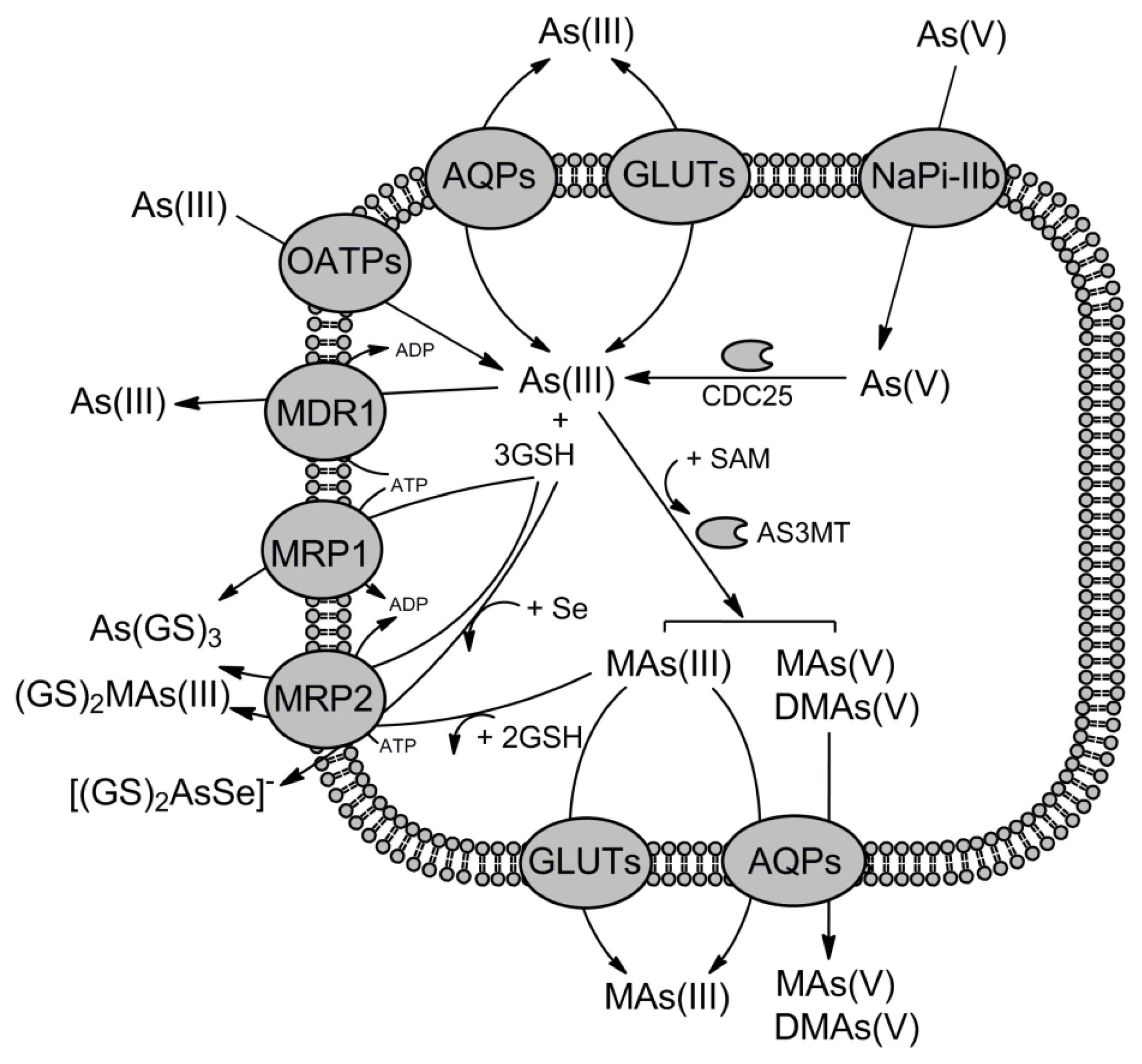
2. Pathways for Arsenic and Antimony Uptake
2.1. Uptake of As(V) via Phosphate Transporters
2.2. Aquaglyceroporins Are the Major Cellular Entrance for As(III) and Sb(III)
2.3. Role of Hexose Transporters in As(III) and Sb(III) Accumulation
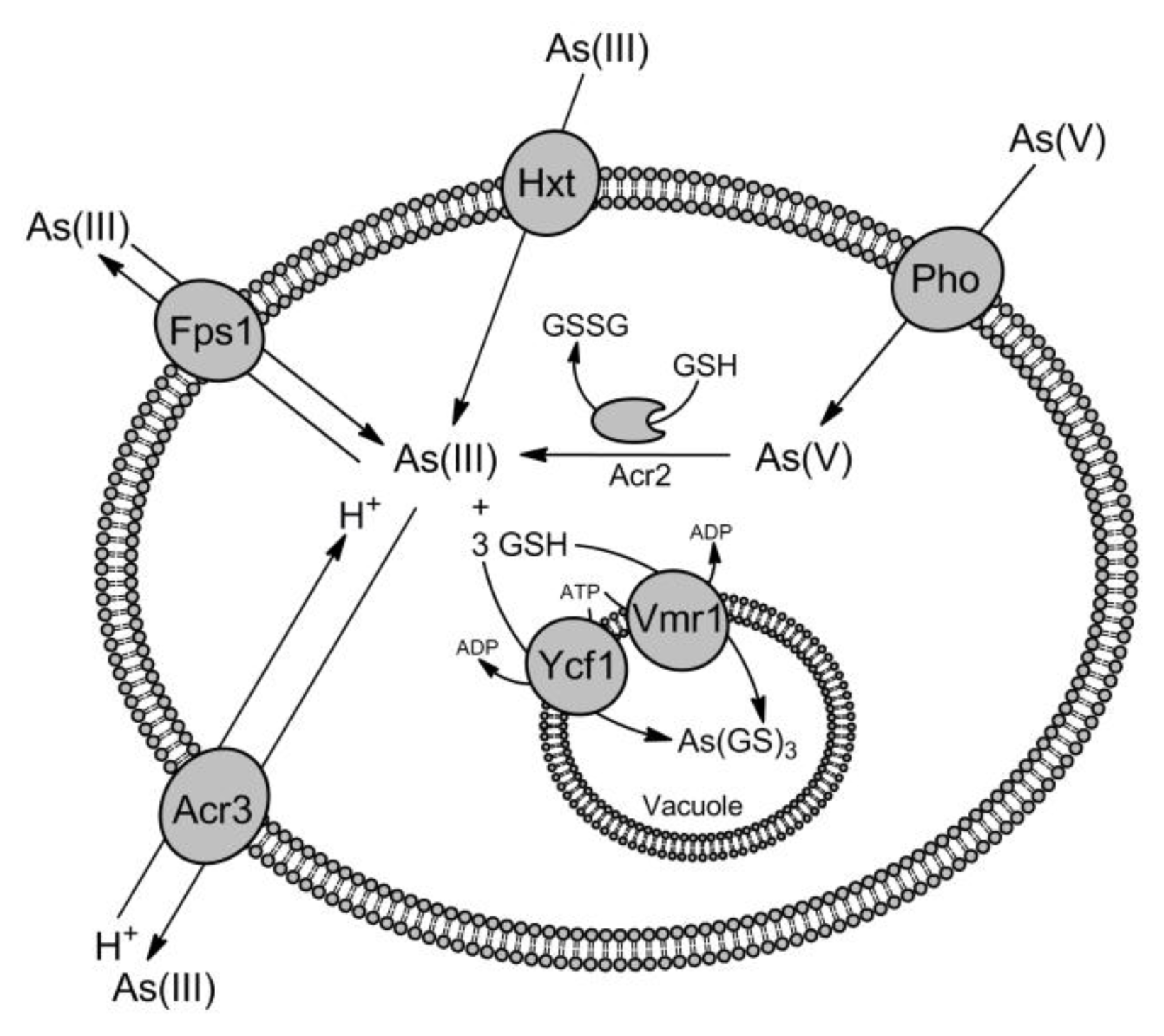
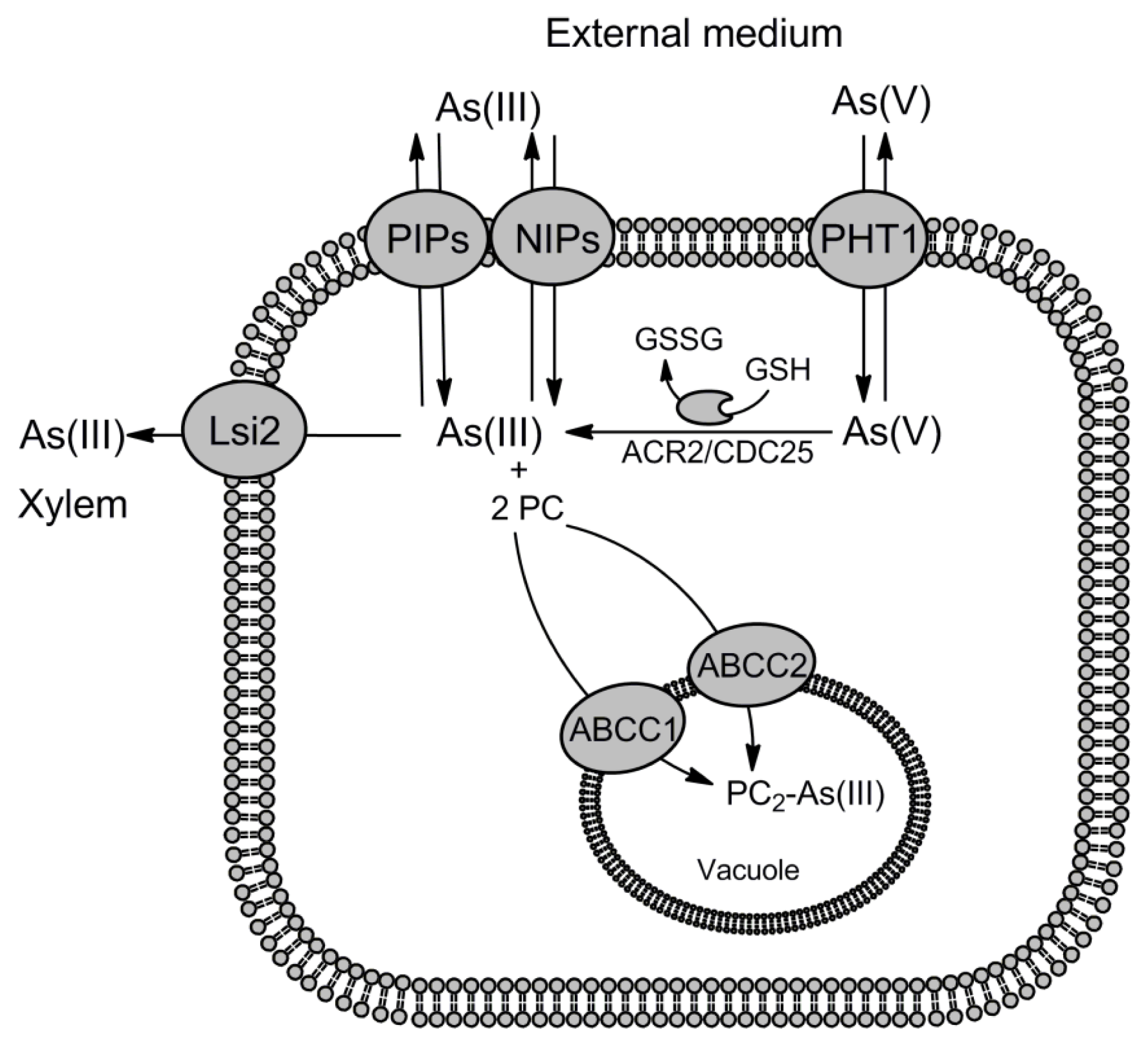
3. Transporters for As(III) and Sb(III) Detoxification
3.1. Involvement of Aquaglyceroporins in As(III) and Sb(III) Efflux
3.2. Acr3 Family of Metalloid Transporters
3.3. Role of ABC Transporters in Metalloid Transport
4. Perspectives
Acknowledgments
References
- Martinez, V.D.; Vucic, E.A.; Becker-Santos, D.D.; Gil, L.; Lam, W.L. Arsenic exposure and the induction of human cancers. J. Toxicol 2011, 2011, 431287. [Google Scholar]
- Garelick, H.; Jones, H.; Dybowska, A.; Valsami-Jones, E. Arsenic pollution sources. Rev. Environ. Contam. Toxicol 2008, 197, 17–60. [Google Scholar]
- Dilda, P.J.; Hogg, P.J. Arsenical-based cancer drugs. Cancer Treat. Rev 2007, 33, 542–564. [Google Scholar]
- Frézard, F.; Demicheli, C. New delivery strategies for the old pentavalent antimonial drugs. Expert. Opin. Drug Deliv 2010, 7, 1343–1358. [Google Scholar]
- Chappuis, F. Melarsoprol-free drug combinations for second-stage Gambian sleeping sickness: the way to go. Clin. Infect. Dis 2007, 45, 1443–1445. [Google Scholar]
- Ali, W.; Isayenkov, S.V.; Zhao, F.J.; Maathuis, F.J. Arsenite transport in plants. Cell. Mol. Life Sci 2009, 66, 2329–2339. [Google Scholar]
- Wysocki, R.; Tamás, M.J. How Saccharomyces cerevisiae copes with toxic metals and metalloids. FEMS Microbiol. Rev 2010, 34, 925–951. [Google Scholar]
- Wysocki, R.; Tamás, M.J. Saccharomyces cerevisiae as a model organism for elucidating arsenic tolerance mechanisms. In Cellular Effects of Heavy Metals, 1st ed; Banfalvi, G., Ed.; Springer: Heidelberg, Germany, 2011; pp. 87–112. [Google Scholar]
- Ramirez-Solis, A.; Mukopadhyay, R.; Rosen, B.P.; Stemmler, T.L. Experimental and theoretical characterization of arsenite in water: insights into the coordination environment of As-O. Inorg. Chem 2004, 43, 2954–2959. [Google Scholar]
- Porquet, A.; Filella, M. Structural evidence of the similarity of Sb(OH)3 and As(OH)3 with glycerol: implications for their uptake. Chem. Res. Toxicol 2007, 20, 1269–1276. [Google Scholar]
- Bienert, G.P.; Schüssler, M.D.; Jahn, T.P. Metalloids: essential, beneficial or toxic? Major intrinsic proteins sort it out. Trends Biochem. Sci 2007, 33, 20–26. [Google Scholar]
- Bhattacharjee, H.; Mukhopadhyay, R.; Thiyagarajan, S.; Rosen, B.P. Aquaglyceroporins: ancient channels for metalloids. J. Biol 2008, 7. [Google Scholar] [CrossRef]
- Brochu, C.; Wang, J.; Roy, G.; Messier, N.; Wang, X.Y.; Saravia, N.G.; Ouellette, M. Antimony uptake systems in the protozoan parasite Leishmania and accumulation differences in antimony-resistant parasites. Antimicrob. Agents Chemother 2003, 47, 3073–3079. [Google Scholar]
- Tschan, M.; Robinson, B.; Schulin, R. Antimony uptake by Zea mays (L.) and Helianthus annuus (L.) from nutrient solution. Environ. Geochem. Health 2008, 30, 187–191. [Google Scholar]
- Persson, B.L.; Lagerstedt, J.O.; Pratt, J.R.; Pattison-Granberg, J.; Lundh, K.; Shokrollahzadeh, S.; Lundh, F. Regulation of phosphate acquisition in Saccharomyces cerevisiae. Curr. Genet 2003, 43, 225–244. [Google Scholar]
- Bun-Ya, M.; Harashima, S.; Oshima, Y. Putative GTP-binding protein, Gtr1, associated with the function of the Pho84 inorganic phosphate transporter in Saccharomyces cerevisiae. Mol. Cell. Biol 1992, 12, 2958–2966. [Google Scholar]
- Bun-ya, M.; Shikata, K.; Nakade, S.; Yompakdee, C.; Harashima, S.; Oshima, Y. Two new genes, PHO86 and PHO87, involved in inorganic phosphate uptake in Saccharomyces cerevisiae. Curr. Genet 1996, 29, 344–351. [Google Scholar]
- Yompakdee, C.; Ogawa, N.; Harashima, S.; Oshima, Y.A. putative membrane protein, Pho88p, involved in inorganic phosphate transport in Saccharomyces cerevisiae. Mol. Gen. Genet 1996, 251, 580–590. [Google Scholar]
- Meharg, A.A.; Macnair, M.R. An altered phosphate uptake system in arsenate tolerant Holcus lanatus. New Phytol 1990, 16, 29–35. [Google Scholar]
- Meharg, A.A.; Macnair, M.R. Mechanisms of arsenate tolerance in Deschampsia cespitosa L. (Beauv.) and Agrostis capillaris L., adaptation of the arsenate uptake system. New Phytol 1991, 119, 291–297. [Google Scholar]
- Abedin, M.J.; Cresser, M.S.; Meharg, A.A.; Feldmann, J.; Cotter-Howells, J. Arsenic accumulation and metabolism in rice (Oryza sativa L.). Environ. Sci. Technol 2002, 36, 962–968. [Google Scholar]
- Shin, H.; Shin, H.S.; Dewbre, G.R.; Harrison, M.J. Phosphate transport in Arabidopsis: Pht1;1 and Pht1;4 play a major role in phosphate acquisition from both low- and high-phosphate environments. Plant J 2004, 39, 629–642. [Google Scholar]
- Gonzáles, E.; Solano, R.; Rubio, V.; Leyva, A.; Paz-Ares, J. PHOSPHATE TRANSPORTER TRAFFIC FACILITATOR1 is a plant-specific SEC12-related protein that enables the endoplasmic reticulum exit of a high-affinity phosphate transporter in Arabidopsis. Plant Cell 2005, 17, 3500–3512. [Google Scholar]
- Wu, Z.; Ren, H.; McGrath, S.P.; Wu, P.; Zhao, F.J. Investigating the contribution of the phosphate transport pathway to arsenic accumulation in rice. Plant Physiol 2011, 157, 498–508. [Google Scholar]
- Villa-Bellosta, R.; Sorribas, V. Arsenate transport by sodium phosphate cotransporter type IIb. Toxicol. Appl. Pharmacol 2010, 247, 36–40. [Google Scholar]
- Beene, L.C.; Halluer, J.; Yoshinaga, M.; Hamdi, M.; Liu, Z. Pentavalent arsenate transport by zebrafish phosphate transporter NaPi-IIb1. Zebrafish 2011, 8, 125–131. [Google Scholar]
- Sanders, O.I.; Rensing, C.; Kuroda, M.; Mitra, B.; Rosen, B.P. Antimonite is accumulated by the glycerol facilitator GlpF in Escherichia coli. J. Bacteriol 1997, 179, 3365–3367. [Google Scholar]
- Wysocki, R.; Chéry, C.C.; Wawrzycka, D.; Van Hulle, M.; Cornelis, R.; Thevelein, J.M.; Tamás, M.J. The glycerol channel Fps1p mediates the uptake of arsenite and antimonite in Saccharomyces cerevisiae. Mol. Microbiol 2001, 40, 1391–1401. [Google Scholar]
- Tamás, M.J.; Luyten, K.; Sutherland, F.C.; Hernandez, A.; Albertyn, J.; Valadi, H.; Li, H.; Prior, B.A.; Kilian, S.G.; Ramos, J.; et al. Fps1p controls the accumulation and release of the compatible solute glycerol in yeast osmoregulation. Mol. Microbiol 1999, 31, 1087–1104. [Google Scholar]
- Thorsen, M.; Di, Y.; Tängemo, C.; Morillas, M.; Ahmadpour, D.; van der Does, C.; Wagner, A.; Johansson, E.; Boman, J.; Posas, F.; et al. The MAPK Hog1p modulates Fps1p-dependent arsenite uptake and tolerance in yeast. Mol. Biol. Cell 2006, 17, 4400–4410. [Google Scholar]
- Beese, S.E.; Negishi, T.; Levin, D.E. Identification of positive regulators of the yeast Fps1 glycerol channel. PLoS Genet 2009, 5, e1000738. [Google Scholar]
- Bienert, G.P.; Thorsen, M.; Schüssler, M.D.; Nilsson, H.R.; Wagner, A.; Tamás, M.J.; Jahn, T.P. A subgroup of plant aquaporins facilitate the bidirectional diffusion of As(OH)3 and Sb(OH)3 across membranes. BMC Biol 2008, 6, 26. [Google Scholar]
- Isayenkov, S.V.; Maathuis, F.J.M. The Arabidopsis thaliana aquaglyceroporin AtNIP7;1 is a pathway for arsenite uptake. FEBS Lett 2008, 582, 1625–1628. [Google Scholar]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Xu, X.Y.; Su, Y.H.; McGrath, S.P.; Zhao, F.J. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad. Sci. USA 2008, 105, 9931–9935. [Google Scholar]
- Maurel, C. Plant aquaporins: novel functions and regulation properties. FEBS Lett 2007, 581, 2227–2236. [Google Scholar]
- Takano, J.; Wada, M.; Ludewig, U.; Schaaf, G.; von Wirén, N.; Fujiwara, T. The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron uptake and plant development under boron limitation. Plant Cell 2006, 18, 1498–1509. [Google Scholar]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar]
- Kamiya, T.; Tanaka, M.; Mitani, N.; Ma, J.F.; Maeshima, M.; Fujiwara, T. NIP1;1, an aquaporin homolog, determines the arsenite sensitivity of Arabidopsis thaliana. J. Biol. Chem 2009, 284, 2114–2120. [Google Scholar]
- Li, R.Y.; Ago, Y.; Liu, W.J.; Mitani, N.; Feldmann, J.; McGrath, S.P.; Ma, J.F.; Zhao, F.J. The rice aquaporin Lsi1 mediates uptake of methylated arsenic species. Plant Physiol 2009, 150, 2071–2080. [Google Scholar]
- Kamiya, T.; Fujiwara, T. Arabidopsis NIP1;1 transports antimonite and determines antimonite sensitivity. Plant Cell Physiol 2009, 50, 1977–1981. [Google Scholar]
- Mosa, K.A.; Kumar, K.; Chhikara, S.; McDermott, J.; Liu, Z.; Musante, C.; White, J.C.; Dhankher, O.P. Members of rice plasma membrane intrinsic proteins subfamily are involved in arsenite permeability and tolerance in plants. Transgenic Res 2012. [Google Scholar] [CrossRef]
- Gourbal, B.; Sonuc, N.; Bhattacharjee, H.; Legare, D.; Sundar, S.; Ouellette, M.; Rosen, B.P.; Mukhopadhyay, R. Drug uptake and modulation of drug resistance in Leishmania by an aquaglyceroporin. J. Biol. Chem 2004, 279, 31010–31017. [Google Scholar]
- Marquis, N.; Gourbal, B.; Rosen, B.P.; Mukhopadhyay, R.; Ouellette, M. Modulation in aquaglyceroporin AQP1 gene transcript levels in drug-resistant Leishmania. Mol. Microbiol 2005, 57, 1690–1699. [Google Scholar]
- Mandal, S.; Maharjan, M.; Singh, S.; Chatterjee, M.; Madhubala, R. Assessing aquaglyceroporin gene status and expression profile in antimony-susceptible and -resistant clinical isolates of Leishmania donovani from India. J. Antimicrob. Chemother 2010, 65, 496–507. [Google Scholar]
- Ishibashi, K.; Hara, S.; Kondo, S. Aquaporin water channels in mammals. Clin. Exp. Nephrol 2009, 13, 107–117. [Google Scholar]
- Liu, Z.; Shen, J.; Carbrey, J.M.; Mukhopadhyay, R.; Agre, P.; Rosen, B.P. Arsenite transport by mammalian aquaglyceroporins AQP7 and AQP9. Proc. Natl. Acad. Sci. USA 2002, 99, 6053–6058. [Google Scholar]
- Liu, Z.; Carbrey, J.M.; Agre, P.; Rosen, B.P. Arsenic trioxide uptake by human and rat aquaglyceroporins. Biochem. Biophys. Res. Commun 2004, 316, 1178–1185. [Google Scholar]
- Bhattacharjee, H.; Carbrey, J.; Rosen, B.P.; Mukhopadhyay, R. Drug uptake and pharmacological modulation of drug sensitivity in leukemia by AQP9. Biochem. Biophys. Res. Commun 2004, 322, 836–841. [Google Scholar]
- Leung, J.; Pang, A.; Yuen, W.H.; Kwong, Y.L.; Tse, E.W. Relationship of expression of aquaglyceroporin 9 with arsenic uptake and sensitivity in leukemia cells. Blood 2007, 109, 740–746. [Google Scholar]
- Lee, T.C.; Ho, I.C.; Lu, W.J.; Huang, J.D. Enhanced expression of multidrug resistance-associated protein 2 and reduced expression of aquaglyceroporin 3 in an arsenic-resistant human cell line. J. Biol. Chem 2006, 281, 18401–18407. [Google Scholar]
- Calatayud, M.; Barrios, J.A.; Vélez, D.; Devesa, V. In vitro study of transporters involved in intestinal absorption of inorganic arsenic. Chem. Res. Toxicol 2012, 25, 446–453. [Google Scholar]
- Hamdi, M.; Sanchez, M.A.; Beene, L.C.; Liu, Q.; Landfear, S.M.; Rosen, B.P.; Liu, Z. Arsenic transport by zebrafish aquaglyceroporins. BMC Mol. Biol 2009, 10, 104. [Google Scholar]
- Liu, Z.; Styblo, M.; Rosen, B.P. Methylarsonous acid transport by aquaglyceroporins. Environ. Health Perspect 2006, 114, 527–531. [Google Scholar]
- McDermott, J.R.; Jiang, X.; Beene, L.C.; Rosen, B.P.; Liu, Z. Pentavalent methylated arsenicals are substrates of human AQP9. Biometals 2010, 23, 119–127. [Google Scholar]
- Liu, Z.; Boles, E.; Rosen, B.P. Arsenic trioxide uptake by hexose permeases in Saccharomyces cerevisiae. J. Biol. Chem 2004, 279, 17312–17318. [Google Scholar]
- Liu, Z.; Sanchez, M.A.; Jiang, X.; Boles, E.; Landfear, S.M.; Rosen, B.P. Mammalian glucose permease GLUT1 facilitates transport of arsenic trioxide and methylarsonous acid. Biochem. Biophys. Res. Commun 2006, 351, 424–430. [Google Scholar]
- Jiang, X.; McDermott, J.R.; Ajees, A.A.; Rosen, B.P.; Liu, Z. Trivalent arsenicals and glucose use different translocation pathways in mammalian GLUT1. Metallomics 2010, 2, 211–219. [Google Scholar]
- Drobná, Z.; Walton, F.S.; Paul, D.S.; Xing, W.; Thomas, D.J.; Stýblo, M. Metabolism of arsenic in human liver: the role of membrane transporters. Arch. Toxicol 2010, 84, 3–16. [Google Scholar]
- Lu, W.; Tamai, I.; Nezu, J.; Lai, M.; Huang, J. Organic anion transportin polypeptide-C mediates arsenic uptake in HEK-293 cells. J. Biomed. Sci 2006, 13, 525–353. [Google Scholar]
- Hagenbuch, B.; Meier, P.J. Organic anion transporting polypeptides of the OATP/SLC21 family: phylogenetic classification as OATP/SLCO superfamily, new nomenclature and molecular/functional properties. Pflugers Arch 2004, 447, 653–665. [Google Scholar]
- Mukhopadhyay, R.; Shi, J.; Rosen, B.P. Purification and characterization of Acr2p, the Saccharomyces cerevisiae arsenate reductase. J. Biol. Chem 2000, 275, 21149–21157. [Google Scholar]
- Zhou, Y.; Messier, N.; Ouellette, M.; Rosen, B.P.; Mukhopadhyay, R. Leishmania major LmACR2 is a pentavalent antimony reductase that confers sensitivity to the drug pentostam. J. Biol. Chem 2004, 279, 37445–37451. [Google Scholar]
- Ellis, D.R.; Gumaelius, L.; Indriolo, E.; Pickering, I.J.; Banks, J.A.; Salt, D.E. A novel arsenate reductase from the arsenic hyperaccumulating fern Pteris vittata. Plant Physiol 2006, 141, 1544–1554. [Google Scholar]
- Duan, G.; Zhou, Y.; Tong, Y.P.; Mukhopadhyay, R.; Rosen, B.P.; Zhu, Y.G. A CDC25 homologue from rice functions as an arsenate reductase. New Phytol 2007, 174, 311–321. [Google Scholar]
- Bhattacharjee, H.; Sheng, J.; Ajees, A.A.; Mukhopadhyay, R.; Rosen, B.P. Adventitious arsenate reductase activity of the catalytic domain of the human Cdc25B and Cdc25C phosphatases. Biochemistry 2010, 49, 802–809. [Google Scholar]
- Yang, H.C.; Cheng, J.; Finan, T.M.; Rosen, B.P.; Bhattacharjee, H. Novel pathway for arsenic detoxification in the legume symbiont Sinorhizobium meliloti. J. Bacteriol 2005, 187, 6991–6997. [Google Scholar]
- Zhao, F.J.; Ago, Y.; Mitani, N.; Li, R.Y.; Su, Y.H.; Yamaji, N.; McGrath, S.P.; Ma, J.F. The role of the rice aquaporin Lsi1 in arsenite efflux from roots. New Phytol 2010, 186, 392–399. [Google Scholar]
- Maciaszczyk-Dziubinska, E.; Migdal, I.; Migocka, M.; Bocer, T.; Wysocki, R. The yeast aquaglyceroporin Fps1p is a bidirectional arsenite channel. FEBS Lett 2010, 584, 726–732. [Google Scholar]
- Jacobson, T.; Thorsen, M.; Vooijs, R.; Schat, H.; Tamás, M.J. Yeast cells export glutathione as an extracellular defence mechanism. In FEBS J, Presented at 35th Congress of the Federation of European Biochemical Societies, Gothenburg, Sweden, 26 June–1 July, 2010; 2010; 277, p. 206. [Google Scholar]
- Dhaoui, M.; Auchère, F.; Blaiseau, P.L.; Lesuisse, E.; Landoulsi, A.; Camadro, J.M.; Haguenauer-Tsapis, R.; Belgareh-Touzé, N. Gex1 is a yeast glutathione exchanger that interferes with pH and redox homeostasis. Mol. Biol. Cell 2011, 22, 2054–2067. [Google Scholar]
- Thomas, D.J.; Li, J.; Waters, S.B.; Xing, W.; Adair, B.M.; Drobna, Z.; Devesa, V.; Styblo, M. Arsenic (+3 oxidation state) methyltransferase and methylation of arsenicals. Exp. Biol. Med 2007, 232, 3–11. [Google Scholar]
- Carbrey, JM.; Song, L.; Zhou, Y.; Yoshinaga, M.; Rojek, A.; Wang, Y.; Liu, Y.; Lujan, H.L.; DiCarlo, S.E.; Nielsen, S.; et al. Reduced arsenic clearance and increased toxicity in aquaglyceroporin-9-null mice. Proc. Natl. Acad. Sci. USA 2009, 106, 15956–15960. [Google Scholar]
- Rosen, B.P.; Tamás, M.J. Arsenic transport in prokaryotes and eukaryotic microbes. Adv. Exp. Med. Biol 2010, 679, 47–55. [Google Scholar]
- Fu, H.L.; Meng, Y.; Ordóñez, E.; Villadangos, A.F.; Bhattacharjee, H.; Gil, J.A.; Mateos, L.M.; Rosen, B.P. Properties of arsenite efflux permeases (Acr3) from Alkaliphilus metalliredigens and Corynebacterium glutamicum. J. Biol. Chem 2009, 284, 19887–19895. [Google Scholar]
- Indriolo, E.; Na, G.; Ellis, D.; Salt, D.E.; Banks, J.A. A vacuolar arsenite transporter necessary for arsenic tolerance in the arsenic hyperaccumulating fern Pteris vittata is missing in flowering plants. Plant Cell 2010, 22, 2045–2057. [Google Scholar]
- Wu, J.; Tisa, L.S.; Rosen, B.P. Membrane topology of the ArsB protein, the membrane subunit of an anion-translocating ATPase. J. Biol. Chem 1992, 267, 12570–12576. [Google Scholar]
- Meng, Y.L.; Liu, Z.; Rosen, B.P. As(III) and Sb(III) uptake by GlpF and efflux by ArsB in Escherichia coli. J. Biol. Chem 2004, 279, 18334–18341. [Google Scholar]
- Mansour, N.M.; Sawhney, M.; Tamang, D.G.; Vogl, C.; Saier, M.H., Jr. The bile/arsenite/riboflavin transporter (BART) superfamily. FEBS J 2007, 274, 612–629. [Google Scholar]
- Bobrowicz, P.; Wysocki, R.; Owsianik, G.; Goffeau, A.; Ulaszewski, S. Isolation of three contiguous genes, ACR1, ACR2 and ACR3, involved in resistance to arsenic compounds in the yeast Saccharomyces cerevisiae. Yeast 1997, 13, 819–828. [Google Scholar]
- Wysocki, R.; Bobrowicz, P.; Ulaszewski, S. The Saccharomyces cerevisiae ACR3 gene encodes a putative membrane protein involved in arsenite transport. J. Biol. Chem 1997, 272, 30061–30066. [Google Scholar]
- Ghosh, M.; Shen, J.; Rosen, B.P. Pathways of As(III) detoxification in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1999, 96, 5001–5006. [Google Scholar]
- Maciaszczyk-Dziubinska, E.; Wawrzycka, D.; Sloma, E.; Migocka, M.; Wysocki, R. The yeast permease Acr3p is a dual arsenite and antimonite plasma membrane transporter. Biochim. Biophys. Acta 2010, 1798, 2170–2175. [Google Scholar]
- Maciaszczyk-Dziubinska, E.; Migocka, M.; Wysocki, R. Acr3p is a plasma membrane antiporter that catalyzes As(III)/H+ and Sb(III)/H+ exchange in Saccharomyces cerevisiae. Biochim. Biophys. Acta 2011, 1808, 1855–1859. [Google Scholar]
- Sato, T.; Kobayashi, Y. The ars operon in the skin element of Bacillus subtilis confers resistance to arsenate and arsenite. J. Bacteriol 1998, 180, 1655–1661. [Google Scholar]
- Villadangos, A.F.; Fu, H.L.; Gil, J.A.; Messens, J.; Rosen, B.P.; Mateos, L.M. Efflux permease CgAcr3-1 of Corynebacterium glutamicum is an arsenite-specific antiporter. J. Biol. Chem 2012, 287, 723–735. [Google Scholar]
- Maciaszczyk, E.; Wysocki, R.; Golik, P.; Lazowska, J.; Ulaszewski, S. Arsenical resistance genes in Saccharomyces douglasii and other yeast species undergo rapid evolution involving genomic rearrangements and duplications. FEMS Yeast Res 2004, 4, 821–832. [Google Scholar]
- López-Maury, L.; Florencio, F.J.; Reyes, J.C. Arsenic sensing and resistance system in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol 2003, 185, 5363–5371. [Google Scholar]
- Xia, X.; Postis, V.L.; Rahman, M.; Wright, G.S.; Roach, P.C.; Deacon, S.E.; Ingram, J.C.; Henderson, P.J.; Findlay, J.B.; Phillips, S.E.; et al. Investigation of the structure and function of a Shewanella oneidensis arsenical-resistance family transporter. Mol. Membr. Biol 2008, 25, 691–705. [Google Scholar]
- Aaltonen, E.K.; Silow, M. Transmembrane topology of the Acr3 family arsenite transporter from Bacillus subtilis. Biochim. Biophys. Acta 2008, 1778, 963–973. [Google Scholar]
- Maciaszczyk-Dziubinska, E.; Wawrzycka, D.; Migocka, M.; Wysocki, R. Mutational analysis of the yeast As(III)/Sb(III) transporter Acr3; University of Wrocław: Wrocław, Poland, Unpublished work; 2012. [Google Scholar]
- Wysocki, R.; Fortier, P.K.; Maciaszczyk, E.; Thorsen, M.; Leduc, A.; Odhagen, A.; Owsianik, G.; Ulaszewski, S.; Ramotar, D.; Tamás, M.J. Transcriptional activation of metalloid tolerance genes in Saccharomyces cerevisiae requires the AP-1-like proteins Yap1p and Yap8p. Mol. Biol. Cell 2004, 15, 2049–2060. [Google Scholar]
- Di, Y.; Tamás, M.J. Regulation of the arsenic-responsive transcription factor Yap8p involves the ubiquitin-proteasome pathway. J. Cell Sci 2007, 120, 256–264. [Google Scholar]
- Ilina, Y.; Sloma, E.; Maciaszczyk-Dziubinska, E.; Novotny, M.; Thorsen, M.; Wysocki, R.; Tamás, M.J. Characterization of the DNA binding motif of the arsenic-responsive transcription factor Yap8p. Biochem. J 2008, 415, 467–475. [Google Scholar]
- Leslie, E.M. Arsenic-glutathione conjugate transport by the human multidrug resistance proteins (MRPs/ABCCs). J. Inorg. Biochem 2011. [Google Scholar] [CrossRef]
- Slot, A.J.; Molinski, S.V.; Cole, S.P. Mammalian multidrug-resistance proteins (MRPs). Essays Biochem 2011, 50, 179–207. [Google Scholar]
- Sharom, F.J. The P-glycoprotein multidrug transporter. Essays Biochem 2011, 50, 161–178. [Google Scholar]
- Wawrzycka, D.; Sobczak, I.; Bartosz, G.; Bocer, T.; Ułaszewski, S.; Goffeau, A. Vmr1p is a novel vacuolar multidrug resistance ABC transporter in Saccharomyces cerevisiae. FEMS Yeast Res 2010, 10, 828–838. [Google Scholar]
- Paumi, C.M.; Chuk, M.; Snider, J.; Stagljar, I.; Michaelis, S. ABC transporters in Saccharomyces cerevisiae and their interactors: new technology advances the biology of the ABCC (MRP) subfamily. Microbiol. Mol. Biol. Rev 2009, 73, 577–593. [Google Scholar]
- Vatamaniuk, O.K.; Bucher, E.A.; Ward, J.T.; Rea, P.A. Wormstake the phyto out of phytochelatins. Trends Biotechnol 2002, 20, 61–64. [Google Scholar]
- Vatamaniuk, O.K.; Mari, S.; Lu, Y.P.; Rea, P.A. Mechanism of heavy metal ion activation of phytochelatin (PC) synthase: blocked thiols are sufficient for PC synthase-catalyzed transpeptidation of glutathione and related thiol peptides. J. Biol. Chem 2000, 275, 31451–31459. [Google Scholar]
- Ortiz, D.F.; Ruscitti, T.; McCue, K.F.; Ow, D.W. Transport of metal-binding peptides by HMT1, a fission yeast ABC-type vacuolar membrane protein. J. Biol. Chem 1995, 270, 4721–4728. [Google Scholar]
- Prévéral, S.; Gayet, L.; Moldes, C.; Hoffmann, J.; Mounicou, S.; Gruet, A.; Reynaud, F.; Lobinski, R.; Verbavatz, J.M.; Vavasseur, A.; et al. A common highly conserved cadmium detoxification mechanism from bacteria to humans: heavy metal tolerance conferred by the ATP-binding cassette (ABC) transporter SpHMT1 requires glutathione but not metal-chelating phytochelatin peptides. J. Biol. Chem 2009, 284, 4936–4943. [Google Scholar]
- Sooksa-Nguan, T.; Yakubov, B.; Kozlovskyy, V.I.; Barkume, C.M.; Howe, K.J.; Thannhauser, T.W.; Rutzke, M.A.; Hart, J.J.; Kochian, L.V.; Rea, P.A.; et al. Drosophila ABC transporter, DmHMT-1, confers tolerance to cadmium. DmHMT-1 and its yeast homolog, SpHMT-1, are not essential for vacuolar phytochelatin sequestration. J. Biol. Chem 2009, 284, 354–362. [Google Scholar]
- Schwartz, M.S.; Benci, J.L.; Selote, D.S.; Sharma, A.K.; Chen, A.G.; Dang, H.; Fares, H.; Vatamaniuk, O.K. Detoxification of multiple heavy metals by a half-molecule ABC transporter, HMT-1, and coelomocytes of Caenorhabditis elegans. PLoS One 2010, 5, e9564. [Google Scholar]
- Song, W.Y.; Park, J.; Mendoza-Cózatl, D.G.; Suter-Grotemeyer, M.; Shim, D.; Hörtensteiner, S.; Geisler, M.; Weder, B.; Rea, P.A.; Rentsch, D.; et al. Arsenic tolerance in Arabidopsis is mediated by two ABCC-type phytochelatin transporters. Proc. Natl. Acad. Sci. USA 2010, 107, 21187–21192. [Google Scholar]
- Mendoza-Cózatl, D.G.; Zhai, Z.; Jobe, T.O.; Akmakjian, G.Z.; Song, W.Y.; Limbo, O.; Russell, M.R.; Kozlovskyy, V.I.; Martinoia, E.; Vatamaniuk, O.K.; et al. Tonoplast-localized Abc2 transporter mediates phytochelatin accumulation in vacuoles and confers cadmium tolerance. J. Biol. Chem 2010, 285, 40416–40426. [Google Scholar]
- Callahan, H.L.; Beverley, S.M. Heavy metal resistance: a new role for P-glycoproteins in Leishmania. Biol. Chem 1991, 266, 18427–18430. [Google Scholar]
- Ouellette, M.; Legare, D.; Haimeur, A.; Grondin, K.; Roy, G.; Brochu, C.; Papadopoulou, B. ABC transporters in Leishmania and their role in drug resistance. Drug Resist Updat 1998, 1, 43–48. [Google Scholar]
- Légaré, D.; Richard, D.; Mukhopadhyay, R.; Stierhof, Y.D.; Rosen, B.P.; Haimeur, A.; Papadopoulou, B.; Ouellette, M. The Leishmania ABC protein PGPA is an intracellular metal-thiol transporter ATPase. J. Biol. Chem 2001, 276, 26301–26307. [Google Scholar]
- Coelho, A.C.; Beverley, S.M.; Cotrim, P.C. Functional genetic identification of PRP1, an ABC transporter superfamily member conferring pentamidine resistance in Leishmania major. Mol. Biochem. Parasitol 2003, 130, 83–90. [Google Scholar]
- Coelho, A.C.; Yamashiro-Kanashiro, E.H.; Bastos, S.F.; Mortara, R.A.; Cotrim, P.C. Intracellular location of the ABC transporter PRP1 related to pentamidine resistance in Leishmania major. Mol. Biochem. Parasitol 2006, 150, 378–383. [Google Scholar]
- Broeks, A.; Gerrard, B.; Allikmets, R.; Dean, M.; Plasterk, R.H. Homologues of the human multidrug resistance genes MRP and MDR contribute to heavy metal resistance in the soil nematode Caenorhabditis elegans. EMBO J 1996, 15, 6132–6143. [Google Scholar]
- Long, Y.; Li, Q.; Wang, Y.; Cui, Z. MRP proteins as potential mediators of heavy metal resistance in zebrafish cells. Comp. Biochem. Physiol. C Toxicol. Pharmacol 2011, 153, 310–317. [Google Scholar]
- Long, Y.; Li, Q.; Cui, Z. Molecular analysis and heavy metal detoxification of ABCC1/MRP1 in zebrafish. Mol. Biol. Rep 2011, 38, 1703–1711. [Google Scholar]
- Chin, K.V.; Tanaka, S.; Darlington, G.; Pastan, I.; Gottesman, M.M. Heat shock and arsenite increase expression of the multidrug resistance (MDR1) gene in human renal carcinoma cells. J. Biol. Chem 1990, 265, 221–226. [Google Scholar]
- Liu, J.; Chen, H.; Miller, D.S.; Saavedra, J.E.; Keefer, L.K.; Johnson, D.R.; Klaassen, C.D.; Waalkes, M.P. Overexpression of glutathione S-transferase II and multidrug resistance transport proteins is associated with acquired tolerance to inorganic arsenic. Mol. Pharmacol 2001, 60, 302–309. [Google Scholar]
- Liu, J.; Liu, Y.; Powell, D.A.; Waalkes, M.P.; Klaassen, C.D. Multidrug-resistance mdr1a/1b double knockout mice are more sensitive than wild type mice to acute arsenic toxicity, with higher arsenic accumulation in tissues. Toxicology 2002, 170, 55–62. [Google Scholar]
- Xie, Y.; Liu, J.; Liu, Y.; Klaassen, C.D.; Waalkes, M.P. Toxicokinetic and genomic analysis of chronic arsenic exposure in multidrug-resistance mdr1a/1b(-/-) double knockout mice. Mol. Cell. Biochem 2004, 255, 11–18. [Google Scholar]
- Cole, S.P.; Sparks, K.E.; Fraser, K.; Loe, D.W.; Grant, C.E.; Wilson, G.M.; Deeley, R.G. Pharmacological characterization of multidrug resistant MRP-transfected human tumor cells. Cancer Res 1994, 54, 5902–5910. [Google Scholar]
- Vernhet, L.; Courtois, A.; Allain, N.; Payen, L.; Anger, J.P.; Guillouzo, A.; Fardel, O. Overexpression of the multidrug resistance-associated protein (MRP1) in human heavy metal-selected tumor cells. FEBS Lett 1999, 443, 321–325. [Google Scholar]
- Vernhet, L.; Allain, N.; Bardiau, C.; Anger, J.P.; Fardel, O. Differential sensitivities of MRP1-overexpressing lung tumor cells to cytotoxic metals. Toxicology 2000, 142, 127–134. [Google Scholar]
- Vernhet, L.; Allain, N.; Payen, L.; Anger, J.P.; Guillouzo, A.; Fardel, O. Resistance of human multidrug resistance-associated protein 1-overexpressing lung tumor cells to the anticancer drug arsenic trioxide. Biochem. Pharmacol 2001, 6, 1387–1391. [Google Scholar]
- Rappa, G.; Lorico, A.; Flavell, R.A.; Sartorelli, A.C. Evidence that the multidrug resistance protein (MRP) functions as a co-transporter of glutathione and natural product toxins. Cancer Res 1997, 57, 5232–5237. [Google Scholar]
- Allen, J.D.; Brinkhuis, R.F.; van Deemter, L.; Wijnholds, J.; Schinkel, A.H. Extensive contribution of the multidrug transporters P-glycoprotein and Mrp1 to basal drug resistance. Cancer Res 2000, 60, 5761–5766. [Google Scholar]
- Leslie, E.M.; Haimeur, A.; Waalkes, M.P. Arsenic transport by the human multidrug resistance protein 1 (MRP1/ABCC1). Evidence that a tri-glutathione conjugate is required. J. Biol. Chem 2004, 279, 32700–32708. [Google Scholar]
- Carew, M.W.; Naranmandura, H.; Shukalek, C.B.; Le, X.C.; Leslie, E.M. Monomethylarsenic diglutathione transport by the human multidrug resistance protein 1 (MRP1/ABCC1). Drug Metab. Dispos 2011, 39, 2298–22304. [Google Scholar]
- Kala, S.V.; Neely, M.W.; Kala, G.; Prater, C.I.; Atwood, D.W.; Rice, J.S.; Lieberman, M.W. The MRP2/cMOAT transporter and arsenic-glutathione complex formation are required for biliary excretion of arsenic. J. Biol. Chem 2000, 275, 33404–33408. [Google Scholar]
- Carew, M.W.; Leslie, E.M. Selenium-dependent and -independent transport of arsenic by the human multidrug resistance protein 2 (MRP2/ABCC2): implications for the mutual detoxification of arsenic and selenium. Carcinogenesis 2010, 31, 1450–1455. [Google Scholar]
- Duan, G.; Kamiya, T.; Ishikawa, S.; Arao, T.; Fujiwara, T. Expressing ScACR3 in rice enhanced arsenite efflux and reduced arsenic accumulation in rice grains. Plant Cell. Physiol 2012, 53, 154–163. [Google Scholar]
- Guo, J.; Xu, W.; Ma, M. The assembly of metals chelation by thiols and vacuolar compartmentalization conferred increased tolerance to and accumulation of cadmium and arsenic in transgenic Arabidopsis thaliana. J. Hazard. Mater 2012, 199–200, 309–313. [Google Scholar]
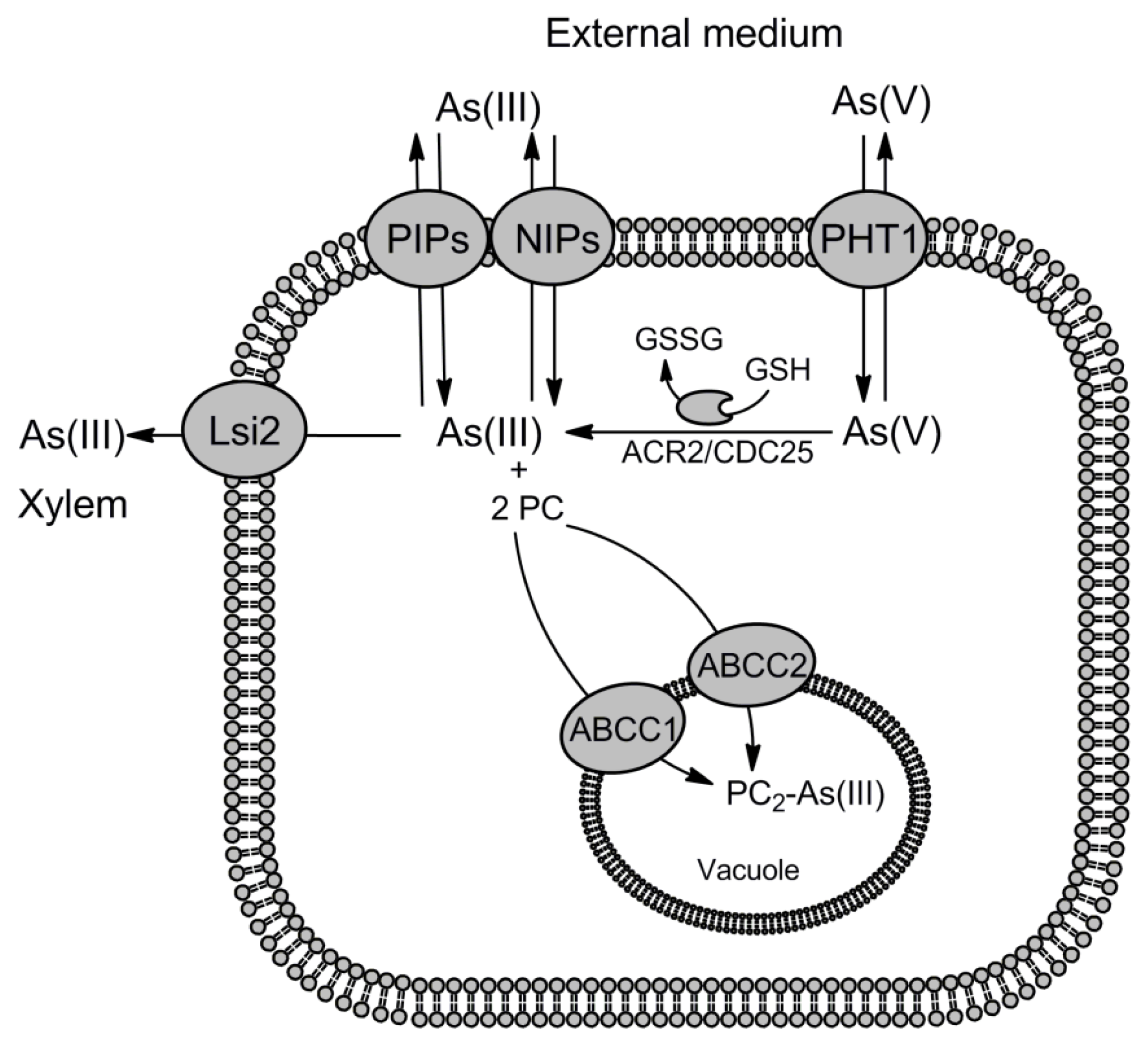
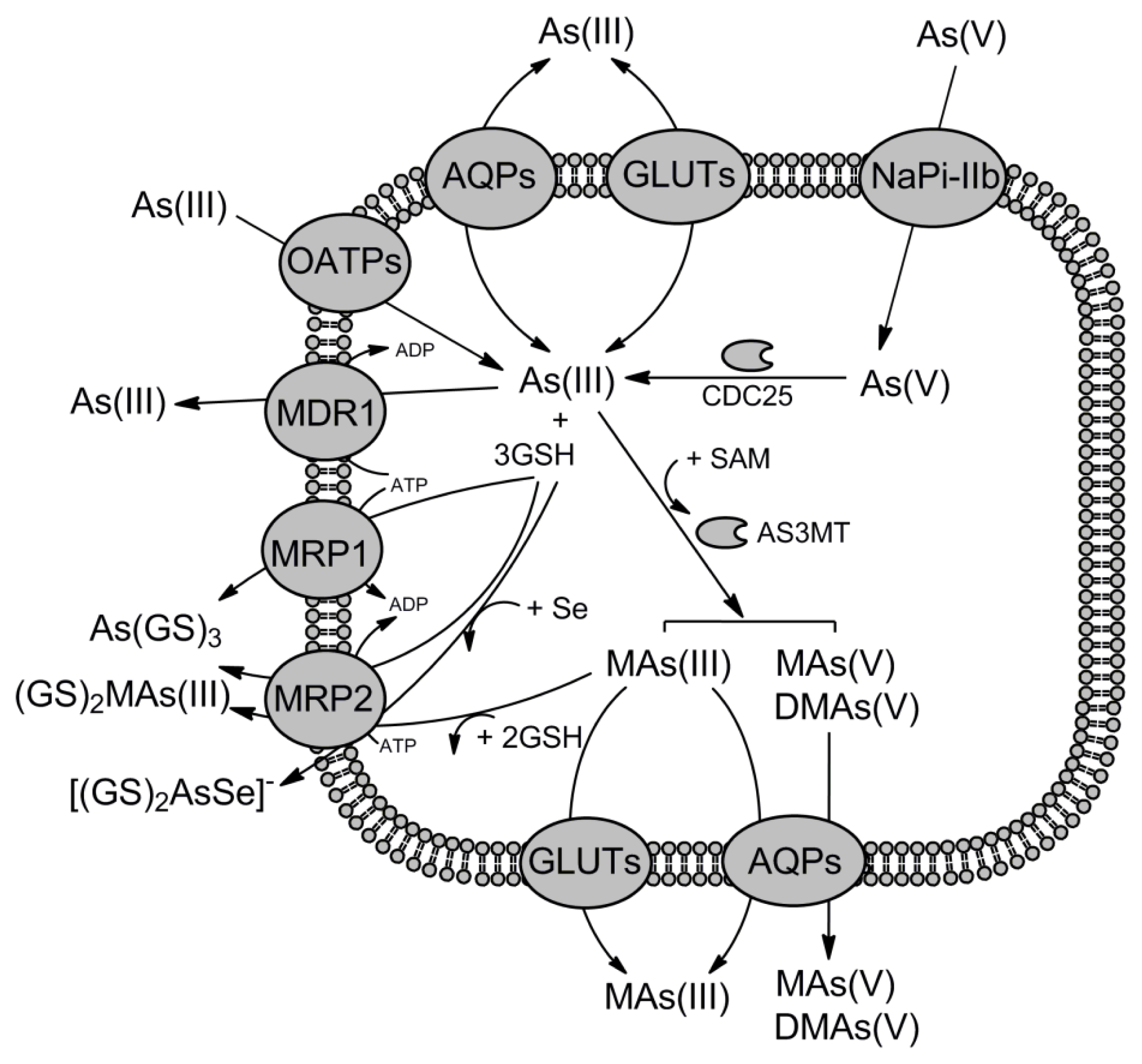
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maciaszczyk-Dziubinska, E.; Wawrzycka, D.; Wysocki, R. Arsenic and Antimony Transporters in Eukaryotes. Int. J. Mol. Sci. 2012, 13, 3527-3548. https://doi.org/10.3390/ijms13033527
Maciaszczyk-Dziubinska E, Wawrzycka D, Wysocki R. Arsenic and Antimony Transporters in Eukaryotes. International Journal of Molecular Sciences. 2012; 13(3):3527-3548. https://doi.org/10.3390/ijms13033527
Chicago/Turabian StyleMaciaszczyk-Dziubinska, Ewa, Donata Wawrzycka, and Robert Wysocki. 2012. "Arsenic and Antimony Transporters in Eukaryotes" International Journal of Molecular Sciences 13, no. 3: 3527-3548. https://doi.org/10.3390/ijms13033527
APA StyleMaciaszczyk-Dziubinska, E., Wawrzycka, D., & Wysocki, R. (2012). Arsenic and Antimony Transporters in Eukaryotes. International Journal of Molecular Sciences, 13(3), 3527-3548. https://doi.org/10.3390/ijms13033527