Phospholipases A1


Abstract
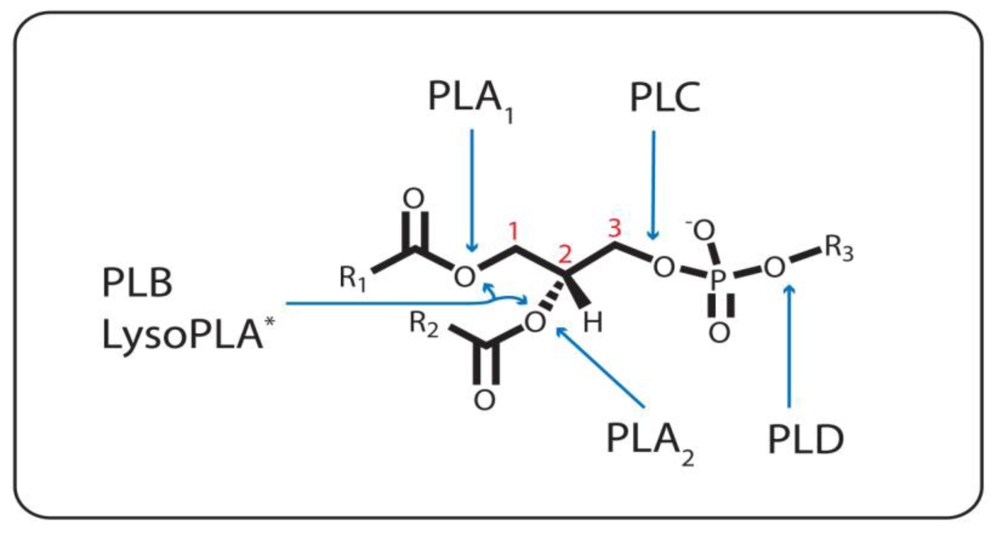
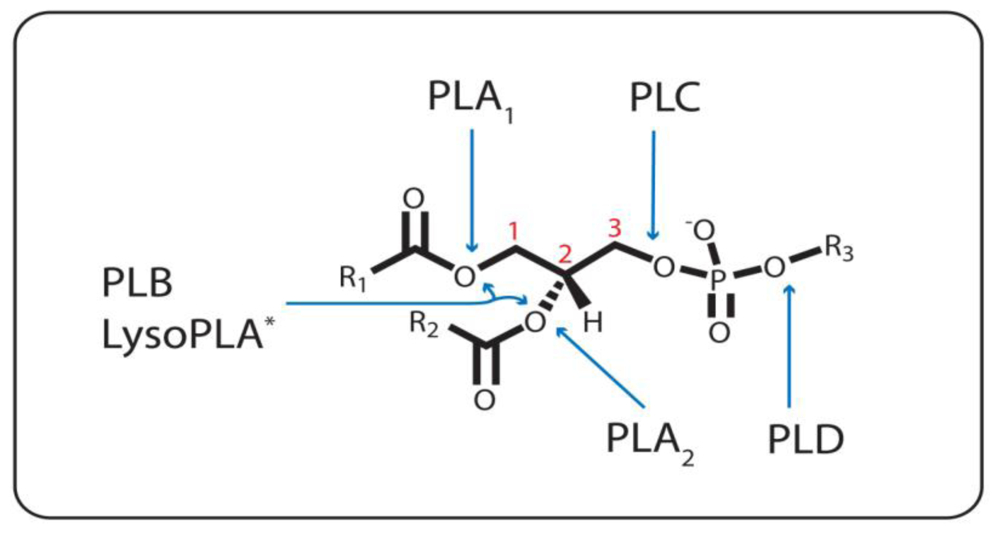
:1. Introduction
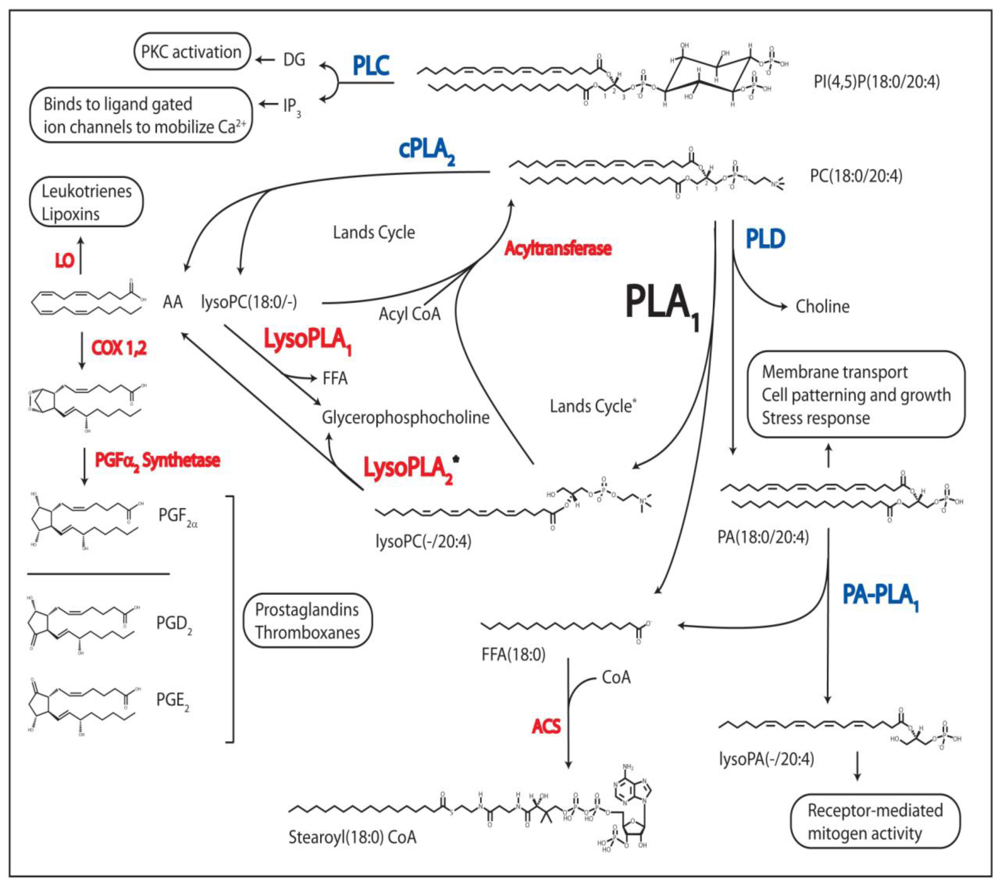
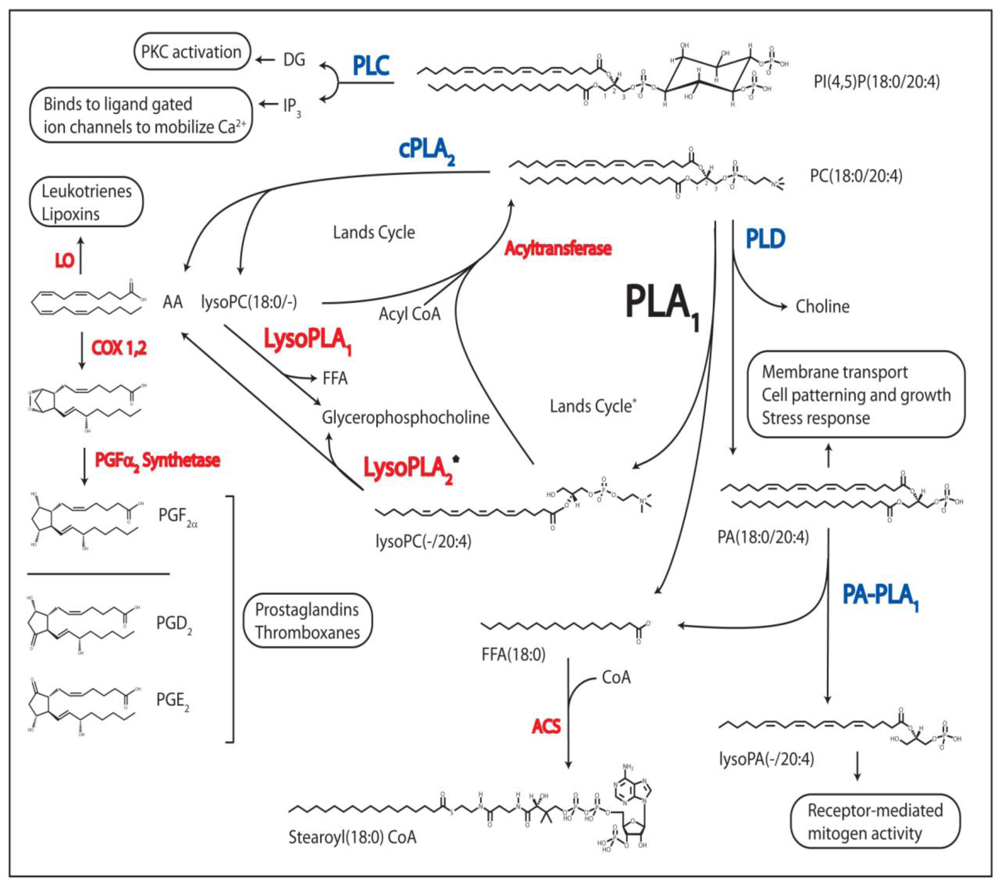
1.1. Phospholipase A1
1.2. The Other Acyl Hydrolases: Phospholipase A2, B, and Lysophospholipase
1.3. The Phosphodiesterases: Phospholipase C and D
2. Classification of Phospholipase A1
3. Mammalian Phospholipase A1
3.1. Bovine Brain PLA1
3.2. Bovine Testis PA-PLA
3.3. p125 and KIAA0725p
3.4. PS-PLA1
3.5. mPA-PLA1
3.6. Guinea-Pig Heart Microsomal PLA1
4. Other PLA1
4.1. Caenorhabditis elegans PLA1
4.2. Venom PLA1
4.3. Trypanosoma brucei PLA1
4.4. Trypanosoma cruzi PLA1
5. Plant Phospholipase A1
5.1. DAD1
5.2. AtLCAT3
6. Bacterial PLA1
7. PLA1 in Biotechnology
8. Perspective
Acknowledgements
References
- Lands, WE. Stories about acyl chains. Biochim. Biophys Acta 2000, 1483, 1–14. [Google Scholar]
- Nishijima, M; Nakaike, S; Tamori, Y; Nojima, S. Detergent-resistant phospholipase A of Escherichia coli K-12, purification and properties. Eur. J. Biochem 1977, 73, 115–124. [Google Scholar]
- Withers-Martinez, C; Carriere, F; Verger, R; Bourgeois, D; Cambillau, C. A pancreatic lipase with a phospholipase A1 activity: crystal structure of a chimeric pancreatic lipase-related protein 2 from guinea pig. Structure 1996, 4, 1363–1374. [Google Scholar]
- Lands, WE. Metabolism of glycerolipids. 2. The enzymatic acylation of lysolecithin. J. Biol. Chem 1960, 235, 2233–2237. [Google Scholar]
- Imae, R; Inoue, T; Kimura, M; Kanamori, T; Tomioka, NH; Kage-Nakadai, E; Mitani, S; Arai, H. Intracellular phospholipase A1 and acyltransferase, which are involved in Caenorhabditis elegans stem cell divisions, determine the sn-1 fatty acyl chain of phosphatidylinositol. Mol Biol Cell 2010, 21, 3114–3124, and reference therein. [Google Scholar]
- Zarini, S; Gijon, MA; Folco, G; Murphy, RC. Effect of arachidonic acid reacylation on leukotriene biosynthesis in human neutrophils stimulated with granulocyte-macrophage colony-stimulating factor and formyl-methionyl-leucyl-phenylalanine. J. Biol Chem 2006, 281, 10134–10142. [Google Scholar]
- Reinhold, SL; Zimmerman, GA; Prescott, SM; McIntyre, TM. Phospholipid remodeling in human neutrophils. Parallel activation of a deacylation/reacylation cycle and platelet-activating factor synthesis. J. Biol. Chem 1989, 264, 21652–21659. [Google Scholar]
- Yamashita, A; Sugiura, T; Waku, K. Acyltransferases and transacylases involved in fatty acid remodeling of phospholipids and metabolism of bioactive lipids in mammalian cells. J. Biochem (Tokyo) 1997, 122, 1–16. [Google Scholar]
- Blank, ML; Smith, ZL; Snyder, F. Contributing factors in the trafficking of [3H]arachidonate between phospholipids. Biochim. Biophys Acta 1992, 1124, 262–272. [Google Scholar]
- Perez, R; Matabosch, X; Llebaria, A; Balboa, MA; Balsinde, J. Blockade of arachidonic acid incorporation into phospholipids induces apoptosis in U937 promonocytic cells. J. Lipid. Res 2006, 47, 484–491. [Google Scholar]
- Waite, M. The Phospholipases (Handbook of Lipid Research); Springer: New York, NY, USA, 1987. [Google Scholar]
- Shimuta, K; Ohnishi, M; Iyoda, S; Gotoh, N; Koizumi, N; Watanabe, H. The hemolytic and cytolytic activities of Serratia marcescens phospholipase A (PhlA) depend on lysophospholipid production by PhlA. BMC Microbiol 2009, 9, 261. [Google Scholar]
- Fahy, E; Subramaniam, S; Brown, HA; Glass, CK; Merrill, AH, Jr; Murphy, RC; Raetz, CR; Russell, DW; Seyama, Y; Shaw, W; et al. A comprehensive classification system for lipids. J. Lipid. Res 2005, 46, 839–862. [Google Scholar]
- Hla, T; Lee, MJ; Ancellin, N; Paik, JH; Kluk, MJ. Lysophospholipids—receptor revelations. Science 2001, 294, 1875–1878. [Google Scholar]
- Ishii, I; Fukushima, N; Ye, X; Chun, J. Lysophospholipid receptors: signaling and biology. Annu. Rev. Biochem 2004, 73, 321–354. [Google Scholar]
- Gauster, M; Rechberger, G; Sovic, A; Horl, G; Steyrer, E; Sattler, W; Frank, S. Endothelial lipase releases saturated and unsaturated fatty acids of high density lipoprotein phosphatidylcholine. J. Lipid. Res 2005, 46, 1517–1525. [Google Scholar]
- Wang, A; Loo, R; Chen, Z; Dennis, EA. Regiospecificity and catalytic triad of lysophospholipase I. J. Biol. Chem 1997, 272, 22030–22036. [Google Scholar]
- Arthur, G. Lysophospholipase A2 activity in guinea-pig heart microsomal fractions displaying high activities with 2-acylglycerophosphocholines with linoleic and arachidonic acids. Biochem. J 1989, 261, 581–586. [Google Scholar]
- Badiani, K; Arthur, G. 2-acyl-sn-glycero-3-phosphoethanolamine lysophospholipase A2 activity in guinea-pig heart microsomes. Biochem. J 1991, 275, 393–398. [Google Scholar]
- Burgoyne, RD; Morgan, A. The control of free arachidonic acid levels. Trends Biochem. Sci 1990, 15, 365–366. [Google Scholar]
- Pete, MJ; Ross, AH; Exton, JH. Purification and properties of phospholipase A1 from bovine brain. J. Biol. Chem 1994, 269, 19494–19500. [Google Scholar]
- Pete, MJ; Wu, DW; Exton, JH. Subcellular fractions of bovine brain degrade phosphatidylcholine by sequential deacylation of the sn-1 and sn-2 positions. Biochim. Biophys Acta 1996, 1299, 325–332. [Google Scholar]
- Ridgley, EL; Ruben, L. Phospholipase from Trypanosoma brucei releases arachidonic acid by sequential sn-1, sn-2 deacylation of phospholipids. Mol. Biochem. Parasitol 2001, 114, 29–40. [Google Scholar]
- van den Bosch, H. Intracellular phospholipases A. Biochim. Biophys Acta 1980, 604, 191–246. [Google Scholar]
- Six, DA; Dennis, EA. The expanding superfamily of phospholipase A2 enzymes: classification and characterization. Biochim. Biophys Acta 2000, 1488, 1–19. [Google Scholar]
- Balsinde, J; Balboa, MA; Dennis, EA. Antisense inhibition of group VI Ca2+-independent phospholipase A2 blocks phospholipid fatty acid remodeling in murine P388D1 macrophages. J. Biol. Chem 1997, 272, 29317–29321. [Google Scholar]
- Tjoelker, LW; Eberhardt, C; Unger, J; Trong, HL; Zimmerman, GA; McIntyre, TM; Stafforini, DM; Prescott, SM; Gray, PW. Plasma platelet-activating factor acetylhydrolase is a secreted phospholipase A2 with a catalytic triad. J. Biol. Chem 1995, 270, 25481–25487. [Google Scholar]
- Gilroy, DW; Newson, J; Sawmynaden, P; Willoughby, DA; Croxtall, JD. A novel role for phospholipase A2 isoforms in the checkpoint control of acute inflammation. FASEB J 2004, 18, 489–498. [Google Scholar]
- Chilton, FH; O'Flaherty, JT; Ellis, JM; Swendsen, CL; Wykle, RL. Selective acylation of lyso platelet activating factor by arachidonate in human neutrophils. J. Biol. Chem 1983, 258, 7268–7271. [Google Scholar]
- Bogatcheva, NV; Sergeeva, MG; Dudek, SM; Verin, AD. Arachidonic acid cascade in endothelial pathobiology. Microvasc. Res 2005, 69, 107–127. [Google Scholar]
- Scott, DL; White, SP; Otwinowski, Z; Yuan, W; Gelb, MH; Sigler, PB. Interfacial catalysis: The mechanism of phospholipase A2. Science 1990, 250, 1541–1546. [Google Scholar]
- Masuda, N; Kitamura, N; Saito, K. Primary structure of protein moiety of Penicillium notatum phospholipase B deduced from the cDNA. Eur. J. Biochem 1991, 202, 783–787. [Google Scholar]
- Lee, KS; Patton, JL; Fido, M; Hines, LK; Kohlwein, SD; Paltauf, F; Henry, SA; Levin, DE. The Saccharomyces cerevisiae PLB1 gene encodes a protein required for lysophospholipase and phospholipase B activity. J. Biol. Chem 1994, 269, 19725–19730. [Google Scholar]
- Cao, YZ; Tam, SW; Arthur, G; Chen, H; Choy, PC. The purification and characterization of a phospholipase A in hamster heart cytosol for the hydrolysis of phosphatidylcholine. J Biol Chem 1987, 262, 16927–16936. [Google Scholar]
- Dekker, N. Outer-membrane phospholipase A: known structure, unknown biological function. Mol. Microbiol 2000, 35, 711–717. [Google Scholar]
- Homma, H; Kobayashi, T; Chiba, N; Karasawa, K; Mizushima, H; Kudo, I; Inoue, K; Ikeda, H; Sekiguchi, M; Nojima, S. The DNA sequence encoding pldA gene, the structural gene for detergent-resistant phospholipase A of E. coli. J. Biochem (Tokyo) 1984, 96, 1655–1664. [Google Scholar]
- Scandella, CJ; Kornberg, A. A membrane-bound phospholipase A1 purified from Escherichia coli. Biochemistry 1971, 10, 4447–4456. [Google Scholar]
- Wang, A; Dennis, EA. Mammalian lysophospholipases. Biochim. Biophys Acta 1999, 1439, 1–16. [Google Scholar]
- Pete, MJ; Exton, JH. Purification of a lysophospholipase from bovine brain that selectively deacylates arachidonoyl-substituted lysophosphatidylcholine. J. Biol. Chem 1996, 271, 18114–18121. [Google Scholar]
- James, SR; Downes, CP. Structural and mechanistic features of phospholipases C: Effectors of inositol phospholipid-mediated signal transduction. Cell. Signal 1997, 9, 329–336. [Google Scholar]
- Katan, M. Families of phosphoinositide-specific phospholipase C: structure and function. Biochim. Biophys Acta 1998, 1436, 5–17. [Google Scholar]
- McDermott, M; Wakelam, MJ; Morris, AJ. Phospholipase D. Biochem. Cell Biol 2004, 82, 225–253. [Google Scholar]
- Sonoda, H; Aoki, J; Hiramatsu, T; Ishida, M; Bandoh, K; Nagai, Y; Taguchi, R; Inoue, K; Arai, H. A novel phosphatidic acid-selective phospholipase A1 that produces lysophosphatidic acid. J. Biol. Chem 2002, 277, 34254–34263. [Google Scholar]
- de Hass, GH; Daemen, FJM; van Deenen, DL. Positional specificity of phosphatide acyl hydrolase (phospholipase A). Nature 1962, 196, 68. [Google Scholar]
- van den Bosch, H; van Deenen, LL. Chemical structure and biochemical significance of lysolecithins from rat liver. Biochim. Biophys Acta 1965, 106, 326–337. [Google Scholar]
- Sato, T; Aoki, J; Nagai, Y; Dohmae, N; Takio, K; Doi, T; Arai, H; Inoue, K. Serine phospholipid-specific phospholipase A that is secreted from activated platelets. A new member of the lipase family. J. Biol. Chem 1997, 272, 2192–2198. [Google Scholar]
- Soldatova, L; Kochoumian, L; King, TP. Sequence similarity of a hornet (D. maculata) venom allergen phospholipase A1 with mammalian lipases. FEBS Lett 1993, 320, 145–149. [Google Scholar]
- Aoki, J; Nagai, Y; Hosono, H; Inoue, K; Arai, H. Structure and function of phosphatidylserine-specific phospholipase A1. Biochim. Biophys Acta 2002, 1582, 26–32. [Google Scholar]
- Richmond, GS; Smith, TK. A novel phospholipase from Trypanosoma brucei. Mol. Microbiol 2007, 63, 1078–1095. [Google Scholar]
- Aoki, J; Inoue, A; Makide, K; Saiki, N; Arai, H. Structure and function of extracellular phospholipase A1 belonging to the pancreatic lipase gene family. Biochimie 2007, 89, 197–204. [Google Scholar]
- Jaeger, KE; Ransac, S; Dijkstra, BW; Colson, C; van Heuvel, M; Misset, O. Bacterial lipases. FEMS Microbiol. Rev 1994, 15, 29–63. [Google Scholar]
- Higgs, HN; Han, MH; Johnson, GE; Glomset, JA. Cloning of a phosphatidic acid-preferring phospholipase A1 from bovine testis. J. Biol. Chem 1998, 273, 5468–5477. [Google Scholar]
- Higgs, HN; Glomset, JA. Identification of a phosphatidic acid-preferring phospholipase A1 from bovine brain and testis. Proc. Natl. Acad. Sci USA 1994, 91, 9574–9578. [Google Scholar]
- Tani, K; Mizoguchi, T; Iwamatsu, A; Hatsuzawa, K; Tagaya, M. p125 is a novel mammalian Sec23p-interacting protein with structural similarity to phospholipid-modifying proteins. J. Biol. Chem 1999, 274, 20505–20512. [Google Scholar]
- Nakajima, K; Sonoda, H; Mizoguchi, T; Aoki, J; Arai, H; Nagahama, M; Tagaya, M; Tani, K. A novel phospholipase A1 with sequence homology to a mammalian Sec23p-interacting protein, p125. J. Biol. Chem 2002, 277, 11329–11335. [Google Scholar]
- Hiramatsu, T; Sonoda, H; Takanezawa, Y; Morikawa, R; Ishida, M; Kasahara, K; Sanai, Y; Taguchi, R; Aoki, J; Arai, H. Biochemical and molecular characterization of two phosphatidic acid-selective phospholipase A1s, mPA-PLA1 alpha and mPA-PLA1 beta. J. Biol. Chem 2003, 278, 49438–49447. [Google Scholar]
- Hosono, H; Homma, M; Ogasawara, Y; Makide, K; Aoki, J; Niwata, H; Watanabe, M; Inoue, K; Ohkohchi, N; Kohda, Y. Expression of Phosphatidylserine-Specific Phospholipase A1 mRNA in Human THP-1-Derived Macrophages. Cell Transplant 2010, 19, 759–764. [Google Scholar]
- Hoffman, DR. Allergens in hymenoptera venom. XXVI: The complete amino acid sequences of two vespid venom phospholipases. Int. Arch. Allergy Immunol 1994, 104, 184–190. [Google Scholar]
- King, TP; Lu, G; Gonzalez, M; Qian, N; Soldatova, L. Yellow jacket venom allergens, hyaluronidase and phospholipase: sequence similarity and antigenic cross-reactivity with their hornet and wasp homologs and possible implications for clinical allergy. J. Allergy Clin. Immunol 1996, 98, 588–600. [Google Scholar]
- Seismann, H; Blank, S; Cifuentes, L; Braren, I; Bredehorst, R; Grunwald, T; Ollert, M; Spillner, E. Recombinant phospholipase A1 (Ves v 1) from yellow jacket venom for improved diagnosis of hymenoptera venom hypersensitivity. Clin. Mol Allergy 2010, 8, 7. [Google Scholar]
- Ishiguro, S; Kawai-Oda, A; Ueda, J; Nishida, I; Okada, K. The DEFECTIVE IN ANTHER DEHISCIENCE1 gene encodes a novel phospholipase A1 catalyzing the initial step of jasmonic acid biosynthesis, which synchronizes pollen maturation, anther dehiscence, and flower opening in Arabidopsis. Plant Cell 2001, 13, 2191–2209. [Google Scholar]
- Noiriel, A; Benveniste, P; Banas, A; Stymne, S; Bouvier-Nave, P. Expression in yeast of a novel phospholipase A1 cDNA from Arabidopsis thaliana. Eur. J. Biochem 2004, 271, 3752–3764. [Google Scholar]
- Kato, T; Morita, MT; Fukaki, H; Yamauchi, Y; Uehara, M; Niihama, M; Tasaka, M. SGR2, a phospholipase-like protein, and ZIG/SGR4, a SNARE, are involved in the shoot gravitropism of Arabidopsis. Plant Cell 2002, 14, 33–46. [Google Scholar]
- Watanabe, I; Koishi, R; Yao, Y; Tsuji, T; Serizawa, N. Molecular cloning and expression of the gene encoding a phospholipase A1 from Aspergillus oryzae. Biosci. Biotechnol. Biochem 1999, 63, 820–826. [Google Scholar]
- Kim, I-H; Garcia, HS; Hill, CG, Jr. Phospholipase A1-catalyzed synthesis of phospholipids enriched in n-3 polyunsaturated fatty acid residues. Enzyme Microb. Technol 2007, 40, 1130–1135. [Google Scholar]
- Richmond, G; Smith, TK. The role and characterization of phospholipase A1 in mediating lysophosphatidylcholine synthesis in Trypanosoma brucei. Biochem. J 2007, 405, 319–329. [Google Scholar]
- Givskov, M; Molin, S. Secretion of Serratia liquefaciens phospholipase from Escherichia coli. Mol. Microbiol 1993, 8, 229–242. [Google Scholar]
- Givskov, M; Olsen, L; Molin, S. Cloning and expression in Escherichia coli of the gene for extracellular phospholipase A1 from Serratia liquefaciens. J. Bacteriol 1988, 170, 5855–5862. [Google Scholar]
- Song, JK; Kim, MK; Rhee, JS. Cloning and expression of the gene encoding phospholipase A1 from Serratia sp. MK1 in Escherichia coli. J. Biotechnol 1999, 72, 103–114. [Google Scholar]
- Schmiel, DH; Wagar, E; Karamanou, L; Weeks, D; Miller, VL. Phospholipase A of Yersinia enterocolitica contributes to pathogenesis in a mouse model. Infect. Immun 1998, 66, 3941–3951. [Google Scholar]
- Kucera, GL; Sisson, PJ; Thomas, MJ; Waite, M. On the substrate specificity of rat liver phospholipase A1. J. Biol. Chem 1988, 263, 1920–1928. [Google Scholar]
- Hirata, K; Dichek, HL; Cioffi, JA; Choi, SY; Leeper, NJ; Quintana, L; Kronmal, GS; Cooper, AD; Quertermous, T. Cloning of a unique lipase from endothelial cells extends the lipase gene family. J. Biol. Chem 1999, 274, 14170–14175. [Google Scholar]
- Pete, MJ; Exton, JH. Phospholipid interactions affect substrate hydrolysis by bovine brain phospholipase A1. Biochim. Biophys Acta 1995, 1256, 367–373. [Google Scholar]
- Wang, X; Devaiah, SP; Zhang, W; Welti, R. Signaling functions of phosphatidic acid. Prog Lipid Res 2006, 45, 250. [Google Scholar]
- Higgs, HN; Glomset, JA. Purification and properties of a phosphatidic acid-preferring phospholipase A1 from bovine testis. Examination of the molecular basis of its activation. J. Biol. Chem 1996, 271, 10874–10882. [Google Scholar]
- Lin, Q; Higgs, HN; Glomset, JA. Membrane lipids have multiple effects on interfacial catalysis by a phosphatidic acid-preferring phospholipase A1 from bovine testis. Biochemistry 2000, 39, 9335–9344. [Google Scholar]
- Han, MH; Han, DK; Aebersold, RH; Glomset, JA. Effects of protein kinase CK2, extracellular signal-regulated kinase 2, and protein phosphatase 2A on a phosphatidic acid preferring phospholipase A1. J. Biol. Chem 2001, 276, 27698–27708. [Google Scholar]
- Mizoguchi, T; Nakajima, K; Hatsuzawa, K; Nagahama, M; Hauri, HP; Tagaya, M; Tani, K. Determination of functional regions of p125, a novel mammalian Sec23p-interacting protein. Biochem. Biophys. Res. Commun 2000, 279, 144–149. [Google Scholar]
- Nakamura, N; Lowe, M; Levine, TP; Rabouille, C; Warren, G. The vesicle docking protein p115 binds GM130, a cis-Golgi matrix protein, in a mitotically regulated manner. Cell 1997, 89, 445–455. [Google Scholar]
- Shimoi, W; Ezawa, I; Nakamoto, K; Uesaki, S; Gabreski, G; Aridor, M; Yamamoto, A; Nagahama, M; Tagaya, M; Tani, K. p125 is localized in endoplasmic reticulum exit sites and involved in their organization. J. Biol. Chem 2005, 280, 10141–10148. [Google Scholar]
- Chernomordik, L. Non-bilayer lipids and biological fusion intermediates. Chem. Phys Lipids 1996, 81, 203–213. [Google Scholar]
- Morikawa, RK; Aoki, J; Kano, F; Murata, M; Yamamoto, A; Tsujimoto, M; Arai, H. Intracellular phospholipase A1 gamma (iPLA1 gamma) is a novel factor involved in coat protein complex I- and Rab6-independent retrograde transport between the endoplasmic reticulum and the Golgi complex. J. Biol. Chem 2009, 284, 26620–26630. [Google Scholar]
- Hosono, H; Aoki, J; Nagai, Y; Bandoh, K; Ishida, M; Taguchi, R; Arai, H; Inoue, K. Phosphatidylserine-specific phospholipase A1 stimulates histamine release from rat peritoneal mast cells through production of 2-acyl-1-lysophosphatidylserine. J. Biol. Chem 2001, 276, 29664–29670. [Google Scholar]
- Nagai, Y; Aoki, J; Sato, T; Amano, K; Matsuda, Y; Arai, H; Inoue, K. An alternative splicing form of phosphatidylserine-specific phospholipase A1 that exhibits lysophosphatidylserine-specific lysophospholipase activity in humans. J. Biol. Chem 1999, 274, 11053–11059. [Google Scholar]
- Badiani, K; Arthur, G. Evidence for receptor and G-protein regulation of a phosphatidylethanolamine-hydrolysing phospholipase A1 in guinea-pig heart microsomes: stimulation of phospholipase A1 activity by DL-isoprenaline and guanine nucleotides. Biochem. J 1995, 312, 805–809. [Google Scholar]
- Badiani, K; Lu, X; Arthur, G. Evidence for the regulation of guinea-pig heart microsomal phosphatidylcholine-hydrolysing phospholipase A1 by guanosine 5′-[gamma-thio]triphosphate. Biochem. J 1992, 288, 965–968. [Google Scholar]
- Yoshihisa, T; Barlowe, C; Schekman, R. Requirement for a GTPase-activating protein in vesicle budding from the endoplasmic reticulum. Science 1993, 259, 1466–1468. [Google Scholar]
- Inoue, A; Aoki, J. Phospholipase A1, struture, distrubition and function. Future Lipidol 2006, 1, 623–636. [Google Scholar]
- Tizard, IR; Mellors, A; Holmes, WL; Nielsen, K. The generation of phospholipase A and hemolytic fatty acids by autolysing suspensions of Trypanosoma congolense. Tropenmed. Parasitol 1978, 29, 127–133. [Google Scholar]
- Hambrey, PN; Mellors, A; Tizard, IR. The phospholipases of pathogenic and non-pathogenic Trypanosoma species. Mol. Biochem. Parasitol 1981, 2, 177–186. [Google Scholar]
- Opperdoes, FR; van Roy, J. The phospholipases of Trypanosoma brucei bloodstream forms and cultured procyclics. Mol. Biochem. Parasitol 1982, 5, 309–319. [Google Scholar]
- Eintracht, J; Maathai, R; Mellors, A; Ruben, L. Calcium entry in Trypanosoma brucei is regulated by phospholipase A2 and arachidonic acid. Biochem. J 1998, 336, 659–666. [Google Scholar]
- Catisti, R; Uyemura, SA; Docampo, R; Vercesi, AE. Calcium mobilization by arachidonic acid in trypanosomatids. Mol. Biochem. Parasitol 2000, 105, 261–271. [Google Scholar]
- Kubata, BK; Duszenko, M; Kabututu, Z; Rawer, M; Szallies, A; Fujimori, K; Inui, T; Nozaki, T; Yamashita, K; Horii, T; Urade, Y; Hayaishi, O. Identification of a novel prostaglandin f(2alpha) synthase in Trypanosoma brucei. J. Exp. Med 2000, 192, 1327–1338. [Google Scholar]
- Wainszelbaum, M; Isola, E; Wilkowsky, S; Cannata, JJ; Florin-Christensen, J; Florin-Christensen, M. Lysosomal phospholipase A1 in Trypanosoma cruzi: An enzyme with a possible role in the pathogenesis of Chagas’ disease. Biochem. J 2001, 355, 765–770. [Google Scholar]
- Tavernier, E; Pugin, A. Phospholipase activities associated with the tonoplast from Acer pseudoplatanus cells: Identification of a phospholipase A1 activity. Biochim. Biophys Acta 1995, 1233, 118–122. [Google Scholar]
- Browse, J. Jasmonate: An oxylipin signal with many roles in plants. Vitam. Horm 2005, 72, 431–456. [Google Scholar]
- Seo, YS; Kim, EY; Mang, HG; Kim, WT. Heterologous expression, and biochemical and cellular characterization of CaPLA1 encoding a hot pepper phospholipase A1 homolog. Plant J 2008, 53, 895–908. [Google Scholar]
- Givskov, M; Molin, S. Expression of extracellular phospholipase from Serratia liquefaciens is growth-phase-dependent, catabolite-repressed and regulated by anaerobiosis. Mol. Microbiol 1992, 6, 1363–1374. [Google Scholar]
- Givskov, M; Eberl, L; Christiansen, G; Benedik, MJ; Molin, S. Induction of phospholipase-and flagellar synthesis in Serratia liquefaciens is controlled by expression of the flagellar master operon flhD. Mol. Microbiol 1995, 15, 445–454. [Google Scholar]
- Miyahara, R; Uehara, K. Recent trends in cosmetics and toiletries. Inform 1995, 6, 672. [Google Scholar]
- Barbara, FH. Lipsomes offer hope as medical tools. Inform 1995, 6, 793. [Google Scholar]
- D’Arrigo, P; Stefano Servi, S. Synthesis of Lysophospholipids. Molecules 2010, 15, 1354–1377. [Google Scholar]
- Guberman, A; Hartmann, M; Tiedtke, A; Florin-Christensen, J; Florin-Christensen, M. A method for the preparation of Tetrahymena thermophila phospholipase A1 suitable for large-scale production. J. Appl. Microbiol 1999, 86, 226–230. [Google Scholar]
- de Maria, L; Vind, J; Oxenball, KM; Svendsen, A; Patkar, S. Phospholipases and their industrial applications. Appl. Microbiol. Biotechnol 2007, 74, 290–300. [Google Scholar]
- Nishihara, M; Kamata, M; Koyama, T; Yazawa, K. New phospholipase A1-producing bacteria from a marine fish. Mar. Biotechnol. (N. Y ) 2008, 10, 382–387. [Google Scholar]
- Shiba, Y; Ono, C; Fukui, F; Watanabe, I; Serizawa, N; Gomi, K; Yoshikawa, H. High-level secretory production of phospholipase A1 by Saccharomyces cerevisiae and Aspergillus oryzae. Biosci. Biotechnol. Biochem 2001, 65, 94–101. [Google Scholar]

{kind=link}
{kind=link}
| Classification | Organism | Name 2 | Location | Cellular Localization | ~ Size (kDa) | Substrate Specificity | Catalytic Properties | Reference |
|---|---|---|---|---|---|---|---|---|
| Eukarya | ||||||||
| Animali: | Bos taurus | □ PA-PLA1 | Testis, Brain | Cytosolic | 98 | PA | SXSXG catalytic serine3 | [52,53] |
| Mus musculus | □ p125 | Ubiquitous | Cytosolic | 111 | nd4 | GXSXG motif | [54] | |
| Homo sapiens | □ KIAA0725p | Ubiquitous | Cytosolic | 81 | PA, PE | GXSXG catalytic serine3 | [55] | |
| Homo sapiens | • mPA-PLA1α | Various tissues | Secreted | 58 | PA | Catalytic triad5 | [43,56] | |
| • mPA-PLA1β | Reproductive tissues | Mem-Ass | 58 | PA | Catalytic triad5 | [43,56] | ||
| PLRP2 | [50] | |||||||
| Rattus norvegicus | • PS-PLA1 | Platelets+various | Secreted | 55 | PS, lysoPS | Catalytic triad5 | [46,48,57] | |
| Dolichovespula maculate | • Dol m I | Venom sac | Secreted | 34 | nd4 | Catalytic triad5 | [47] | |
| Vespula spp. | • Ves v I | Venom sac | Secreted | 34 | nd4 | Catalytic triad 5 | [58–60] | |
| • Ves m I | Venom sac | Secreted | 34 | nd4 | Catalytic triad 5 | [58–60] | ||
| Caenorhabditis Elegans | IPLA-1 | ER | 87 | PI | Catalytic triad 5 | [5] | ||
| Plantae: | Arabidopsis thaliana | DAD1 | Anthers | Chloroplast | 45 | PC6 | GXSXG motif | [61] |
| AtLCAT3 | nd4 | Microsomes | 46 | PC, PE, PA | SXSXG-catalytic triad3 | [62,63] | ||
| Fungi: | Aspergillus oryzae | AoPLA1 | n/a4 | Secreted | 36 | nd4 | GXSXG motif | [64] |
| Thermomyces lanuginosus | n/a4 | nd4 | PC | [65] | ||||
| Protozoa: | Trypanosoma brucei | TbPLA1 | n/a4 | Cytosolic | 34 | PC | GXSXG catalytic serine 3 | [49,66] |
| Prokarya | ||||||||
| Bacteria: | Serratia spp. | ♦ PhlA | n/a | Secreted | 34 | nd4 | GXSXG motif | [67,68] |
| ♦ PlaA | n/a | Secreted | 34 | nd4 | GXSXG motif | [69] | ||
| Yersinia enterocolitica | ♦ YplA | n/a | Secreted | 34 | nd4 | GXSXG motif | [70] | |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Richmond, G.S.; Smith, T.K. Phospholipases A1. Int. J. Mol. Sci. 2011, 12, 588-612. https://doi.org/10.3390/ijms12010588
Richmond GS, Smith TK. Phospholipases A1. International Journal of Molecular Sciences. 2011; 12(1):588-612. https://doi.org/10.3390/ijms12010588
Chicago/Turabian StyleRichmond, Gregory S., and Terry K. Smith. 2011. "Phospholipases A1" International Journal of Molecular Sciences 12, no. 1: 588-612. https://doi.org/10.3390/ijms12010588
APA StyleRichmond, G. S., & Smith, T. K. (2011). Phospholipases A1. International Journal of Molecular Sciences, 12(1), 588-612. https://doi.org/10.3390/ijms12010588