Author Contributions
Conceptualization, B.E.O., B.O.A. and O.I.O.; Methodology, E.A.A. and O.T.I.; Validation, R.T.A. and T.N.P.D.; Investigation, B.E.O., D.E.S., S.A.O., C.S.U., T.N.P.D. and D.T.E.; Data curation, O.T.I.; Writing—original draft, M.S.M.-G.; Writing—review and editing, D.E.S. and E.A.A.; Supervision, O.V.O.; Project administration, R.T.A. All authors have read and agreed to the published version of the manuscript.
Figure 1.
The three reference ligands, Oxamniquine, Praziquantel, and Artemisinin, and identified ligands (Andrographolide, Nicotiflorin, and Isoquercitrin).
Figure 1.
The three reference ligands, Oxamniquine, Praziquantel, and Artemisinin, and identified ligands (Andrographolide, Nicotiflorin, and Isoquercitrin).
Figure 2.
The superimposed structures of the co-crystalised ligands in their co-crystallised (magenta) and re-docked poses (green) [RMSD = 0.73 and 0.87 A for 2X8H and 6UY4, respectively].
Figure 2.
The superimposed structures of the co-crystalised ligands in their co-crystallised (magenta) and re-docked poses (green) [RMSD = 0.73 and 0.87 A for 2X8H and 6UY4, respectively].
Figure 3.
The 3D (top) and 2D (bottom) ligand interactions of the Artemisinin–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 3.
The 3D (top) and 2D (bottom) ligand interactions of the Artemisinin–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
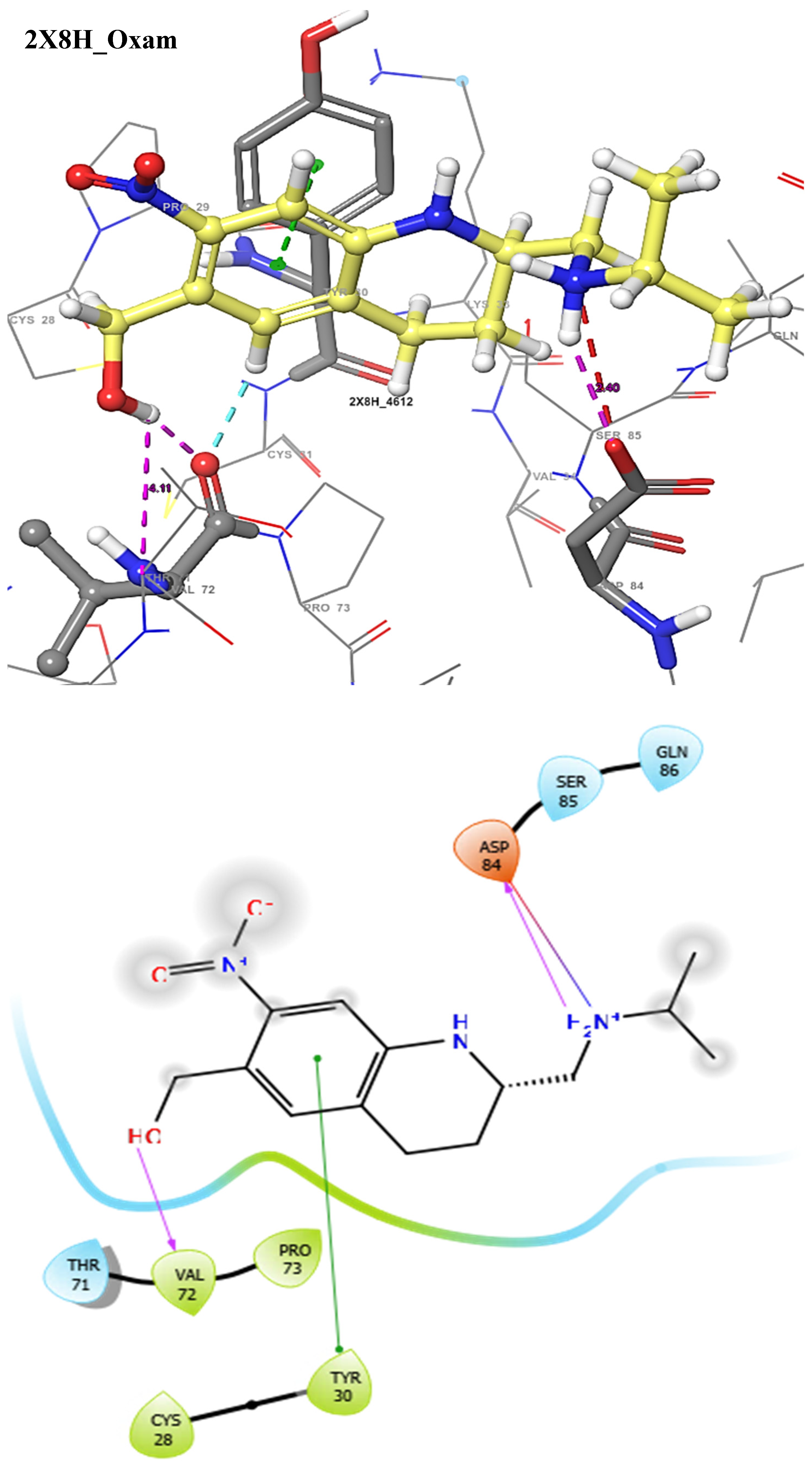
Figure 4.
The 3D (top) and 2D (bottom) ligand interactions of the Oxamniquine–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 4.
The 3D (top) and 2D (bottom) ligand interactions of the Oxamniquine–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 5.
The 3D (top) and 2D (bottom) ligand interactions of the Praziquantel–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 5.
The 3D (top) and 2D (bottom) ligand interactions of the Praziquantel–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 6.
The 3D (top) and 2D (bottom) ligand interactions of the Andrographolide–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 6.
The 3D (top) and 2D (bottom) ligand interactions of the Andrographolide–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 7.
The 3D (top) and 2D (bottom) ligand interactions of the Nicotiflorin–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 7.
The 3D (top) and 2D (bottom) ligand interactions of the Nicotiflorin–thioredoxin glutathione reductase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 8.
The 3D (top) and 2D (bottom) ligand interactions of the Oxamniquine–Schistosoma mansoni Arginase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 8.
The 3D (top) and 2D (bottom) ligand interactions of the Oxamniquine–Schistosoma mansoni Arginase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 9.
The 3D (top) and 2D (bottom) ligand interactions of the Andrographolide–Schistosoma mansoni Arginase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 9.
The 3D (top) and 2D (bottom) ligand interactions of the Andrographolide–Schistosoma mansoni Arginase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 10.
The 3D (top) and 2D (bottom) ligand interactions of the Andrographolide–Schistosoma mansoni dihydroorotate dehydrogenase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 10.
The 3D (top) and 2D (bottom) ligand interactions of the Andrographolide–Schistosoma mansoni dihydroorotate dehydrogenase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 11.
The 3D (top) and 2D (bottom) ligand interactions of the Artemisinin–Schistosoma mansoni dihydroorotate dehydrogenase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 11.
The 3D (top) and 2D (bottom) ligand interactions of the Artemisinin–Schistosoma mansoni dihydroorotate dehydrogenase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 12.
The 3D (top) and 2D (bottom) ligand interactions of the Isoquercitrin–Schistosoma mansoni dihydroorotate dehydrogenase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 12.
The 3D (top) and 2D (bottom) ligand interactions of the Isoquercitrin–Schistosoma mansoni dihydroorotate dehydrogenase complex. The binding site residues’ charges are indicated in the 2D image by the colours red for negative, blue for positive, and white for neutral.
Figure 13.
MD simulation of 4Q3T, 6UY4, and 2X8H complexed to 5318517: Andrographolide, 5318767: Nicotiflorin, and 5280804: Isoquercitrin. (A) RMSF and RMSD graphical illustration plot. (B) rGyr representation and SASA diagram. All simulations were carried out using Schrödinger’s Maestro suite.
Figure 13.
MD simulation of 4Q3T, 6UY4, and 2X8H complexed to 5318517: Andrographolide, 5318767: Nicotiflorin, and 5280804: Isoquercitrin. (A) RMSF and RMSD graphical illustration plot. (B) rGyr representation and SASA diagram. All simulations were carried out using Schrödinger’s Maestro suite.
Table 1.
Hit compounds from Azadirachta indica with their physicochemical properties.
Table 1.
Hit compounds from Azadirachta indica with their physicochemical properties.
| Ligand | MW | #HA | #AHA | #RB | #HBA | #HBD |
|---|
| 4612 | 279.33 | 20 | 6 | 4 | 5 | 3 |
| 4891 | 312.41 | 23 | 6 | 2 | 2 | 0 |
| 68827 | 282.33 | 20 | 0 | 0 | 5 | 0 |
| 5318517 | 350.45 | 25 | 0 | 3 | 5 | 3 |
| 5318767 | 594.52 | 42 | 16 | 6 | 15 | 9 |
| 5280804 | 464.38 | 33 | 16 | 4 | 12 | 8 |
Table 2.
Medicinal chemistry profile and drug-likeness of reference drugs and hit compounds from Azadirachta indica.
Table 2.
Medicinal chemistry profile and drug-likeness of reference drugs and hit compounds from Azadirachta indica.
| Ligand | Lipinski #Violations | Ghose #Violations | Veber #Violations | Egan #Violations | TPSA | BS | PAINS #Alerts |
|---|
| 4612 | 0 | 0 | 0 | 0 | 90.11 | 0.55 | 0 |
| 4891 | 0 | 0 | 0 | 0 | 40.62 | 0.55 | 0 |
| 68827 | 0 | 0 | 0 | 0 | 53.99 | 0.55 | 0 |
| 5318517 | 0 | 0 | 0 | 0 | 86.99 | 0.55 | 0 |
| 5318767 | 3 | 4 | 1 | 1 | 249.2 | 0.17 | 0 |
| 5280804 | 2 | 1 | 1 | 1 | 210.15 | 0.17 | 1 |
Table 3.
Absorptivity, distribution, and metabolism properties of identified ligands compared with reference ligands.
Table 3.
Absorptivity, distribution, and metabolism properties of identified ligands compared with reference ligands.
| Ligand | GI Absorption | BBB Permeant | PGP Substrate | CYP1A2 Inhibitor | CYP2C19 Inhibitor | CYP2C9 Inhibitor | CYP2D6 Inhibitor |
|---|
| 4612 | High | No | Yes | No | No | No | No |
| 4891 | High | Yes | Yes | No | Yes | No | Yes |
| 68827 | High | Yes | No | Yes | No | No | No |
| 5318517 | High | No | Yes | No | No | No | No |
| 5318767 | Low | No | Yes | No | No | No | No |
| 5280804 | Low | No | No | No | No | No | No |
Table 4.
Molecular interaction of the docking output of the complexes.
Table 4.
Molecular interaction of the docking output of the complexes.
| Receptor | Ligand | Glide Score (kcal/mol) | MMGBSA (kcal/mol) | H-Bond | Salt Bridge | PI-PI | PI Cation |
|---|
| 2X8H | 68827 | −3.86 | −33.27 | SER85 | TYR30 | - | - |
| 4612 | −3.49 | −31.67 | ASP84, VAL72 | ASP84 | TYR30 | - |
| 4891 | −2.58 | −28.18 | SER85 | - | - | - |
| 5318767 | −5.97 | −46.40 | THR25, GLN60, SER85 | - | - | LYS25 |
| 5318517 | −4.65 | −30.49 | SER85, ASP84, GLN86 | - | - | - |
| 4Q3T | 68827 | −2.87 | −26.16 | - | - | - | - |
| 4891 | −3.41 | −33.61 | SER167 | - | - | - |
| 4612 | −5.97 | −28.78 | THR276, ASN160, ASP158, ASP213, ASP211 | ASP211, ASP213, ASP158 | HIS156 | - |
| 5318767 | −8.67 | −41.44 | ASP211, ASP213,ALA156, SER165, GLU216,SER167, | - | - | - |
| 5318517 | −4.34 | −30.05 | ASP211, SER165, ASN160 | - | - | - |
| 6UY4 | 68827 | −5.37 | −39.33 | - | - | - | - |
| 4891 | −6.80 | −40.64 | SER53 | - | PHE35 | - |
| 4612 | −3.23 | −44.21 | PHE357 | - | - | - |
| 5280804 | −12.80 | −52.05 | HIS50, SER53, ARG130, PHE61, ALA39 | - | - | - |
| 5318517 | −10.19 | −45.75 | PHE61, ARG130, ALA49, SER53 | - | - | - |
Table 5.
Binding energies of known inhibitors for Schistosoma mansoni thioredoxin glutathione reductase (2X8H) retrieved from the CHEMBL database.
Table 5.
Binding energies of known inhibitors for Schistosoma mansoni thioredoxin glutathione reductase (2X8H) retrieved from the CHEMBL database.
| S/N | Compounds | Docking Score (Kcal/mol) |
|---|
| 1. | CHEMBL3322286 | −5.46 |
| 2. | CHEMBL1486739 | −4.26 |
| 3. | CHEMBL3322292 | −4.01 |
| 4. | CHEMBL1455957 | −3.81 |
| 5. | CHEMBL4846043 | −3.73 |
| 6. | CHEMBL1449349 | −3.59 |
| 7. | CHEMBL568961 | −3.46 |
| 8. | CHEMBL4858362 | −3.41 |
| 9. | CHEMBL3322287 | −3.36 |
| 10. | CHEMBL571936 | −3.33 |
| 11. | CHEMBL500868 | −3.30 |
| 12. | CHEMBL567641 | −3.27 |
| 13. | CHEMBL582970 | −3.24 |
| 14. | CHEMBL578810 | −3.22 |
| 15. | CHEMBL1325877 | −3.21 |
| 16. | CHEMBL576118 | −3.14 |
| 17. | CHEMBL574577 | −3.13 |
| 18. | CHEMBL570130 | −3.09 |
| 19. | CHEMBL1428415 | −3.07 |
| 20. | CHEMBL4852477 | −3.06 |
Table 6.
Binding energies of known inhibitors for Schistosoma mansoni dihydroorotate dehydrogenase (6UY4) retrieved from the CHEMBL database.
Table 6.
Binding energies of known inhibitors for Schistosoma mansoni dihydroorotate dehydrogenase (6UY4) retrieved from the CHEMBL database.
| S/N | Compounds | Docking Score (Kcal/mol) |
|---|
| 1. | CHEMBL1450 | −10.35 |
| 2. | CHEMBL4586212 | −9.77 |
| 3. | CHEMBL38434 | −9.51 |
| 4. | CHEMBL38434 | −9.51 |
| 5. | CHEMBL4472078 | −9.33 |
| 6. | CHEMBL4474026 | −9.18 |
| 7. | CHEMBL2023282 | −9.10 |
| 8. | CHEMBL4524841 | −9.05 |
| 9. | CHEMBL4560384 | −8.73 |
| 10. | CHEMBL4540838 | −8.63 |
| 11. | CHEMBL2408379 | −8.58 |
| 12. | CHEMBL4452960 | −8.45 |
| 13. | CHEMBL4457147 | −8.44 |
| 14. | CHEMBL2041119 | −8.38 |
| 15. | CHEMBL4566973 | −8.18 |
| 16. | CHEMBL2408378 | −8.15 |
| 17. | CHEMBL973 | −8.13 |
| 18. | CHEMBL4546952 | −7.99 |
| 19. | CHEMBL1738786 | −7.94 |
| 20. | CHEMBL4527976 | −7.81 |
Table 7.
Interactive properties of MDs of the native receptors and protein–ligand interactions.
Table 7.
Interactive properties of MDs of the native receptors and protein–ligand interactions.
| Receptor | Ligand | RSMF | RSMD | rGyr | MolSA | SASA | PSA |
|---|
| 4Q3T | 5318767 | 2.017 ± 0.70 | 1.189 ± 0.38 | 4.270 ± 0.10 | 430.446 ± 11.42 | 295.193 ± 38.10 | 406.801 ± 11.76 |
| 5318517 | 1.317 ± 0.55 | 1.151 ± 0.42 | 4.022 ± 0.13 | 316.260 ± 2.22 | 300.554 ± 63.60 | 178.618 ± 4.80 |
| 6UY4 | 5318517 | 0.895 ± 0.64 | 1.301 ± 0.36 | 3.728 ± 0.08 | 310.596 ± 240 | 99.332 ± 38.14 | 169.829 ± 4.90 |
| 5280804 | 0.738 ± 0.44 | 0.580 ± 0.09 | 4.180 ± 0.03 | 356.443 ± 3.10 | 26.187 ± 9.24 | 383.859 ± 9.98 |
| 2X8H | 5318767 | 1.365 ± 0.73 | 4.348 ± 0.59 | 4.265 ± 0.18 | 428.864 ± 10.88 | 434.628 ± 42.04 | 414.392 ± 10.01 |
| 5318517 | 1.417 ± 0.84 | 1.137 ± 0.45 | 3.399 ± 0.13 | 315.950 ± 3.01 | 437.568 ± 134.21 | 179.797 ± 5.18 |

 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}