Inflammation, Oxidative Stress, and Obesity

 , , ,
, , ,
Abstract
:1. Introduction
2. Etiological Factors
3. Epidemiology
4. Adipose Tissue
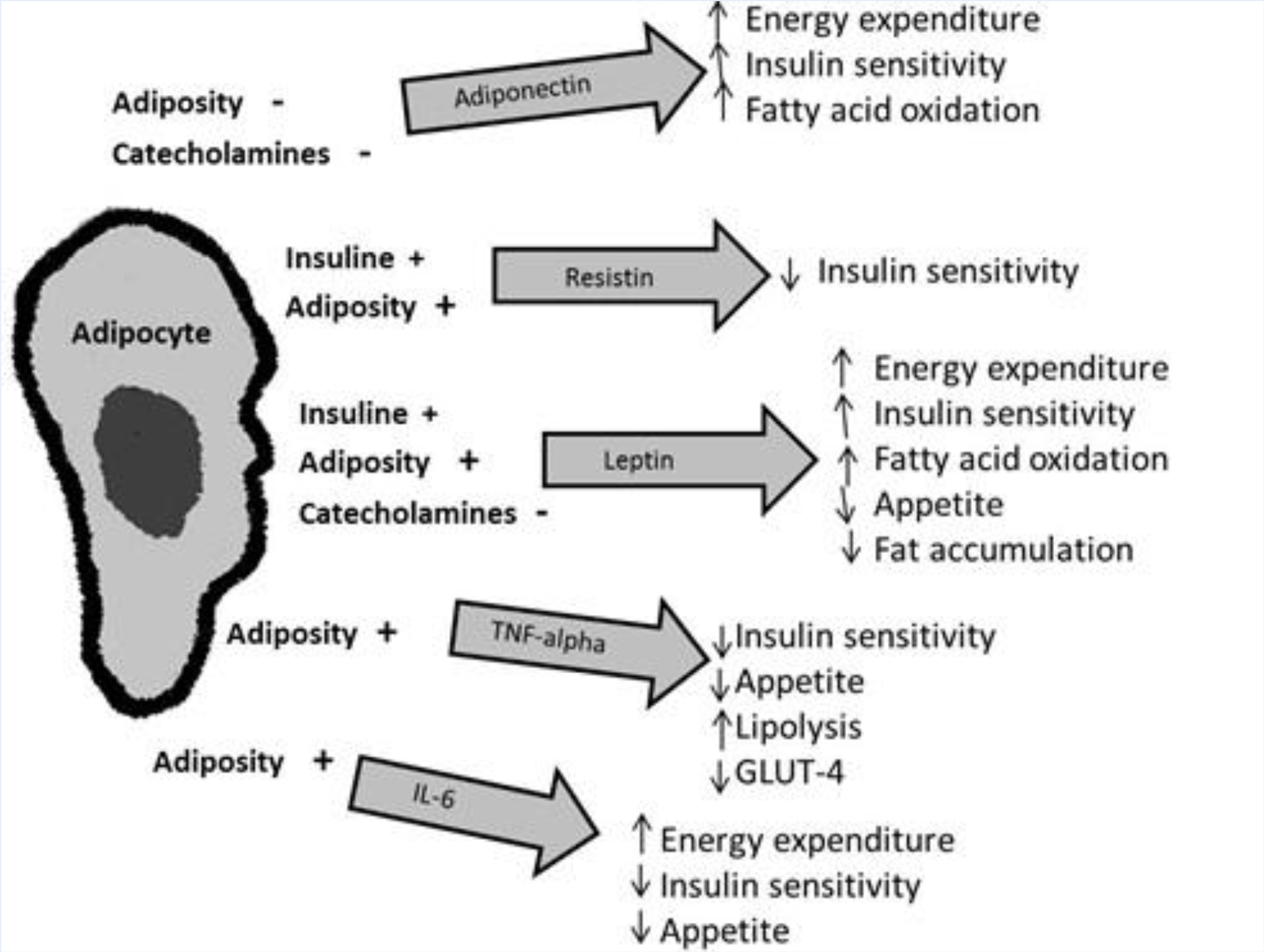
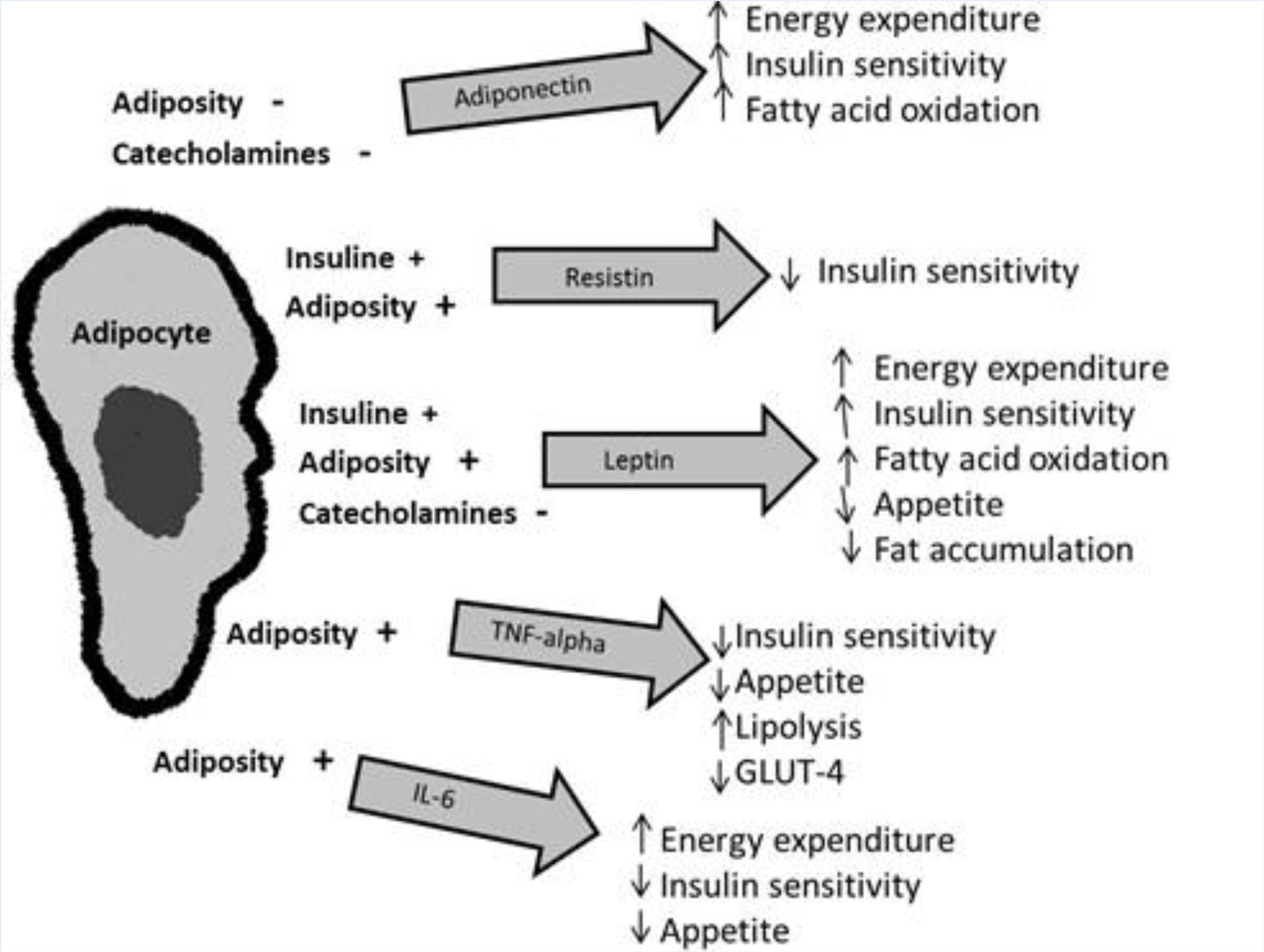
5. Adipokines and Metabolic Homeostasis
5.1. Leptin
5.2. Tumor Necrosis Factor Alpha (TNF-α)
5.3. Interleukin 6 (IL-6)
5.4. Angiotensinogen/PAI-1
5.5. Adiponectin
5.6. Adipsin
5.7. Resistin
5.8. Other Adipokines
6. Lipotoxicity
7. Obesity and Oxidative Stress
- Peroxisomal fatty acid metabolism, in which H2O2 is formed as a byproduct, and despite that peroxisomes contain high catalase activity, they may cause OS under certain pathological conditions.
- Cytochrome P450 microsomal reactions, which catalyze the metabolism of xenobiotic compounds by oxidoreducers, forming superoxide anion as a byproduct, which can cause OS.
- Phagocyte cells, which attack invasive pathogens with a mixture of ROS and other oxidants. This is an immune response, but also damages surrounding tissues, producing inflammation.
- The mitochondrial respiratory chain. It is considered that the mitochondria are the site within the cell where the largest amount of ROS are generated, causing defects in mitochondrial metabolism and diseases.
8. Mechanisms of Formation of Free Radicals during Obesity
8.1. Adipose Tissue
8.2. Fatty Acid Oxidation
8.3. Overconsumption of Oxygen
8.4. Accumulation of Cellular Damage
8.5. Type of Diet
8.6. Role of Mitochondria in the Development of OS in Obesity
9. Complications-generated Oxidative Stress in Obesity
10. Obesity and Antioxidant Capacity
11. Nitric Oxide in Obesity
12. Inflammation and Obesity
13. Endothelial Dysfunction
14. Conclusions
References
- Kaufer, M; Tavano, L; Ávila, H. Obesidad en el adulto. In Nutriología Médica, 1st ed; Casanueva, E, Kaufer, M, Pérez, A, Arroyo, P, Eds.; Editorial Médica Panamericana: México, México, 2001. [Google Scholar]
- Sikaris, K. The clinical biochemistry of obesity. Clin. Biochem. Rev 2004, 25, 165–181. [Google Scholar]
- Lastra, G; Manrique, CM; Hayden, MR. The role of beta-cell dysfunction in the cardiometabolic syndrome. J. Cardiometab. Syndr 2006, 1, 41–46. [Google Scholar]
- Barquera, S. Obesidad: La epidemia mundial. In Sobrepeso y Obesidad, 1st ed; Barquera, S, Tolentino, L, Rivera, J, Eds.; Instituto Nacional de Salud Pública: México, México, 2006. [Google Scholar]
- Bravo, P; Morse, S; Borne, D; Aguílar, E; Reisin, E. Leptin and hypertension in obesity. Vasc. Health Risk Manage 2006, 2, 163–169. [Google Scholar]
- Amirkhizi, F; Siassi, F; Minaie, S; Djalali, M; Rahimi, A; Chamari, M. Is obesity associated with increased plasma lipid peroxidación and oxidative stress in women. ARYA Atheroscler. J 2007, 2, 189–192. [Google Scholar]
- Chan, RS; Woo, J. Prevention of overweight and obesity: How effective is the current public health approach. Int. J. Environ. Res. Public Health 2010, 7, 765–783. [Google Scholar]
- Dulloo, AG; Jacquet, J; Solinas, G; Montani, JP; Schutz, Y. Body composition phenotypes in pathways to obesity and the metabolic syndrome. Int J Obes 2010, 34(Suppl. 2), S4–S17. [Google Scholar]
- Sánchez, F; García, R; Alarcón, F; Cruz, M. Adipocinas, tejido adiposo y su relación con células del sistema inmune. Gac. Méd. Méx 2005, 141, 505–512. [Google Scholar]
- Deng, Y; Scherer, PE. Adipokines as novel biomarkers and regulators of the metabolic syndrome. Ann. N. Y. Acad. Sci 2010, 1212, E1–E19. [Google Scholar]
- Fonseca-Alaniz, MH; Takada, J; Alonso-Vale, MI; Lima, FB. Adipose tissue as an endocrine organ: From theory to practice. J Pediatr 2007, 83(Suppl. 5), S192–S203. [Google Scholar]
- Steffes, M; Gross, M; Lee, D; Schreiner, P; Jacobs, D. Adiponectin, visceral fat, oxidative stress and early macrovascular disease: The coronary artery risk development in young adults study. Obesity 2006, 14, 319–326. [Google Scholar]
- Hukshorn, CJ; Lindeman, JH; Toet, KH; Saris, WH; Eilers, PH; Westerterp-Plantenga, MS; Kooistra, T. Leptin and the proinflammatory state associated with human obesity. J. Clin. Endocrinol. Metab 2004, 89, 1773–1778. [Google Scholar]
- Cachofeiro, V; Miana, M; Martín, B. Obesidad, inflamación y disfunción endotelial. Rev. Esp. Obes 2006, 4, 195–204. [Google Scholar]
- Ouchi, N; Parker, JL; Lugus, JJ; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol 2011, 11, 85–97. [Google Scholar]
- Ouedraogo, R; Gong, Y; Berzins, B; Wu, X; Mahadev, K; Hough, K; Chan, L; Goldstein, BJ; Scalia, R. Adiponectin deficiency increases leukocyte-endothelium interactions via upregulation of endothelial cell adhesion molecules in vivo. J. Clin. Invest 2007, 117, 1718–1761. [Google Scholar]
- Recansens, M; Ricart, W; Fernández, J. Obesidad e inflamación. Rev. Med. Univ. Navarra 2004, 48, 49–54. [Google Scholar]
- Pyrzak, B; Ruminska, M; Popko, K; Demkow, U. Adiponectin as a biomarker of the metabolic syndrome in children and adolescents. Eur J Med Res 2010, 15(Suppl.), 147–151. [Google Scholar]
- Steppan, CM; Bailey, ST; Bhat, S; Brown, EJ; Banerjee, RR; Wright, CM; Patel, HR; Ahima, RS; Lazar, MA. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar]
- Steppan, CM; Lazar, MA. The current biology of resistin. J. Int. Med 2004, 255, 439–447. [Google Scholar]
- Sonoli, SS; Shivprasad, S; Prasad, CV; Patil, AB; Desai, PB; Somannavar, MS. Visfatin—A review. Eur. Rev. Med. Pharmacol. Sci 2011, 15, 9–14. [Google Scholar]
- Barth, S; Klein, P; Horbach, T; Dötsch, J; Rauh, M; Rascher, W; Knerr, I. Expression of neuropeptide Y, omentin and visfatin in visceral and subcutaneous adipose tissues in humans: Relation to endocrine and clinical parameters. Obes. Facts 2010, 3, 245–251. [Google Scholar]
- Kluth, O; Mirhashemi, F; Scherneck, S; Kaiser, D; Kluge, R; Neschen, S; Joost, HG; Schürmann, A. Dissociation of lipotoxicity and glucotoxicity in a mouse model of obesity associated diabetes: Role of forkhead box O1 (FOXO1) in glucose-induced beta cell failure. Diabetologia 2011, 54, 605–616. [Google Scholar]
- Esposito, K; Ciotola, M; Giugliano, D. Oxidative stress in the Metabolic Syndrome. J. Endocrinol. Invest 2006, 29, 791–795. [Google Scholar]
- Pihl, E; Zilmer, K; Kullisaar, T; Kairane, C; Magi, A; Zilmer, M. Atherogenic inflammatory and oxidative stress markers in relation to overweight values in male former athletes. Int. J. Obesity 2006, 30, 141–146. [Google Scholar]
- Chrysohoou, C; Panagiotakos, DB; Pitsavos, C; Skoumas, I; Papademetriou, L; Economou, M; Stefanadis, C. The implication of obesity on total antioxidant capacity apparently healthy men and women: The ATTICA study. Nutr. Metab. Cardiovasc. Dis 2007, 17, 590–597. [Google Scholar]
- Hartwich, J; Goralska, J; Siedlecka, D; Gruca, A; Trzos, M; Dembinska-Kiec, A. Effec of supplementation with vitamin E and C on plasma hsCPR level and cobalt-albumin binding score as markers of plasma oxidative stress in obesity. Genes Nutr 2007, 2, 151–154. [Google Scholar]
- Patel, C; Ghanim, H; Ravishankar, S; Sia, CL; Viswanathan, P; Mohantym, P; Dandona, P. Prolonged reactive oxygen species generation and Nuclear Factor- kB activation after a high-fat, high-carbohydrate meal in the obese. J. Clin. Endocrinol. Metab 2007, 92, 4476–4479. [Google Scholar]
- Block, G; Dietrich, M; Norkus, EP; Morrow, JD; Hudes, M; Caan, B; Packer, L. Factors associated with oxidative stress in human populations. Am. J. Epidemiol 2002, 156, 274–285. [Google Scholar]
- Keaney, JF, Jr; Larson, MG; Vasan, RS; Wilson, PWF; Lipinska, I; Corey, D; Massaro, JM; Sutherland, P; Vita, JA; Benjamin, EJ. Obesity and systemic oxidative stress: Clinical correlates of oxidative stress in the Framingham study. Arterioscler. Tromb. Vasc. Biol 2003, 23, 434–439. [Google Scholar]
- Morrow, J. Is a oxidative stress a connection between obesity and atherosclerosis. Arterioscler. Tromb. Vasc. Biol 2003, 23, 368–370. [Google Scholar]
- Duvnjak, M; Lerotic, I; Barsic, N; Tomasic, V; Jukic, L; Velagic, V. Pathogenesis and management issues for non-alcoholic fatty liver disease. World. J. Gastroenterol 2007, 13, 4539–4550. [Google Scholar]
- Khan, N; Naz, L; Yasmeen, G. Obesity: An independent risk factor systemic oxidative stress. Park. J. Pharm. Sci 2006, 19, 62–69. [Google Scholar]
- Monteiro, R; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediators. Inflamm 2010, 2010, 289645. [Google Scholar]
- Martínez, J. Mitocondrial oxidative stress and inflammation: A slalom to obesity and insulin resistance. J. Physiol. Biochem 2006, 62, 303–306. [Google Scholar]
- Mainese, K; Morhan, S; Chong, Z. Oxidative stress biology and cell injury during type 1 and 2 diabetes mellitus. Curr. Neoruvasc. Res 2007, 4, 63–71. [Google Scholar]
- Ozata, M; Mergen, M; Oktenli, C; Aydin, A; Sanisoglu, SY; Bolu, E; Yilmaz, MI; Sayal, A; Isimer, A; Ozdemir, IC. Increased oxidative stress and hypozincemia in male obesity. Clin. Biochem 2002, 35, 627–631. [Google Scholar]
- Capel, I; Dorrell, H. Abnormal antioxidant defense in some tissues of congenitally obese mice. Biochemistry 1984, 219, 41–49. [Google Scholar]
- Vincent, H; Vincent, K; Vourguignon, C; Braith, R. Obesity and postexercise oxidative stress in older women. Med. Sci. Sports Exer 2005, 37, 213–219. [Google Scholar]
- Furukawa, S; Fujita, T; Shimabukuro, M; Iwaki, M; Yamada, Y; Nakajima, Y; Nakayama, O; Makishima, M; Matsuda, M; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Invest 2004, 114, 1752–1761. [Google Scholar]
- Higdon, J; Frei, B. Obesity and oxidative stress: A direct link to CVD? Arterioscler. Tromb. Vasc. Biol 2003, 23, 365–367. [Google Scholar]
- Shimabukuro, M; Ohneda, M; Lee, Y; Unger, R. Role of nitric oxide in obesity-induced β cell disease. J. Clin. Invest 1997, 100, 290–295. [Google Scholar]
- Chakraborty, K; Khan, GA; Banerjee, P; Ray, U; Sinha, AK. Inhibition of human blood platelet aggregation and the stimulation of nitric oxide synthesis by aspirin. Platelets 2003, 14, 421–427. [Google Scholar]
- De Souza, C; Van Guilder, G; Greiner, J; Smith, D; Hoetzer, G; Stauffer, B. Basal endothelial nitric oxide release is preserved in overweight and obese adults. Obes. Res 2005, 13, 1303–1306. [Google Scholar]
- Dobrian, A; Schriver, S; Lynch, T; Prewitt, R. Effect of salt on hypertension and oxidative stress in a rat model of diet-induced obesity. Am. J. Physiol. Renal. Physiol 2003, 285, 619–628. [Google Scholar]
- Flores, M; Barquera, S; Carrión, C. Inflamación obesidad y diabetes mellitus tipo 2. In Sobrepeso y Obesidad, 1st ed; Barquera, S, Tolentino, L, Rivera, J, Eds.; Instituto Nacional de Salud Pública: México, México, 2006. [Google Scholar]
- Bastarrachea, R; López, J; Bolado, N; Téllez, J; Laviada, H; Comuzzie, A. Macrófagos, inflamación, tejido adiposo, obesidad y resistencia a la insulina. Gac. Méd. Méx 2007, 143, 505–512. [Google Scholar]
- Nishimura, S; Manabe, I; Nagasaki, M; Eto, K; Yamashita, H; Ohsugi, M; Otsu, M; Hara, K; Ueki, K; Sugiura, S; Yoshimura, K; Kadowaki, T; Nagai, R. CD8 + effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med 2009, 15, 914–920. [Google Scholar]
- Cani, PD; Bibiloni, R; Knauf, C; Waget, A; Neyrinck, AM; Delzenne, NM; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar]
- Hadi, H; Carr, C; Suwaidi, J. Endothelial dysfunction: Cardiovascular risk factors, therapy, and outcome. Vasc. Health Risk Manage 2005, 1, 183–198. [Google Scholar]
- Couillard, C; Ruel, G; Archer, WR; Pomerleau, S; Bergeron, J; Couture, P; Lamarche, B; Bergeron, N. Circulating levels of oxidative stress markers and endotelial adhesión molecules in men with abdominal obesity. J. Clin. Endocrinol. Metab 2005, 90, 6454–6459. [Google Scholar]
- Galili, O; Versari, D; Sattler, KJ; Olson, ML; Mannheim, D; McConnell, JP; Chade, AR; Lerman, LO; Lerman, A. Early experimental obesity is associated with endothelial dysfunction and oxidative stress. Am J Physiol Heart Circ Physiol 2007, 292, H904–H911. [Google Scholar]

{kind=link}
| Insulin resistance and diabetes |
| Systemic arterial hypertension |
| Ischemic heart diseases |
| Obstructive sleep apnea, asthma |
| Gout |
| Peripheral vascular disease |
| Psychology problems (social stigmatization) |
| Rheumatological and orthopedics problems |
| Oncology problems |
| Liver failure |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, Oxidative Stress, and Obesity. Int. J. Mol. Sci. 2011, 12, 3117-3132. https://doi.org/10.3390/ijms12053117
Fernández-Sánchez A, Madrigal-Santillán E, Bautista M, Esquivel-Soto J, Morales-González Á, Esquivel-Chirino C, Durante-Montiel I, Sánchez-Rivera G, Valadez-Vega C, Morales-González JA. Inflammation, Oxidative Stress, and Obesity. International Journal of Molecular Sciences. 2011; 12(5):3117-3132. https://doi.org/10.3390/ijms12053117
Chicago/Turabian StyleFernández-Sánchez, Alba, Eduardo Madrigal-Santillán, Mirandeli Bautista, Jaime Esquivel-Soto, Ángel Morales-González, Cesar Esquivel-Chirino, Irene Durante-Montiel, Graciela Sánchez-Rivera, Carmen Valadez-Vega, and José A. Morales-González. 2011. "Inflammation, Oxidative Stress, and Obesity" International Journal of Molecular Sciences 12, no. 5: 3117-3132. https://doi.org/10.3390/ijms12053117
APA StyleFernández-Sánchez, A., Madrigal-Santillán, E., Bautista, M., Esquivel-Soto, J., Morales-González, Á., Esquivel-Chirino, C., Durante-Montiel, I., Sánchez-Rivera, G., Valadez-Vega, C., & Morales-González, J. A. (2011). Inflammation, Oxidative Stress, and Obesity. International Journal of Molecular Sciences, 12(5), 3117-3132. https://doi.org/10.3390/ijms12053117