Abstract
Aging is associated with disturbances in iron metabolism and storage. During the last decade, remarkable progress has been made toward understanding their cellular and molecular mechanisms in aging and age-associated diseases using both cultured cells and animal models. The field has moved beyond descriptive studies to potential intervention studies focusing on iron chelation and removal. However, some findings remain controversial and inconsistent. This review summarizes important features of iron dyshomeostasis in aging research with a particular emphasis on current knowledge of the mechanisms underlying age-associated disorders in rodent models.
1. Introduction
Iron is an essential nutrient. Disturbances of iron metabolism may have deleterious consequences in severe pathological conditions such as cardiovascular diseases [1–3], diabetes [4], cancer [5–7] and neurodegenerative diseases [8–12]. It has been widely documented that aging is associated with dyshomeostasis of iron metabolism and regulation in both rodents [13–25] (Table 1) and humans [26–32]. The elderly are more prone to becoming anemic [33–37], which adversely affects muscle strength [38], physical performance [39], cognition [40] and longevity [41]. In contrast, age-related iron overload is also increasingly being recognized as a public health concern [42–45]. Despite the prevalence and adverse health effects associated with these disorders, the mechanisms are still not well defined and many questions remain to be answered [2,46,47].

Table 1.
Summary of studies reporting organ-specific changes in iron content with age in rodents.
2. Impaired Iron Status with Age in Rodent Models
2.1. Organ-Specific Changes in Iron Content with Age
During the last decade, a number of studies have documented age-related iron accumulation in rodents (Table 1). One of the earliest studies (by Massie et al. [20]) revealed age-related changes in iron content in young (1.5–7 months), middle-aged (21 months) and aged (30 months) male C57BL/6J mice. They showed that total iron concentrations were significantly elevated in the liver, heart, kidney and brain of aged animals. In further support of the finding of iron dyshomeostasis in aging, Sohal et al. [24] reported that there was an age-associated increase in non-heme iron levels in liver, kidney, brain and heart, which, however, is independent of the increases in redox-active iron determined using bleomycin-detectable iron assay. In addition, the observation in the same study that life-long 40% caloric restriction had no effect on iron levels in heart and brain and even exacerbated iron accumulation in liver and kidney does not support the hypothesis that labile iron plays an essential role underlying the age-associated increase in oxidative damage. On the contrary, a successful attempt to ameliorate age-related iron accumulation by life-long 40% caloric restriction was published by Cook and Yu [19] in 1998. The results of their study in male Fischer 344 rats showed a remarkable age-related increase in non-heme iron levels in liver, kidney and brain of animals fed ad libitum. Their finding that caloric restriction markedly mitigated iron accumulation in multiple tissue systems of aged animals as well as our recent study [17] suggests that caloric restriction beneficially modulates iron dyshomeostasis.
Since iron accumulation is widely accepted as a feature of the aging process particularly in post-mitotic tissues by emerging research in the intervening decade [14–18,21,22,25], a substantial research effort has been directed at exploring potential iron chelation therapies. Recently, deferiprone and deferasirox emerged as promising orally active iron-sequestering agents [23,48,49]. Arvapalli et al. [21] reported that deferasirox, administrated at a dose of 100 mg/kg body weight on alternate days for 6 months, was effective in reducing total iron levels in the heart and liver as well as attenuating cardiomyocyte apoptosis in 27-month-old Fischer 344 x Brown Norway rats. However, limited information is provided in the same study to warrant that the use of chelator did not exacerbate the low serum ferritin usually observed in aged animals. A non-toxic iron chelator or potential treatment strategy that locally removes excess iron in particular tissues without affecting systemic iron utilization, storage and transport, may represent an ideal therapeutic intervention.
In contrast to the above-mentioned studies, Ahluwalia et al. [13] showed that total non-heme iron levels in the liver, spleen, and bone marrow of Lewis rats declined with age. The conflicting study findings may stem from several factors involved in rodent aging research, such as strain, species, diets, and life stages of animals.
2.2. Life Stage, Species, Sex and Strain Differences across Studies
Though research scientists have made considerable progress in defining rodent life stages across species and strains, crucial definitional problems remain unsolved. It could be considered as a central challenge in investigating age-associated changes in iron homeostasis and metabolism primarily because the average lifespan varies greatly depending on sex, strain, and breeding system.
Outbred strains, such as Sprague Dawley and Wistar rats, have been widely used to investigate iron homeostasis and metabolism in aging research, while C57BL/6 mice, Fischer 344 rats, and Lewis rats as inbred strains are excellent models as well. Recently, Fischer 344 x Brown Norway rat, a F1 hybrid strain, has been proposed as a potential model for aging since it most closely reproduces healthy aging in humans [55,56]. A comparative study on the muscle mass and contractile properties between Fischer 344 x Brown Norway and Fischer 344 rats by Rice et al. [55] indicated that there were ageassociated decreases in both of the two sub-populations of muscle fibers in Fischer 344 x Brown Norway rats, suggesting that the F1 hybrid strain is a better model of sarcopenia than Fischer 344. Besides the differences in age-associated physiological or pathological alterations, outbred and F1 hybrid animals exhibit hybrid vigor with long lifespans. The median survival ages for male and female Fischer 344 rats are 24 and 26 months, respectively; however it extends to 34 months for male and 30 months for female Fischer 344 x Brown Norway rats [50]. In the basic science of aging, two or three age cohorts were commonly selected and referred to representative life stages as young and old animals or young, middle-aged, and old animals, which may dramatically limit the power of investigations and the universality of conclusions. If indeed iron is a contributing factor in the aging process, a significant alterations in iron levels or metabolism between young and aged animals will be detectable at the point that iron dyshomeostasis has occurred in the study population and remains relatively stable. Given the fact that aged rats and mice past the 25% survival age are more prone to underlying age-associated diseases and are not a good research model of healthy aging for most purposes, the cut-off age at which point iron status has substantially altered while the incidence of pathologies is relatively low is crucial in aging research using rodent models.
Table 1 lists the animal studies reporting organ-specific changes in iron content with age in rodents. The median survival age of each strain has been employed to estimate and compare animal life stages across studies. Aged cohorts at their median survival ages have been included in these studies except for the one provided by Takeda et al. [51], who reported an age-related iron accumulation using young (3-week-old) and mature (6-month-old) Wistar female rats. However, other investigators using male Fischer 344 [19] and male Fischer 344 x Brown Norway rats [17] indicated that iron levels in liver remained unchanged until late middle age. Recently, Hahn et al. [13,57] reported age- and sex-dependent changes in tissue iron levels among C57BL/6, DBA/2J, and BALB/c mouse strains and further confirmed that there were age-dependent and sex-specific changes in mouse tissue iron by strain. Taken together, these observations suggest the onset of impaired iron status in rodent models highly depends on sex and strain.
2.3. Age-Associated Decrease in Heme Iron Levels vs. Increase in Non-Heme Iron Levels
Measurements of iron levels in rodent models usually fall into two categories: total iron determined by spectrometry techniques, such as atomic absorption spectroscopy [20] or inductively coupled plasma emission spectrometry [21,51], and non-heme iron measured by colorimetric methods [13–19]. Total iron includes both heme and non-heme iron. The first indication that heme biosynthesis declines with age was provided by Bitar and Weiner [58], who examined age-related changes in heme and heme proteins in male Sprague-Dawley rats. This finding was further confirmed by studies with emphasis on heme deficiency in both neurodegenerative disorders [59–62] and normal aging [19]. In considering the findings of age-associated decline in heme biosynthesis and increases in non-heme iron levels, total iron measurements per se may not fully reflect iron dyshomeostasis in aging research, in particular when the conclusion of unaltered iron levels over time was reached using spectrometry techniques.
2.4. Ferroportin—The Only Way out for Cellular Iron
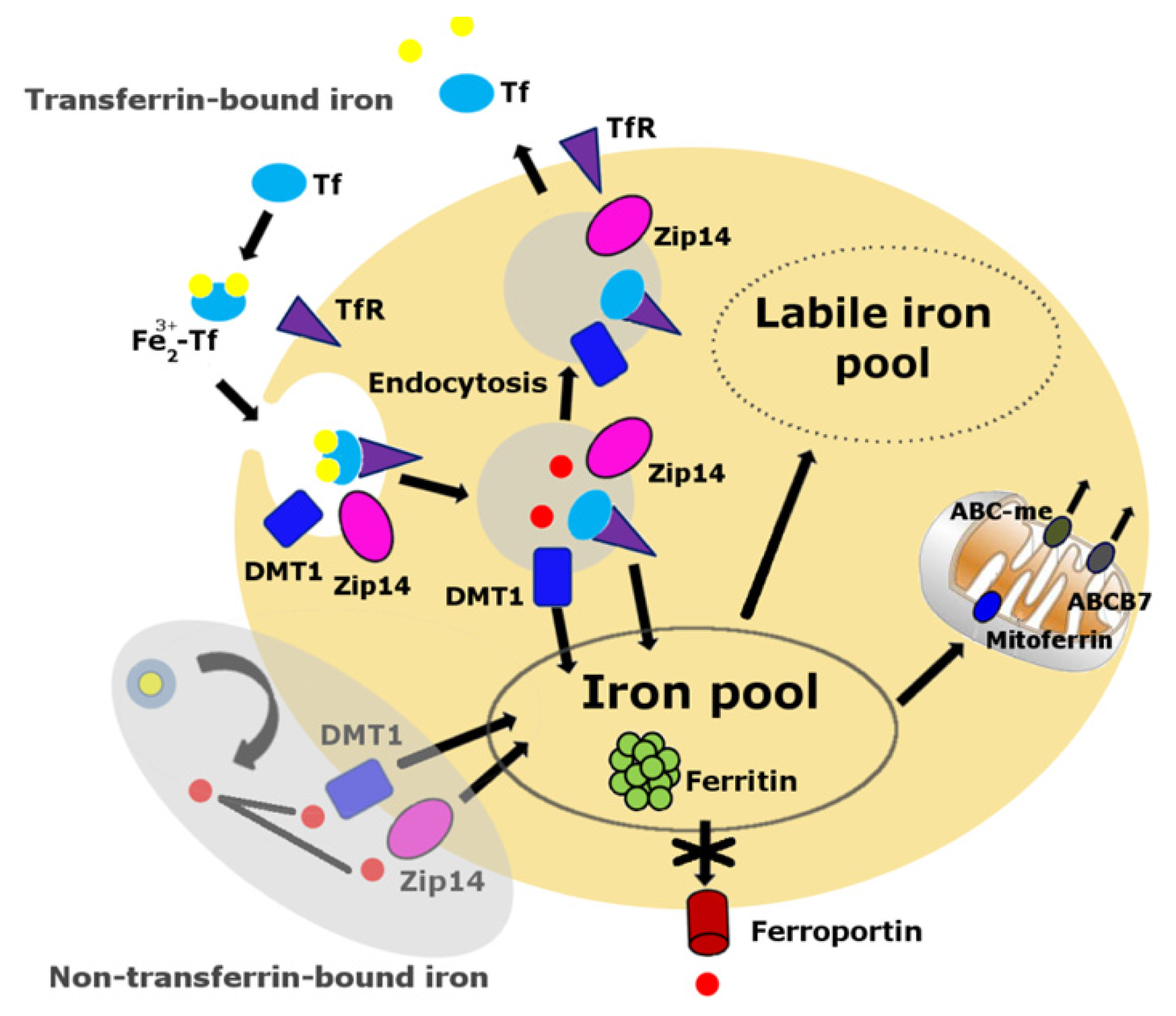
Cellular iron balance is coordinated by iron uptake, storage and export [63–65]. Iron cannot diffuse through cellular membranes unassisted. Either a receptor-mediated or non-receptor-mediated pathway is required to facilitate cellular iron import into the cytoplasm (Figure 1). The primary route of cellular iron acquisition is through receptor-mediated endocytosis of transferrin (Tf) [66]. Cells take up Tf-bound iron in proportion to their cell-surface expression of transferrin receptor (TfR) [67]. Divalent metal transporter-1 (DMT1), a ferrous iron transporter, can import iron into the cell, a mechanism which is essential for intestinal uptake of inorganic sources of dietary iron [65]. Zip14, a member of the SLC39 metal-ion transporter family, has also been shown to mediate iron uptake by cells [68,69]. Cellular iron export is mediated by ferroportin, the only known iron exporter in mammals [70].
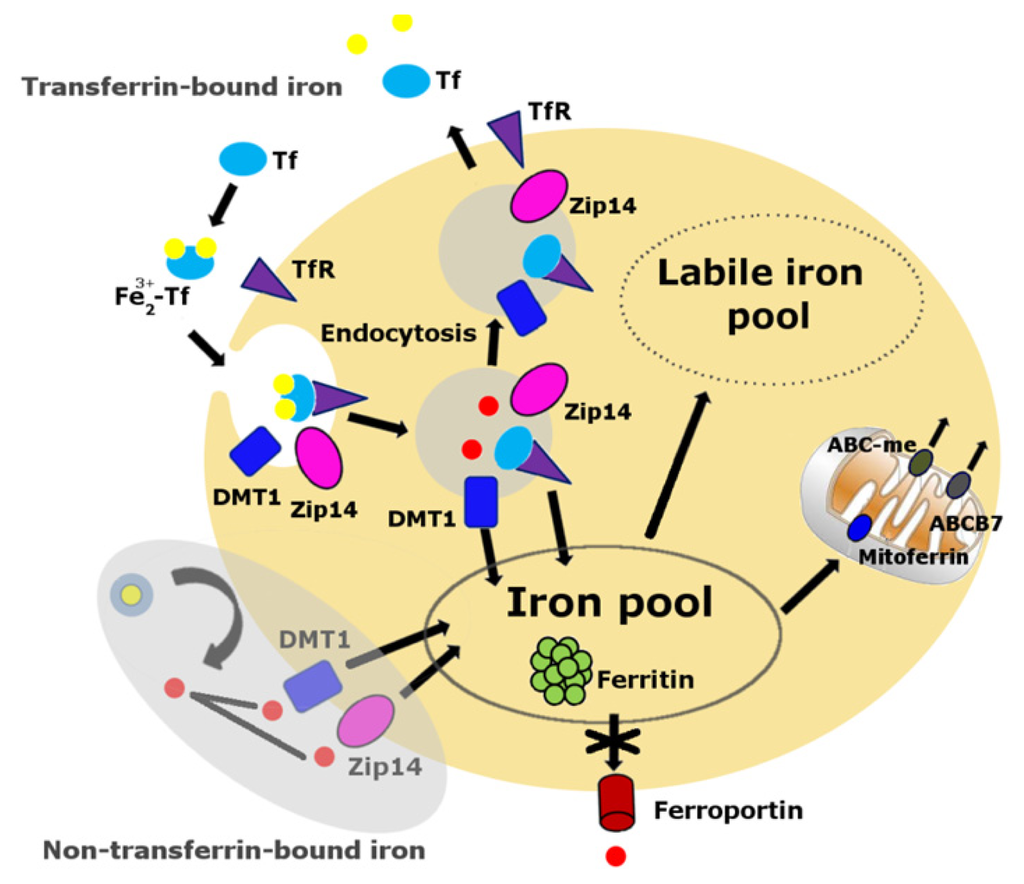
Figure 1.
Cellular labile iron pool. The transferrin-transferrin receptor (Tf-TfR) pathway is the primary route of cellular iron acquisition [67]. Cells assimilate iron when Fe3+-Tf binds to TfR at the cell surface, and the complex is internalized into endosomes. Endosomal acidification promotes iron to dissociate from Tf, and the metal is then reduced to Fe2+ and transported into the cytosol by the transmembrane protein divalent metal transporter 1 (DMT1) and Zip14. The non-Tf-bound iron pathway, the shaded area, appears mainly during states of iron overload. Much of the iron normally assimilated by cells is destined to the mitochondria via mitoferrin, the site of heme and iron-sulfur cluster biosynthesis. Iron is exported from the mitochondria in the form of iron-sulfur clusters or heme. Export of iron-sulfur clusters involves ABCB7. Cells export iron through ferroportin. The absence of ferroportin in skeletal muscles and other post-mitotic tissues may result in iron accumulation over time.
Ferroportin is a transmembrane protein expressed on the surface of absorptive enterocytes, macrophages, hepatocytes, and placental cells [70]. Tissue-specific ablation of ferroportin results in embryonic lethality [70]. McKie et al. [71] reported that ferroportin was weakly expressed in kidney, liver, and testis and absent in brain, heart, lung and skeletal muscle of mice fed on a normal diet [71]. A recent study on muscle iron metabolism in Fischer 344 x Brown Norway rats by us [15] further supported the finding that the absence of ferroportin in skeletal muscles significantly contributes to the iron accumulation in aged animals. Thus, the lack of a cellular iron export mechanism in post-mitotic tissues could be one of the essential factors contributing to iron accumulation in aging.
2.5. Animal Diets
Although iron balance is tightly regulated at the site of absorption (duodenum) [72], rodent diets with different iron levels may alter iron homeostasis across studies. The AIN-76 diet (American Institute of Nutrition, 1977) or AIN-93 [73], a substitute for the original AIN-76 diet to improve the performance of animals, is a widely used purified diet for laboratory rodents formulated with 35 mg iron/kg diet, an amount considered to meet the minimum requirement of iron for normal growth and hematopoiesis [74]. Natural-ingredient and typical rodent diets, which usually contain 198 to 270 mg iron/kg diet, have also been used to provide good health and reproduction in laboratory rodents [74,75]. In early 1970s, Sorbie and Valberg [76] observed that 25 to 100 mg iron/kg diet was associated with low iron storage in liver of male C57BL/6J mice and that higher concentrations may be necessary for reproduction. A recent study published by Cooksey et al. [77] also indicated that the mice on the 35 mg·iron/kg diet did exhibit remarkable decreases in hepatic iron and serum ferritin compared with mice on the 500 mg·iron/kg diet. In agreement with these observations, a long-term study on AIN-93M (maintenance formulation) diet by Ahluwalia et al. [13] showed that iron status and stores in liver, spleen and femur marrow decline with age in male Lewis rats. Despite the low iron stores in animals fed AIN purified diets, Jung et al. [22] demonstrated that non-heme iron and ferritin levels significantly increased with age in the plantaris muscle of male Fischer 344 rats during short-term feeding with AIN-93M diet, suggesting that skeletal muscles are extremely vulnerable to iron accumulation in aging. Long-term studies on dietary modification or adjustments are warranted to create an optimal diet containing a maintenance-level of iron that is suitable for rodent models at different life stages.
3. Iron Accumulation and Labile Iron
Cells maintain a pool of available labile iron [78] (Figure 1) identified by several terms, including “transition iron”, “free iron”, “low-molecular-weight iron”, “redox-active iron” or “chelatable iron” [79], which exists in dynamic equilibrium with various cellular components. Optimal function of cells highly depends on the maintenance of cellular iron levels [80]. When iron prevails over cellular iron sequestration, labile iron may be released from either loosely bound iron proteins or storage sites, particularly under conditions of cellular stress [81]. Labile iron is highly reactive and has the potential to catalyze the formation of harmful reactive oxygen species, ultimately leading to oxidative damage and cell death [82,83]. In light of previous studies showing catastrophic cellular damage by labile iron, Simunek et al. [84] showed that H2O2-induced collapse of mitochondrial membrane potential was completely prevented by pre-treatment with the lipophilic iron chelator, salicylaldehyde isonicotinoyl hydrazone (SIH), in cultured H9c2 cardiac myoblasts, suggesting that hydrogen peroxide per se is not harmful, but it may become highly toxic if labile iron coexists. Furthermore, the observation that iron chelation by an iron chelator, deferoxamine, mitigated immobilization-induced muscle loss in male Wistar rats implies that labile iron could be one of several potential contributors to accelerate muscle atrophy during prolonged inactivity [85].
4. Iron Dyshomeostasis in Age-Associated Disorders in Humans
Clinical and epidemiological studies have shown that iron plays important roles in multiple agingassociated disorders, such as cardiovascular diseases [1,2], inflammatory diseases [86], neurodegenerative diseases [10,11], and cancer [5–7,87]. Salonen et al. [88] reported that men with serum ferritin greater than or equal to 200 μg/L had a 2.2-fold increased risk of acute myocardial infarction compared with men with a lower serum ferritin at age of 42 to 60 in eastern Finland. In agreement with the epidemiological finding, Tuomainen et al. [89] demonstrated that men with high body iron stores were at a 2- to 3-fold increased risk of the first acute myocardial infarction. In healthy subjects [90] and anemic patients [91], the level of serum ferritin showed an age-related tendency to increase. Recently, Tull et al. [92] indicated that these subjects are likely to have anemia of chronic diseases with adequate iron stores and unable to utilize iron from storage sites. Therefore, the most common cause of anemia in the elderly is anemia of chronic disease, which has been identified as impaired iron status rather than iron deficiency.
Age-associated decline in hematologic variables has been the subject of extensive investigation in animal models [13,17,93–95] and humans [28,32,36,96]. A number of studies have shown that aging is associated with an erythropoietic decline [93,97] as well as a reduced reserve capacity in the hematopoietic system [98–100]. However, an early study reported by Boggs and Patrene [94] using B6D2 F1 female mice argued that an expanded plasma volume in aged animals substantially contributed to the decrease in hematocrit, whereas circulating red cell mass remained unchanged in aged animals, suggesting an age-related “dilutional” anemia. In a follow-up study using a mathematical model of erythropoiesis, Loeffler and Pantel [101] revealed that the lower hematocrits in aged mice were due to plasma volume expansion, rather than changes in red cell mass between young and aged animals. A clinical study of dietary iron intake and excretion in healthy elderly subjects aged 70 to 85 years indicated that hemoglobin levels were within the established reference range for adult individuals.
It has been proposed that age-related anemia may be associated with increased hepcidin levels in response to elevated interleukin-6 levels [102]. Hepcidin is the principal regulatory hormone produced by hepatocytes in response to iron loading [103] or inflammation [104]. Under iron overload conditions, hepcidin downregulates ferroportin expression in enterocytes and macrophages, thereby reducing serum iron levels via decreasing intestinal iron absorption and macrophage iron recycling [105]. Despite the important role of hepcidin in systemic iron homeostasis, quantification of plasma hepcidin has proved to be technically difficult. The development of the first validated serum enzyme-linked immunosorbent assay (ELISA) by Ganz et al. [106] has allowed significant advances in studies of ageassociated alterations in plasma hepcidin levels. A recent study using the ELISA assay in anemic patients by Lee et al. [107] showed that anemia in the elderly was not associated with increased plasma hepcidin levels. The observation in the same study that both the mean and median hepcidin levels were lower in anemic elderly patients suggests that elevated plasma hepcidin levels may be secondary to age-associated pathology, acute or chronic infections and inflammation. The findings further support the conclusion reported by Tull et al. [92] that aging is associated with impaired iron status, a most common cause of anemia in the elderly.
5. Iron and Mitochondrial Function in Aging
The mitochondrion is the central site of heme and iron-sulfur cluster biosynthesis [108]. Recent studies in both yeast and mammalian systems have shown that mitochondrial iron increase with age, in particular under conditions of cellular stress, which may be a potential causative factor in age-related mitochondrial dysfunction [18,109–111]. Rauen et al. [79] have developed a selective mitochondrial iron fluorescent probe, rhodamine B 4-[(2,20-bipyridin-4-yl)aminocarbonyl]benzyl ester (RDA), which shows that labile iron was about 16.0 μM in rat hepatocyte mitochondria. A study on muscle mitochondrial function in aged rats from our group [18] showed that aging was associated with elevated mitochondrial non-heme iron levels in skeletal muscle, which is significantly correlated with mitochondrial susceptibility to permeability transition pore opening, an important factor in the pathogenesis of cell death. Moreover, Veatch et al. [109] established a link between defects in ironsulfur cluster biosynthesis and genomic instability in yeast aging research. It has been shown that yeast cell aging was associated with an impairment of mitochondrial DNA integrity, which in turn affects the transport efficiency of iron-sulfur proteins between cytoplasm and mitochondria. Furthermore, impaired mitochondrial iron-sulfur biosynthesis contributed to increased cellular iron acquisition, iron regulon activation and mitochondrial iron accumulation. These observations highlight the mechanism of altered iron homeostasis in mitochondria, which may cause multiple defects in mitochondrial heme and iron-sulfur cluster biosynthesis as well as iron accumulation.
6. Future Research
Age-associated iron dyshomeostasis is a process of progressive changes in multiple organ systems. Much research effort is directed at developing therapeutics or interventions to combat these changes. Some impressive successes have been achieved in non-mammalian models using iron chelators to mitigate iron overload and iron-related disorders, such as Alzheimer’s disease [112,113], Parkinson’s disease [114,115], Friedreich’s ataxia [116,117] and retinal disease [118,119]. A major concern arises from iron chelation therapy against the aging process is that compounds available to date cannot specifically target individual organs or systems. This may dramatically limit the use of iron chelators in elderly persons, in particular when considering the finding that altered iron status is characterized by adequate iron stores and low hematologic variables in both rodent [17] and human studies [92,120].
A major research challenge will be to develop novel, safe and feasible interventions that mitigate age-associated iron dyshomeostasis. Indeed, calorie restriction has been shown to be effective in modulating the age-associated iron accumulation in rat muscle, liver, brain and kidney [17,19,121]. Late-onset caloric restriction has proven to be less effective [122,123]. Dietary compounds that inhibit iron absorption (e.g., polyphenols in tea and coffee [124,125]) may offer alternative approaches to mitigate iron accumulation during the aging process. Future research is warranted to test dietary interventions.
7. Conclusions
In summary, impaired iron status and iron dyshomeostasis are associated with organ-specific changes in iron levels in multiple organ systems with age. Lack of ferroportin expression, at least in part, exacerbates iron accumulation over time in various tissues, such as skeletal muscle. Future research can be directed to late-onset therapeutics or interventions for modulating impaired iron status in aging.
Acknowledgements
This research was supported by NIH grants to C.L. (NIH AG17994), M.D.K. (NIH DK080706), a fellowship award from the American Heart Association to J.X. (09POST2060112), and the University of Florida Institute on Aging and Claude D. Pepper Older Americans Independence Center (1 P30AG028740).
References
- Roijers, R.B.; Debernardi, N.; Cleutjens, J.P.; Schurgers, L.J.; Mutsaers, P.H.; van der Vusse, G.J. Microcalcifications in early intimal lesions of atherosclerotic human coronary arteries. Am. J. Pathol 2011, 178, 2879–2887. [Google Scholar]
- Sullivan, J.L. Iron in arterial plaque: Modifiable risk factor for atherosclerosis. Biochim. Biophys. Acta 2009, 1790, 718–723. [Google Scholar]
- Carpenter, J.P.; He, T.; Kirk, P.; Roughton, M.; Anderson, L.J.; de Noronha, S.V.; Sheppard, M.N.; Porter, J.B.; Walker, J.M.; Wood, J.C.; et al. On t2* magnetic resonance and cardiac iron. Circulation 2011, 123, 1519–1528. [Google Scholar]
- Tajima, S.; Ikeda, Y.; Sawada, K.; Yamano, N.; Horinouchi, Y.; Kihira, Y.; Ishizawa, K.; Izawa-Ishizawa, Y.; Kawazoe, K.; Tomita, S.; et al. Iron reduction by deferoxamine leads to amelioration of adiposity via the regulation of oxidative stress and inflammation in obese and type 2 diabetes kkay mice. Am. J. Physiol. Endocrinol. Metab 2011. [Google Scholar] [CrossRef]
- Toyokuni, S. Role of iron in carcinogenesis: Cancer as a ferrotoxic disease. Cancer Sci 2009, 100, 9–16. [Google Scholar]
- Toyokuni, S. Iron as a target of chemoprevention for longevity in humans. Free Radic Res 2011, 45, 906–917. [Google Scholar]
- Torti, S.V.; Torti, F.M. Ironing out cancer. Cancer Res 2011, 71, 1511–1514. [Google Scholar]
- Morgan, N.V.; Westaway, S.K.; Morton, J.E.; Gregory, A.; Gissen, P.; Sonek, S.; Cangul, H.; Coryell, J.; Canham, N.; Nardocci, N.; et al. Pla2g6, encoding a phospholipase a2, is mutated in neurodegenerative disorders with high brain iron. Nat. Genet 2006, 38, 752–754. [Google Scholar]
- Pandolfo, M.; Pastore, A. The pathogenesis of friedreich ataxia and the structure and function of frataxin. J. Neurol 2009, 256, 9–17. [Google Scholar]
- Oshiro, S.; Morioka, M.S.; Kikuchi, M. Dysregulation of iron metabolism in alzheimer’s disease, parkinson’s disease, and amyotrophic lateral sclerosis. Adv. Pharmacol. Sci 2011, 2011, 378278. [Google Scholar]
- He, Q.; Du, T.; Yu, X.; Xie, A.; Song, N.; Kang, Q.; Yu, J.; Tan, L.; Xie, J.; Jiang, H. Dmt1 polymorphism and risk of parkinson’s disease. Neurosci. Lett 2011, 501, 128–131. [Google Scholar]
- Lei, P.; Ayton, S.; Finkelstein, D.I.; Spoerri, L.; Ciccotosto, G.D.; Wright, D.K.; Wong, B.X.; Adlard, P.A.; Cherny, R.A.; Lam, L.Q.; et al. Tau deficiency induces parkinsonism with dementia by impairing app-mediated iron export. Nat. Med 2012, 18, 291–295. [Google Scholar]
- Ahluwalia, N.; Gordon, M.A.; Handte, G.; Mahlon, M.; Li, N.Q.; Beard, J.L.; Weinstock, D.; Ross, A.C. Iron status and stores decline with age in lewis rats. J. Nutr 2000, 130, 2378–2383. [Google Scholar]
- Altun, M.; Edstrom, E.; Spooner, E.; Flores-Moralez, A.; Bergman, E.; Tollet-Egnell, P.; Norstedt, G.; Kessler, B.M.; Ulfhake, B. Iron load and redox stress in skeletal muscle of aged rats. Muscle Nerve 2007, 36, 223–233. [Google Scholar]
- Xu, J.; Hwang, J.C.; Lees, H.A.; Wohlgemuth, S.E.; Knutson, M.D.; Judge, A.R.; Dupont-Versteegden, E.E.; Marzetti, E.; Leeuwenburgh, C. Long-term perturbation of muscle iron homeostasis following hindlimb suspension in old rats is associated with high levels of oxidative stress and impaired recovery from atrophy. Exp. Gerontol 2012, 47, 100–108. [Google Scholar]
- Hofer, T.; Marzetti, E.; Xu, J.; Seo, A.Y.; Gulec, S.; Knutson, M.D.; Leeuwenburgh, C.; Dupont-Versteegden, E.E. Increased iron content and rna oxidative damage in skeletal muscle with aging and disuse atrophy. Exp.Gerontol 2008, 43, 563–570. [Google Scholar]
- Xu, J.; Knutson, M.D.; Carter, C.S.; Leeuwenburgh, C. Iron accumulation with age, oxidative stress and functional decline. PLoS One 2008, 3, e2865. [Google Scholar]
- Seo, A.Y.; Xu, J.; Servais, S.; Hofer, T.; Marzetti, E.; Wohlgemuth, S.E.; Knutson, M.D.; Chung, H.Y.; Leeuwenburgh, C. Mitochondrial iron accumulation with age and functional consequences. Aging Cell 2008, 7, 706–716. [Google Scholar]
- Cook, C.I.; Yu, B.P. Iron accumulation in aging: Modulation by dietary restriction. Mech. Ageing Dev 1998, 102, 1–13. [Google Scholar]
- Massie, H.R.; Aiello, V.R.; Banziger, V. Iron accumulation and lipid peroxidation in aging c57bl/6j mice. Exp. Gerontol 1983, 18, 277–285. [Google Scholar]
- Arvapalli, R.K.; Paturi, S.; Laurino, J.P.; Katta, A.; Kakarla, S.K.; Gadde, M.K.; Wu, M.; Rice, K.M.; Walker, E.M.; Wehner, P.; et al. Deferasirox decreases age-associated iron accumulation in the aging f344xbn rat heart and liver. Cardiovasc. Toxicol 2010, 10, 108–116. [Google Scholar]
- Jung, S.H.; DeRuisseau, L.R.; Kavazis, A.N.; Deruisseau, K.C. Plantaris muscle of aged rats demonstrates iron accumulation and altered expression of iron regulation proteins. Exp. Physiol 2008, 93, 407–414. [Google Scholar]
- Sohn, Y.S.; Breuer, W.; Munnich, A.; Cabantchik, Z.I. Redistribution of accumulated cell iron: A modality of chelation with therapeutic implications. Blood 2008, 111, 1690–1699. [Google Scholar]
- Sohal, R.S.; Wennberg-Kirch, E.; Jaiswal, K.; Kwong, L.K.; Forster, M.J. Effect of age and caloric restriction on bleomycin-chelatable and nonheme iron in different tissues of c57bl/6 mice. Free Radic. Biol. Med 1999, 27, 287–293. [Google Scholar]
- Bulvik, B.E.; Berenshtein, E.; Konijn, A.M.; Grinberg, L.; Vinokur, V.; Eliashar, R.; Chevion, M.M. Aging is an organ-specific process: Changes in homeostasis of iron and redox proteins in the rat. Age (Dordr.) 2011. [Google Scholar] [CrossRef]
- Aquino, D.; Bizzi, A.; Grisoli, M.; Garavaglia, B.; Bruzzone, M.G.; Nardocci, N.; Savoiardo, M.; Chiapparini, L. Age-related iron deposition in the basal ganglia: Quantitative analysis in healthy subjects. Radiology 2009, 252, 165–172. [Google Scholar]
- Gregory, A.; Polster, B.J.; Hayflick, S.J. Clinical and genetic delineation of neurodegeneration with brain iron accumulation. J. Med. Genet 2009, 46, 73–80. [Google Scholar]
- Gaskell, H.; Derry, S.; Andrew Moore, R.; McQuay, H.J. Prevalence of anaemia in older persons: Systematic review. BMC Geriatr 2008, 8, 1. [Google Scholar]
- Peran, P.; Cherubini, A.; Luccichenti, G.; Hagberg, G.; Demonet, J.F.; Rascol, O.; Celsis, P.; Caltagirone, C.; Spalletta, G.; Sabatini, U. Volume and iron content in basal ganglia and thalamus. Hum. Brain Mapp 2009, 30, 2667–2675. [Google Scholar]
- Cherubini, A.; Peran, P.; Caltagirone, C.; Sabatini, U.; Spalletta, G. Aging of subcortical nuclei: Microstructural, mineralization and atrophy modifications measured in vivo using mri. Neuroimage 2009, 48, 29–36. [Google Scholar]
- Zecca, L.; Gallorini, M.; Schunemann, V.; Trautwein, A.X.; Gerlach, M.; Riederer, P.; Vezzoni, P.; Tampellini, D. Iron, neuromelanin and ferritin content in the substantia nigra of normal subjects at different ages: Consequences for iron storage and neurodegenerative processes. J. Neurochem 2001, 76, 1766–1773. [Google Scholar]
- Inelmen, E.M.; D’Alessio, M.; Gatto, M.R.; Baggio, M.B.; Jimenez, G.; Bizzotto, M.G.; Enzi, G. Descriptive analysis of the prevalence of anemia in a randomly selected sample of elderly people living at home: Some results of an italian multicentric study. Aging (Milano) 1994, 6, 81–89. [Google Scholar]
- Thomson, C.A.; Stanaway, J.D.; Neuhouser, M.L.; Snetselaar, L.G.; Stefanick, M.L.; Arendell, L.; Chen, Z. Nutrient intake and anemia risk in the women’s health initiative observational study. J. Am. Diet Assoc 2011, 111, 532–541. [Google Scholar]
- Tussing-Humphreys, L.; Braunschweig, C. Anemia in postmenopausal women: Dietary inadequacy or nondietary factors? J. Am. Diet Assoc 2011, 111, 528–531. [Google Scholar]
- Przybyszewska, J.; Zekanowska, E.; Kedziora-Kornatowska, K.; Boinska, J.; Cichon, R.; Porzych, K. Prohepcidin and iron metabolism parameters in the obese elderly patients with anemia. J. Nutr. Health Aging 2011, 15, 259–264. [Google Scholar]
- Price, E.A.; Mehra, R.; Holmes, T.H.; Schrier, S.L. Anemia in older persons: Etiology and evaluation. Blood Cells Mol. Dis 2011, 46, 159–165. [Google Scholar]
- House, M.J.; St Pierre, T.G.; Milward, E.A.; Bruce, D.G.; Olynyk, J.K. Relationship between brain r(2) and liver and serum iron concentrations in elderly men. Magn. Reson. Med 2010, 63, 275–281. [Google Scholar]
- Penninx, B.W.; Pahor, M.; Cesari, M.; Corsi, A.M.; Woodman, R.C.; Bandinelli, S.; Guralnik, J.M.; Ferrucci, L. Anemia is associated with disability and decreased physical performance and muscle strength in the elderly. J. Am. Geriatr. Soc 2004, 52, 719–724. [Google Scholar]
- Roy, C.N. Anemia in frailty. Clin. Geriatr. Med 2011, 27, 67–78. [Google Scholar]
- Chaves, P.H.; Carlson, M.C.; Ferrucci, L.; Guralnik, J.M.; Semba, R.; Fried, L.P. Association between mild anemia and executive function impairment in community-dwelling older women: The women’s health and aging study ii. J. Am. Geriatr. Soc 2006, 54, 1429–1435. [Google Scholar]
- Denny, S.D.; Kuchibhatla, M.N.; Cohen, H.J. Impact of anemia on mortality, cognition, and function in community-dwelling elderly. Am. J. Med 2006, 119, 327–334. [Google Scholar]
- Blasiak, J.; Szaflik, J.; Szaflik, J.P. Implications of altered iron homeostasis for age-related macular degeneration. Front Biosci 2011, 16, 1551–1559. [Google Scholar]
- Franzini, C.; Berlusconi, A.; Favarelli, C.; Brambilla, S. Low frequency of elevated serum transferrin saturation in elderly subjects. Clin. Chim. Acta 2000, 298, 181–186. [Google Scholar]
- Richer, S.; Rudy, D.; Statkute, L.; Karofty, K.; Frankowski, J. Serum iron, transferrin saturation, ferritin, and dietary data in age-related macular degeneration. Am. J. Ther 2002, 9, 25–28. [Google Scholar]
- Zekanowska, E.; Boinska, J.; Kwapisz, J.; Kedziora-Kornatowska, K.; Porzych, K.; Ratajczak, M. Serum Prohepcidin and Other Iron Metabolism Parameters in Healthy Adults. Przegl. Lek 2011, 68, 82–86. [Google Scholar]
- Schrag, M.; Mueller, C.; Oyoyo, U.; Smith, M.A.; Kirsch, W.M. Iron, zinc and copper in the alzheimer’s disease brain: A quantitative meta-analysis. Some insight on the influence of citation bias on scientific opinion. Prog. Neurobiol 2011, 94, 296–306. [Google Scholar]
- Sullivan, J.L. Is stored iron safe? J. Lab. Clin. Med 2004, 144, 280–284. [Google Scholar]
- Anderson, L.J.; Wonke, B.; Prescott, E.; Holden, S.; Walker, J.M.; Pennell, D.J. Comparison of effects of oral deferiprone and subcutaneous desferrioxamine on myocardial iron concentrations and ventricular function in beta-thalassaemia. Lancet 2002, 360, 516–520. [Google Scholar]
- Pennell, D.J.; Berdoukas, V.; Karagiorga, M.; Ladis, V.; Piga, A.; Aessopos, A.; Gotsis, E.D.; Tanner, M.A.; Smith, G.C.; Westwood, M.A.; et al. Randomized controlled trial of deferiprone or deferoxamine in beta-thalassemia major patients with asymptomatic myocardial siderosis. Blood 2006, 107, 3738–3744. [Google Scholar]
- Turturro, A.; Witt, W.W.; Lewis, S.; Hass, B.S.; Lipman, R.D.; Hart, R.W. Growth curves and survival characteristics of the animals used in the biomarkers of aging program. J. Gerontol. A Biol. Sci. Med. Sci 1999, 54, B492–B501. [Google Scholar]
- Takeda, T.; Kimura, M.; Yokoi, K.; Itokawa, Y. Effect of age and dietary protein level on tissue mineral levels in female rats. Biol. Trace Elem. Res 1996, 54, 55–74. [Google Scholar]
- Koolhaas, J.M. The Ufaw Handbook on the Care and Management of Laboratory Animals; The Universities Federation for Animal Welfare: Hertfordshire, UK, 2010; pp. 311–322. [Google Scholar]
- Feldman, J.D.; Woda, B.A. Pathology and tumor incidence in aged lewis and bn rats. Clin. Immunol. Immunopathol 1980, 15, 331–343. [Google Scholar]
- Keenan, K.P.; Smith, P.F.; Hertzog, P.; Soper, K.; Ballam, G.C.; Clark, R.L. The effects of overfeeding and dietary restriction on sprague-dawley rat survival and early pathology biomarkers of aging. Toxicol. Pathol 1994, 22, 300–315. [Google Scholar]
- Rice, K.M.; Linderman, J.K.; Kinnard, R.S.; Blough, E.R. The fischer 344/nniahsd x brown norway/binia is a better model of sarcopenia than the fischer 344/nniahsd: A comparative analysis of muscle mass and contractile properties in aging male rat models. Biogerontology 2005, 6, 335–343. [Google Scholar]
- Rice, K.M.; Wu, M.; Blough, E.R. Aortic aging in the fischer 344/nniahsd x brown norway/binia rat. J. Pharmacol. Sci 2008, 108, 393–398. [Google Scholar]
- Hahn, P.; Song, Y.; Ying, G.S.; He, X.; Beard, J.; Dunaief, J.L. Age-dependent and gender-specific changes in mouse tissue iron by strain. Exp.Gerontol 2009, 44, 594–600. [Google Scholar]
- Bitar, M.; Weiner, M. Modification of age-induced changes in heme and hemoproteins by testosterone in male rats. Mech. Ageing Dev 1983, 23, 285–296. [Google Scholar]
- Atamna, H. Heme, iron, and the mitochondrial decay of ageing. Ageing Res. Rev 2004, 3, 303–318. [Google Scholar]
- Atamna, H.; Killilea, D.W.; Killilea, A.N.; Ames, B.N. Heme deficiency may be a factor in the mitochondrial and neuronal decay of aging. Proc. Natl. Acad. Sci. USA 2002, 99, 14807–14812. [Google Scholar]
- Atamna, H.; Liu, J.; Ames, B.N. Heme deficiency selectively interrupts assembly of mitochondrial complex iv in human fibroblasts: Revelance to aging. J. Biol. Chem 2001, 276, 48410–48416. [Google Scholar]
- Atamna, H.; Walter, P.B.; Ames, B.N. The role of heme and iron-sulfur clusters in mitochondrial biogenesis, maintenance, and decay with age. Arch. Biochem. Biophys 2002, 397, 345–353. [Google Scholar]
- Chua, A.C.; Graham, R.M.; Trinder, D.; Olynyk, J.K. The regulation of cellular iron metabolism. Crit. Rev. Clin. Lab. Sci 2007, 44, 413–459. [Google Scholar]
- Kohgo, Y.; Ikuta, K.; Ohtake, T.; Torimoto, Y.; Kato, J. Body iron metabolism and pathophysiology of iron overload. Int. J. Hematol 2008, 88, 7–15. [Google Scholar]
- Dunn, L.L.; Rahmanto, Y.S.; Richardson, D.R. Iron uptake and metabolism in the new millennium. Trends Cell Biol 2007, 17, 93–100. [Google Scholar]
- Leverence, R.; Mason, A.B.; Kaltashov, I.A. Noncanonical interactions between serum transferrin and transferrin receptor evaluated with electrospray ionization mass spectrometry. Proc. Natl. Acad. Sci. USA 2010, 107, 8123–8128. [Google Scholar]
- Hofer, T.; Marzetti, E.; Seo, A.Y.; Xu, J.; Knutson, M.D.; Leeuwenburgh, C. Mechanisms of iron regulation and oxidative stress in sarcopenia and neurodegenerative diseases. In Free Radicals in Biology and Medicine; Gutierrez-Merino, C., Leeuwenburgh, C., Eds.; Research Signpost: Kerala, India, 2008; pp. 1–22. [Google Scholar]
- Liuzzi, J.P.; Aydemir, F.; Nam, H.; Knutson, M.D.; Cousins, R.J. Zip14 (slc39a14) mediates non-transferrin-bound iron uptake into cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13612–13617. [Google Scholar]
- Zhao, N.; Gao, J.; Enns, C.A.; Knutson, M.D. Zrt/irt-like protein 14 (zip14) promotes the cellular assimilation of iron from transferrin. J. Biol. Chem 2010, 285, 32141–32150. [Google Scholar]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The iron exporter ferroportin/slc40a1 is essential for iron homeostasis. Cell Metab 2005, 1, 191–200. [Google Scholar]
- McKie, A.T.; Marciani, P.; Rolfs, A.; Brennan, K.; Wehr, K.; Barrow, D.; Miret, S.; Bomford, A.; Peters, T.J.; Farzaneh, F.; et al. A novel duodenal iron-regulated transporter, ireg1, implicated in the basolateral transfer of iron to the circulation. Mol. Cell 2000, 5, 299–309. [Google Scholar]
- Galy, B.; Ferring, D.; Minana, B.; Bell, O.; Janser, H.G.; Muckenthaler, M.; Schumann, K.; Hentze, M.W. Altered body iron distribution and microcytosis in mice deficient in iron regulatory protein 2 (irp2). Blood 2005, 106, 2580–2589. [Google Scholar]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. Ain-93 purified diets for laboratory rodents: Final report of the american institute of nutrition ad hoc writing committee on the reformulation of the ain-76a rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar]
- Nutrient requirements of the mouse. In Nutrient Requirements of Laboratory Animals, 4th ed; Subcommittee on Laboratory Animal Nutrition; Commitee on Animal Nutrition, Board on Agriculture, National Research Council, National Academy Press: Washington, DC, USA, 1995; p. 90.
- Knapka, J.J.; Smith, K.P.; Judge, F.J. Effect of open and closed formula rations on the performance of three strains of laboratory mice. Lab. Anim. Sci 1974, 24, 480–487. [Google Scholar]
- Sorbie, J.; Valberg, L.S. Iron balance in the mouse. Lab. Anim. Sci 1974, 24, 900–904. [Google Scholar]
- Cooksey, R.C.; Jones, D.; Gabrielsen, S.; Huang, J.; Simcox, J.A.; Luo, B.; Soesanto, Y.; Rienhoff, H.; Abel, E.D.; McClain, D.A. Dietary iron restriction or iron chelation protects from diabetes and loss of beta-cell function in the obese (ob/ob lep-/-) mouse. Am. J. Physiol. Endocrinol. Metab 2010, 298, E1236–E1243. [Google Scholar]
- Breuer, W.; Shvartsman, M.; Cabantchik, Z.I. Intracellular labile iron. Int. J. Biochem. Cell Biol 2007, 40, 350–354. [Google Scholar]
- Rauen, U.; Springer, A.; Weisheit, D.; Petrat, F.; Korth, H.G.; de, G.H.; Sustmann, R. Assessment of chelatable mitochondrial iron by using mitochondrion-selective fluorescent iron indicators with different iron-binding affinities. ChemBioChem 2007, 8, 341–352. [Google Scholar]
- Wang, J.; Pantopoulos, K. Regulation of cellular iron metabolism. Biochem. J 2011, 434, 365–381. [Google Scholar]
- Cantu, D.; Schaack, J.; Patel, M. Oxidative inactivation of mitochondrial aconitase results in iron and h2o2-mediated neurotoxicity in rat primary mesencephalic cultures. PLoS One 2009, 4, e7095. [Google Scholar]
- Cabantchik, Z.I.; Kakhlon, O.; Epsztejn, S.; Zanninelli, G.; Breuer, W. Intracellular and extracellular labile iron pools. Adv. Exp. Med. Biol 2002, 509, 55–75. [Google Scholar]
- Kruszewski, M. Labile iron pool: The main determinant of cellular response to oxidative stress. Mutat. Res 2003, 531, 81–92. [Google Scholar]
- Simunek, T.; Boer, C.; Bouwman, R.A.; Vlasblom, R.; Versteilen, A.M.; Sterba, M.; Gersl, V.; Hrdina, R.; Ponka, P.; de Lange, J.J.; et al. Sih—a novel lipophilic iron chelator—protects h9c2 cardiomyoblasts from oxidative stress-induced mitochondrial injury and cell death. J. Mol. Cell Cardiol 2005, 39, 345–354. [Google Scholar]
- Kondo, H.; Miura, M.; Kodama, J.; Ahmed, S.M.; Itokawa, Y. Role of iron in oxidative stress in skeletal muscle atrophied by immobilization. Pflugers Arch 1992, 421, 295–297. [Google Scholar]
- Cherayil, B.J.; Ellenbogen, S.; Shanmugam, N.N. Iron and intestinal immunity. Curr. Opin. Gastroenterol 2011, 27, 523–528. [Google Scholar]
- Weijl, N.I.; Elsendoorn, T.J.; Moison, R.M.; Lentjes, E.G.; Brand, R.; Berger, H.M.; Osanto, S. Non-protein bound iron release during chemotherapy in cancer patients. Clin. Sci. (Lond.) 2004, 106, 475–484. [Google Scholar]
- Salonen, J.T.; Nyyssonen, K.; Korpela, H.; Tuomilehto, J.; Seppanen, R.; Salonen, R. High stored iron levels are associated with excess risk of myocardial infarction in eastern finnish men. Circulation 1992, 86, 803–811. [Google Scholar]
- Tuomainen, T.P.; Punnonen, K.; Nyyssonen, K.; Salonen, J.T. Association between body iron stores and the risk of acute myocardial infarction in men. Circulation 1998, 97, 1461–1466. [Google Scholar]
- Casale, G.; Bonora, C.; Migliavacca, A.; Zurita, I.E.; de Nicola, P. Serum ferritin and ageing. Age Ageing 1981, 10, 119–122. [Google Scholar]
- Joosten, E.; Van Loon, R.; Billen, J.; Blanckaert, N.; Fabri, R.; Pelemans, W. Serum transferrin receptor in the evaluation of the iron status in elderly hospitalized patients with anemia. Am. J. Hematol 2002, 69, 1–6. [Google Scholar]
- Tull, K.I.; Hirani, V.; Ali, A.; Chua, E.; Mindell, J.S. Impact of different diagnostic thresholds and the anaemia-ferritin-transferrin receptor model on the prevalence of anaemia and impaired iron status in older people. Age Ageing 2009, 38, 609–613. [Google Scholar]
- Boggs, D.R.; Patrene, K. Hematopoiesis and aging. V. A decline in hematocrit occurs in all aging female b6d2f1 mice. Exp. Aging Res 1986, 12, 131–134. [Google Scholar]
- Boggs, D.R.; Patrene, K.D. Hematopoiesis and aging iii: Anemia and a blunted erythropoietic response to hemorrhage in aged mice. Am. J. Hematol 1985, 19, 327–338. [Google Scholar]
- Finch, C.E.; Foster, J.R. Hematologic and serum electrolyte values of the c57bl-6j male mouse in maturity and senescence. Lab. Anim. Sci 1973, 23, 339–349. [Google Scholar]
- Seaverson, E.L.; Buell, J.S.; Fleming, D.J.; Bermudez, O.I.; Potischman, N.; Wood, R.J.; Chasan-Taber, L.; Tucker, K.L. Poor iron status is more prevalent in hispanic than in non-hispanic white older adults in massachusetts. J. Nutr 2007, 137, 414–420. [Google Scholar]
- Eisenstaedt, R.; Penninx, B.W.; Woodman, R.C. Anemia in the elderly: Current understanding and emerging concepts. Blood Rev 2006, 20, 213–226. [Google Scholar]
- Saitoh, T.; Morimoto, K.; Kumagai, T.; Tsuboi, I.; Aikawa, S.; Horie, T. Comparison of erythropoietic response to androgen in young and old senescence accelerated mice. Mech. Ageing Dev 1999, 109, 125–139. [Google Scholar]
- Lipschitz, D.A. Age-related declines in hematopoietic reserve capacity. Semin. Oncol 1995, 22, 3–5. [Google Scholar]
- Balducci, L.; Hardy, C.L.; Lyman, G.H. Hemopoiesis and aging. Cancer Treat Res 2005, 124, 109–134. [Google Scholar]
- Loeffler, M.; Pantel, K. A mathematical model of erythropoiesis suggests an altered plasma volume control as cause for anemia in aged mice. Exp. Gerontol 1990, 25, 483–495. [Google Scholar]
- Nemeth, E.; Valore, E.V.; Territo, M.; Schiller, G.; Lichtenstein, A.; Ganz, T. Hepcidin, a putative mediator of anemia of inflammation, is a type ii acute-phase protein. Blood 2003, 101, 2461–2463. [Google Scholar]
- Pigeon, C.; Ilyin, G.; Courselaud, B.; Leroyer, P.; Turlin, B.; Brissot, P.; Loreal, O. A new mouse liver-specific gene, encoding a protein homologous to human antimicrobial peptide hepcidin, is overexpressed during iron overload. J. Biol. Chem 2001, 276, 7811–7819. [Google Scholar]
- Nemeth, E.; Ganz, T. The role of hepcidin in iron metabolism. Acta Haematol 2009, 122, 78–86. [Google Scholar]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar]
- Ganz, T.; Olbina, G.; Girelli, D.; Nemeth, E.; Westerman, M. Immunoassay for human serum hepcidin. Blood 2008, 112, 4292–4297. [Google Scholar]
- Lee, P.; Gelbart, T.; Waalen, J.; Beutler, E. The anemia of ageing is not associated with increased plasma hepcidin levels. Blood Cells Mol. Dis 2008, 41, 252–254. [Google Scholar]
- Levi, S.; Rovida, E. The role of iron in mitochondrial function. Biochim. Biophys. Acta 2009, 1790, 629–636. [Google Scholar]
- Veatch, J.R.; McMurray, M.A.; Nelson, Z.W.; Gottschling, D.E. Mitochondrial dysfunction leads to nuclear genome instability via an iron-sulfur cluster defect. Cell 2009, 137, 1247–1258. [Google Scholar]
- Liang, L.P.; Jarrett, S.G.; Patel, M. Chelation of mitochondrial iron prevents seizure-induced mitochondrial dysfunction and neuronal injury. J. Neurosci 2008, 28, 11550–11556. [Google Scholar]
- Duvigneau, J.C.; Piskernik, C.; Haindl, S.; Kloesch, B.; Hartl, R.T.; Huttemann, M.; Lee, I.; Ebel, T.; Moldzio, R.; Gemeiner, M.; et al. A novel endotoxin-induced pathway: Upregulation of heme oxygenase 1, accumulation of free iron, and free iron-mediated mitochondrial dysfunction. Lab. Invest 2008, 88, 70–77. [Google Scholar]
- Weinberg, E.D.; Miklossy, J. Iron withholding: A defense against disease. J. Alzheimers. Dis 2008, 13, 451–463. [Google Scholar]
- Liu, G.; Men, P.; Perry, G.; Smith, M.A. Nanoparticle and iron chelators as a potential novel alzheimer therapy. Methods Mol. Biol 2010, 610, 123–144. [Google Scholar]
- Kaur, D.; Andersen, J. Does cellular iron dysregulation play a causative role in parkinson’s disease? Ageing Res. Rev 2004, 3, 327–343. [Google Scholar]
- Ghosh, B.; Antonio, T.; Reith, M.E.; Dutta, A.K. Discovery of 4-(4-(2-((5-hydroxy-1,2,3,4- tetrahydronaphthalen-2-yl)(propyl)amino)ethyl) piperazin-1-yl)quinolin-8-ol and its analogues as highly potent dopamine d2/d3 agonists and as iron chelator: In vivo activity indicates potential application in symptomatic and neuroprotective therapy for parkinson’s disease. J. Med. Chem 2010, 53, 2114–2125. [Google Scholar]
- Whitnall, M.; Rahmanto, Y.S.; Sutak, R.; Xu, X.; Becker, E.M.; Mikhael, M.R.; Ponka, P.; Richardson, D.R. The mck mouse heart model of friedreich’s ataxia: Alterations in iron-regulated proteins and cardiac hypertrophy are limited by iron chelation. Proc. Natl. Acad. Sci. USA 2008, 105, 9757–9762. [Google Scholar]
- Goncalves, S.; Paupe, V.; Dassa, E.P.; Rustin, P. Deferiprone targets aconitase: Implication for friedreich’s ataxia treatment. BMC Neurol 2008, 8, 20. [Google Scholar]
- Dunaief, J.L. Iron induced oxidative damage as a potential factor in age-related macular degeneration: The cogan lecture. Invest Ophthalmol. Vis. Sci 2006, 47, 4660–4664. [Google Scholar]
- Lukinova, N.; Iacovelli, J.; Dentchev, T.; Wolkow, N.; Hunter, A.; Amado, D.; Ying, G.S.; Sparrow, J.R.; Dunaief, J.L. Iron chelation protects the retinal pigment epithelial cell line arpe-19 against cell death triggered by diverse stimuli. Invest Ophthalmol. Vis. Sci 2009, 50, 1440–1447. [Google Scholar]
- Darnton-Hill, I.; Webb, P.; Harvey, P.W.; Hunt, J.M.; Dalmiya, N.; Chopra, M.; Ball, M.J.; Bloem, M.W.; De, B.B. Micronutrient deficiencies and gender: Social and economic costs. Am. J. Clin. Nutr 2005, 81, 1198S–1205S. [Google Scholar]
- Choi, J.H.; Kim, D.W.; Yu, B. Modulation of age-related alterations of iron, ferritin, and lipid peroxidation in rat brain synaptosomes. J. Nutr. Health Aging 1998, 2, 133–137. [Google Scholar]
- Weindruch, R.; Walford, R.L. Dietary restriction in mice beginning at 1 year of age: Effect on life-span and spontaneous cancer incidence. Science 1982, 215, 1415–1418. [Google Scholar]
- Olshansky, S.J.; Hayflick, L.; Carnes, B.A. Position statement on human aging. J. Gerontol. A Biol. Sci. Med. Sci 2002, 57, B292–B297. [Google Scholar]
- Brune, M.; Rossander, L.; Hallberg, L. Iron absorption and phenolic compounds: Importance of different phenolic structures. Eur. J. Clin. Nutr 1989, 43, 547–557. [Google Scholar]
- Hurrell, R.F.; Reddy, M.; Cook, J.D. Inhibition of non-haem iron absorption in man by polyphenolic-containing beverages. Br. J. Nutr 1999, 81, 289–295. [Google Scholar]
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).