
Intracellular Protective Functions and Therapeutical Potential of Trehalose


Abstract
1. Introduction
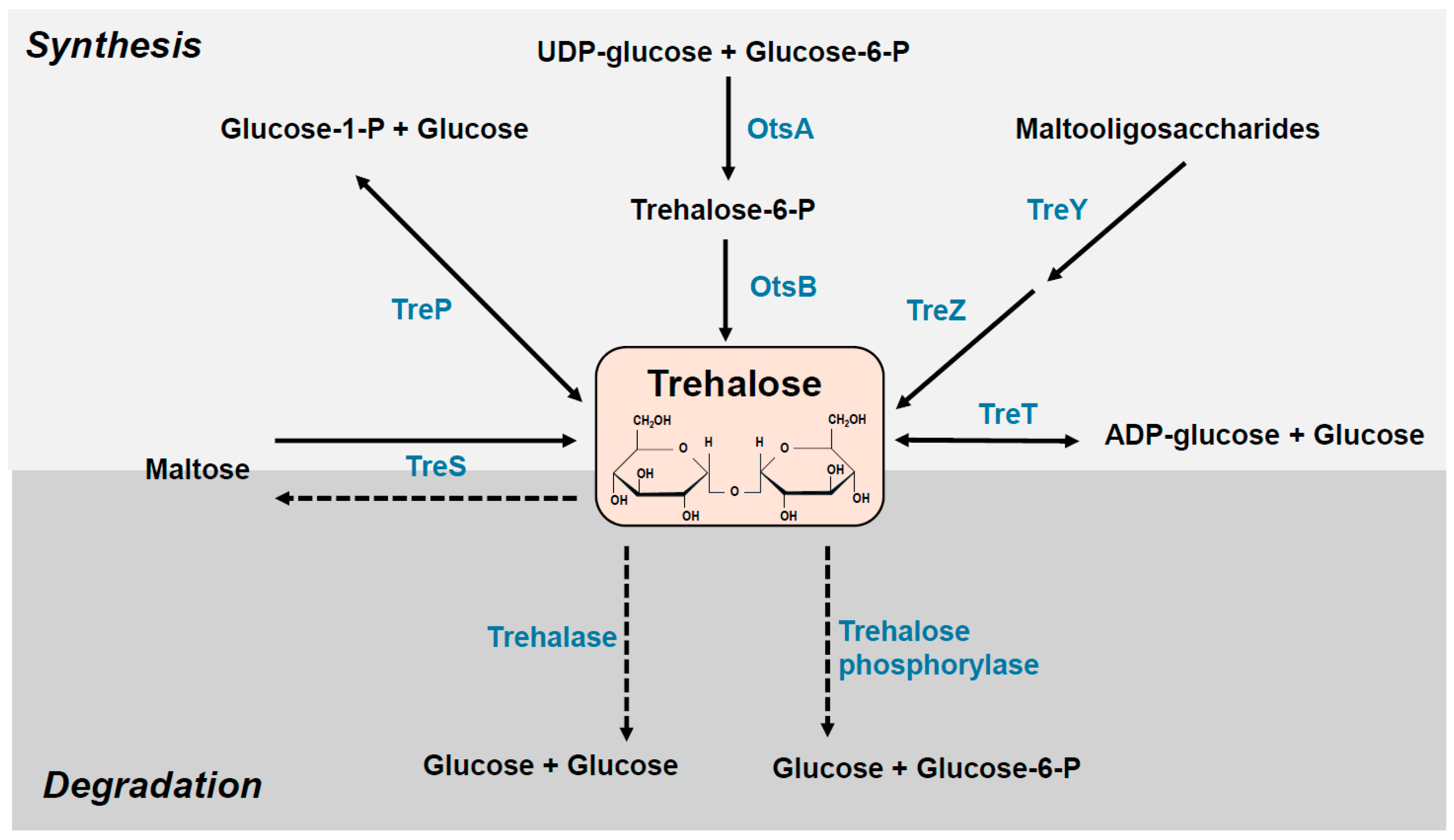
2. Mechanisms Governing the Accumulation of Trehalose
3. Intracellular Protective Functions of Trehalose
3.1. Trehalose as an Osmoprotectant and Chemical Chaperone
3.2. Trehalose as a Free Radicals Scavenger
3.3. Regulatory Functions of Trehalose
3.4. Trehalose and Neuroprotection
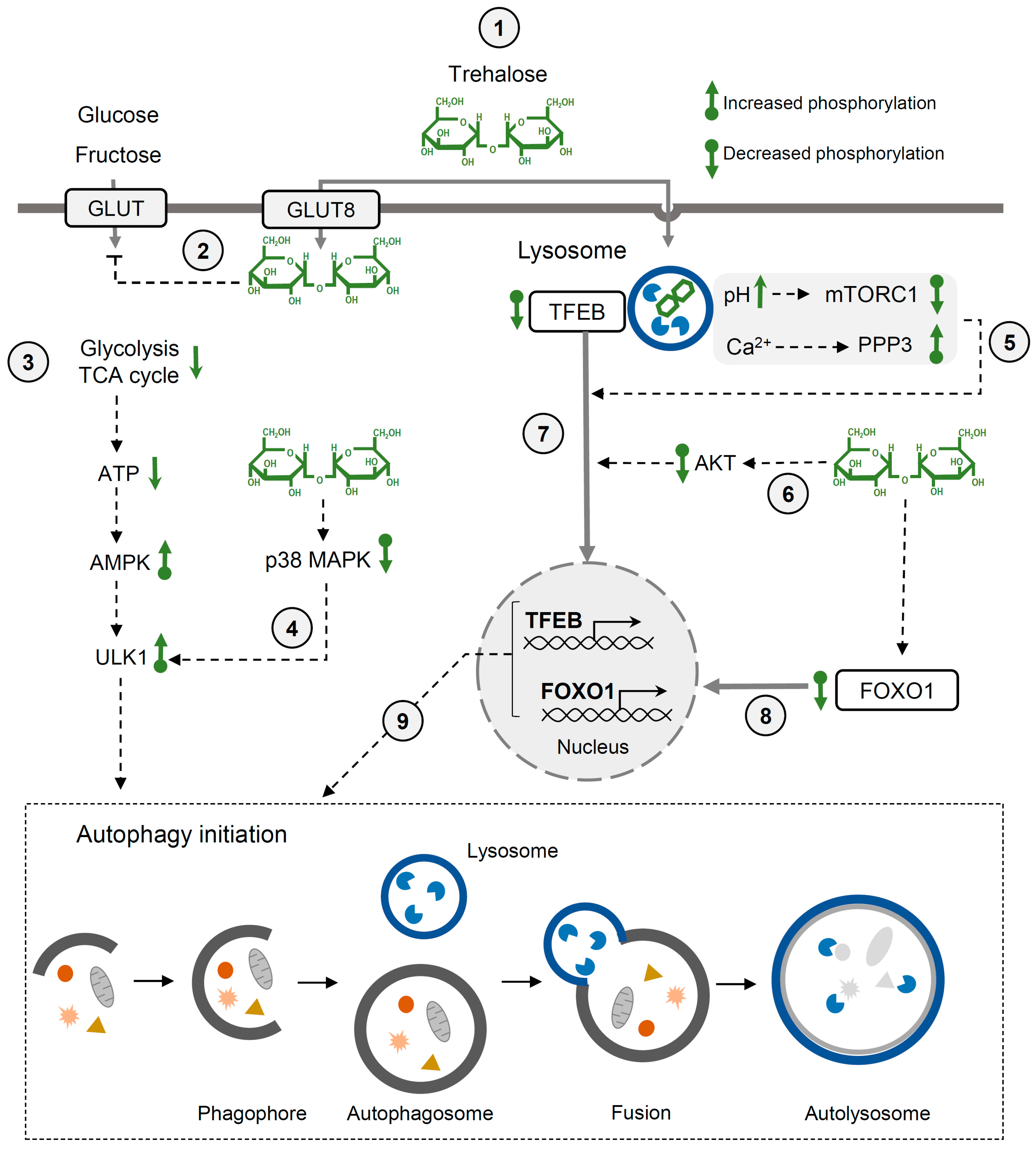
3.5. Trehalose and Autophagy
3.6. Trehalose and Glucose Homeostasis
4. Trehalose and Pathogenicity
5. Applications of Trehalose
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kumar, R. Role of Naturally Occurring Osmolytes in Protein Folding and Stability. Arch. Biochem. Biophys. 2009, 491, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.; Choi, I. Roles of Osmolytes in Protein Folding and Aggregation in Cells and Their Biotechnological Applications. Int. J. Biol. Macromol. 2018, 109, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Sleator, R.D.; Hill, C. Bacterial Osmoadaptation: The Role of Osmolytes in Bacterial Stress and Virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef]
- Teramoto, N.; Sachinvala, N.D.; Shibata, M. Trehalose and Trehalose-Based Polymers for Environmentally Benign, Biocompatible and Bioactive Materials. Molecules 2008, 13, 1773–1816. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.B.; Krakowka, S.; Dexter, L.B.; Schmid, H.; Wolterbeek, A.P.M.; Waalkens-Berendsen, D.H.; Shigoyuki, A.; Kurimoto, M. Trehalose: A Review of Properties, History of Use and Human Tolerance, and Results of Multiple Safety Studies. Food Chem. Toxicol. 2002, 40, 871–898. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Akiba, C.; Izawa, M.; Iwami, M. Steroid Hormone-Dependent Changes in Trehalose Physiology in the Silkworm, Bombyx mori. J. Comp. Physiol. B 2023, 193, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New Insights on Trehalose: A Multifunctional Molecule. Glycobiology 2003, 13, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Attfield, P.V. Trehalose Accumulates in Saccharomyces cerevisiae during Exposure to Agents That Induce Heat Shock Response. FEBS Lett. 1987, 225, 259–263. [Google Scholar] [CrossRef]
- De Virgilio, C.; Hottiger, T.; Dominguez, J.; Boller, T.; Wiemken, A. The Role of Trehalose Synthesis for the Acquisition of Thermotolerance in Yeast. Eur. J. Biochem. 1994, 219, 179–186. [Google Scholar] [CrossRef]
- Hohmann, S. Osmotic Stress Signaling and Osmoadaptation in Yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef]
- Maicas, S.; Guirao-Abad, J.P.; Argüelles, J.-C. Yeast Trehalases: Two Enzymes, One Catalytic Mission. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2016, 1860, 2249–2254. [Google Scholar] [CrossRef]
- Horlacher, R.; Uhland, K.; Klein, W.; Ehrmann, M.; Boos, W. Characterization of a Cytoplasmic Trehalase of Escherichia coli. J. Bacteriol. 1996, 178, 6250–6257. [Google Scholar] [CrossRef] [PubMed]
- Strom, A.R.; Kaasen, I. Trehalose Metabolism in Escherichia coli: Stress Protection and Stress Regulation of Gene Expression. Mol. Microbiol. 1993, 8, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Kandror, O.; DeLeon, A.; Goldberg, A.L. Trehalose Synthesis Is Induced upon Exposure of Escherichia coli to Cold and Is Essential for Viability at Low Temperatures. Proc. Natl. Acad. Sci. USA 2002, 99, 9727–9732. [Google Scholar] [CrossRef] [PubMed]
- Hengge-Aronis, R.; Klein, W.; Lange, R.; Rimmele, M.; Boos, W. Trehalose Synthesis Genes Are Controlled by the Putative Sigma Factor Encoded by RpoS and Are Involved in Stationary-Phase Thermotolerance in Escherichia coli. J. Bacteriol. 1991, 173, 7918–7924. [Google Scholar] [CrossRef] [PubMed]
- Iordachescu, M.; Imai, R. Trehalose Biosynthesis in Response to Abiotic Stresses. J. Integr. Plant Biol. 2008, 50, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Ruhal, R.; Kataria, R.; Choudhury, B. Trends in Bacterial Trehalose Metabolism and Significant Nodes of Metabolic Pathway in the Direction of Trehalose Accumulation. Microb. Biotechnol. 2013, 6, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Thammahong, A.; Puttikamonkul, S.; Perfect, J.R.; Brennan, R.G.; Cramer, R.A. Central Role of the Trehalose Biosynthesis Pathway in the Pathogenesis of Human Fungal Infections: Opportunities and Challenges for Therapeutic Development. Microbiol. Mol. Biol. Rev. 2017, 81, e00053-16. [Google Scholar] [CrossRef] [PubMed]
- De Smet, K.A.L.; Weston, A.; Brown, I.N.; Young, D.B.; Robertson, B.D. Three Pathways for Trehalose Biosynthesis in Mycobacteria. Microbiology 2000, 146, 199–208. [Google Scholar] [CrossRef]
- Wolf, A.; Krämer, R.; Morbach, S. Three Pathways for Trehalose Metabolism in Corynebacterium glutamicum ATCC13032 and Their Significance in Response to Osmotic Stress. Mol. Microbiol. 2003, 49, 1119–1134. [Google Scholar] [CrossRef]
- Tapia, H.; Young, L.; Fox, D.; Bertozzi, C.R.; Koshland, D. Increasing Intracellular Trehalose Is Sufficient to Confer Desiccation Tolerance to Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2015, 112, 6122–6127. [Google Scholar] [CrossRef]
- Gechev, T.S.; Dinakar, C.; Benina, M.; Toneva, V.; Bartels, D. Molecular Mechanisms of Desiccation Tolerance in Resurrection Plants. Cell Mol. Life Sci. 2012, 69, 3175–3186. [Google Scholar] [CrossRef]
- Koshland, D.; Tapia, H. Desiccation Tolerance: An Unusual Window into Stress Biology. Mol. Biol. Cell 2019, 30, 737–741. [Google Scholar] [CrossRef]
- França, M.B.; Panek, A.D.; Eleutherio, E.C.A. Oxidative Stress and Its Effects during Dehydration. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.; KC, S.; Gonzalez, T.; Tapia, H.; Boothby, T.C. Trehalose and Tardigrade CAHS Proteins Work Synergistically to Promote Desiccation Tolerance. Commun. Biol. 2022, 5, 1046. [Google Scholar] [CrossRef] [PubMed]
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.M.; Mundree, S.; Williams, B.; Bewley, J.D. Desiccation Tolerance: Avoiding Cellular Damage during Drying and Rehydration. Annu. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef] [PubMed]
- Corradini, D.; Strekalova, E.G.; Eugene Stanley, H.; Gallo, P. Microscopic Mechanism of Protein Cryopreservation in an Aqueous Solution with Trehalose. Sci. Rep. 2013, 3, 1218. [Google Scholar] [CrossRef]
- Shao, Q.; Wang, J.; Zhu, W. Trehalose Stabilizing Protein in a Water Replacement Scenario: Insights from Molecular Dynamics Simulation. bioRxiv 2019. [Google Scholar] [CrossRef]
- Diamant, S.; Eliahu, N.; Rosenthal, D.; Goloubinoff, P. Chemical Chaperones Regulate Molecular Chaperones in Vitro and in Cells under Combined Salt and Heat Stresses. J. Biol. Chem. 2001, 276, 39586–39591. [Google Scholar] [CrossRef]
- Tapia, H.; Koshland, D.E. Trehalose Is a Versatile and Long-Lived Chaperone for Desiccation Tolerance. Curr. Biol. 2014, 24, 2758–2766. [Google Scholar] [CrossRef]
- Leszczynska, D.; Matuszewska, E.; Kuczynska-Wisnik, D.; Furmanek-Blaszk, B.; Laskowska, E. The Formation of Persister Cells in Stationary-Phase Cultures of Escherichia coli Is Associated with the Aggregation of Endogenous Proteins. PLoS ONE 2013, 8, e54737. [Google Scholar] [CrossRef] [PubMed]
- Łupkowska, A.; Monem, S.; Dębski, J.; Stojowska-Swędrzyńska, K.; Kuczyńska-Wiśnik, D.; Laskowska, E. Protein Aggregation and Glycation in Escherichia coli Exposed to Desiccation-Rehydration Stress. Microbiol. Res. 2023, 270, 127335. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.R.; Poddar, N.K.; Dar, T.A.; Kumar, R.; Ahmad, F. Protein and DNA Destabilization by Osmolytes: The Other Side of the Coin. Life Sci. 2011, 88, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, W.-M.; Wang, W. Trehalose: Protector of Antioxidant Enzymes or Reactive Oxygen Species Scavenger under Heat Stress? Environ. Exp. Bot. 2008, 63, 378–384. [Google Scholar] [CrossRef]
- Benaroudj, N.; Lee, D.H.; Goldberg, A.L. Trehalose Accumulation during Cellular Stress Protects Cells and Cellular Proteins from Damage by Oxygen Radicals. J. Biol. Chem. 2001, 276, 24261–24267. [Google Scholar] [CrossRef]
- Herdeiro, R.S.; Pereira, M.D.; Panek, A.D.; Eleutherio, E.C.A. Trehalose Protects Saccharomyces Cerevisiae from Lipid Peroxidation during Oxidative Stress. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2006, 1760, 340–346. [Google Scholar] [CrossRef]
- Prasad, K.H.D.; Ebenezer, C.; Solomon, R.V.; Iyyappan, E. Deciphering the Structure-Property Relationship and Antioxidant Mechanisms of Trehalose–An in-Silico Approach. J. Mol. Struct. 2023, 1291, 135957. [Google Scholar] [CrossRef]
- Jain, N.K.; Roy, I. Effect of Trehalose on Protein Structure. Prot. Sci. 2009, 18, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Kuczyńska-Wiśnik, D.; Stojowska, K.; Matuszewska, E.; Leszczyńska, D.; Algara, M.M.; Augustynowicz, M.; Laskowska, E. Lack of Intracellular Trehalose Affects Formation of Escherichia coli Persister Cells. Microbiology 2015, 161, 786–796. [Google Scholar] [CrossRef]
- Moon, J.E.; Heo, W.; Lee, S.H.; Lee, S.H.; Lee, H.G.; Lee, J.H.; Kim, Y.J. Trehalose Protects the Probiotic Yeast Saccharomyces boulardii against Oxidative Stress-Induced Cell Death. J. Microbiol. Biotechnol. 2020, 30, 54–61. [Google Scholar] [CrossRef]
- da Costa Morato Nery, D.; da Silva, C.G.; Mariani, D.; Fernandes, P.N.; Pereira, M.D.; Panek, A.D.; Eleutherio, E.C.A. The Role of Trehalose and Its Transporter in Protection against Reactive Oxygen Species. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2008, 1780, 1408–1411. [Google Scholar] [CrossRef]
- Rohman, M.M.; Islam, M.R.; Monsur, M.B.; Amiruzzaman, M.; Fujita, M.; Hasanuzzaman, M. Trehalose Protects Maize Plants from Salt Stress and Phosphorus Deficiency. Plants 2019, 8, 568. [Google Scholar] [CrossRef] [PubMed]
- Blázquez, M.A.; Lagunas, R.; Gancedo, C.; Gancedo, J.M. Trehalose-6-phosphate, a New Regulator of Yeast Glycolysis That Inhibits Hexokinases. FEBS Lett. 1993, 329, 51–54. [Google Scholar] [CrossRef]
- Petitjean, M.; Teste, M.-A.; François, J.M.; Parrou, J.-L. Yeast Tolerance to Various Stresses Relies on the Trehalose-6P Synthase (Tps1) Protein, Not on Trehalose. J. Biol. Chem. 2015, 290, 16177–16190. [Google Scholar] [CrossRef] [PubMed]
- Ratnakumar, S.; Tunnacliffe, A. Intracellular Trehalose Is Neither Necessary nor Sufficient for Desiccation Tolerance in Yeast. FEMS Yeast Res. 2006, 6, 902–913. [Google Scholar] [CrossRef]
- Chen, A.; Smith, J.R.; Tapia, H.; Gibney, P.A. Characterizing Phenotypic Diversity of Trehalose Biosynthesis Mutants in Multiple Wild Strains of Saccharomyces cerevisiae. G3 2022, 12, jkac196. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Zhao, H.; Li, J.; Gong, Y.; Li, X. The Devastating Rice Blast Airborne Pathogen Magnaporthe Oryzae—A Review on Genes Studied with Mutant Analysis. Pathogens 2023, 12, 379. [Google Scholar] [CrossRef]
- Wilson, R.A.; Jenkinson, J.M.; Gibson, R.P.; Littlechild, J.A.; Wang, Z.-Y.; Talbot, N.J. Tps1 Regulates the Pentose Phosphate Pathway, Nitrogen Metabolism and Fungal Virulence. EMBO J. 2007, 26, 3673–3685. [Google Scholar] [CrossRef]
- Moruno Algara, M.; Kuczyńska-Wiśnik, D.; Dębski, J.; Stojowska-Swędrzyńska, K.; Sominka, H.; Bukrejewska, M.; Laskowska, E. Trehalose Protects Escherichia coli against Carbon Stress Manifested by Protein Acetylation and Aggregation. Mol. Microbiol. 2019, 112, 866–880. [Google Scholar] [CrossRef]
- Kuczyńska-Wiśnik, D.; Moruno-Algara, M.; Stojowska-Swȩdrzyńska, K.; Laskowska, E. The Effect of Protein Acetylation on the Formation and Processing of Inclusion Bodies and Endogenous Protein Aggregates in Escherichia coli Cells. Microb. Cell Fact. 2016, 15, 189. [Google Scholar] [CrossRef]
- Hentchel, K.L.; Escalante-Semerena, J.C. Acylation of Biomolecules in Prokaryotes: A Widespread Strategy for the Control of Biological Function and Metabolic Stress. Microbiol. Mol. Biol. Rev. 2015, 79, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Guo, L.; Fu, Y.; Huo, M.; Qi, Q.; Zhao, G. Bacterial Protein Acetylation and Its Role in Cellular Physiology and Metabolic Regulation. Biotechnol. Adv. 2021, 53, 107842. [Google Scholar] [CrossRef] [PubMed]
- Laskowska, E.; Kuczyńska-Wiśnik, D. New Insight into the Mechanisms Protecting Bacteria during Desiccation. Curr. Genet. 2020, 66, 313–318. [Google Scholar] [CrossRef]
- Béranger, F.; Crozet, C.; Goldsborough, A.; Lehmann, S. Trehalose Impairs Aggregation of PrPSc Molecules and Protects Prion-Infected Cells against Oxidative Damage. Biochem. Biophys. Res. Commun. 2008, 374, 44–48. [Google Scholar] [CrossRef]
- Tanaka, M.; Machida, Y.; Niu, S.; Ikeda, T.; Jana, N.R.; Doi, H.; Kurosawa, M.; Nekooki, M.; Nukina, N. Trehalose Alleviates Polyglutamine-Mediated Pathology in a Mouse Model of Huntington Disease. Nat. Med. 2004, 10, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Barkhordarian, H.; Emadi, S.; Park, C.B.; Sierks, M.R. Trehalose Differentially Inhibits Aggregation and Neurotoxicity of Beta-Amyloid 40 and 42. Neurobiol. Dis. 2005, 20, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.-B.; Jiang, T.; Lan, D.-M.; Lu, J.-H.; Yue, Z.-Y.; Wang, J.; Zhou, P. Trehalose Inhibits Fibrillation of A53T Mutant Alpha-Synuclein and Disaggregates Existing Fibrils. Arch. Biochem. Biophys. 2012, 523, 144–150. [Google Scholar] [CrossRef]
- Casarejos, M.J.; Solano, R.M.; Gómez, A.; Perucho, J.; de Yébenes, J.G.; Mena, M.A. The Accumulation of Neurotoxic Proteins, Induced by Proteasome Inhibition, Is Reverted by Trehalose, an Enhancer of Autophagy, in Human Neuroblastoma Cells. Neurochem. Int. 2011, 58, 512–520. [Google Scholar] [CrossRef]
- Rusmini, P.; Cortese, K.; Crippa, V.; Cristofani, R.; Cicardi, M.E.; Ferrari, V.; Vezzoli, G.; Tedesco, B.; Meroni, M.; Messi, E.; et al. Trehalose Induces Autophagy via Lysosomal-Mediated TFEB Activation in Models of Motoneuron Degeneration. Autophagy 2019, 15, 631–651. [Google Scholar] [CrossRef]
- Maruf, A.; Milewska, M.; Varga, M.; Wandzik, I. Trehalose-Bearing Carriers to Target Impaired Autophagy and Protein Aggregation Diseases. J. Med. Chem. 2023, 66, 15613–15628. [Google Scholar] [CrossRef]
- Palmieri, M.; Pal, R.; Nelvagal, H.R.; Lotfi, P.; Stinnett, G.R.; Seymour, M.L.; Chaudhury, A.; Bajaj, L.; Bondar, V.V.; Bremner, L.; et al. MTORC1-Independent TFEB Activation via Akt Inhibition Promotes Cellular Clearance in Neurodegenerative Storage Diseases. Nat. Commun. 2017, 8, 14338. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Yoon, Y.-S.; Lee, S.-J. Mechanism of Neuroprotection by Trehalose: Controversy Surrounding Autophagy Induction. Cell Death Dis. 2018, 9, 712. [Google Scholar] [CrossRef]
- Khalifeh, M.; Barreto, G.; Sahebkar, A. Therapeutic Potential of Trehalose in Neurodegenerative Diseases: The Knowns and Unknowns. Neural Regen. Res. 2021, 16, 2026. [Google Scholar] [CrossRef]
- Chen, A.; Gibney, P.A. Dietary Trehalose as a Bioactive Nutrient. Nutrients 2023, 15, 1393. [Google Scholar] [CrossRef] [PubMed]
- Hui Yap, K.; Azmin, S.; Makpol, S.; Damanhuri, H.; Mustapha, M.; Hamzah, J.; Ibrahim, N. Profiling Neuroprotective Potential of Trehalose in Animal Models of Neurodegenerative Diseases: A Systematic Review. Neural Regen. Res. 2023, 18, 1179. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.M.; Moura, I.B.; Arai, N.; Spittal, W.; Clark, E.; Nishida, Y.; Harris, H.C.; Bentley, K.; Davis, G.; Wang, D.; et al. Trehalose-Induced Remodelling of the Human Microbiota Affects Clostridioides difficile Infection Outcome in an In Vitro Colonic Model: A Pilot Study. Front. Cell. Infect. Microbiol. 2021, 11, 670935. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L.; Mazmanian, S.K. The Gut Microbiota–Brain Axis in Behaviour and Brain Disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Loh, J.S.; Mak, W.Q.; Tan, L.K.S.; Ng, C.X.; Chan, H.H.; Yeow, S.H.; Foo, J.B.; Ong, Y.S.; How, C.W.; Khaw, K.Y. Microbiota–Gut–Brain Axis and Its Therapeutic Applications in Neurodegenerative Diseases. Signal Transduct. Target. Ther. 2024, 9, 37. [Google Scholar]
- Hosseinpour-Moghaddam, K.; Caraglia, M.; Sahebkar, A. Autophagy Induction by Trehalose: Molecular Mechanisms and Therapeutic Impacts. J. Cell Physiol. 2018, 233, 6524–6543. [Google Scholar] [CrossRef]
- Del Bello, B.; Gamberucci, A.; Marcolongo, P.; Maellaro, E. The Autophagy Inducer Trehalose Stimulates Macropinocytosis in NF1-Deficient Glioblastoma Cells. Cancer Cell Int. 2022, 22, 232. [Google Scholar] [CrossRef] [PubMed]
- DeBosch, B.J.; Heitmeier, M.R.; Mayer, A.L.; Higgins, C.B.; Crowley, J.R.; Kraft, T.E.; Chi, M.; Newberry, E.P.; Chen, Z.; Finck, B.N.; et al. Trehalose Inhibits Solute Carrier 2A (SLC2A) Proteins to Induce Autophagy and Prevent Hepatic Steatosis. Sci. Signal 2016, 9, ra21. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.J.; Stitham, J.; Evans, T.D.; Zhang, X.; Rodriguez-Velez, A.; Yeh, Y.S.; Tao, J.; Takabatake, K.; Epelman, S.; Lodhi, I.J.; et al. Trehalose Causes Low-Grade Lysosomal Stress to Activate TFEB and the Autophagy-Lysosome Biogenesis Response. Autophagy 2021, 17, 3740–3752. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ren, J. MTOR-Independent Autophagy Inducer Trehalose Rescues against Insulin Resistance-Induced Myocardial Contractile Anomalies: Role of P38 MAPK and Foxo1. Pharmacol. Res. 2016, 111, 357–373. [Google Scholar] [CrossRef] [PubMed]
- Belzile, J.-P.; Sabalza, M.; Craig, M.; Clark, E.; Morello, C.S.; Spector, D.H. Trehalose, an MTOR-Independent Inducer of Autophagy, Inhibits Human Cytomegalovirus Infection in Multiple Cell Types. J. Virol. 2016, 90, 1259–1277. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.L.; Grose, C. Variable Effects of Autophagy Induction by Trehalose on Herpesviruses Depending on Conditions of Infection. Yale J. Biol. Med. 2017, 90, 25–33. [Google Scholar] [PubMed]
- Rawat, P.; Hon, S.; Teodorof-Diedrich, C.; Spector, S.A. Trehalose Inhibits Human Immunodeficiency Virus Type 1 Infection in Primary Human Macrophages and CD4 + T Lymphocytes through Two Distinct Mechanisms. J. Virol. 2020, 94, e00237-20. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Yaribeygi, A.; Sathyapalan, T.; Sahebkar, A. Molecular Mechanisms of Trehalose in Modulating Glucose Homeostasis in Diabetes. Diabetes Metab. Syndr. 2019, 13, 2214–2218. [Google Scholar] [CrossRef] [PubMed]
- Arai, C.; Arai, N.; Mizote, A.; Kohno, K.; Iwaki, K.; Hanaya, T.; Arai, S.; Ushio, S.; Fukuda, S. Trehalose Prevents Adipocyte Hypertrophy and Mitigates Insulin Resistance. Nutr. Res. 2010, 30, 840–848. [Google Scholar] [CrossRef]
- Arai, C.; Miyake, M.; Matsumoto, Y.; Mizote, A.; Yoshizane, C.; Hanaya, Y.; Koide, K.; Yamada, M.; Hanaya, T.; Arai, S.; et al. Trehalose Prevents Adipocyte Hypertrophy and Mitigates Insulin Resistance in Mice with Established Obesity. J. Nutr. Sci. Vitaminol. 2013, 59, 393–401. [Google Scholar] [CrossRef]
- Eckstein, S.S.; Weigert, C.; Lehmann, R. Divergent Roles of IRS (Insulin Receptor Substrate) 1 and 2 in Liver and Skeletal Muscle. Curr. Med. Chem. 2017, 24, 1827–1852. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, Regulation, and Involvement in Disease. Int. Immunopharmacol. 2021, 101 Pt B, 107598. [Google Scholar] [CrossRef]
- Higgins, C.B.; Zhang, Y.; Mayer, A.L.; Fujiwara, H.; Stothard, A.I.; Graham, M.J.; Swarts, B.M.; DeBosch, B.J. Hepatocyte ALOXE3 Is Induced during Adaptive Fasting and Enhances Insulin Sensitivity by Activating Hepatic PPARγ. JCI Insight 2018, 3, e120794. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.J. Trehalose Synthesis and Metabolism Are Required at Different Stages of Plant Infection by Magnaporthe grisea. EMBO J. 2003, 22, 225–235. [Google Scholar] [CrossRef]
- Zhu, X.; Fang, D.; Li, D.; Zhang, J.; Jiang, H.; Guo, L.; He, Q.; Zhang, T.; Macho, A.P.; Wang, E.; et al. Phytophthora Sojae Boosts Host Trehalose Accumulation to Acquire Carbon and Initiate Infection. Nat. Microbiol. 2023, 8, 1561–1573. [Google Scholar] [CrossRef] [PubMed]
- Al-Bader, N.; Vanier, G.; Liu, H.; Gravelat, F.N.; Urb, M.; Hoareau, C.M.-Q.; Campoli, P.; Chabot, J.; Filler, S.G.; Sheppard, D.C. Role of Trehalose Biosynthesis in Aspergillus fumigatus Development, Stress Response, and Virulence. Infect. Immun. 2010, 78, 3007–3018. [Google Scholar] [CrossRef] [PubMed]
- Puttikamonkul, S.; Willger, S.D.; Grahl, N.; Perfect, J.R.; Movahed, N.; Bothner, B.; Park, S.; Paderu, P.; Perlin, D.S.; Cramer, R.A., Jr. Trehalose 6-phosphate Phosphatase Is Required for Cell Wall Integrity and Fungal Virulence but Not Trehalose Biosynthesis in the Human Fungal Pathogen Aspergillus fumigatus. Mol. Microbiol. 2010, 77, 891–911. [Google Scholar] [CrossRef] [PubMed]
- Doehlemann, G.; Berndt, P.; Hahn, M. Trehalose Metabolism Is Important for Heat Stress Tolerance and Spore Germination of Botrytis cinerea. Microbiology 2006, 152, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Esparza, M.; Aguinaga, A.; González-Párraga, P.; García-Peñarrubia, P.; Jouault, T.; Argüelles, J.C. Role of Trehalose in Resistance to Macrophage Killing: Study with a Tps1/Tps1 Trehalose-Deficient Mutant of Candida albicans. Clin. Microbiol. Inf. 2007, 13, 384–394. [Google Scholar] [CrossRef]
- Martínez-Esparza, M.; Martínez-Vicente, E.; González-Párraga, P.; Ros, J.M.; García-Peñarrubia, P.; Argüelles, J.-C. Role of Trehalose-6P Phosphatase (TPS2) in Stress Tolerance and Resistance to Macrophage Killing in Candida albicans. Int. J. Med. Microbiol. 2009, 299, 453–464. [Google Scholar] [CrossRef]
- Petzold, E.W.; Himmelreich, U.; Mylonakis, E.; Rude, T.; Toffaletti, D.; Cox, G.M.; Miller, J.L.; Perfect, J.R. Characterization and Regulation of the Trehalose Synthesis Pathway and Its Importance in the Pathogenicity of Cryptococcus neoformans. Infect. Immun. 2006, 74, 5877–5887. [Google Scholar] [CrossRef] [PubMed]
- Song, X.-S.; Li, H.-P.; Zhang, J.-B.; Song, B.; Huang, T.; Du, X.-M.; Gong, A.-D.; Liu, Y.-K.; Feng, Y.-N.; Agboola, R.S.; et al. Trehalose 6-Phosphate Phosphatase Is Required for Development, Virulence and Mycotoxin Biosynthesis Apart from Trehalose Biosynthesis in Fusarium graminearum. Fungal Genet. Biol. 2014, 63, 24–41. [Google Scholar] [CrossRef]
- Chen, L.; Ma, X.; Sun, T.; Zhu, Q.-H.; Feng, H.; Li, Y.; Liu, F.; Zhang, X.; Sun, J.; Li, Y. VdPT1 Encoding a Neutral Trehalase of Verticillium dahliae Is Required for Growth and Virulence of the Pathogen. Int. J. Mol. Sci. 2023, 25, 294. [Google Scholar] [CrossRef] [PubMed]
- Vanaporn, M.; Sarkar-Tyson, M.; Kovacs-Simon, A.; Ireland, P.M.; Pumirat, P.; Korbsrisate, S.; Titball, R.W.; Butt, A. Trehalase Plays a Role in Macrophage Colonization and Virulence of Burkholderia pseudomallei in Insect and Mammalian Hosts. Virulence 2017, 8, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Pavanelo, D.B.; Houle, S.; Matter, L.B.; Dozois, C.M.; Horn, F. The Periplasmic Trehalase Affects Type 1 Fimbria Production and Virulence of Extraintestinal Pathogenic Escherichia coli Strain MT78. Infect. Immun. 2018, 86, e00241-18. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-C.; Lin, T.-L.; Hsieh, P.-F.; Yang, H.-C.; Wang, J.-T. Isolation of Genes Involved in Biofilm Formation of a Klebsiella pneumoniae Strain Causing Pyogenic Liver Abscess. PLoS ONE 2011, 6, e23500. [Google Scholar] [CrossRef] [PubMed]
- Ells, T.C.; Truelstrup Hansen, L. Increased Thermal and Osmotic Stress Resistance in Listeria monocytogenes 568 Grown in the Presence of Trehalose Due to Inactivation of the Phosphotrehalase-Encoding Gene TreA. Appl. Environ. Microbiol. 2011, 77, 6841–6851. [Google Scholar] [CrossRef] [PubMed]
- Murphy, H.N.; Stewart, G.R.; Mischenko, V.V.; Apt, A.S.; Harris, R.; McAlister, M.S.B.; Driscoll, P.C.; Young, D.B.; Robertson, B.D. The OtsAB Pathway Is Essential for Trehalose Biosynthesis in Mycobacterium Tuberculosis. J. Biol. Chem. 2005, 280, 14524–14529. [Google Scholar] [CrossRef]
- Djonović, S.; Urbach, J.M.; Drenkard, E.; Bush, J.; Feinbaum, R.; Ausubel, J.L.; Traficante, D.; Risech, M.; Kocks, C.; Fischbach, M.A.; et al. Trehalose Biosynthesis Promotes Pseudomonas aeruginosa Pathogenicity in Plants. PLoS Pathog. 2013, 9, e1003217. [Google Scholar] [CrossRef]
- Hamilton, C.D.; Steidl, O.R.; MacIntyre, A.M.; Hendrich, C.G.; Allen, C. Ralstonia Solanacearum Depends on Catabolism of Myo-Inositol, Sucrose, and Trehalose for Virulence in an Infection Stage–Dependent Manner. Mol. Plant-Microbe Interact. 2021, 34, 669–679. [Google Scholar] [CrossRef]
- McIntyre, H.J.; Davies, H.; Hore, T.A.; Miller, S.H.; Dufour, J.-P.; Ronson, C.W. Trehalose Biosynthesis in Rhizobium Leguminosarum Bv. Trifolii and Its Role in Desiccation Tolerance. Appl. Environ. Microbiol. 2007, 73, 3984–3992. [Google Scholar] [CrossRef] [PubMed]
- Howells, A.M.; Bullifent, H.L.; Dhaliwal, K.; Griffin, K.; García de Castro, A.; Frith, G.; Tunnacliffe, A.; Titball, R.W. Role of Trehalose Biosynthesis in Environmental Survival and Virulence of Salmonella enterica Serovar Typhimurium. Res. Microbiol. 2002, 153, 281–287. [Google Scholar] [CrossRef]
- Cánovas, D.; Fletcher, S.A.; Hayashi, M.; Csonka, L.N. Role of Trehalose in Growth at High Temperature of Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2001, 183, 3365–3371. [Google Scholar] [CrossRef] [PubMed]
- Ledermann, R.; Emmenegger, B.; Couzigou, J.-M.; Zamboni, N.; Kiefer, P.; Vorholt, J.A.; Fischer, H.-M. Bradyrhizobium diazoefficiens Requires Chemical Chaperones To Cope with Osmotic Stress during Soybean Infection. mBio 2021, 12, e00390-21. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.C.; Chen, C.; Beattie, G.A. Identification of the Trehalose Biosynthetic Loci of Pseudomonas syringae and Their Contribution to Fitness in the Phyllosphere. Environ. Microbiol. 2010, 12, 1486–1497. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jeong, B.; Bae, H.R.; Jang, H.A.; Kim, J.K. Trehalose Biosynthesis Gene OtsA Protects against Stress in the Initial Infection Stage of Burkholderia -Bean Bug Symbiosis. Microbiol. Spectr. 2023, 11, e0351022. [Google Scholar] [CrossRef]
- Piazza, A.; Zimaro, T.; Garavaglia, B.S.; Ficarra, F.A.; Thomas, L.; Marondedze, C.; Feil, R.; Lunn, J.E.; Gehring, C.; Ottado, J.; et al. The Dual Nature of Trehalose in Citrus Canker Disease: A Virulence Factor for Xanthomonas citri Subsp. Citri and a Trigger for Plant Defence Responses. J. Exp. Bot. 2015, 66, 2795–2811. [Google Scholar] [CrossRef]
- MacIntyre, A.M.; Barth, J.X.; Pellitteri Hahn, M.C.; Scarlett, C.O.; Genin, S.; Allen, C. Trehalose Synthesis Contributes to Osmotic Stress Tolerance and Virulence of the Bacterial Wilt Pathogen Ralstonia solanacearum. Mol. Plant Microbe 2020, 33, 462–473. [Google Scholar] [CrossRef]
- Poueymiro, M.; Cazalé, A.C.; François, J.M.; Parrou, J.L.; Peeters, N.; Genin, S. A Ralstonia Solanacearum Type III Effector Directs the Production of the Plant Signal Metabolite Trehalose-6-Phosphate. mBio 2014, 5, e02065-14. [Google Scholar] [CrossRef]
- Sachdeva, K.; Goel, M.; Sudhakar, M.; Mehta, M.; Raju, R.; Raman, K.; Singh, A.; Sundaramurthy, V. Mycobacterium tuberculosis (Mtb) Lipid Mediated Lysosomal Rewiring in Infected Macrophages Modulates Intracellular Mtb Trafficking and Survival. J. Biol. Chem. 2020, 295, 9192–9210. [Google Scholar] [CrossRef]
- Kalscheuer, R.; Koliwer-Brandl, H. Genetics of Mycobacterial Trehalose Metabolism. Microbiol. Spectr. 2014, 2, MGM2-0002-2013. [Google Scholar] [CrossRef] [PubMed]
- Harland, C.W.; Rabuka, D.; Bertozzi, C.R.; Parthasarathy, R. The Mycobacterium Tuberculosis Virulence Factor Trehalose Dimycolate Imparts Desiccation Resistance to Model Mycobacterial Membranes. Biophys. J. 2008, 94, 4718–4724. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Fresneda, R.; Martínez-Esparza, M.; Maicas, S.; Argüelles, J.-C.; Valentín, E. In Candida parapsilosis the ATC1 Gene Encodes for an Acid Trehalase Involved in Trehalose Hydrolysis, Stress Resistance and Virulence. PLoS ONE 2014, 9, e99113. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Fresneda, R.; Muñoz-Megías, M.L.; Yagüe, G.; Solano, F.; Maicas, S.; Argüelles, J.C. Lack of Functional Trehalase Activity in Candida parapsilosis Increases Susceptibility to Itraconazole. J. Fungi 2022, 8, 371. [Google Scholar] [CrossRef] [PubMed]
- Gollan, B.; Grabe, G.; Michaux, C.; Helaine, S. Bacterial Persisters and Infection: Past, Present, and Progressing. Annu. Rev. Microbiol. 2019, 73, 359–385. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and Guidelines for Research on Antibiotic Persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister Cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.-H.; Ryu, C.-M.; Kim, J.-S. Bacterial Persistence: Fundamentals and Clinical Importance. J. Microbiol. 2019, 57, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Bollen, C.; Dewachter, L.; Michiels, J. Protein Aggregation as a Bacterial Strategy to Survive Antibiotic Treatment. Front. Mol. Biosci. 2021, 8, 669664. [Google Scholar] [CrossRef]
- Vega, N.M.; Allison, K.R.; Khalil, A.S.; Collins, J.J. Signaling-Mediated Bacterial Persister Formation. Nat. Chem. Biol. 2012, 8, 431–433. [Google Scholar] [CrossRef]
- Hu, Y.; Kwan, B.W.; Osbourne, D.O.; Benedik, M.J.; Wood, T.K. Toxin YafQ Increases Persister Cell Formation by Reducing Indole Signalling. Environ. Microbiol. 2015, 17, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tian, T.; Zhang, J.; Jin, X.; Yue, H.; Zhang, X.-H.; Du, L.; Bai, F. Indole Reverses Intrinsic Antibiotic Resistance by Activating a Novel Dual-Function Importer. mBio 2019, 10, e00676-19. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Lee, S.-K.; Song, N.; Nathan, T.O.; Swarts, B.M.; Eum, S.-Y.; Ehrt, S.; Cho, S.-N.; Eoh, H. Transient Drug-Tolerance and Permanent Drug-Resistance Rely on the Trehalose-Catalytic Shift in Mycobacterium tuberculosis. Nat. Commun. 2019, 10, 2928. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, S.; Wang, Y.J. Trehalose: Current Use and Future Applications. J. Pharm. Sci. 2011, 100, 2020–2053. [Google Scholar] [CrossRef] [PubMed]
- Vinciguerra, D.; Gelb, M.B.; Maynard, H.D. Synthesis and Application of Trehalose Materials. JACS Au 2022, 2, 1561–1587. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Tapia, H.; Goddard, J.M.; Gibney, P.A. Trehalose and Its Applications in the Food Industry. Compr. Rev. Food Sci. Food Saf. 2022, 21, 5004–5037. [Google Scholar] [CrossRef] [PubMed]
- Crowley, C.A.; Smith, W.P.W.; Seah, K.T.M.; Lim, S.-K.; Khan, W.S. Cryopreservation of Human Adipose Tissues and Adipose-Derived Stem Cells with DMSO and/or Trehalose: A Systematic Review. Cells 2021, 10, 1837. [Google Scholar] [CrossRef] [PubMed]
- Cagini, C.; Di Lascio, G.; Torroni, G.; Mariniello, M.; Meschini, G.; Lupidi, M.; Messina, M. Dry Eye and Inflammation of the Ocular Surface after Cataract Surgery: Effectiveness of a Tear Film Substitute Based on Trehalose/Hyaluronic Acid vs Hyaluronic Acid to Resolve Signs and Symptoms. J. Cataract. Refract. Surg. 2021, 47, 1430–1435. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Sánchez, A.; Martinez-Perez, C.; Alvarez-Peregrina, C.; Sánchez-Tena, M.Á.; De-Hita-Cantalejo, C.; Sánchez-González, M.C.; Sánchez-González, J.-M. Trehalose and Dry Eye Disease: A Comprehensive Systematic Review of Randomized Controlled Trials. J. Clin. Med. 2023, 12, 7301. [Google Scholar] [CrossRef]
- Laihia, J.; Kaarniranta, K. Trehalose for Ocular Surface Health. Biomolecules 2020, 10, 809. [Google Scholar] [CrossRef]
- Morales-Carrizales, D.A.; Gopar-Cuevas, Y.; Loera-Arias, M.d.J.; Saucedo-Cardenas, O.; Montes de Oca-Luna, R.; Garcia-Garcia, A.; Rodriguez-Rocha, H. A Neuroprotective Dose of Trehalose Is Harmless to Metabolic Organs: Comprehensive Histopathological Analysis of Liver, Pancreas, and Kidney. DARU J. Pharm. Sci. 2023, 31, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Honda, Y.; Tanaka, M.; Honda, S. Trehalose Extends Longevity in the Nematode Caenorhabditis elegans. Aging Cell 2010, 9, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.; Marconi, M.; Musillo, C.; Chiarotti, F.; Bellisario, V.; Matarrese, P.; Gambardella, L.; Vona, R.; Lombardi, M.; Foglieni, C.; et al. Trehalose Administration in C57BL/6N Old Mice Affects Healthspan Improving Motor Learning and Brain Anti-Oxidant Defences in a Sex-Dependent Fashion: A Pilot Study. Exp. Gerontol. 2020, 129, 110755. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Kingsley, S.; Walker, G.; Mondoux, M.A.; Tissenbaum, H.A. Metabolic Shift from Glycogen to Trehalose Promotes Lifespan and Healthspan in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2018, 115, E2791–E2800. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R.; Tenor, J.L.; Miao, Y.; Brennan, R.G. Trehalose Pathway as an Antifungal Target. Virulence 2017, 8, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Guirao-Abad, J.P. Analysis of Validamycin as a Potential Antifungal Compound against Candida albicans. Int. Microbiol. 2013, 16, 217–225. [Google Scholar] [PubMed]
- Danielson, N.D.; Collins, J.; Stothard, A.I.; Dong, Q.Q.; Kalera, K.; Woodruff, P.J.; DeBosch, B.J.; Britton, R.A.; Swarts, B.M. Degradation-Resistant Trehalose Analogues Block Utilization of Trehalose by Hypervirulent Clostridioides difficile. Chem. Comm. 2019, 55, 5009–5012. [Google Scholar] [CrossRef]
- Wolber, J.M.; Urbanek, B.L.; Meints, L.M.; Piligian, B.F.; Lopez-Casillas, I.C.; Zochowski, K.M.; Woodruff, P.J.; Swarts, B.M. The Trehalose-Specific Transporter LpqY-SugABC Is Required for Antimicrobial and Anti-Biofilm Activity of Trehalose Analogues in Mycobacterium smegmatis. Carbohydr. Res. 2017, 450, 60–66. [Google Scholar] [CrossRef]
- Govind, S.R.; Jogaiah, S.; Abdelrahman, M.; Shetty, H.S.; Tran, L.-S.P. Exogenous Trehalose Treatment Enhances the Activities of Defense-Related Enzymes and Triggers Resistance against Downy Mildew Disease of Pearl Millet. Front. Plant Sci. 2016, 7, 1593. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Species | Mutation | Virulence | Survival under Abiotic Stress Conditions | Ref. |
|---|---|---|---|---|
| Fungi | ||||
| Aspergillus fumigatus | tpsA tpsB (trehalose-6-phosphate synthase genes) | Hypervirulent in a murine model of invasive aspergillosis | Delayed germination at 37 °C, reduced thermotolerance at 50 °C and increased susceptibility to oxidative shock | [86] |
| orlA (trehalose-6P phosphatase) | Virulence attenuation in murine models of invasive pulmonary aspergillosis, increased sensitivity to cell wall perturbing agents (Calcofluor white, Congo Red or nikkomycin Z) | [87] | ||
| Botrytis cinerea | tps1 (trehalose-6-phosphate synthase) | Similar to the wild-type strain | Normal vegetative growth; growth inhibition at higher temperature | [88] |
| tre1 (neutral trehalase) | Delayed germination in the presence of sugars at low concentrations | Increased heat resistance | ||
| Candida albicans | tps1 | Lower infection rate in mice; more sensitive to macrophage killing | [89] | |
| tps2 (trehalose-6P phosphatase) | Reduces virulence; more susceptible to murine and human macrophages phagocytosis | Reduced growth at high temperatures, sensitivity to heat shock (42 °C) and oxidative exposure (50 mM H2O2) | [90] | |
| Cryptococcus neoformans | tps1 tps2 | Avirulent in rabbits and mice | Temperature sensitivity at 37 °C in the presence of glucose | [91] |
| nth1 (neutral trehalase) | Similar to the wild-type strain | |||
| Fusarium graminearum | TPS2 | Reduction in mycotoxin production; 99% lower virulence on wheat | [92] | |
| Magnaporthe grisea | TPS | Reduced sporulation and virulence | [84] | |
| NTH1 | Decreased ability to colonize plant tissue | |||
| Verticillium dahliae | Vdpt1 (neutral trehalase) | Reduced colony growth rate, delayed mycelial growth and conidial germination and reduced pathogenicity | Increased resistance to high temperature, NaCl, sorbitol, and validamycin A | [93] |
| Bacteria | ||||
| Burkholderia pseudomallei | treA (trehalase) | Decreased biofilm formation; reduced virulence in G. mellonella and in mice; reduced growth in murine macrophages | Increased tolerance to thermal stress | [94] |
| Escherichia coli (ExPEC) strain MT78 | treA | Reduces cell invasion and colonization of the bladder in a murine urinary tract infection model; reduced production of type 1 fimbriae | Increased resistance to 0.6 M urea | [95] |
| Escherichia coli | otsA (trehalose-6-phosphate synthase) | Increased formation of antibiotic (Amp and Ofx) tolerance cells after heat shock and in stationary cultures | Reduced ability to survive at 4 °C Increased oxidative stress damage | [14,40] |
| otsA otsB double mutation | Decreased stationary phase thermotolerance at 55 °C | [15] | ||
| Klebsiella pneumoniae NTUH-K2044 | treC (trehalose-6-phosphate hydrolase) | Decreased capsule production; decreased biofilm formation; attenuated ability to colonize the gastrointestinal tract in a murine model | [96] | |
| Listeria monocytogenes | treA (phosphotrehalase) | Enhanced resistance to heat, high osmolarity, desiccation, and freeze–thaw cycling stresses. | [97] | |
| Mycobacterium tuberculosis | otsB2 (trehalose-6-phosphate phosphatase) | Lethal | [98] | |
| otsA | Growth defect during in vitro culture and murine infection | |||
| treS (trehalose synthase) | Reduced growth in the mouse model | |||
| Pseudomonas aeruginosa strain PA14 | treYZ (maltooligosyl trehalose synthase and maltooligosyl trehalose trehalohydrolase | Severe attenuation in virulence in Arabidopsis leaves | [99] | |
| treS | Attenuation in virulence in Arabidopsis leaves | |||
| double ΔtreYZΔtreS mutant | Severe attenuation in virulence in Arabidopsis leaves; full virulence in nematodes, insects, or mice | |||
| Ralstonia solanacearum | treA | Defective in tomato colonization, reduced virulence | [100] | |
| Rhizobium leguminosarum bv. trifolii | otsA treY | Less competitive for nodule occupancy than the wild-type strain | Decreased resistance to drying and subsequent storage at 25 °C | [101] |
| Salmonella enterica serovar Typhimurium | otsA | The ability to colonize spleen tissues in mice similar to the wild-type strain | Growth defect at 45 °C in minimal medium containing 0.2 M NaCl | [102,103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuczyńska-Wiśnik, D.; Stojowska-Swędrzyńska, K.; Laskowska, E. Intracellular Protective Functions and Therapeutical Potential of Trehalose. Molecules 2024, 29, 2088. https://doi.org/10.3390/molecules29092088
Kuczyńska-Wiśnik D, Stojowska-Swędrzyńska K, Laskowska E. Intracellular Protective Functions and Therapeutical Potential of Trehalose. Molecules. 2024; 29(9):2088. https://doi.org/10.3390/molecules29092088
Chicago/Turabian StyleKuczyńska-Wiśnik, Dorota, Karolina Stojowska-Swędrzyńska, and Ewa Laskowska. 2024. "Intracellular Protective Functions and Therapeutical Potential of Trehalose" Molecules 29, no. 9: 2088. https://doi.org/10.3390/molecules29092088
APA StyleKuczyńska-Wiśnik, D., Stojowska-Swędrzyńska, K., & Laskowska, E. (2024). Intracellular Protective Functions and Therapeutical Potential of Trehalose. Molecules, 29(9), 2088. https://doi.org/10.3390/molecules29092088