Abstract
The function of the nuclear receptor (NR) in breast cancer progression has been investigated for decades. The majority of the nuclear receptors have well characterized natural ligands, but a few of them are orphan receptors for which no ligand has been identified. RORα, one member of the retinoid orphan nuclear receptor (ROR) subfamily of orphan receptors, regulates various cellular and pathological activities. RORα is commonly down-regulated and/or hypoactivated in breast cancer compared to normal mammary tissue. Expression of RORα suppresses malignant phenotypes in breast cancer cells, in vitro and in vivo. Activity of RORα can be categorized into the canonical and non-canonical nuclear receptor pathways, which in turn regulate various breast cancer cellular function, including cell proliferation, apoptosis and invasion. This information suggests that RORα is a potent tumor suppressor and a potential therapeutic target for breast cancer.
1. Introduction
Inactivation of tumor suppressors is essential for cancer development and progression. It has been shown that a wide variety of tumor suppressors, such as P53 [1], PTEN [2] and some microRNA [3], have the potential to be used as therapeutic targets. Breast cancer is one of the most common malignancies of women worldwide. In 2010, global incidence of breast cancer was about 1,643,000 cases and breast cancer-related women deaths were about 425,000 [4]. Therefore, there is urgent need to identify novel therapeutic targets to fight this mortal disease. We and others recently showed that the orphan nuclear receptor RORα is downregulated in cancer tissues and cell lines and that expression of RORα results in tumor suppressive activities [5–7], suggesting that RORα is a potential drug target for breast cancer treatment.
Aberrant activation of nuclear receptors (NR) during breast cancer progression was observed many years ago. The clinical value of NR as a therapeutic target has already been demonstrated. For example, estrogen receptor-α (ERα), overexpressed in ~70% of breast cancers, is an effective target for the treatment of breast cancer [8]. In contrast, most breast cancers show a down-regulation of retinoic acid receptor (RAR) expression [9], while activation of RAR in breast cancer cells appears to have growth-inhibitory activity [10]. These findings raise hope that perhaps NR may provide new options to prevent progression in human breast cancer.
There are 48 members in the human NR superfamily, which includes receptors for thyroid hormone, steroid hormones, various lipids and oxysterols. The majority of nuclear receptors have well characterized natural ligands, but a few of them are orphan receptors for which no ligand has been identified [11]. Retinoid orphan nuclear receptor (ROR), a subfamily of the orphan nuclear factor family, is so-named because of sequence similarities to the retinoic acid receptor (RAR) and the retinoid X receptor (RXR) [12,13]. In the early 1990s, RORα was identified as the first member of ROR subfamily of orphan receptors. Expression of RORα was found in multiple tissues and cells, including brain, muscle, colon, heart, skin, lung, spleen, leukocytes and mammary epithelial cells [14,15]. Aberrant activation of RORα influences various cellular pathologies, such as osteoporosis, autoimmune diseases, asthma and obesity [16–19]. Furthermore, reduced expression and hypoactivation of RORα in several human tumors, combined with their functional role as tumor suppressors, make RORα an attractive target for cancer therapy.
2. RORα Structure
RORα shows a domain structure similar to other NRs with four major functional domains (Figure 1A). The A/B region refers to amino-terminal of RORα. The C region, highly conserved among the ROR family members, is the DNA binding domain (DBD). A relatively short region, D, or the hinge domain, links the C region to the E region. The E region is the ligand binding domain (LBD); in addition to ligand recognition and binding, the LBD also regulates ligand-dependent transcriptional activity. The F region, a carboxy-terminal to the LBD, exists in some NRs [20]. There are four human RORα isoforms, referred to as ROR (α1–α4), while only two isoforms, α1 and α4, have been identified for mice [11]. Isoforms of RORα vary in their A/B domains and display different DNA recognition and transactivation features [13]. Crystallographic studies of RORα suggest that sterols, such as cholesterol, cholesterol sulfate and 7-dehydrocholesterol, may act as a natural ligand of this receptor [21,22]. Recent research has demonstrated that, in human endometrial cells, cholesterol sulfate can regulate expression of the RORα responsive gene NR1D1 without binding to the RORα receptor itself, suggesting that cholesterol sulfate may regulate RORα responsive gene expression, not as a ligand for RORα [23].
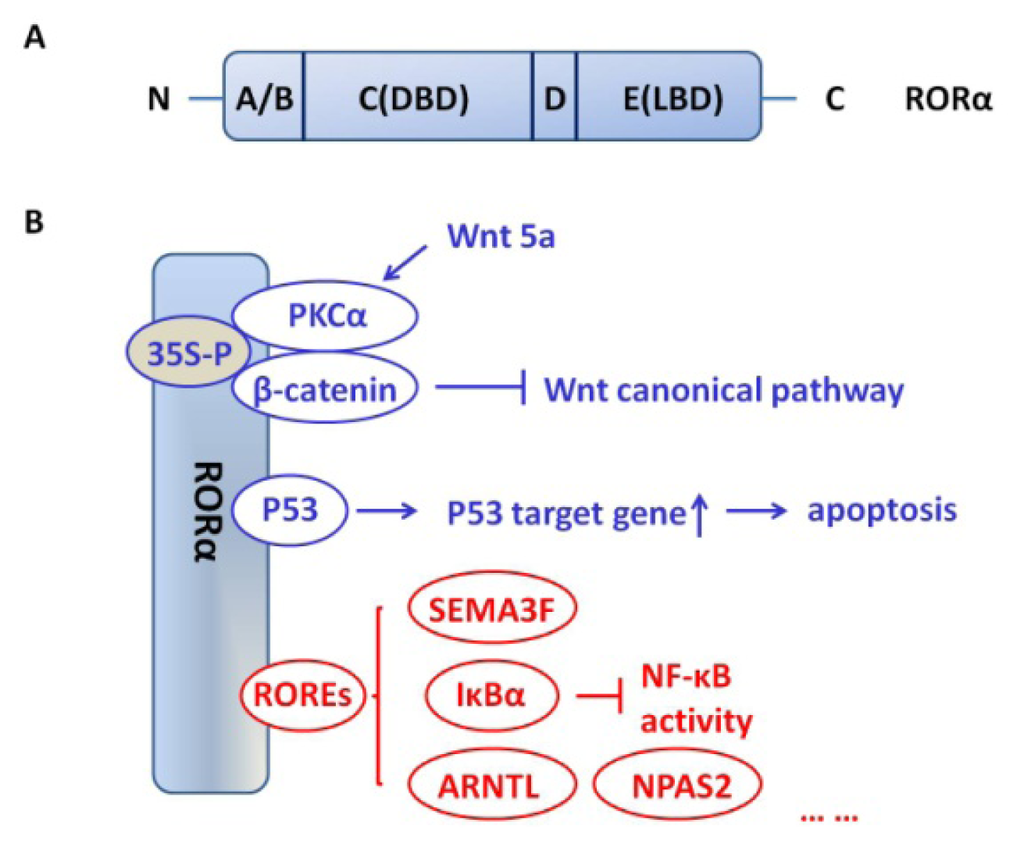
Figure 1.
Schematic structure of RORα and interaction of RORα with other proteins andpathways. (A) RORα shows a typical domain structure with four major functional domains. The A/B region refers to amino-terminal of RORα. The C region, highly conserved among the ROR family members, is the DNA binding domain (DBD). D is the hinge domain and links the C region to the E region. The E region is the ligand binding domain (LBD); (B) Canonical (red) and non-canonical (blue) nuclear receptor activities that may contribute to tumor suppressor function of RORα.
3. RORα Function in Human Breast Cancer
The RORα gene, which is comprised of 15 exons, covers a relatively large 730 kb genomic region. It maps to the middle of chromosome 15q22.2, a region that is highly unstable with frequent breaks and gene rearrangements [24]. Microarray data showed that mRNA levels of RORα are significantly reduced in many cancers (Table 1). RORα has also been identified as one of the methylation-silenced genes in gastric cancer cell lines [25], which favors the concept that reduced RORα expression promotes cancer progression. Downregulation of RORα phosphorylation was observed in colon cancer [26]. While RORα mRNA has been detected in both ER-positive and ER-negative human breast cancer cells [27], the RORα gene appears to be down-regulated in breast cancer compared to normal mammary tissue [24,28]. These results suggest that deregulation of RORα contributes to the development of breast cancer.

Table 1.
Analyzing published microarray datasets show that the mRNA levels of RORα is downregulated in various cancers; numbers in the table show how many datasets passed the threshold (cancer vs. normal: 1.5 fold change and p < 0.05). Blue represents the datasets in which the mRNA levels of RORα are downregulated in cancer tissues compared to normal tissues, while the datasets with upregulated RORα in cancer tissue are shown in red.
RORα plays an important role in suppressing malignant phenotypes in culture and in vivo. Recently, we reported that inhibition of RORα expression was associated with disruption of polarized acinar structure, the normal cytoarchitecture for breast tissue. Restoration of RORα expression in breast cancer cells resulted in morphologic characteristics associated with less aggressive tumor types: non-branched round spheroid structures in 3D culture, with a colony size and invasive capacity that was significantly reduced [5]. Since disruption of polarized acinar structure is an important early event for breast cancer development, this study suggested that reduced RORα expression contributes to the earliest stages of breast cancer development. In addition, expression of RORα in the mammary epithelial cell line MCF12F significantly inhibited cell proliferation [24]. Activation of RORα in prostate cancer cells affected cell cycle distribution, inducing a decrease in the S phase and a significant decrease of cell proliferation [7]. A recently study showed that introduction of RORα led to an increase of Dox-induced apoptosis in HCT-116 p53+/+ colon cancer cells [6]. Together, these results indicate that RORα is a potent tumor suppressor.
4. Potential Pathways that Mediate the Tumor Suppressive Activities of RORα
4.1. Canonical versus Non-Canonical Pathways
RORα activates nuclear receptor pathways in cancer cells that can be categorized as canonical and non-canonical (Figure 1B). Through these pathways, RORα regulates a variety of cellular activities, such as proliferation, invasion and cell polarization. The canonical RORα pathway involves binding of RORα to ROR response elements (ROREs). ROREs are the specific DNA sequences, AT-rich consensus motifs, in the regulatory region of the target gene [13]. Binding of RORα to the RORE modulates gene transcription and ultimately results in a change in the amount of protein produced. The most distinctive difference between the canonical and non-canonical pathways is the ability of the non-canonical pathway to influence gene expression without binding to ROREs. The mechanism by which RORα influences gene transcription is post-translational modifications and interaction. The significance of this pathway has been emphasized in recent studies.
4.2. Role of SEMA3F
SEMA3F is a tumor-suppressive microenvironmental factor that is often inactivated in metastatic cancer [29,30]. This factor has recently been characterized as a RORα-targeted gene [5]. Expression of RORα in breast cancer cells significantly induces SEMA3F transcription and inhibits the mammary tumor invasion in 3D culture [5]. RORE have been identified in the promoter region of the SEMA3F gene. Deletion of the RORE in the SEMA3F promoter significantly reduced the transcriptional activation driven by the SEMA3F promoter, indicating that RORα regulates transcription of SEMA3F through canonical nuclear receptor pathways. Moreover, silencing SEMA3F expression in RORα-expressing breast cancer cells rescues the invasive phenotypes in 3D culture, suggesting that tumor suppressor function of RORα is at least partially conferred by SEMA3F. On the other hand, reducing SEMA3F expression has little effect on tumor growth, suggesting that the tumor suppressor function of RORα involves other target genes and pathways as well [5].
4.3. Role of Wnt/β-Catenin
RORα activity is regulated by various post-translational modifications, including phosphorylation, ubiquitination and SUMOylation. Lee and colleagues showed that Wnt5a/PKC induces phosphorylation of RORα on serine residue 35 [26]. Wnt signaling can use the canonical (β-catenin dependent) and non-canonical (β-catenin independent) pathways. The canonical Wnt signaling pathway has been implicated in supporting breast transformation to cancer and in tumor progression [31,32]. Wnt5a activates non-canonical Wnt signaling and directs a breast cancer-suppressing effect [33,34]. Phosphorylated RORα, induced by Wnt5a/PKC pathway activation, attenuates the canonical Wnt signaling pathway. The inhibition is accomplished through binding of RORα to β-catenin, which suppresses the transcription of Wnt/β-catenin target genes. The transrepression mechanism of RORα on β-catenin is achieved, at least in part, by competition with a subset of coactivators for β-catenin binding and, possibly, recruitment of histone lysine methyltransferases, which results in transcriptional repression [26]. Therefore, RORα may suppress breast cancer progression by inhibiting Wnt/β-catenin target genes.
4.4. Role of p53
It is well-established that p53-regulated apoptosis and DNA repair are important in preventing cancers and that aberrant p53 function promotes breast cancer development and progression [35,36]. RORα has recently been identified as a direct p53 target gene. DNA damaging agents, such as doxorubicin and ionizing radiation, induce RORα expression in a p53-dependent manner [6]. Interestingly, RORα can also enhance DNA damage-induced apoptosis through p53 in colon cancer cells. It is revealed by genome-wide analysis that RORα could regulate p53-responsive genes, which mainly influence apoptosis. Further study also showed that RORα regulates p53 stability and p53 transcription activation in a HAUSP/Usp7-dependent manner [6]. Although enhancing p53 target gene by RORα is also reported in hepatocellular carcinoma cells [37], it remains to be determined whether RORα could stimulate breast cancer cell apoptosis via such an interaction with p53.
4.5. Role of Hypoxia/Angiogenesis
Clinical evidence showed that hypoxia is associated with angiogenesis and a poor prognosis in patients with invasive breast cancer [38]. Other in vivo studies demonstrated that ischemia-induced angiogenesis was enhanced in RORα-deficient mice. RORα (sg/sg) mice had an increased angiogenic score and capillary density within the ischemic hindlimb, suggesting that RORα is a potential inhibitor of angiogenesis. In addition, more extensive angiogenesis correlated with an increased expression of endothelial nitric oxide synthetase (eNOS ) protein, whereas the level of the anti-angiogenic cytokine IL-12 was significantly reduced [39]. These observations suggest that RORα may participate in the control of gene transcription in response to hypoxic stress and functions as an important negative modulator of angiogenesis in breast cancer. HIF-1α is involved in tumor angiogenesis and metastasis by regulating genes involved in response to hypoxia [40]. Transcriptional activation of RORα4, but not RORα1, is induced under hypoxic conditions by HIF-1α in human hepatoma cells [41,42]. These studies suggest that RORα may be a potential target of hypoxic stress and is involved in the regulation of angiogenesis.
4.6. Role of NF-κβ
Emerging evidence demonstrates that RORα is a crucial regulator of the NF-κB pathway [43,44]. Ectopic expression of RORα in human primary smooth-muscle cells inhibits NF-κB-dependent promoter activity and NF-κB-responsive genes, such as IL-6, IL-8 and COX-2. Further analysis showed that RORα negatively interferes with the NF-κB signaling pathway by activating IκBα transcription [44]. In addition, it has been shown that NF-κB-responsive genes IL-6 and COX-2 can be up-regulated to Rev-ERBα [45], while the activity of Rev-ERBα can be competitively inhibited by RORα [46]. Transcription factor NF-κB regulates a variety of cancer related processes, including immune-response, cell survival and cancer invasion [47]. Elevated NF-κB binding activity has been observed in both breast cancer cell lines and primary human breast cancer tissues and contributes to the activation of cell-cycle related genes and various microenvironmental cues [48–50]. Thus, it is worthwhile to explore whether the RORα suppresses breast cancer progression through inhibition of the NF-κB signaling pathway.
4.7. Role of Circadian-Related Genes
Disruption of circadian rhythms is associated with an elevated risk of breast cancer [51,52]. It has been demonstrated that SNPs of NPAS2 and downregulation of PERs correlates with breast cancer development and progression [53,54]. Furthermore, PER2 deficient mice are prone to develop cancer in response to radiation [55]. These results suggest that aberrant activation of circadian genes contributes to breast cancer development. RORα-deficient mice exhibit aberrant circadian behavior, indicating that RORα is a potent regulator of circadian rhythms. It has been shown that RORα regulates Bmal1 expression and consolidates daily locomotor activity in the suprachiasmatic nucleus [56]. Moreover, RORE has been identified in the promoter regions of BMAL1 and NPAS2 [57,58], indicating that the RORα regulates circadian genes expression through the canonical pathway. However, it remains to be determined whether RORα modulates circadian rhythms in breast cancer cells and how disruption of circadian rhythms promote breast cancer progression.
4.8. Interaction with Other NR
Cross-talk with or modulation of other nuclear receptors, such as estrogen receptor (ER), is another important function of RORα. It has been shown that RORα cooperates with ER to induce cyclin D1 expression in the ER-positive breast cancer cell line MCF-7 [59]. RORα also significantly augmented the expression and activity of aromatase (an enzyme complex that catalyzes the conversion of androgens to estrogens) in MCF-7 cells [60]. Although RORα appears to be a potential ERα partner, RORα seems to be expressed differently than ER in breast cancer cells; no correlation was found between RORα expression and ERα status [61]. Interestingly, we found that RORα imparts some cancer-suppressive activities in the ER-negative breast cancer cell lines MDA-MB-231, MDA-MB-157 and T4-2, such as inhibition of cell migration and proliferation. In vivo, tumors formed by RORα-expressing MDA-MB-231 cells were also much smaller than tumors formed from the wild-type cells [5]. But, the same treatment has little effect on ER positive cell lines (data not shown). Thus RORα may have different activity in ER-positive and -negative breast cancer cells, and the mechanism whereby RORα differentially regulates cellular response in ER-positive and -negative cells remains to be elucidated.
It is most likely that tumor suppressor function of RORα is mediated by multiple pathways and involves canonical and non-canonical nuclear receptor activity. In addition, crosstalk among those pathways has been observed in vitro and in vivo; therefore, an integrated view of RORα downstream signaling is crucial for our understanding of roles of this protein in breast cancer progression.
5. ROR α as a Drug Target
RORα-targeted therapeutics may efficiently suppress certain types of tumors, thus it is crucial to identify potent ligands or agonists that have the potential to be used in cancer treatment. In fact, a recent pharmacokinetic study indicates that SR1078, a synthetic agonist for the orphan nuclear receptors RORα and RORγ, induces expression of two ROR target genes, glucose-6-phosphatase and FGF21 in mice [62]. Treatment with SR1078 enhances apoptosis of liver cancer cells in culture, suggesting that the RORα agonist may be a potent inhibitor of cancer progression [37]. In addition, melatonin, secreted by the pineal gland, has been suggested as the natural ligand for RORα [63,64]. Increasing evidence suggests that melatonin has the potential be used in breast cancer prevention and therapeutically [52,65]. Melatonin treatment induced apoptosis in the murine colonic cancer; the effect was diminished by RZR/RORα antagonist CGP 55644 [66,67]. Thus, it is important to explore whether the RORα plays a key role in melatonin-mediated inhibition of cell invasion and proliferation of breast cancer cells. Hopefully, RORα-specific, clinically-useful agonists for breast cancer treatment will be identified and tested in the future.
6. Conclusions
The orphan nuclear receptor RORα has recently been identified as a potent tumor suppressor [5,7,26,67]. Expression of RORα is downregulated in breast cancer tissues and cell lines. Restoration of RORα expression in cancer cells suppresses the malignant phenotypes in culture and in vivo[5]. Based on these observations and given the recent progress characterizing RORα agonists, further investigations of tumor suppressor activities by RORα in breast cancers may lead to the discovery of novel therapeutic targets for this mortal disease.
Acknowledgments
This study was supported by grants from NCI (P30 CA147886 to B.M. Evers), ACS (IRG 85-001-22 to R. Xu) and AHA (12SDG8600000 to R. Xu). We thank Catherine Anthony for the scientific editing.
References
- Harris, C.C. Structure and function of the p53 tumor suppressor gene: Clues for rational cancer therapeutic strategies. J. Natl. Cancer Inst 1996, 88, 1442–1455. [Google Scholar]
- Saito, Y.; Gopalan, B.; Mhashilkar, A.M.; Roth, J.A.; Chada, S.; Zumstein, L.; Ramesh, R. Adenovirus-mediated PTEN treatment combined with caffeine produces a synergistic therapeutic effect in colorectal cancer cells. Cancer Gene Ther 2003, 10, 803–813. [Google Scholar]
- Wiggins, J.F.; Ruffino, L.; Kelnar, K.; Omotola, M.; Patrawala, L.; Brown, D.; Bader, A.G. Development of a lung cancer therapeutic based on the tumor suppressor microRNA-34. Cancer Res 2010, 70, 5923–5930. [Google Scholar]
- Forouzanfar, M.H.; Foreman, K.J.; Delossantos, A.M.; Lozano, R.; Lopez, A.D.; Murray, C.J.; Naghavi, M. Breast and cervical cancer in 187 countries between 1980 and 2010: A systematic analysis. Lancet 2011, 378, 1461–1484. [Google Scholar]
- Xiong, G.; Wang, C.; Evers, B.M.; Zhou, B.P.; Xu, R. RORalpha suppresses breast tumor invasion by inducing SEMA3F expression. Cancer Res 2012, 72, 1728–1739. [Google Scholar]
- Kim, H.; Lee, J.M.; Lee, G.; Bhin, J.; Oh, S.K.; Kim, K.; Pyo, K.E.; Lee, J.S.; Yim, H.Y.; Kim, K.I.; et al. DNA damage-induced RORalpha is crucial for p53 stabilization and increased apoptosis. Mol. Cell 2011, 44, 797–810. [Google Scholar]
- Moretti, R.M.; Marelli, M.M.; Motta, M.; Polizzi, D.; Monestiroli, S.; Pratesi, G.; Limonta, P. Activation of the orphan nuclear receptor RORalpha induces growth arrest in androgen-independent DU 145 prostate cancer cells. Prostate 2001, 46, 327–335. [Google Scholar]
- Ariazi, E.A.; Ariazi, J.L.; Cordera, F.; Jordan, V.C. Estrogen receptors as therapeutic targets in breast cancer. Curr. Top. Med. Chem 2006, 6, 181–202. [Google Scholar]
- Widschwendter, M.; Berger, J.; Hermann, M.; Muller, H.M.; Amberger, A.; Zeschnigk, M.; Widschwendter, A.; Abendstein, B.; Zeimet, A.G.; Daxenbichler, G.; et al. Methylation and silencing of the retinoic acid receptor-beta2 gene in breast cancer. J. Natl. Cancer Inst 2000, 92, 826–832. [Google Scholar]
- Liu, Y.; Lee, M.O.; Wang, H.G.; Li, Y.; Hashimoto, Y.; Klaus, M.; Reed, J.C.; Zhang, X. Retinoic acid receptor beta mediates the growth-inhibitory effect of retinoic acid by promoting apoptosis in human breast cancer cells. Mol. Cell. Biol 1996, 16, 1138–1149. [Google Scholar]
- Jetten, A.M.; Kurebayashi, S.; Ueda, E. The ROR nuclear orphan receptor subfamily: Critical regulators of multiple biological processes. Prog. Nucleic Acid Res. Mol. Biol 2001, 69, 205–247. [Google Scholar]
- Becker-Andre, M.; Andre, E.; DeLamarter, J.F. Identification of nuclear receptor mRNAs by RT-PCR amplification of conserved zinc-finger motif sequences. Biochem. Biophys. Res. Commun 1993, 194, 1371–1379. [Google Scholar]
- Giguere, V.; Tini, M.; Flock, G.; Ong, E.; Evans, R.M.; Otulakowski, G. Isoform-specific amino-terminal domains dictate DNA-binding properties of ROR alpha, a novel family of orphan hormone nuclear receptors. Genes Dev 1994, 8, 538–553. [Google Scholar]
- Hamilton, B.A.; Frankel, W.N.; Kerrebrock, A.W.; Hawkins, T.L.; FitzHugh, W.; Kusumi, K.; Russell, L.B.; Mueller, K.L.; van Berkel, V.; Birren, B.W.; et al. Disruption of the nuclear hormone receptor RORalpha in staggerer mice. Nature 1996, 379, 736–739. [Google Scholar]
- Steinmayr, M.; Andre, E.; Conquet, F.; Rondi-Reig, L.; Delhaye-Bouchaud, N.; Auclair, N.; Daniel, H.; Crepel, F.; Mariani, J.; Sotelo, C.; et al. staggerer phenotype in retinoid-related orphan receptor alpha-deficient mice. Proc. Natl. Acad. Sci. USA 1998, 95, 3960–3965. [Google Scholar]
- Jarvis, C.I.; Staels, B.; Brugg, B.; Lemaigre-Dubreuil, Y.; Tedgui, A.; Mariani, J. Age-related phenotypes in the staggerer mouse expand the RORalpha nuclear receptor’s role beyond the cerebellum. Mol. Cell. Endocrinol 2002, 186, 1–5. [Google Scholar]
- Wang, N.S.; McHeyzer-Williams, L.J.; Okitsu, S.L.; Burris, T.P.; Reiner, S.L.; McHeyzer-Williams, M.G. Divergent transcriptional programming of class-specific B cell memory by T-bet and RORalpha. Nat. Immunol 2012, 13, 604–611. [Google Scholar]
- Wong, S.H.; Walker, J.A.; Jolin, H.E.; Drynan, L.F.; Hams, E.; Camelo, A.; Barlow, J.L.; Neill, D.R.; Panova, V.; Koch, U.; et al. Transcription factor RORalpha is critical for nuocyte development. Nat. Immunol 2012, 13, 229–236. [Google Scholar]
- Kang, H.S.; Okamoto, K.; Takeda, Y.; Beak, J.Y.; Gerrish, K.; Bortner, C.D.; DeGraff, L.M.; Wada, T.; Xie, W.; Jetten, A.M. Transcriptional profiling reveals a role for RORalpha in regulating gene expression in obesity-associated inflammation and hepatic steatosis. Physiol. Genomics 2011, 43, 818–828. [Google Scholar]
- Jetten, A.M. Retinoid-related orphan receptors (RORs): Critical roles in development, immunity, circadian rhythm, and cellular metabolism. Nucl. Recept. Signal 2009, 7, e003. [Google Scholar]
- Kallen, J.; Schlaeppi, J.M.; Bitsch, F.; Delhon, I.; Fournier, B. Crystal structure of the human RORalpha Ligand binding domain in complex with cholesterol sulfate at 2.2 A. J. Biol. Chem 2004, 279, 14033–14038. [Google Scholar]
- Kallen, J.A.; Schlaeppi, J.M.; Bitsch, F.; Geisse, S.; Geiser, M.; Delhon, I.; Fournier, B. X-ray structure of the hRORalpha LBD at 1.63 A: Structural and functional data that cholesterol or a cholesterol derivative is the natural ligand of RORalpha. Structure 2002, 10, 1697–1707. [Google Scholar]
- Zenri, F.; Hiroi, H.; Momoeda, M.; Tsutsumi, R.; Hosokawa, Y.; Koizumi, M.; Nakae, H.; Osuga, Y.; Yano, T.; Taketani, Y. Expression of retinoic acid-related orphan receptor alpha and its responsive genes in human endometrium regulated by cholesterol sulfate. J. Steroid Biochem. Mol. Biol 2012, 128, 21–28. [Google Scholar]
- Zhu, Y.; McAvoy, S.; Kuhn, R.; Smith, D.I. RORA, a large common fragile site gene, is involved in cellular stress response. Oncogene 2006, 25, 2901–2908. [Google Scholar]
- Yamashita, S.; Tsujino, Y.; Moriguchi, K.; Tatematsu, M.; Ushijima, T. Chemical genomic screening for methylation-silenced genes in gastric cancer cell lines using 5-aza-2′-deoxycytidine treatment and oligonucleotide microarray. Cancer Sci 2006, 97, 64–71. [Google Scholar]
- Lee, J.M.; Kim, I.S.; Kim, H.; Lee, J.S.; Kim, K.; Yim, H.Y.; Jeong, J.; Kim, J.H.; Kim, J.Y.; Lee, H.; et al. RORalpha attenuates Wnt/beta-catenin signaling by PKCalpha-dependent phosphorylation in colon cancer. Mol. Cell 2010, 37, 183–195. [Google Scholar]
- Dai, J.; Ram, P.T.; Yuan, L.; Spriggs, L.L.; Hill, S.M. Transcriptional repression of RORalpha activity in human breast cancer cells by melatonin. Mol. Cell. Endocrinol 2001, 176, 111–120. [Google Scholar]
- Lu, Y.; Yi, Y.; Liu, P.; Wen, W.; James, M.; Wang, D.; You, M. Common human cancer genes discovered by integrated gene-expression analysis. PLoS One 2007, 2, e1149. [Google Scholar]
- Shimizu, A.; Mammoto, A.; Italiano, J.E., Jr; Pravda, E.; Dudley, A.C.; Ingber, D.E.; Klagsbrun, M. ABL2/ARG tyrosine kinase mediates SEMA3F-induced RhoA inactivation and cytoskeleton collapse in human glioma cells. J. Biol. Chem. 2008, 283, 27230–27238. [Google Scholar]
- Potiron, V.A.; Sharma, G.; Nasarre, P.; Clarhaut, J.A.; Augustin, H.G.; Gemmill, R.M.; Roche, J.; Drabkin, H.A. Semaphorin SEMA3F affects multiple signaling pathways in lung cancer cells. Cancer Res 2007, 67, 8708–8715. [Google Scholar]
- Benhaj, K.; Akcali, K.C.; Ozturk, M. Redundant expression of canonical Wnt ligands in human breast cancer cell lines. Oncol. Rep 2006, 15, 701–707. [Google Scholar]
- Yan, L.; Della Coletta, L.; Powell, K.L.; Shen, J.; Thames, H.; Aldaz, C.M.; MacLeod, M.C. Activation of the canonical Wnt/beta-catenin pathway in ATF3-induced mammary tumors. PLoS One 2011, 6, e16515. [Google Scholar]
- Kremenevskaja, N.; von Wasielewski, R.; Rao, A.S.; Schofl, C.; Andersson, T.; Brabant, G. Wnt-5a has tumor suppressor activity in thyroid carcinoma. Oncogene 2005, 24, 2144–2154. [Google Scholar]
- Leris, A.C.; Roberts, T.R.; Jiang, W.G.; Newbold, R.F.; Mokbel, K. WNT5A expression in human breast cancer. Anticancer Res 2005, 25, 731–734. [Google Scholar]
- Knappskog, S.; Lonning, P.E. p53 and its molecular basis to chemoresistance in breast cancer. Expert Opin. Ther. Targets 2012, 16, S23–S30. [Google Scholar]
- Lacroix, M.; Toillon, R.A.; Leclercq, G. p53 and breast cancer, an update. Endocr. Relat. Cancer 2006, 13, 293–325. [Google Scholar]
- Wang, Y.; Solt, L.A.; Kojetin, D.J.; Burris, T.P. Regulation of p53 stability and apoptosis by a ROR agonist. PLoS One 2012, 7, e34921. [Google Scholar]
- Pugh, C.W.; Gleadle, J.; Maxwell, P.H. Hypoxia and oxidative stress in breast cancer. Hypoxia signalling pathways. Breast Cancer Res 2001, 3, 313–317. [Google Scholar]
- Besnard, S.; Silvestre, J.S.; Duriez, M.; Bakouche, J.; Lemaigre-Dubreuil, Y.; Mariani, J.; Levy, B.I.; Tedgui, A. Increased ischemia-induced angiogenesis in the staggerer mouse, a mutant of the nuclear receptor Roralpha. Circ. Res 2001, 89, 1209–1215. [Google Scholar]
- Dales, J.P.; Garcia, S.; Meunier-Carpentier, S.; Andrac-Meyer, L.; Haddad, O.; Lavaut, M.N.; Allasia, C.; Bonnier, P.; Charpin, C. Overexpression of hypoxia-inducible factor HIF-1alpha predicts early relapse in breast cancer: retrospective study in a series of 745 patients. Int. J. Cancer 2005, 116, 734–739. [Google Scholar]
- Miki, N.; Ikuta, M.; Matsui, T. Hypoxia-induced activation of the retinoic acid receptor-related orphan receptor alpha4 gene by an interaction between hypoxia-inducible factor-1 and Sp1. J. Biol. Chem 2004, 279, 15025–15031. [Google Scholar]
- Chauvet, C.; Bois-Joyeux, B.; Danan, J.L. Retinoic acid receptor-related orphan receptor (ROR) alpha4 is the predominant isoform of the nuclear receptor RORalpha in the liver and is up-regulated by hypoxia in HepG2 human hepatoma cells. Biochem. J 2002, 364, 449–456. [Google Scholar]
- Journiac, N.; Jolly, S.; Jarvis, C.; Gautheron, V.; Rogard, M.; Trembleau, A.; Blondeau, J.P.; Mariani, J.; Vernet-der Garabedian, B. The nuclear receptor ROR(alpha) exerts a bi-directional regulation of IL-6 in resting and reactive astrocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 21365–21370. [Google Scholar]
- Delerive, P.; Monte, D.; Dubois, G.; Trottein, F.; Fruchart-Najib, J.; Mariani, J.; Fruchart, J.C.; Staels, B. The orphan nuclear receptor ROR alpha is a negative regulator of the inflammatory response. EMBO Rep 2001, 2, 42–48. [Google Scholar]
- Migita, H.; Morser, J.; Kawai, K. Rev-erbalpha upregulates NF-kappaB-responsive genes in vascular smooth muscle cells. FEBS Lett 2004, 561, 69–74. [Google Scholar]
- Guillaumond, F.; Dardente, H.; Giguere, V.; Cermakian, N. Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors. J. Biol. Rhythms 2005, 20, 391–403. [Google Scholar]
- Hayden, M.S.; Ghosh, S. Signaling to NF-kappaB. Genes Dev 2004, 18, 2195–2224. [Google Scholar]
- Nakshatri, H.; Bhat-Nakshatri, P.; Martin, D.A.; Goulet, R.J., Jr; Sledge, G.W., Jr. Constitutive activation of NF-kappaB during progression of breast cancer to hormone-independent growth. Mol. Cell. Biol. 1997, 17, 3629–3639. [Google Scholar]
- Huber, M.A.; Azoitei, N.; Baumann, B.; Grunert, S.; Sommer, A.; Pehamberger, H.; Kraut, N.; Beug, H.; Wirth, T. NF-kappaB is essential for epithelial-mesenchymal transition and metastasis in a model of breast cancer progression. J. Clin. Invest 2004, 114, 569–581. [Google Scholar]
- Cao, Y.; Karin, M. NF-kappaB in mammary gland development and breast cancer. J. Mammary Gland Biol. Neoplasia 2003, 8, 215–223. [Google Scholar]
- Sahar, S.; Sassone-Corsi, P. Circadian clock and breast cancer: A molecular link. Cell Cycle 2007, 6, 1329–1331. [Google Scholar]
- Stevens, R.G. Circadian disruption and breast cancer: from melatonin to clock genes. Epidemiology 2005, 16, 254–258. [Google Scholar]
- Zhu, Y.; Stevens, R.G.; Leaderer, D.; Hoffman, A.; Holford, T.; Zhang, Y.; Brown, H.N.; Zheng, T. Non-synonymous polymorphisms in the circadian gene NPAS2 and breast cancer risk. Breast Cancer Res. Treat 2008, 107, 421–425. [Google Scholar]
- Winter, S.L.; Bosnoyan-Collins, L.; Pinnaduwage, D.; Andrulis, I.L. Expression of the circadian clock genes Per1 and Per2 in sporadic and familial breast tumors. Neoplasia 2007, 9, 797–800. [Google Scholar]
- Fu, L.; Pelicano, H.; Liu, J.; Huang, P.; Lee, C. The circadian gene Period2 plays an important role in tumor suppression and DNA damage response in vivo. Cell 2002, 111, 41–50. [Google Scholar]
- Sato, T.K.; Panda, S.; Miraglia, L.J.; Reyes, T.M.; Rudic, R.D.; McNamara, P.; Naik, K.A.; FitzGerald, G.A.; Kay, S.A.; Hogenesch, J.B. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron 2004, 43, 527–537. [Google Scholar]
- Akashi, M.; Takumi, T. The orphan nuclear receptor RORalpha regulates circadian transcription of the mammalian core-clock Bmal1. Nat. Struct. Mol. Biol 2005, 12, 441–448. [Google Scholar]
- Crumbley, C.; Wang, Y.; Kojetin, D.J.; Burris, T.P. Characterization of the core mammalian clock component, NPAS2, as a REV-ERBalpha/RORalpha target gene. J. Biol. Chem 2010, 285, 35386–35392. [Google Scholar]
- Dong, C.; Yuan, L.; Dai, J.; Lai, L.; Mao, L.; Xiang, S.; Rowan, B.; Hill, S.M. Melatonin inhibits mitogenic cross-talk between retinoic acid-related orphan receptor alpha (RORalpha) and ERalpha in MCF-7 human breast cancer cells. Steroids 2010, 75, 944–951. [Google Scholar]
- Odawara, H.; Iwasaki, T.; Horiguchi, J.; Rokutanda, N.; Hirooka, K.; Miyazaki, W.; Koibuchi, Y.; Shimokawa, N.; Iino, Y.; Takeyoshi, I.; et al. Activation of aromatase expression by retinoic acid receptor-related orphan receptor (ROR) alpha in breast cancer cells: Identification of a novel ROR response element. J. Biol. Chem 2009, 284, 17711–17719. [Google Scholar]
- Gu, F.; Hsu, H.K.; Hsu, P.Y.; Wu, J.; Ma, Y.; Parvin, J.; Huang, T.H.; Jin, V.X. Inference of hierarchical regulatory network of estrogen-dependent breast cancer through ChIP-based data. BMC Syst. Biol 2010, 4, 170. [Google Scholar]
- Wang, Y.; Kumar, N.; Nuhant, P.; Cameron, M.D.; Istrate, M.A.; Roush, W.R.; Griffin, P.R.; Burris, T.P. Identification of SR1078, a synthetic agonist for the orphan nuclear receptors RORalpha and RORgamma. ACS Chem. Biol 2010, 5, 1029–1034. [Google Scholar]
- Wiesenberg, I.; Missbach, M.; Kahlen, J.P.; Schrader, M.; Carlberg, C. Transcriptional activation of the nuclear receptor RZR alpha by the pineal gland hormone melatonin and identification of CGP 52608 as a synthetic ligand. Nucleic Acids Res 1995, 23, 327–333. [Google Scholar]
- Carlberg, C. Gene regulation by melatonin. Ann. NY Acad. Sci 2000, 917, 387–396. [Google Scholar]
- Korkmaz, A.; Sanchez-Barcelo, E.J.; Tan, D.X.; Reiter, R.J. Role of melatonin in the epigenetic regulation of breast cancer. Breast Cancer Res. Treat 2009, 115, 13–27. [Google Scholar]
- Winczyk, K.; Pawlikowski, M.; Karasek, M. Melatonin and RZR/ROR receptor ligand CGP 52608 induce apoptosis in the murine colonic cancer. J. Pineal Res 2001, 31, 179–182. [Google Scholar]
- Winczyk, K.; Pawlikowski, M.; Guerrero, J.M.; Karasek, M. Possible involvement of the nuclear RZR/ROR-alpha receptor in the antitumor action of melatonin on murine Colon 38 cancer. Tumor Biol 2002, 23, 298–302. [Google Scholar]
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).