Model Protocells from Single-Chain Lipids

{kind=link}
{kind=link}
Abstract
:1. Introduction
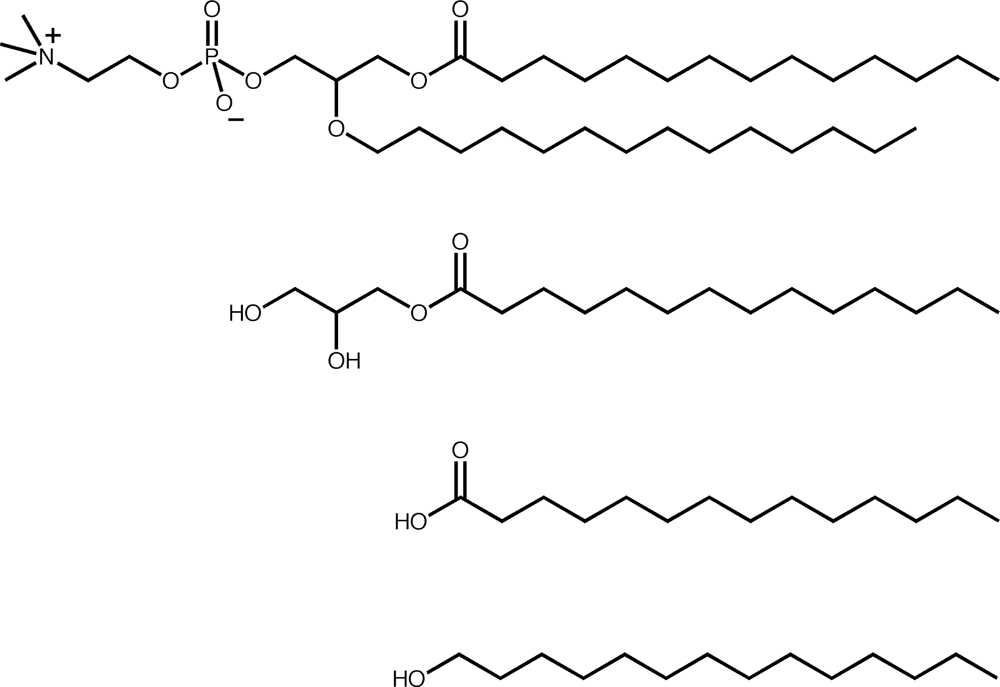
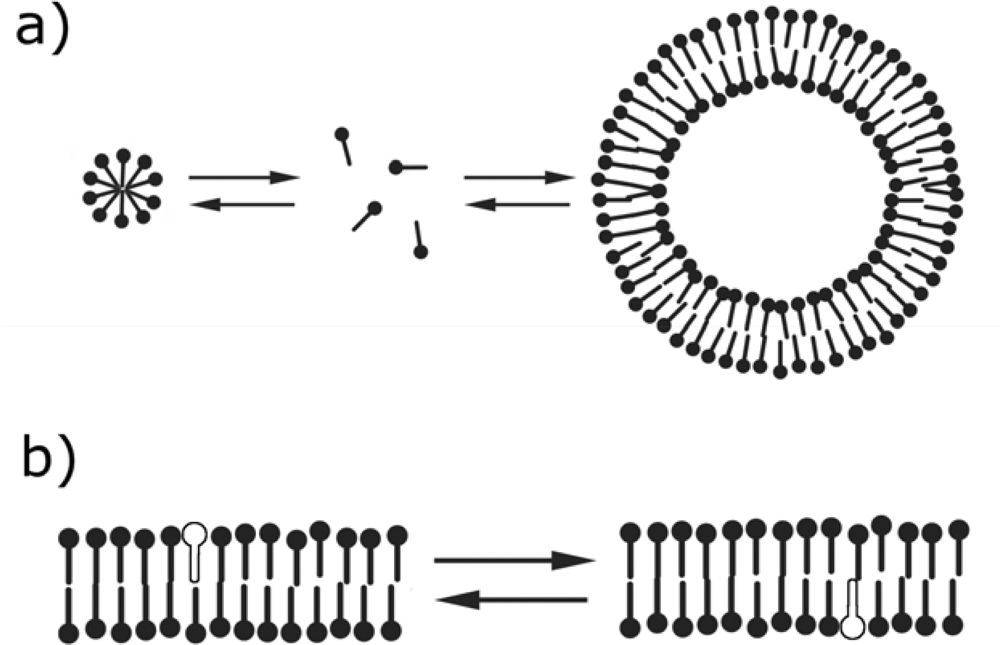
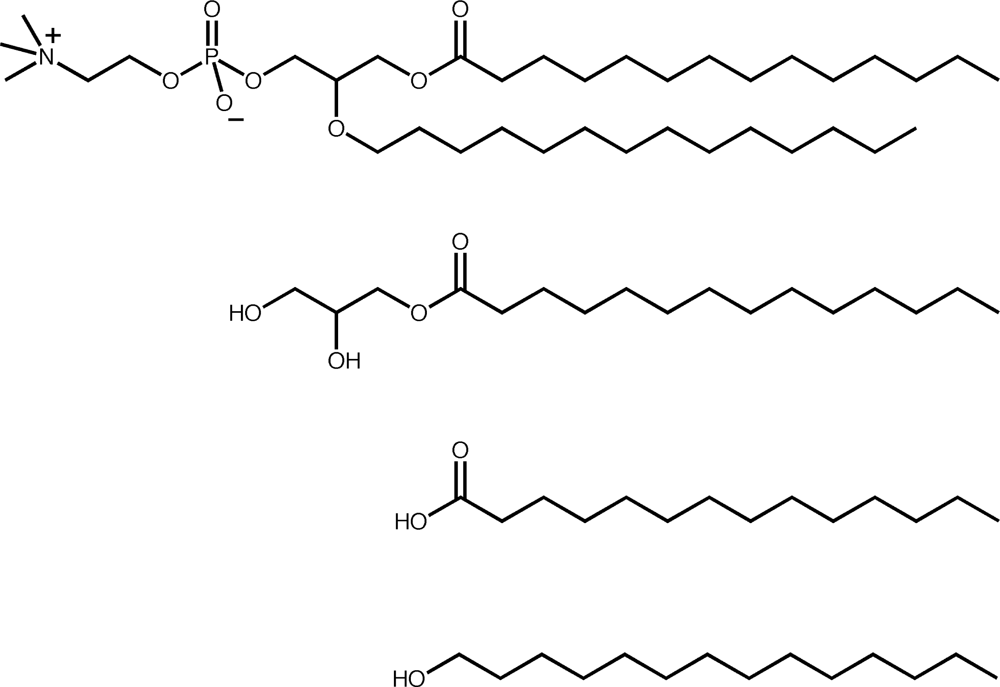
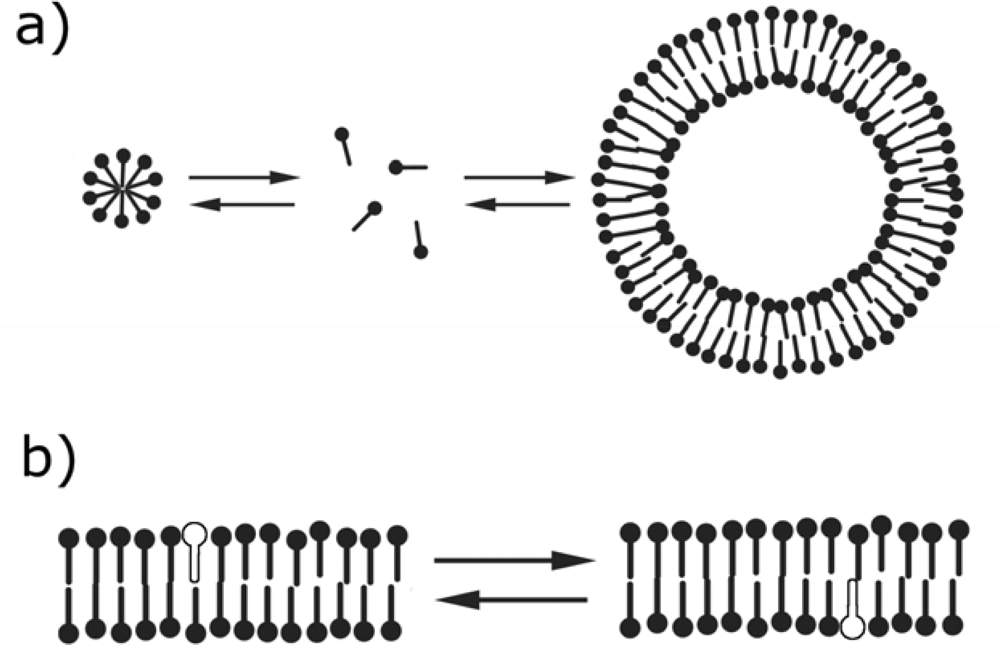
2. Similarities between fatty acid and phospholipid membranes
3. Differences between fatty acid and phospholipid membranes: lipid hydrophobicity
4. Differences between fatty acid and phospholipid membranes: lipid flip-flop
5. Membranes composed of mixtures of single-chain lipids
4. Conclusions
Acknowledgments
References
- de Duve, C. Singularities: Landmarks on the Pathways of Life; Cambridge University Press: New York, USA, 2005; p. 258. [Google Scholar]
- Morowitz, HJ. Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis; Yale University Press: New Haven, USA, 1992; p. 195. [Google Scholar]
- Szostak, JW; Bartel, DP; Luisi, PL. Synthesizing life. Nature 2001, 409, 387–390. [Google Scholar]
- Martin, W; Russell, MJ. On the origins of cells: A hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryoties, and from prokaryotes to nucleated cells. Philos. Trans. R Soc. Lond. B Biol. Sci 2003, 358, 59–83. [Google Scholar]
- McCollom, TM; Ritter, G; Simoneit, BR. Lipid synthesis under hydrothermal conditions by Fischer-Tropsch-type reactions. Orig. Life Evol. Biosph 1999, 29, 153–166. [Google Scholar]
- Pizzarello, S. The chemistry of life’s origin: A carbonaceous meteorite perspective. Acc. Chem. Res 2006, 39, 231–237. [Google Scholar]
- Deamer, DW. Boundary structures are formed by organic components of the Murchison carbonaceous chondrite. Nature 1985, 317, 792–794. [Google Scholar]
- Deamer, DW; Pasley, RM. Amphiphilic components of the Murchison carbonaceous chondrite: Surface properties and membrane formation. Orig. Life Evol. Biosphere 1989, 19, 21–38. [Google Scholar]
- Lawless, JG; Yuen, GU. Quantification of monocarboxylic acids in the Murchison carbonaceous meteorite. Nature 1979, 282, 396–398. [Google Scholar]
- Gebicki, JM; Hicks, M. Ufasomes are stable particles surrounded by unsaturated fatty acid membranes. Nature 1973, 243, 232–234. [Google Scholar]
- Gebicki, JM; Hicks, M. Preparation and properties of vesicles enclosed by fatty acid membranes. Chem. Phys. Lipids 1976, 16, 142–146. [Google Scholar]
- Hargreaves, WR; Deamer, DW. Liposomes from ionic, single-chain amphiphiles. Biochemistry 1978, 17, 3759–3768. [Google Scholar]
- Monnard, PA; Deamer, DW. Membrane self-assembly processes: Steps toward the first cellular life. Anat. Rec 2002, 268, 196–207. [Google Scholar]
- Morigaki, K; Walde, P. Fatty acid vesicles. Curr. Opin. Colloid Interface Sci 2007, 12, 75–80. [Google Scholar]
- Hanzyc, MM; Toyota, T; Ikegami, T; Packard, N; Sugawara, T. Fatty acid chemistry at the oil-water interface: Self-propelled oil droplets. J. Am. Chem. Soc 2007, 129, 9386–9391. [Google Scholar]
- Mansy, SS; Szostak, JW. Thermostability of model protocell membranes. Proc. Natl. Acad. Sci. USA 2008, 105, 13351–13355. [Google Scholar]
- Walde, P; Goto, A; Monnard, PA; Wessicken, M; Luisi, PL. Oparin’s reactions revisited: enzymatic synthesis of poly(adenylic acid) in micelles and self-reproducing vesicles. J. Am. Chem. Soc 1994, 116, 7541–7547. [Google Scholar]
- Chen, IA; Salehi-Ashtiani, K; Szostak, JW. RNA catalysis in model protocell vesicles. J. Am. Chem. Soc 2005, 127, 13213–13219. [Google Scholar]
- Chen, IA; Roberts, RW; Szostak, JW. The emergence of competition between model protocells. Science 2004, 305, 1474–1476. [Google Scholar]
- Sacerdote, MG; Szostak, JW. Semipermeable lipid bilayers exhibit diastereoselectivity favoring ribose. Proc. Natl. Acad. Sci. USA 2005, 102, 6004–6008. [Google Scholar]
- Deamer, DW. The first living systems: A bioenergetic perspective. Microbiol. Mol. Biol. Rev 1997, 61, 239–261. [Google Scholar]
- Israelachvili, JN. Intermolecular & Surface Forces, 2nd ed; Academic Press: London, UK, 1991; p. 450. [Google Scholar]
- Dejanovic, B; Mirosavljevic, K; Noethig-laslo, V; Pecar, S; Sentjurc, M; Walde, P. An ESR characterization of micelles and vesicles formed in aqueous decanoic acid/sodium decanoate systems using different spin labels. Chem. Phys. Lipids 2008, 156, 17–25. [Google Scholar]
- Fukuda, H; Goto, A; Yoshioka, H; Goto, R; Morigaki, K; Walde, P. Electron Spin Resonance study of the pH-induced transformation of micelles to vesicles in an aqueous oleic acid/oleate system. Langmuir 2001, 17, 4223–4231. [Google Scholar]
- Morigaki, K; Walde, P; Misran, M; Robinson, BH. Thermodynamic and kinetic stability. Properties of micelles and vesicles formed by the decanoic acid/decanoate system. Colloids Surf. A 2003, 213, 37–44. [Google Scholar]
- Chen, IA. The emergence of cells during the origin of life. Science 2006, 314, 1558–1559. [Google Scholar]
- McLean, LR; Phillips, MC. Mechanism of cholesterol and phosphatidylcholine exchange or transfer between unilamellar vesicles. Biochemistry 1981, 20, 2893–2900. [Google Scholar]
- Chen, IA; Szostak, JW. Membrane growth can generate a transmembrane pH gradient in fatty acid vesicles. Proc. Natl. Acad. Sci. USA 2004, 101, 7965–7970. [Google Scholar]
- Hamilton, JA. Fast flip-flop of cholesterol and fatty acids in membranes: Implications for membrane transport proteins. Curr. Opin. Lipidol 2003, 14, 263–271. [Google Scholar]
- Streiff, S; Ribeiro, N; Wu, Z; Gumienna-Kontecka, E; Elhabiri, M; Albrecht-Gary, AM; Ourisson, G; Nakatani, Y. “Primitive” membrane from polyprenyl phosphates and polyprenyl alcohols. Chem. Biol 2007, 14, 313–319. [Google Scholar]
- Walde, P; Wessicken, M; Radler, U; Berclaz, N; Conde-Frieboes, K; Luisi, PL. Preparation and characterization of vesicles from mono-n-alkyl phosphates and phosphonates. J. Phys. Chem. B 1997, 101, 7390–7397. [Google Scholar]
- Hanczyc, MM; Fujikawa, SM; Szostak, JW. Experimental models of primitative cellular compartments: encapsulation, growth, and division. Science 2003, 302, 618–622. [Google Scholar]
- Walde, P; Wick, R; Fresta, M; Mangone, A; Luisi, PL. Autopoietic self-reproduction of fatty acid vesicles. J. Am. Chem. Soc 1994, 116, 11649–11654. [Google Scholar]
- Luisi, PL; Stano, P; Rasi, S; Mavelli, F. A possible route to prebiotic vesicle reproduction. Artifical Life 2004, 10, 297–308. [Google Scholar]
- Chen, IA; Szostak, JW. A kinetic study of the growth of fatty acid vesicles. Biophys. J 2004, 87, 988–998. [Google Scholar]
- Mansy, SS; Schrum, JP; Krishnamurthy, M; Tobe, S; Treco, DA; Szostak, JW. Replication of a genetic polymer inside of a model protocell. Nature 2008, 454, 122–125. [Google Scholar]
- Cisse, I; Okumus, B; Joo, C; Ha, T. Fueling protein-DNA interactions inside porous nanocontainers. Proc. Natl. Acad. Sci. USA 2007, 104, 12646–12650. [Google Scholar]
- Monnard, PA; Deamer, DW. Nutrient uptake by protocells: a liposome model system. Orig. Life Evol. Biosph 2001, 31, 147–155. [Google Scholar]
- Monnard, PA; Luptak, A; Deamer, DW. Models of primitive cellular life: polymerases and templates in liposomes. Philos. Trans. R Soc. Lond. B Biol. Sci 2007, 362, 1741–1750. [Google Scholar]
- Apel, CL; Deamer, DW; Mautner, MN. Self-assembled vesicles of monocarboxylic acids and alcohols: conditions for stability and for the encapsulation of biopolymers. Biochim. et Biophys. Acta 2002, 1559, 1–9. [Google Scholar]
- Monnard, PA; Apel, CL; Kanavarioti, A; Deamer, DW. Influence of ionic inorganic solutes on self-assembly and polymerization processes related to early forms of life: Implications for a prebiotic aqueous medium. Astrobiology 2002, 2, 139–152. [Google Scholar]
- Namani, T; Deamer, DW. Stability of model membranes in extreme environments. Orig. Life Evol. Biosph 2008, 38, 329–341. [Google Scholar]
- Chen, IA; Hanzyc, MM; Sazani, PL; Szostak, JW. Protocells: Genetic polymers inside membrane vesicles; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, USA, 2006; pp. 57–88. [Google Scholar]
- Cowan, JA. Inorganic Biochemistry, 2nd ed; Wiley-VCH: New York, USA, 1997; p. 440. [Google Scholar]
- Wilson, MA; Pohorille, A. Mechanism of unassisted ion transport across membrane bilayers. J. Am. Chem. Soc 1996, 118, 6580–6587. [Google Scholar]
- Yoyoshima, Y; Thompson, TE. Chloride flux in bilayer membranes: Chloride permeability in aqueous dispersions of single-walled, bilayer vesicles. Biochemistry 1975, 14, 1525. [Google Scholar]
- Luisi, PL. Autopoiesis: A review and a reappraisal. Naturwissenschaften 2003, 90, 49–59. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mansy, S.S. Model Protocells from Single-Chain Lipids. Int. J. Mol. Sci. 2009, 10, 835-843. https://doi.org/10.3390/ijms10030835
Mansy SS. Model Protocells from Single-Chain Lipids. International Journal of Molecular Sciences. 2009; 10(3):835-843. https://doi.org/10.3390/ijms10030835
Chicago/Turabian StyleMansy, Sheref S. 2009. "Model Protocells from Single-Chain Lipids" International Journal of Molecular Sciences 10, no. 3: 835-843. https://doi.org/10.3390/ijms10030835
APA StyleMansy, S. S. (2009). Model Protocells from Single-Chain Lipids. International Journal of Molecular Sciences, 10(3), 835-843. https://doi.org/10.3390/ijms10030835