Effects of Soybean Agglutinin on Intestinal Barrier Permeability and Tight Junction Protein Expression in Weaned Piglets

Abstract
:1. Introduction
2. Results and Discussion
2.1. Purification of Glycinin and β-Conglycinin
2.2. Experimental Design
2.3. Determination of d-Lactic Acid in Plasma
2.4. Determination of Diamine Oxidase (DAO) in Ileal Mucosa
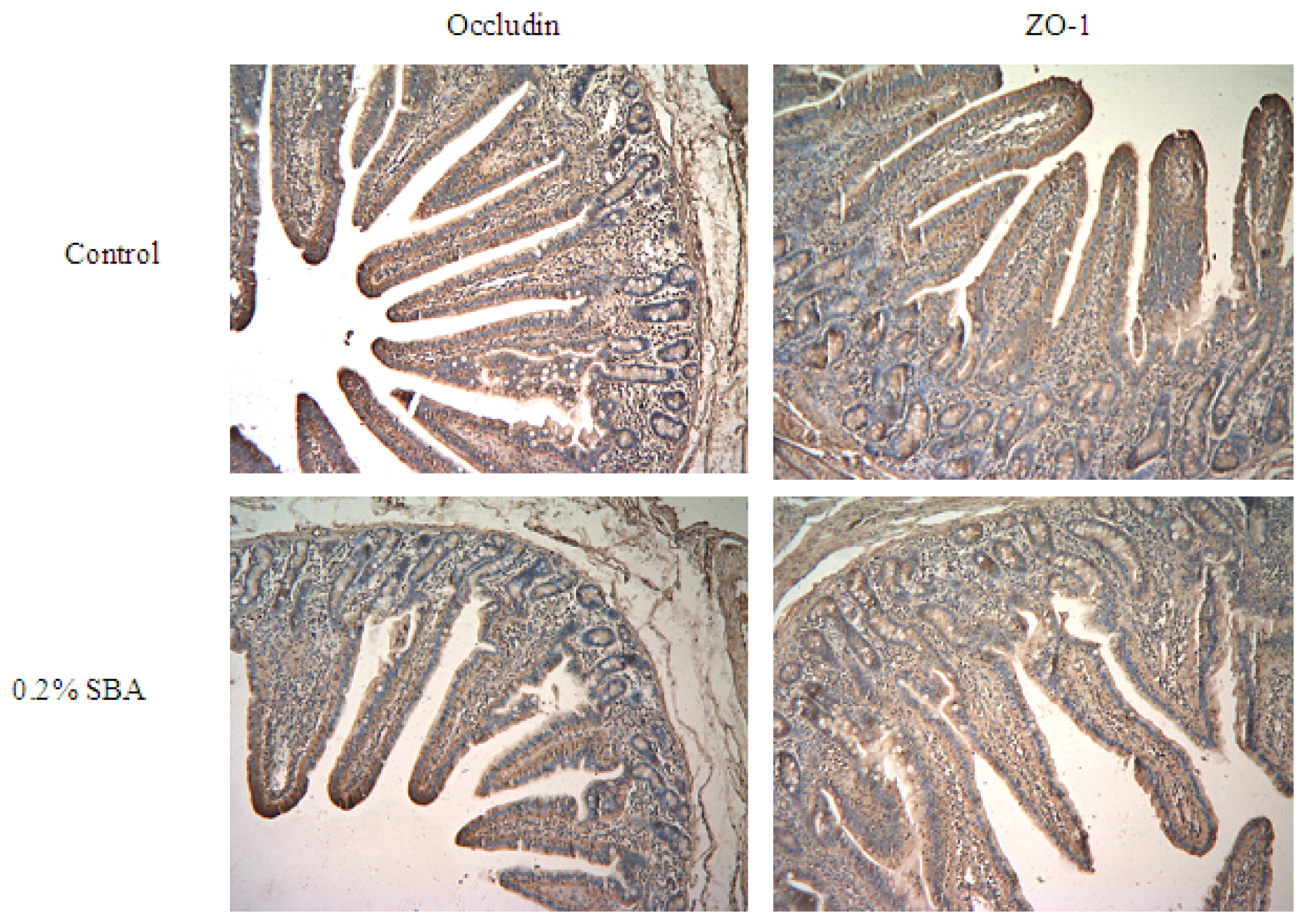
2.5. Immunohistochemistry
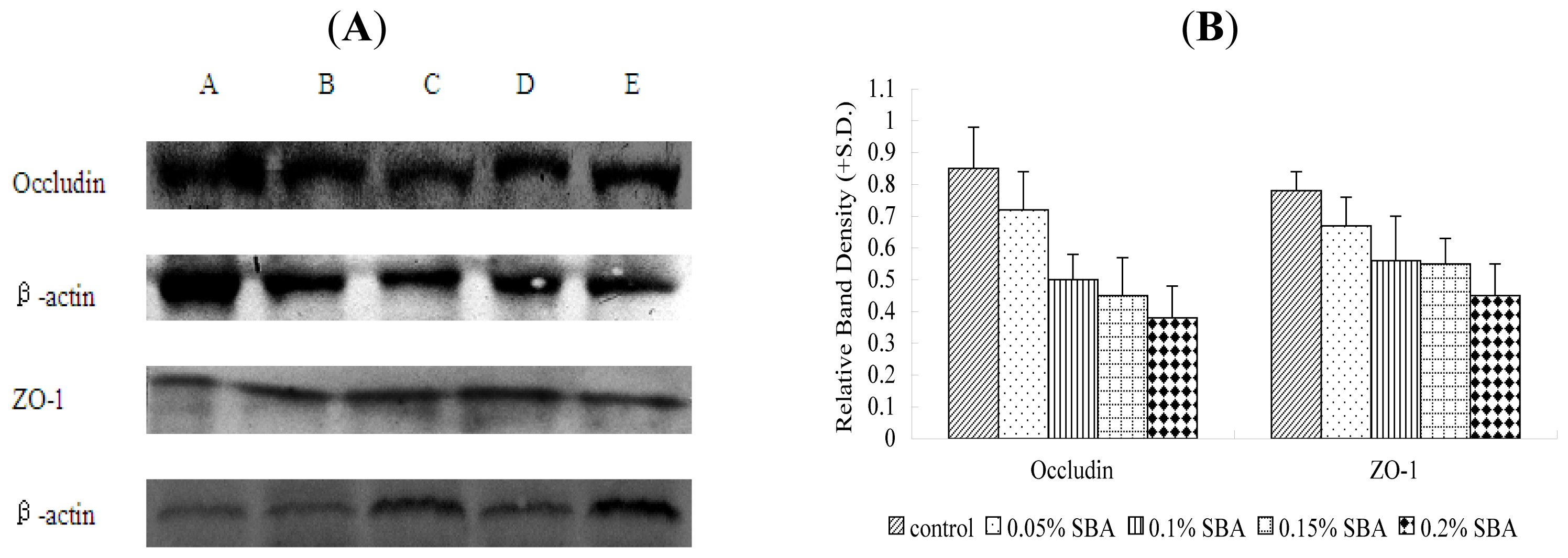
2.6. Immunoblotting
2.7. Statistical Analysis
3. Results
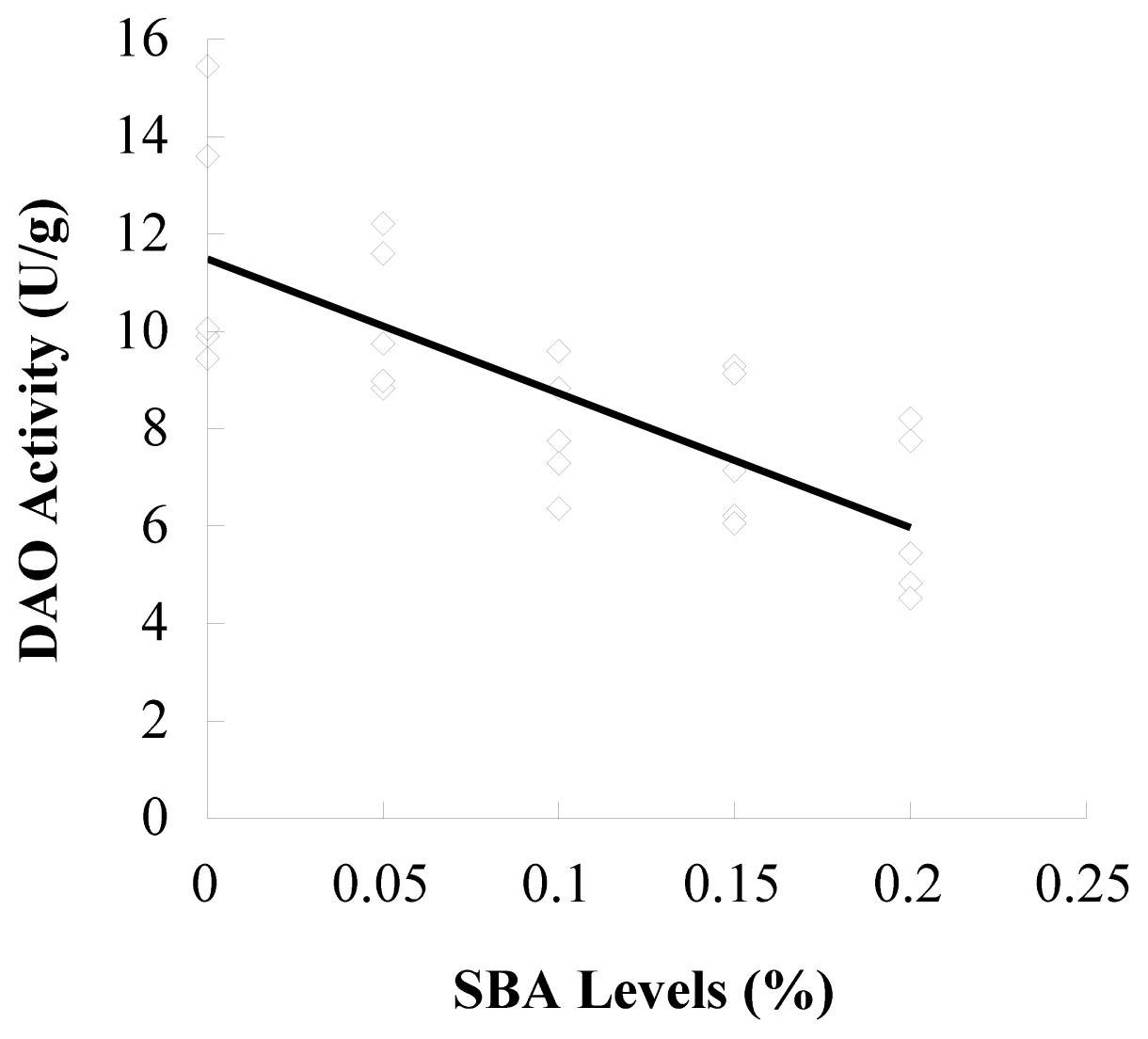
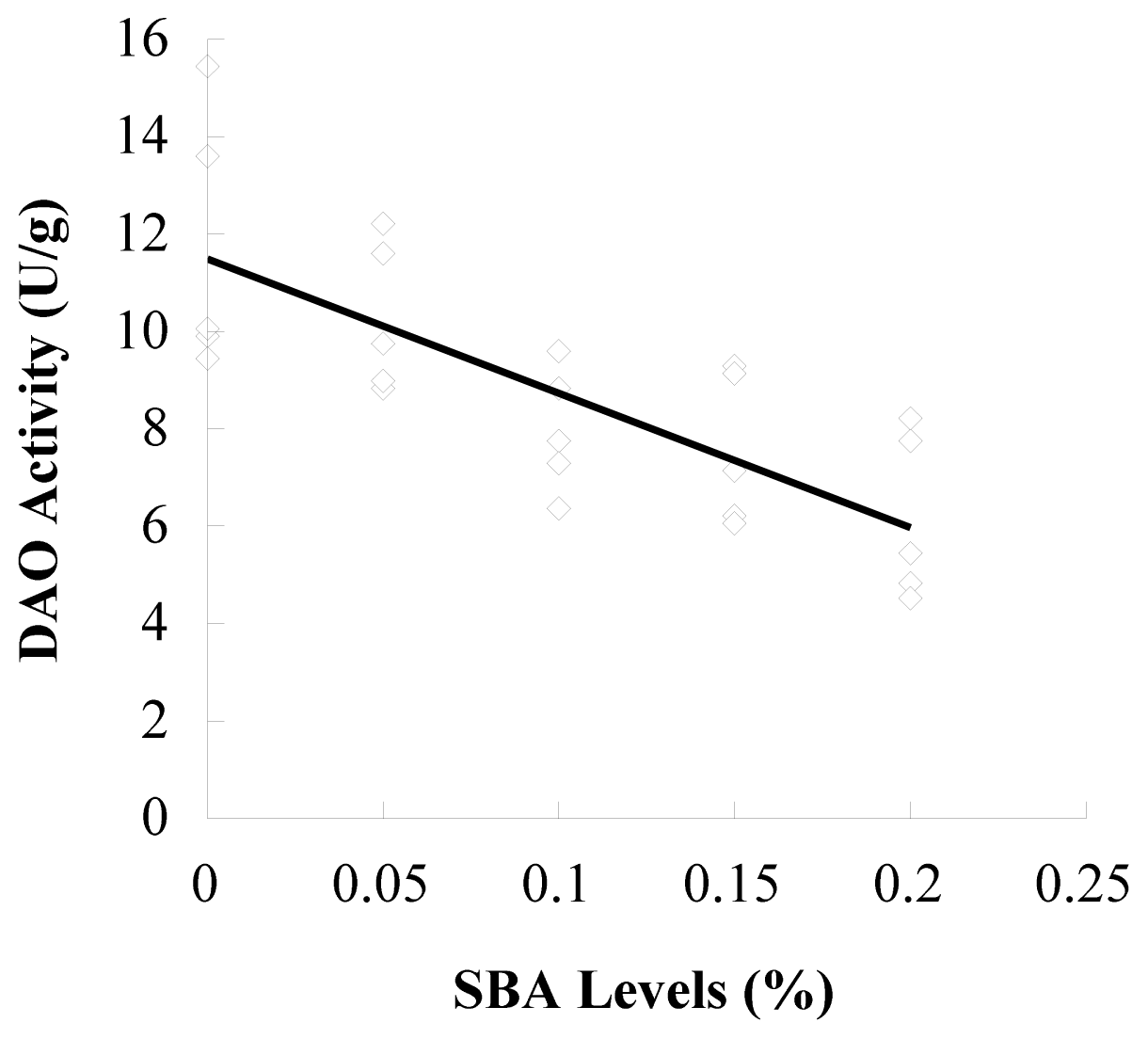
3.1. d-Lactic Acid in Plasma and DAO in Ileal Mucosa
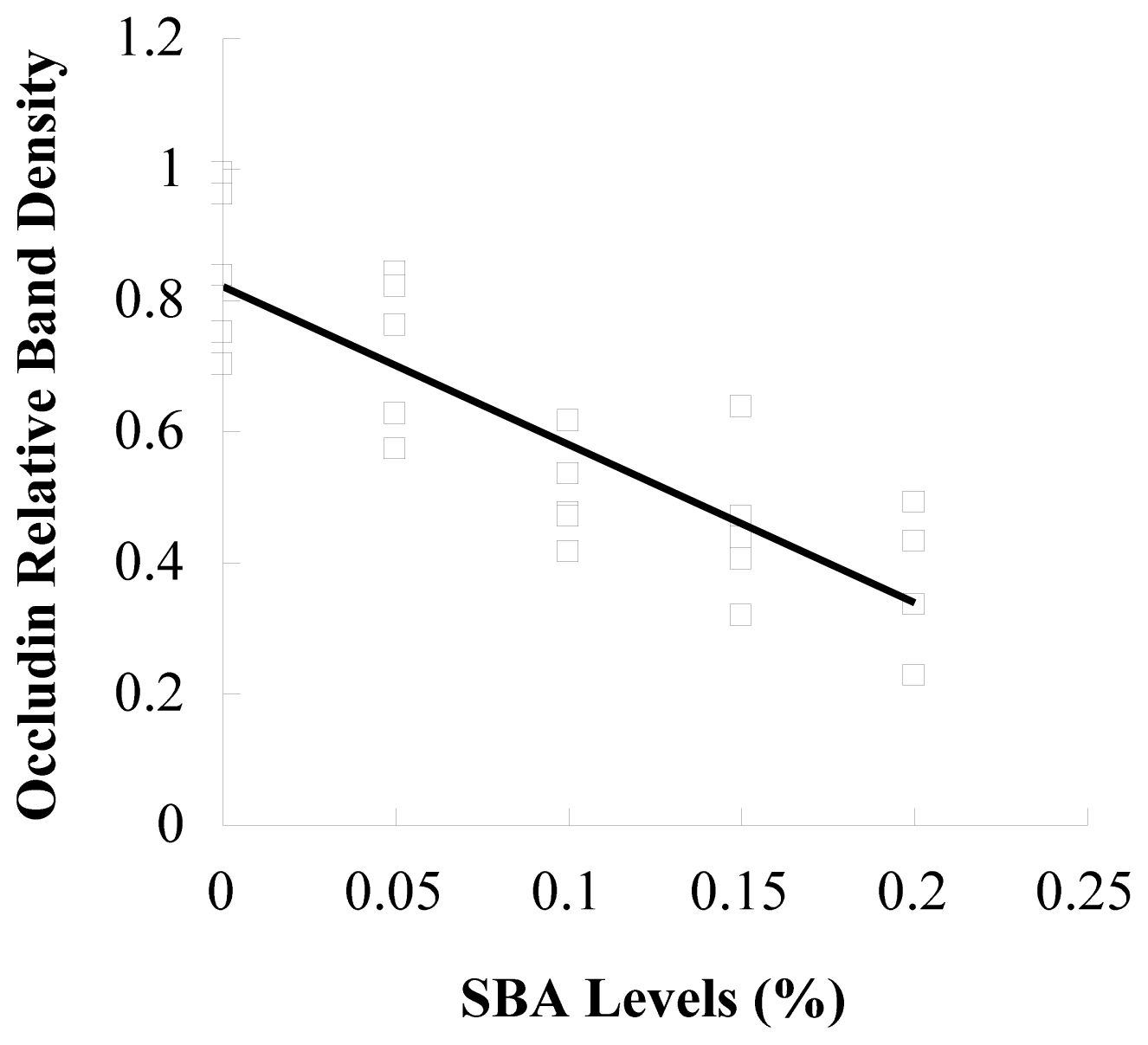
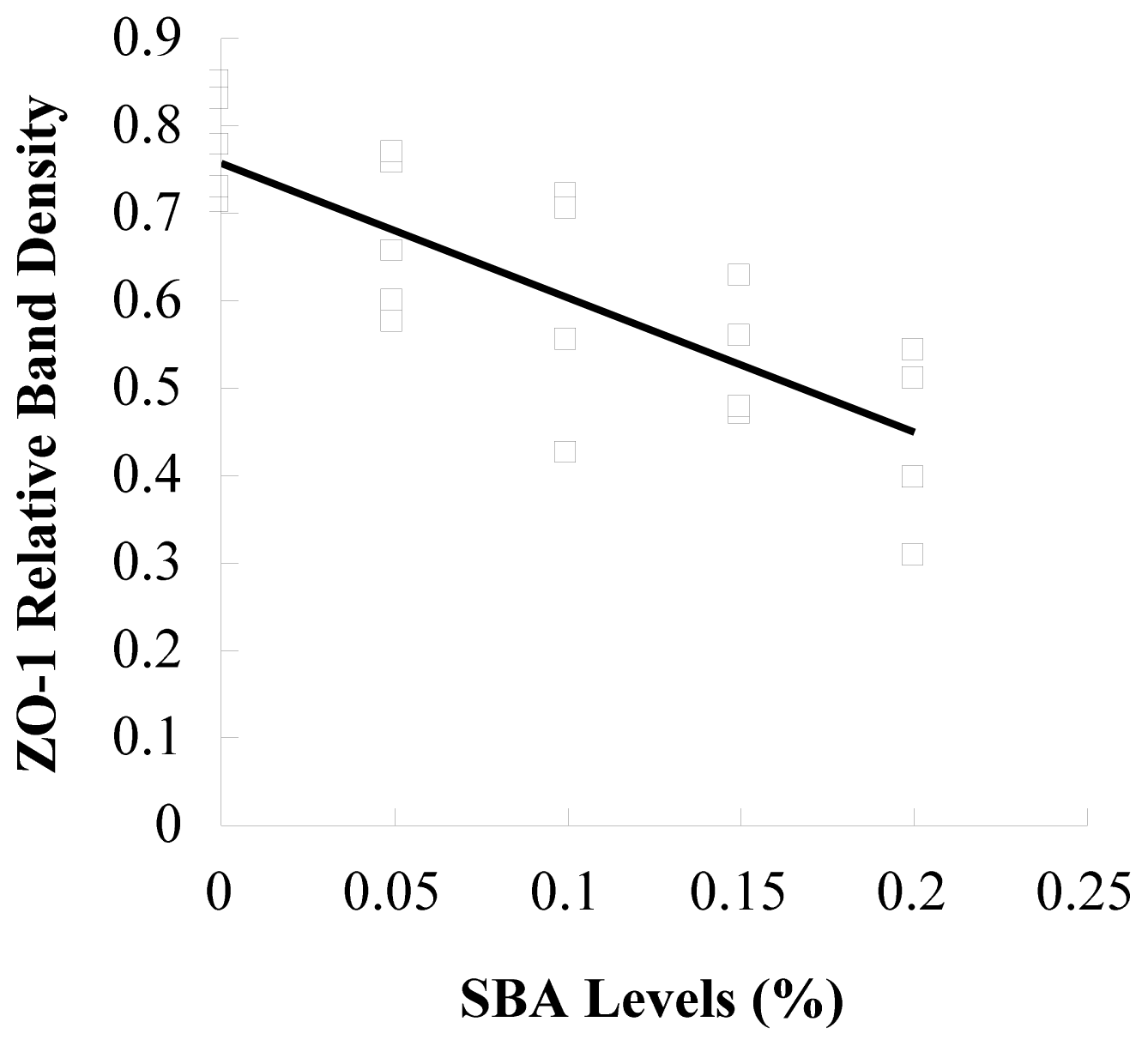
3.2. Occludin or ZO-1 Expression in Mid-Jejunum Tissue
4. Discussions
5. Conclusions
Acknowledgments
References
- Gatel, F. Protein quality of legume seeds for non-ruminant animals: A literature review. Anim. Feed Sci. Tech 1994, 45, 317–348. [Google Scholar]
- King, T.P.; Begbie, R.; Cadenhead, A. Nutritional toxicity of raw kidney beans in pigs. Immunocytochemical and cytopathological studies on the gutand the pancreas. J. Sci. Food Agric 1983, 34, 1404–1412. [Google Scholar]
- Sharon, N.; Lis, H. Legume lectins—A large family of homologous proteins. FASEB J 1990, 4, 3198–3208. [Google Scholar]
- Valdebouze, P.; Bergeron, E.; Gaborit, T.; Delort-Laval, J. Content and distribution of trypsin inhibitors and haemagglutinins in some legume seeds. Can. J. Plant Sci 1980, 60, 695–701. [Google Scholar]
- Gupta, Y.P. Anti-nutritional and toxic factors in food legumes: A review. Plant Foods Hum. Nutr 1987, 37, 201–228. [Google Scholar]
- Huisman, J.; Tolman, G.H. Antinutritional factors in the plant proteins of diets for non-ruminants. In Recent Developments in Pig Nutrition; Garnsworthy, P.C., Wiseman, J., Eds.; Nottingham University Press: Nottingham, UK, 2001; Volume 3, pp. 261–322. [Google Scholar]
- Friedman, M.; Brandon, D.L. Nutritional and health benefits of soy proteins. J. Agric. Food Chem 2001, 49, 1069–1077. [Google Scholar]
- Grant, G.; Watt, W.B.; Stewart, J.C.; Bardocz, S.; Pusztai, A. Intestinal and pancreatic responses to dietary soybean (Glycine-max) proteins. Biochem. Soc. Trans 1988, 16, 610–611. [Google Scholar]
- Zang, J.; Li, D.; Piao, X.; Tang, S. Effects of soybean agglutinin on body composition and organ weights in rats. Arch. Anim. Nutr 2006, 60, 245–253. [Google Scholar]
- Sharon, N.; Lis, H. Specificity and affinity. In Lectins, 2nd ed.; Sharon, N., Lis, H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 63–104. [Google Scholar]
- Yosbida, Y.; Ban, Y.; Kinosbita, S. Tight junction transmembrane protein claudin subtype expression and distribution in human corneal and conjunctival epithelium. Invest. Ophthalmol. Vis. Sci 2009, 50, 2103–2108. [Google Scholar]
- Liu, H.P.; Hu, C.H.; Xu, Y. Effects of early weaning on intestinal permeability and tight junction protein occludin mRDA expression levels of piglets. Chin. J. Anim. Nutr 2008, 20, 442–446. [Google Scholar]
- Hu, C.H.; Qian, Z.C.; Liu, H.P.; Xu, Y. Effect of high level of zinc oxide on tight junction expression in intestinal epithelial cells and intestinal mucosal barrier in early weaning piglets. Chin. J. Anim. Vet. Sci 2009, 40, 1638–1644. [Google Scholar]
- Li, J.Y.; Yu, Y.; Hao, Y.; Jin, H.; Xu, H. Determination of diamine oxidase activity in intestinal tissue and blood using spectrophotometry. Amino Acid Biot. Res 1996, 18, 28–30. [Google Scholar]
- Thompson, J.S. The effect of the route of nutrient delivery on gut structure and diaminoxidase levels. JPEN 1987, 11, 28–32. [Google Scholar]
- Bjanason, I.; Macpherson, A.; Hollander, D. Intestinal permeability: An overview. Gastroenterology 1995, 10, 1566–1581. [Google Scholar]
- Smith, S.M.; Eng, H.K.; Buccini, F. Use of d-lactic acid measurement in the diagnosis of bacterial infections. J. Infect. Dis 1985, 154, 658–664. [Google Scholar]
- Ohno, Y.; Naganuma, T.; Ogawa, T.; Muramoto, K. Effect of lectins on the transport of food factors in Caco-2 cell monolayers. J. Agric. Food Chem 2006, 54, 548–553. [Google Scholar]
- Cereijido, M. Evolution of ideas on the tight junction. In Tight Junctions; Cereijido, M., Ed.; CRC Press: Boca Raton, FL, USA, 1992; pp. 1–13. [Google Scholar]
- Furuse, M.; Hirase, T.; Itoh, M.; Nagafuchi, A.; Yonemura, S.; Tsukita, S. Occludin: A novel integral membrane protein localizing at tight junctions. J. Cell Biol 1993, 123, 1777–1788. [Google Scholar]
- Ando-Akatsuka, Y.; Saitou, M.; Hirase, T.; Kishi, M.; Sakakibara, A.; Itoh, M.; Yonemura, S.; Furuse, M.; Tsukita, S. Interspecies diversity of the occludin sequence: cDNA cloning of human, mouse, dog, and rat-kangaroo homologues. J. Cell Biol 1996, 133, 43–47. [Google Scholar]
- Saitou, M.; Furuse, M.; Sasaki, H.; Schulzke, J.D.; Fromm, M.; Takano, H.; Noda, T.; Tsukita, S. Complex phenotype of mice lacking occludin, a component of tight junction strands. Mol. Biol. Cell 2000, 11, 4131–4142. [Google Scholar]
- Schulzke, J.D.; Gitter, A.H.; Mankertz, J.; Spiegel, S.; Seidler, U.; Amasheh, S.; Saitou, M.; Tsukita, S.; Fromm, M. Epithelial transport and barrier function in occludin-deficient mice. Biochim. Biophys. Acta 2005, 1669, 34–42. [Google Scholar]
- Yu, A.S.; McCarthy, K.M.; Francis, S.A.; McCormack, J.M.; Lai, J.; Rogers, R.A.; Lynch, R.D.; Schneeberger, E.E. Knockdown of occludin expression leads to diverse phenotypic alterations in epithelial cells. Am. J. Physiol. Cell Physiol 2005, 288, C1231–C1241. [Google Scholar]
- Willott, E.; Balda, M.S.; Heintzelman, M.; Jameson, B.; Anderson, J.M. Localization and differential expression of two isoforms of the tight junction protein ZO-1. Am. J. Phys 1992, 262, C1119–C1124. [Google Scholar]
- Zhao, J.H.; Wang, J.H.; Dong, L.; Shi, H.; Wang, Z.; Ding, H.; Shi, H.; Lu, X. A protease inhibitor against acute stress-induced visceral hypersensitivity and paracellular permeability in rats. Eur. J. Pharmacol 2011, 654, 289–294. [Google Scholar]
- Pellegrina, C.D.; Rizzi, C.; Mosconi, S.; Zoccatelli, G.; Peruffo, A.; Chignola, R. Plant lectin as carriers for oral drugs: Is wheat germ agglutinin a suitable candidate? Toxicol. Appl. Pharmcol 2005, 207, 170–178. [Google Scholar]
- Pellegrina, C.D.; Perbellini, O.; Scupoli, M.T.; Tomelleri, C.; Zanetti, C.; Zoccatell, G.; Fusi, M.; Peruffo, A.; Rizzi, C.; Chignola, R. Effects of wheat germ agglutinin on human gastrointestinal epithelium: Insights from an experimental model of immunue/epithelial cell interaction. Toxicol. Appl. Pharmacol 2009, 237, 146–153. [Google Scholar]
- Pinton, P.; Nougayrède, J.P.; Rio, J.C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharmacol 2009, 237, 41–48. [Google Scholar] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Nutrient contents 3 (%) | ||
|---|---|---|---|
| Maize | 22.00 | Crude protein | 23.92 |
| Fish meal | 11.20 | Digestive energy (MJ/kg) | 16.91 |
| Whey powder | 18.00 | Calcium | 0.86 |
| Milk powder | 48.00 | Phosphorus | 0.83 |
| Salt | 0.20 | Lysine | 2.00 |
| Vitamin premix 1 | 0.10 | ||
| Vitamin premix 2 | 0.20 | ||
| Limestone | 0.30 |
| Groups | 0% SBA | 0.05% SBA | 0.1% SBA | 0.15% SBA | 0.2 % SBA |
|---|---|---|---|---|---|
| DAO/(U·g−1) | 11.69 ± 2.67 a | 10.28 ± 1.54 a,b | 7.97 ± 1.27 bc | 7.57 ± 1.56 c | 6.15 ± 1.71 c |
| d-lactic acid/(μg·mL−1) | 2.11 ± 0.26 a | 2.09 ± 0.29 a | 2.29 ± 0.12 ab | 2.45 ± 0.09 b,c | 2.62 ± 0.18 c |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, Y.; Qin, G.; Sun, Z.; Che, D.; Bao, N.; Zhang, X. Effects of Soybean Agglutinin on Intestinal Barrier Permeability and Tight Junction Protein Expression in Weaned Piglets. Int. J. Mol. Sci. 2011, 12, 8502-8512. https://doi.org/10.3390/ijms12128502
Zhao Y, Qin G, Sun Z, Che D, Bao N, Zhang X. Effects of Soybean Agglutinin on Intestinal Barrier Permeability and Tight Junction Protein Expression in Weaned Piglets. International Journal of Molecular Sciences. 2011; 12(12):8502-8512. https://doi.org/10.3390/ijms12128502
Chicago/Turabian StyleZhao, Yuan, Guixin Qin, Zewei Sun, Dongsheng Che, Nan Bao, and Xiaodong Zhang. 2011. "Effects of Soybean Agglutinin on Intestinal Barrier Permeability and Tight Junction Protein Expression in Weaned Piglets" International Journal of Molecular Sciences 12, no. 12: 8502-8512. https://doi.org/10.3390/ijms12128502
APA StyleZhao, Y., Qin, G., Sun, Z., Che, D., Bao, N., & Zhang, X. (2011). Effects of Soybean Agglutinin on Intestinal Barrier Permeability and Tight Junction Protein Expression in Weaned Piglets. International Journal of Molecular Sciences, 12(12), 8502-8512. https://doi.org/10.3390/ijms12128502