
Angiotensin-Converting Enzyme (ACE)-Inhibitor Activity of Novel Peptides Derived from Porcine Liver and Placenta



Abstract
1. Introduction
2. Results
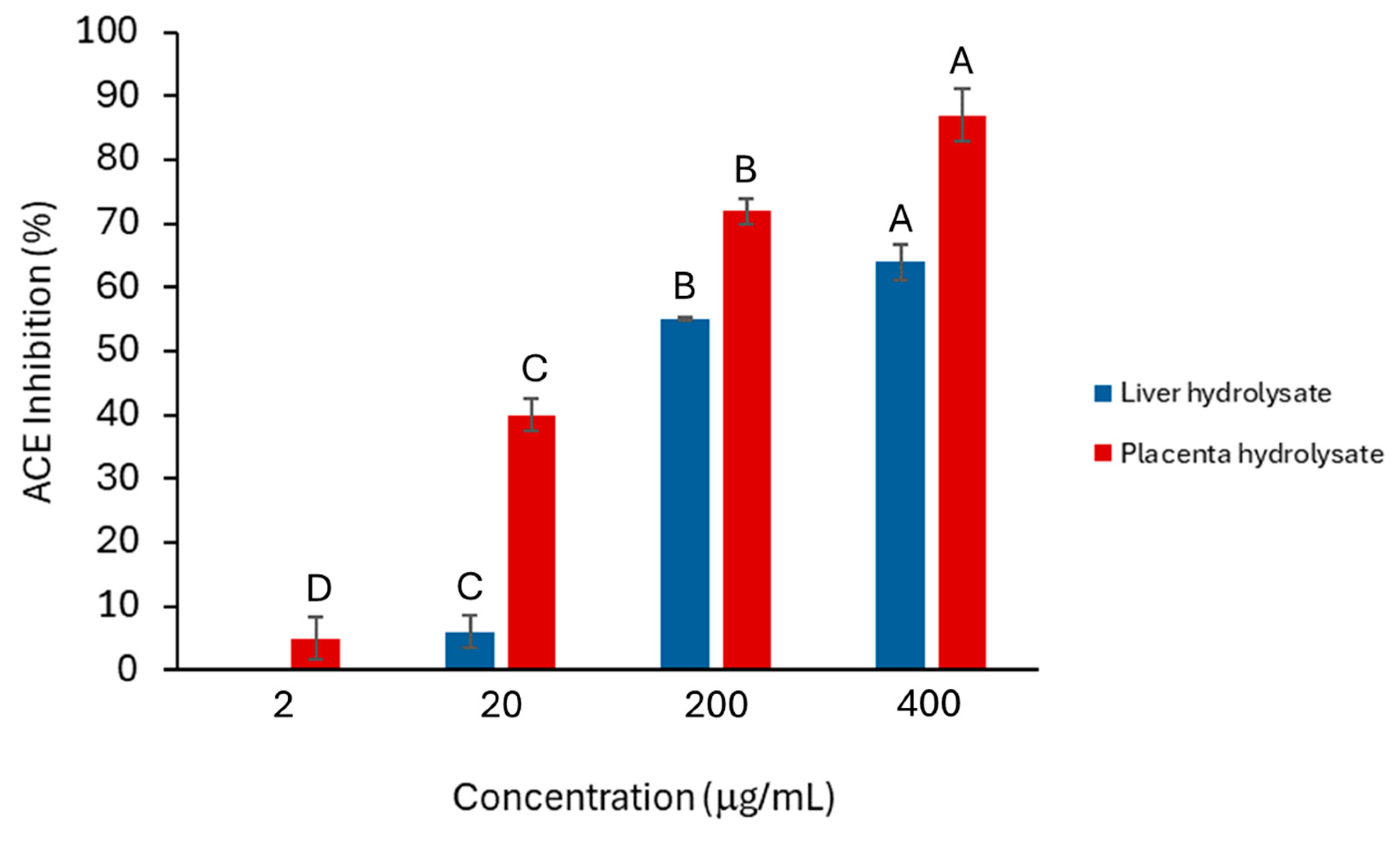
2.1. ACE-Inhibition Activity of Crude Liver and Placenta Hydrolysates
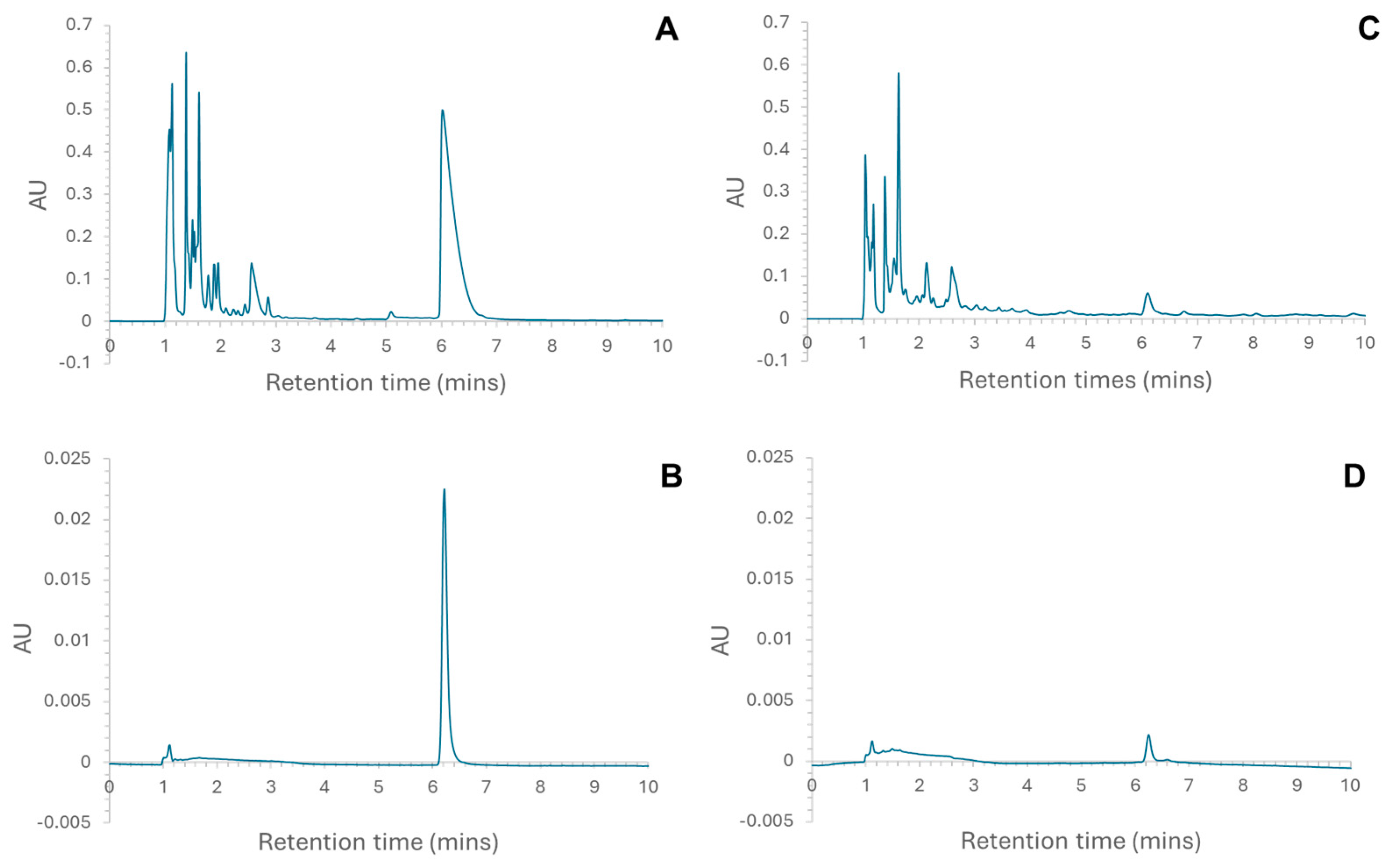
2.2. Peptide Sequencing Using MS/MS
2.3. In Silico Papain Digestion and Activity Prediction
2.4. Synthesised Peptides
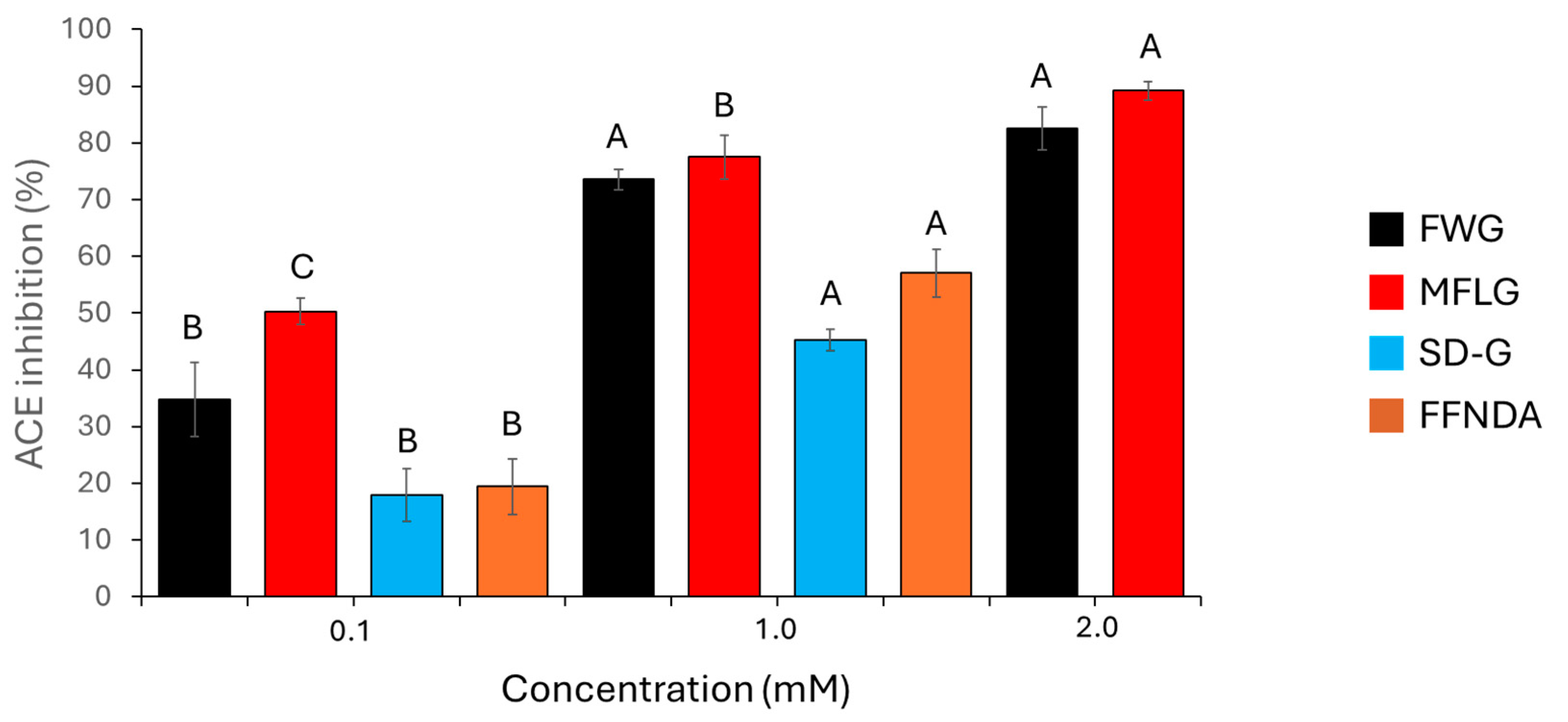
2.5. ACE Inhibition Assay of Synthetic Peptides
3. Discussion
4. Materials and Methods
4.1. Isolation of Individual Fractions for MS Analysis
4.2. Peptide Sequencing Using MS/MS
4.3. In Silico Digestion, and Activity Prediction
4.4. Peptide Synthesis and Preparation
- FWG: 10 mg/mL = 24.48 mM therefore 10 mM = 4.085 mg/mL
- MFLG: 10 mg/mL = 21.43 mM therefore10 mM = 4.666 mg/mL
- SDPPLVFVG: 10 mg/mL = 10.75 mM therefore 10 mM = 9.302 mg/mL
- FFNDA: 10 mg/mL = 16.32 mM or10 mM = 6.127 mg/mL
4.5. Angiotensin-Converting Enzyme (ACE)-Inhibition Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rawlings, N.D.; Barrett, A.J. Families of cysteine peptidases. Methods Enzymol. 1994, 244, 461–486. [Google Scholar] [PubMed]
- Arshad, Z.I.; Amid, A.; Yusof, F.; Jaswir, I.; Ahmad, K.; Loke, S.P. Bromelain: An overview of industrial application and purification strategies. Appl. Microbiol. Biotechnol. 2014, 98, 7283–7297. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.S.; El-Bassiony, T.; Elmalt, L.M.; Ibrahim, H.R. Identification of potent antioxidant bioactive peptides from goat milk proteins. Food Res. Int. 2015, 74, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, A.P.F.; Daroit, D.J.; Fontoura, R.; Meira, S.M.M.; Segalin, J.; Brandelli, A. Hydrolysates of sheep cheese whey as a source of bioactive peptides with antioxidant and angiotensin-converting enzyme inhibitory activities. Peptides 2014, 61, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Mooney, C.; Haslam, N.J.; Pollastri, G.; Shields, D.C. Towards the Improved Discovery and Design of Functional Peptides: Common Features of Diverse Classes Permit Generalized Prediction of Bioactivity. PLoS ONE 2012, 7, e45012. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef]
- Jang, A.; Lee, M. Purification and identification of angiotensin converting enzyme inhibitory peptides from beef hydrolysates. Meat Sci. 2005, 69, 653–661. [Google Scholar] [CrossRef]
- Pearman, N.A.; Ronander, E.; Smith, A.M.; Morris, G.A. The identification and characterisation of novel bioactive peptides derived from porcine liver. Curr. Res. Food Sci. 2020, 3, 314–321. [Google Scholar] [CrossRef]
- Samaei, S.P.; Martini, S.; Tagliazucchi, D.; Gianotti, A.; Babini, E. Antioxidant and angiotensin I-converting enzyme (ACE) Inhibitory peptides obtained from alcalase protein hydrolysate fractions of hemp (Cannabis sativa L.) bran. J. Agric. Food Chem. 2021, 69, 9220–9228. [Google Scholar] [CrossRef]
- Vulupala, H.R.; Sajja, Y.; Bagul, P.K.; Bandla, R.; Nagarapu, L.; Benerjee, S.K. Potent ACE inhibitors from 5-hydroxy indanone derivatives. Bioorg. Chem. 2018, 77, 660–665. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2016, 45, D158–D169. [Google Scholar]
- Himaya, S.W.A.; Ngo, D.-H.; Ryu, B.; Kim, S.-K. An active peptide purified from gastrointestinal enzyme hydrolysate of Pacific cod skin gelatin attenuates angiotensin-1 converting enzyme (ACE) activity and cellular oxidative stress. Food Chem. 2012, 132, 1872–1882. [Google Scholar] [CrossRef]
- Vercruysse, L.; Van Camp, J.; Smagghe, G. ACE inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem. 2005, 53, 8106–8115. [Google Scholar] [CrossRef] [PubMed]
- Arihara, K.; Nakashima, Y.; Mukai, T.; Ishikawa, S.; Itoh, M. Peptide inhibitors for angiotensin I-converting enzyme from enzymatic hydrolysates of porcine skeletal muscle proteins. Meat Sci. 2001, 57, 319–324. [Google Scholar] [CrossRef]
- Katayama, K.; Tomatsu, M.; Fuchu, H.; Sugiyama, M.; Kawahara, S.; Yamauchi, K.; Kawamura, Y.; Muguruma, M. Purification and characterization of an angiotensin I-converting enzyme inhibitory peptide derived from porcine troponin C. Anim. Sci. J. 2003, 74, 53–58. [Google Scholar] [CrossRef]
- Muguruma, M.; Ahhmed, A.M.; Katayama, K.; Kawahara, S.; Maruyama, M.; Nakamura, T. Identification of pro-drug type ACE inhibitory peptide sourced from porcine myosin B: Evaluation of its antihypertensive effects in vivo. Food Chem. 2009, 114, 516–522. [Google Scholar] [CrossRef]
- Miyoshi, S.; Ishikawa, H.; Kaneko, T.; Fukui, F.; Tanaka, H.; Maruyama, S. Structures and activity of angiotensin-converting enzyme inhibitors in an α-zein hydrolysate. Agric. Biol. Chem. 1991, 55, 1313–1318. [Google Scholar]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin i-converting enzyme inhibitory peptides: Quantitative structure−activity relationship study of di- and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef]
- Hatakenaka, T.; Kato, T.; Okamoto, K. In vitro and in silico studies on angiotensin I-converting enzyme inhibitory peptides found in hydrophobic domains of porcine elastin. Molecules 2023, 28, 3337. [Google Scholar] [CrossRef]
- Mora, L.M.G.; Toldrá, F. ACEI-inhibitory peptides naturally generated in meat and meat products and their health relevance. Nutrients 2018, 10, 1259. [Google Scholar] [CrossRef]
- Katayama, K.; Anggraeni, H.E.; Mori, T.; Ahhmed, A.M.; Kawahara, S.; Sugiyama, M.; Nakayama, T.; Maruyama, M.; Muguruma, M. Porcine skeletal muscle troponin is a good source of peptides with angiotensin-I converting enzyme inhibitory activity and antihypertensive effects in spontaneously hypertensive rats. J. Agric. Food Chem. 2008, 56, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Mirdhayati, I.; Hermanianto, J.; Wijaya, C.H.; Sajuthi, D.; Arihara, K. Angiotensin converting enzyme (ACE) inhibitory and antihypertensive activities of protein hydrolysate from meat of Kacang goat (Capra aegagrus hircus). J. Sci. Food Agric. 2016, 96, 3536–3542. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, T.; Yamanaka, T.A.; Otsuka, T.; Yamashita, E.; Maruyama, S. Antihypertensive effect of enzymatic hydrolysate of collagen and Gly-Pro in spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2009, 73, 2317–2319. [Google Scholar] [CrossRef] [PubMed]
- Escudero, E.; Toldrá, F.; Sentandreu, M.A.; Nishimura, H.; Arihara, K. Antihypertensive activity of peptides identified in the in vitro gastrointestinal digest of pork meat. Meat Sci. 2012, 91, 382–384. [Google Scholar] [CrossRef]
- Aydin, F.; Turkoglu, V.; Bas, Z. Purification and characterization of angiotensin-converting enzyme (ACE) from sheep lung. Mol. Biol. Rep. 2021, 48, 4191–4199. [Google Scholar] [CrossRef]
- Lassoued, I.; Mora, L.; Barkia, A.; Aristoy, M.-C.; Nasri, M.; Toldrá, F. Bioactive peptides identified in thornback ray skin’s gelatin hydrolysates by proteases from Bacillus subtilis and Bacillus amyloliquefaciens. J. Proteom. 2015, 128, 8–17. [Google Scholar] [CrossRef]
- Lafarga, T.; O’Connor, P.; Hayes, M. Identification of novel dipeptidyl peptidase-IV and angiotensin-I-converting enzyme inhibitory peptides from meat proteins using in silico analysis. Peptides 2014, 59, 53–62. [Google Scholar] [CrossRef]
- Cheung, I.W.Y.; Nakayama, S.; Hsu, M.N.K.; Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Angiotensin-I converting enzyme inhibitory activity of hydrolysates from oat (Avena sativa) proteins by in silico and in vitro analyses. J. Agric. Food Chem. 2009, 57, 9234–9242. [Google Scholar] [CrossRef]
- Chatterjee, A.; Kanawjia, S.K.; Khetra, Y.; Saini, P. Discordance between in silico & in vitro analyses of ACE inhibitory & antioxidative peptides from mixed milk tryptic whey protein hydrolysate. J. Food Sci. Technol. 2015, 52, 5621–5630. [Google Scholar]
- Burkhart, J.M.; Schumbrutzki, C.; Wortelkamp, S.; Sickmann, A.; Zahedi, R.P. Systematic and quantitative comparison of digest efficiency and specificity reveals the impact of trypsin quality on MS-based proteomics. J. Proteom. 2012, 75, 1454–1462. [Google Scholar] [CrossRef]
- Picotti, P.; Aebersold, R.; Domon, B. The implications of proteolytic background for shotgun proteomics. Mol. Cell. Proteom. 2007, 6, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-converting enzyme (ACE)-Inhibitory peptides from plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, B.J.; Mugesh, G. Effect of peptide-based captopril analogues on angiotensin converting enzyme activity and peroxynitrite-mediated tyrosine nitration. Org. Biomol. Chem. 2011, 9, 5185–5192. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.S.; Ko, W.C. Angiotensin I- converting enzyme inhibitory activity of protein hydrolysate from Tuna broth. J. Food Compos. Anal. 2004, 12, 232–237. [Google Scholar] [CrossRef]
- Lee, S.Y.; Hur, S.J. Purification of novel angiotensin converting enzyme inhibitory peptides from beef myofibrillar proteins and analysis of their effect in spontaneously hypertensive rat model. Biomed. Pharmacother. 2019, 116, 109046. [Google Scholar] [CrossRef]
- Hayes, M.; Mora, L. Alternative proteins as a source of bioactive peptides: The edible snail and generation of hydrolysates containing peptides with bioactive potential for use as functional foods. Foods 2021, 10, 276. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM database of bioactive peptides: Current opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Raffaella, C.; Pellicorio, V.; Barberis, M.; Frosi, I.; Papetti, A. Recent advances in the valorization of seed wastes as source of bioactive peptides with multifunctional properties. Trends Food Sci. 2024, 144, 104322. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Papain Digestion | ||
|---|---|---|
| Cytosol Aminopeptidase- Derived Peptides | Haemoglobin Subunit Alpha-Derived Peptides | Type IV Collagen Alpha-1 Chain Partial-Derived Peptides |
| FWG (1.00) * | MFLG (0.96) * | FFNDA (0.77) * |
| FLLR (0.86) | FPH (0.94) | FSSPA (0.65) |
| WQR (0.84) | FLA (0.86) | - |
| FLK (0.81) | - | - |
| NWH (0.78) | - | - |
| SDPPLVFVG (0.75) * | - | - |
| Peptide * | ACE Inhibition IC50 (μM) | Protein Source | References |
|---|---|---|---|
| FWG | 470 | Cytosol aminopeptidase | This study |
| MFLG | 70 | Haemoglobin subunit alpha | This study |
| SDPPLVFVG | 1160 | Cytosol aminopeptidase | This study |
| FFNDA | 830 | Type IV collagen alpha-1 chain | This study |
| ITTNP | 549 | Myosin | [13,14] |
| MNPPK | 946 | Myosin | [13,14] |
| MNP | 67 | Myosin | [13,14] |
| NPP | 291 | Myosin | [13,14] |
| PPK | >1000 | Myosin | [13,14] |
| ITT | 678 | Myosin | [13,14] |
| TTN | 673 | Myosin | [13,14] |
| TNP | 207 | Myosin | [13,14] |
| RMLGQTPTK | 34 | Troponin C | [13,15] |
| RMLGQTP | 503 | Troponin C | [13,15] |
| KRVITY | 6 | Myosin | [16] |
| VKAGF | 20 | Actin | [16] |
| VAP | 1 | Tropoelastin | [17] |
| APG | 1194 | Tropoelastin | [17] |
| GAP | 35 | Tropoelastin | [17] |
| VSP | 10 | Tropoelastin | [17] |
| TRP | 1 | Tropoelastin | [17] |
| LSP | 2 | Tropoelastin | [17,18] |
| VGP | 26 | Tropoelastin | [17,18] |
| VLP | 4 | Tropoelastin | [17,19] |
| GVG | 369 | Tropoelastin | [17] |
| GLG | 485 | Tropoelastin | [17] |
| VKKVLGNP | 29 | Myosin (light chain) | [20,21] |
| GF(Hyp)GP | 91 | Collagen | [22] |
| KRQKYDI | 27 | Troponin | [17,20] |
| RPR | 382 | Nebulin | [20,23] |
| KAPVA | 47 | Titin | [20,24] |
| PTPVP | 256 | Titin | [20,24] |
| Captopril | 5 × 10−4 | Commercial | [25] |
| Reagents | Volume (μL) | ||
|---|---|---|---|
| A | B | C | |
| Milli-Q Water | 0 | 40 | 15 |
| Inhibitor/Peptide | 40 | 0 | 40 |
| 0.5 mM HHL | 145 | 145 | 145 |
| Incubate at 37 °C for 3 min | |||
| 200 mU/mL ACE | 15 | 15 | 0 |
| Incubate at 37 °C for 30 min | |||
| 1 M HCl | 200 | 200 | 200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pearman, N.A.; Morris, G.A.; Smith, A.M. Angiotensin-Converting Enzyme (ACE)-Inhibitor Activity of Novel Peptides Derived from Porcine Liver and Placenta. Molecules 2025, 30, 754. https://doi.org/10.3390/molecules30030754
Pearman NA, Morris GA, Smith AM. Angiotensin-Converting Enzyme (ACE)-Inhibitor Activity of Novel Peptides Derived from Porcine Liver and Placenta. Molecules. 2025; 30(3):754. https://doi.org/10.3390/molecules30030754
Chicago/Turabian StylePearman, Nicholas A., Gordon A. Morris, and Alan M. Smith. 2025. "Angiotensin-Converting Enzyme (ACE)-Inhibitor Activity of Novel Peptides Derived from Porcine Liver and Placenta" Molecules 30, no. 3: 754. https://doi.org/10.3390/molecules30030754
APA StylePearman, N. A., Morris, G. A., & Smith, A. M. (2025). Angiotensin-Converting Enzyme (ACE)-Inhibitor Activity of Novel Peptides Derived from Porcine Liver and Placenta. Molecules, 30(3), 754. https://doi.org/10.3390/molecules30030754