
Isolation of Extracellular Outer Membrane Vesicles (OMVs) from Escherichia coli Using EVscore47 Beads

Abstract
1. Introduction
2. Results and Discussion
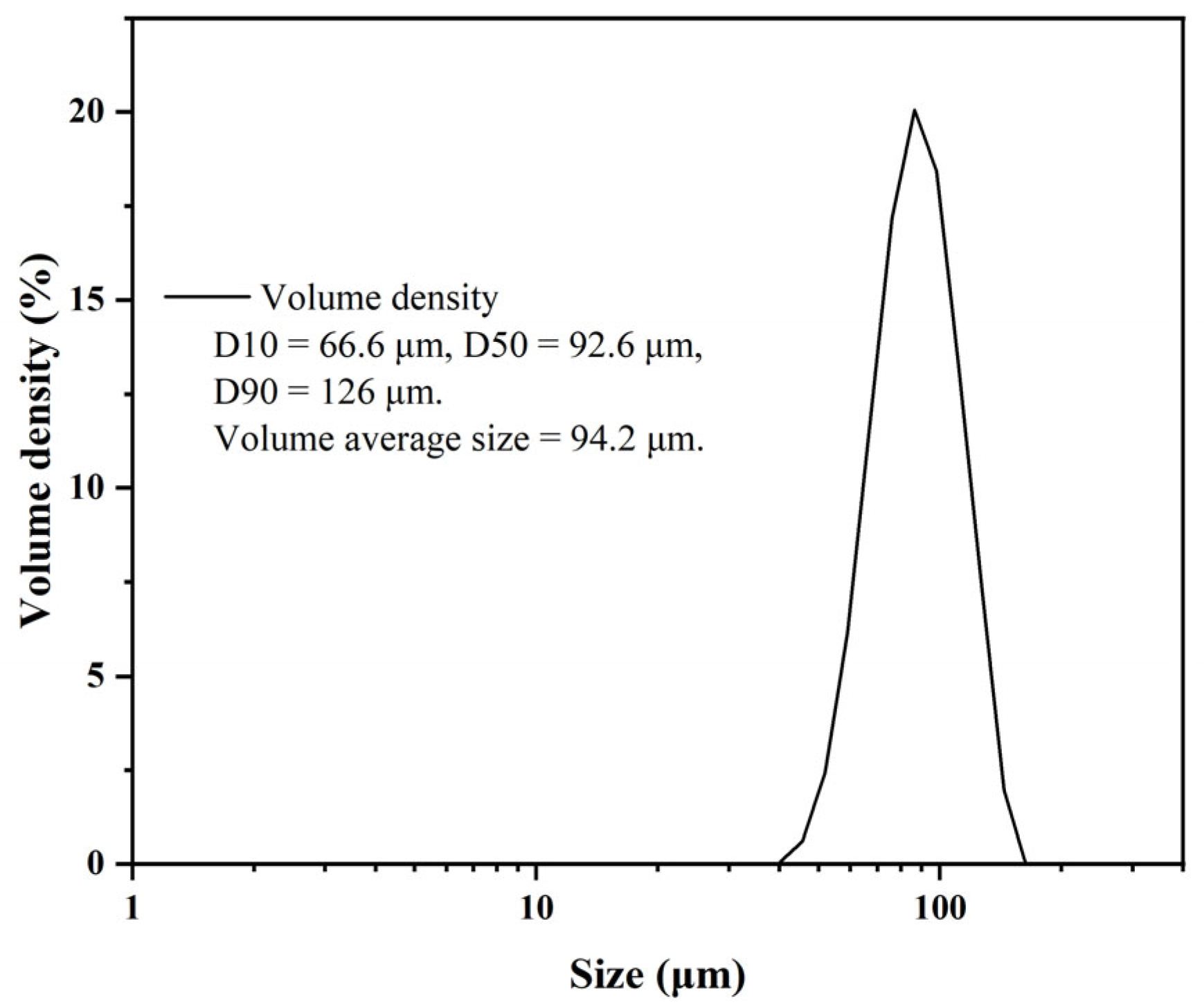
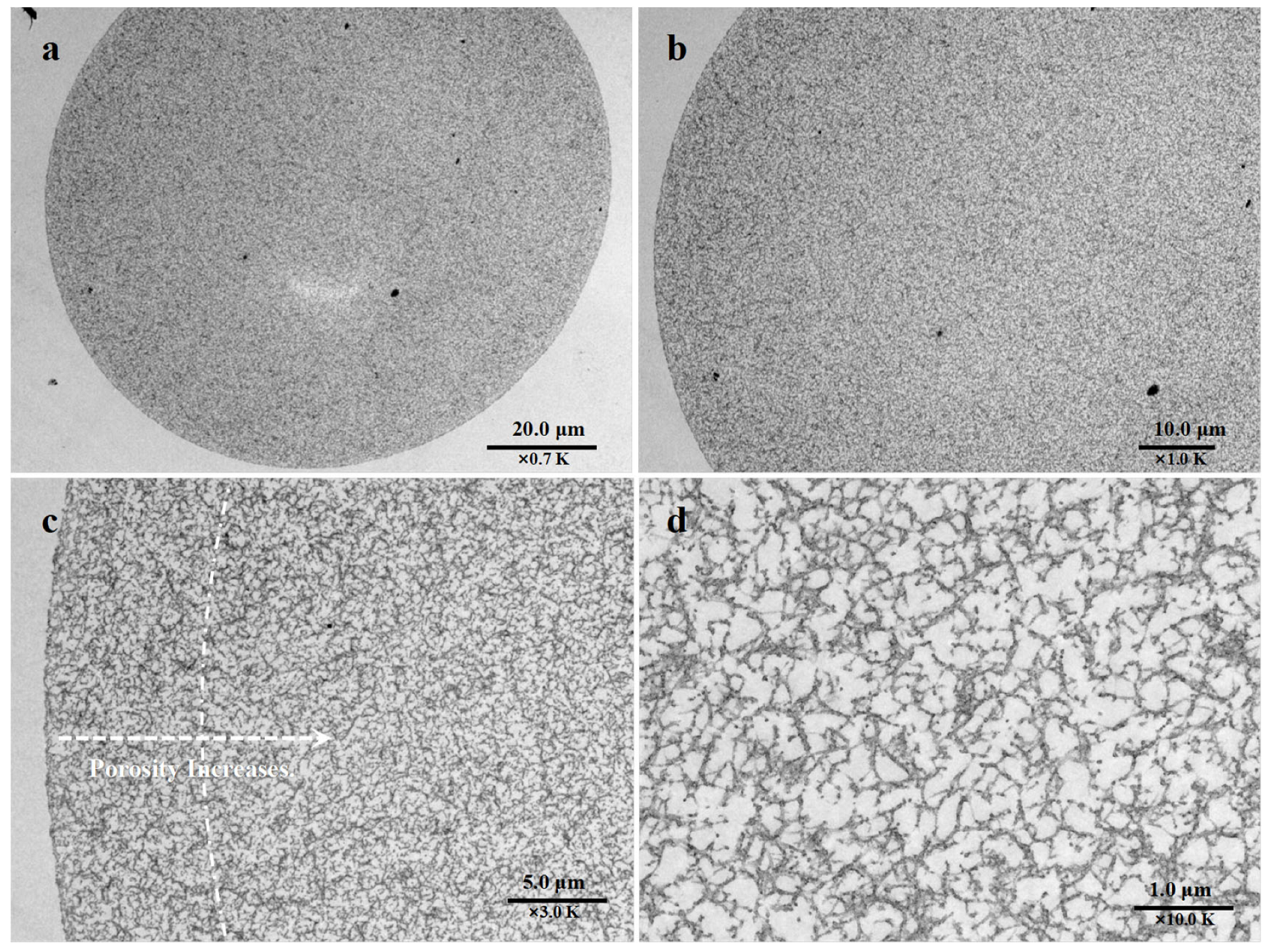
2.1. Preparation and Characterization of EVscore47 Microspheres
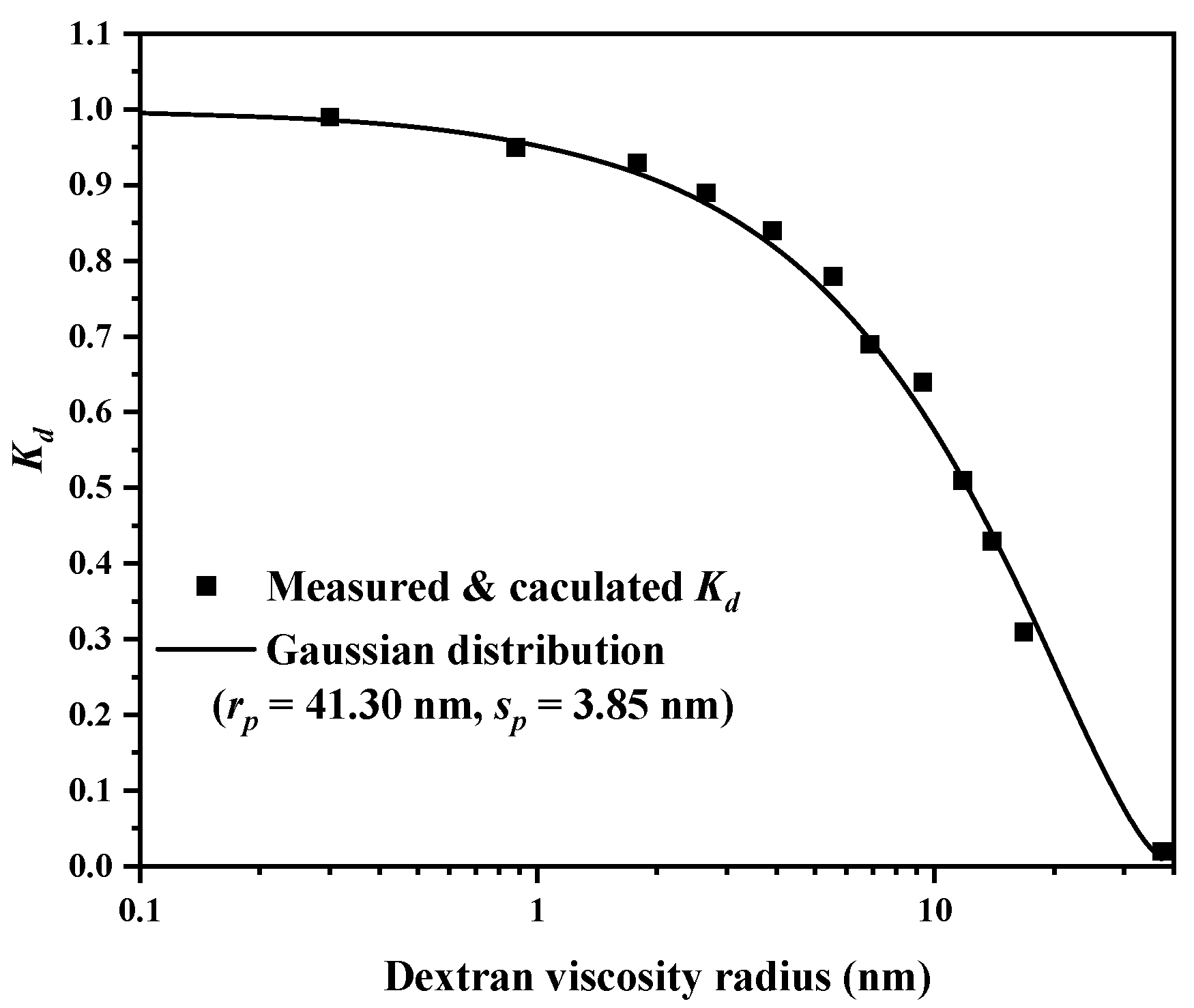
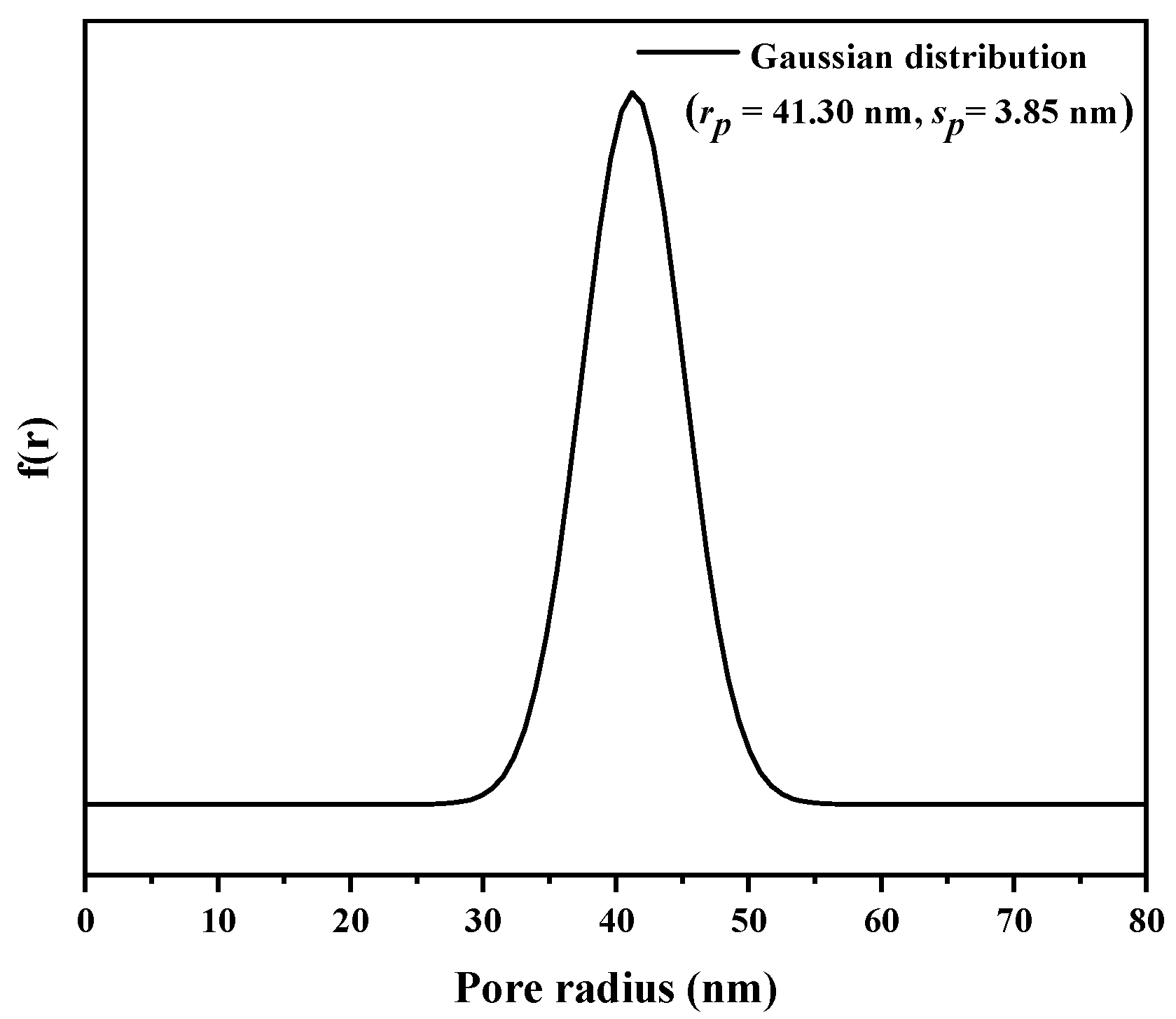
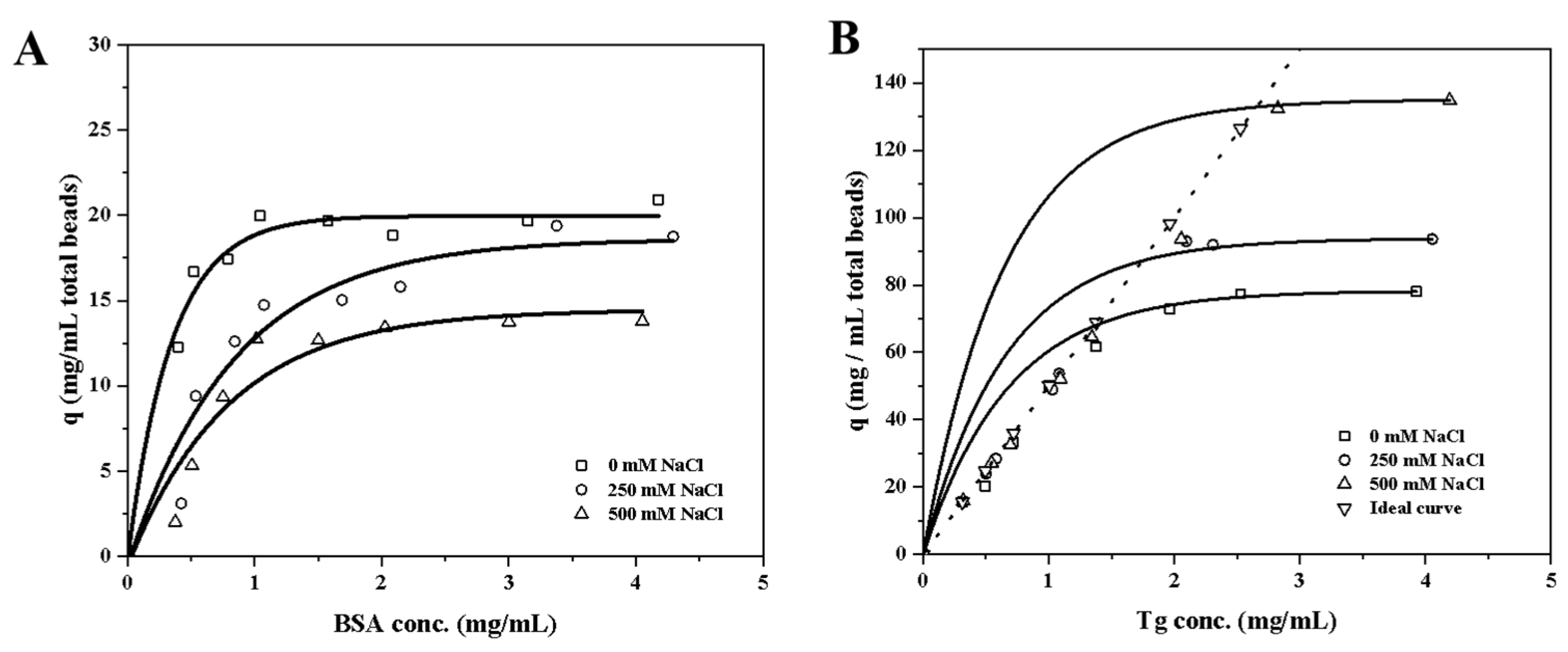
2.2. Adsorption Isotherms
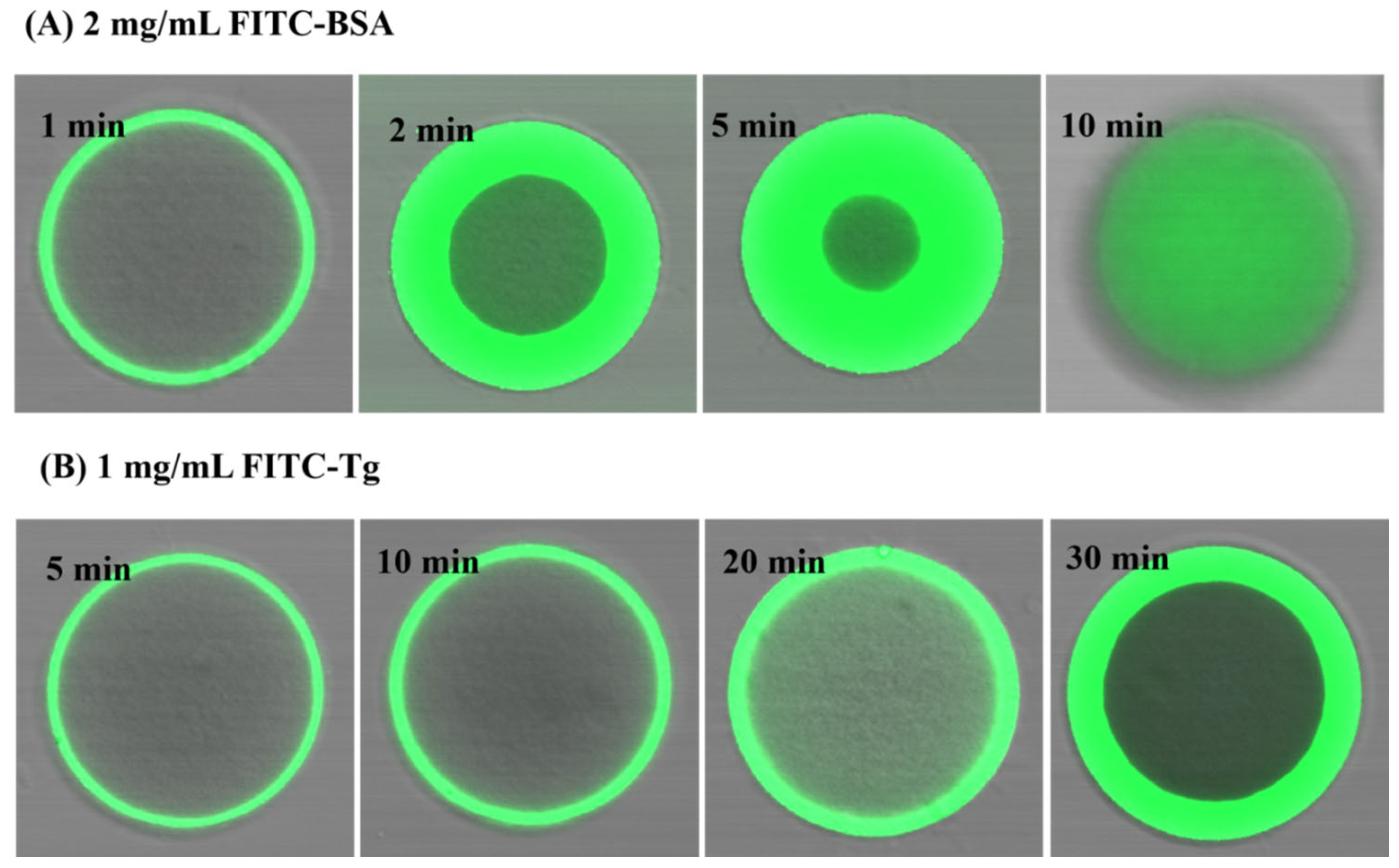
2.3. Confocal Laser Scanning Microscopy (CLSM) Results
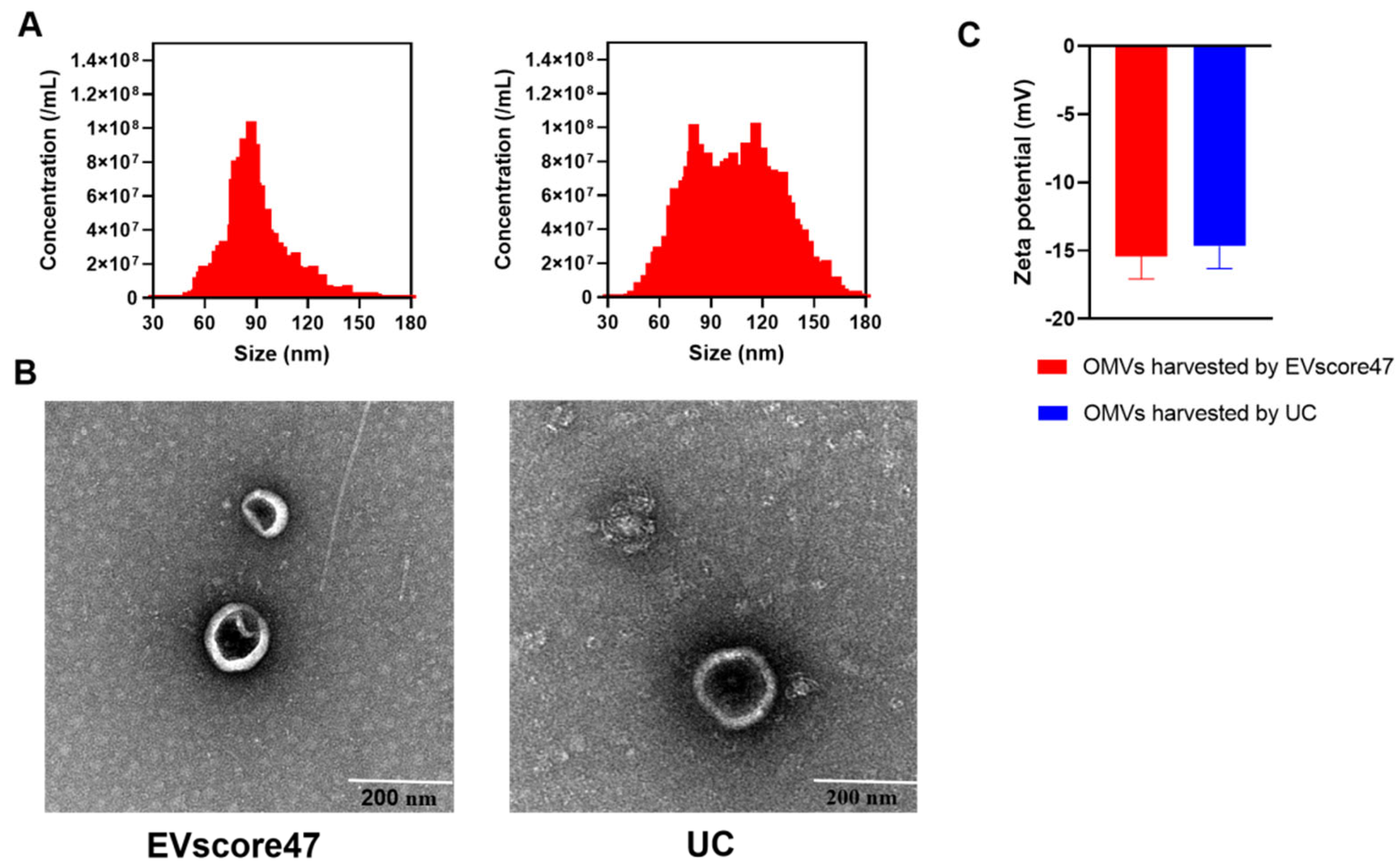
2.4. Isolation and Characterization of OMVs
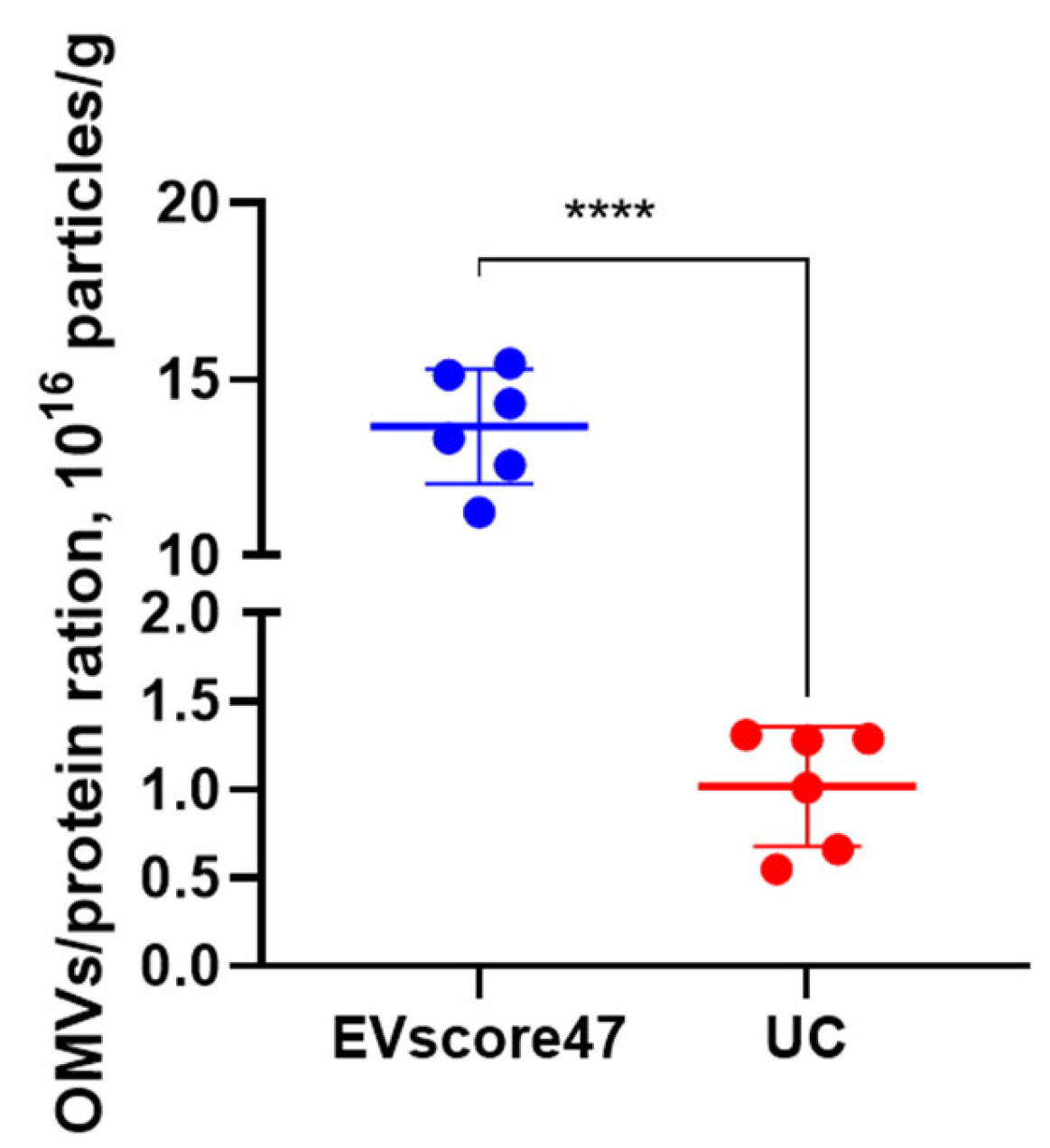
2.5. Amounts of OMVs
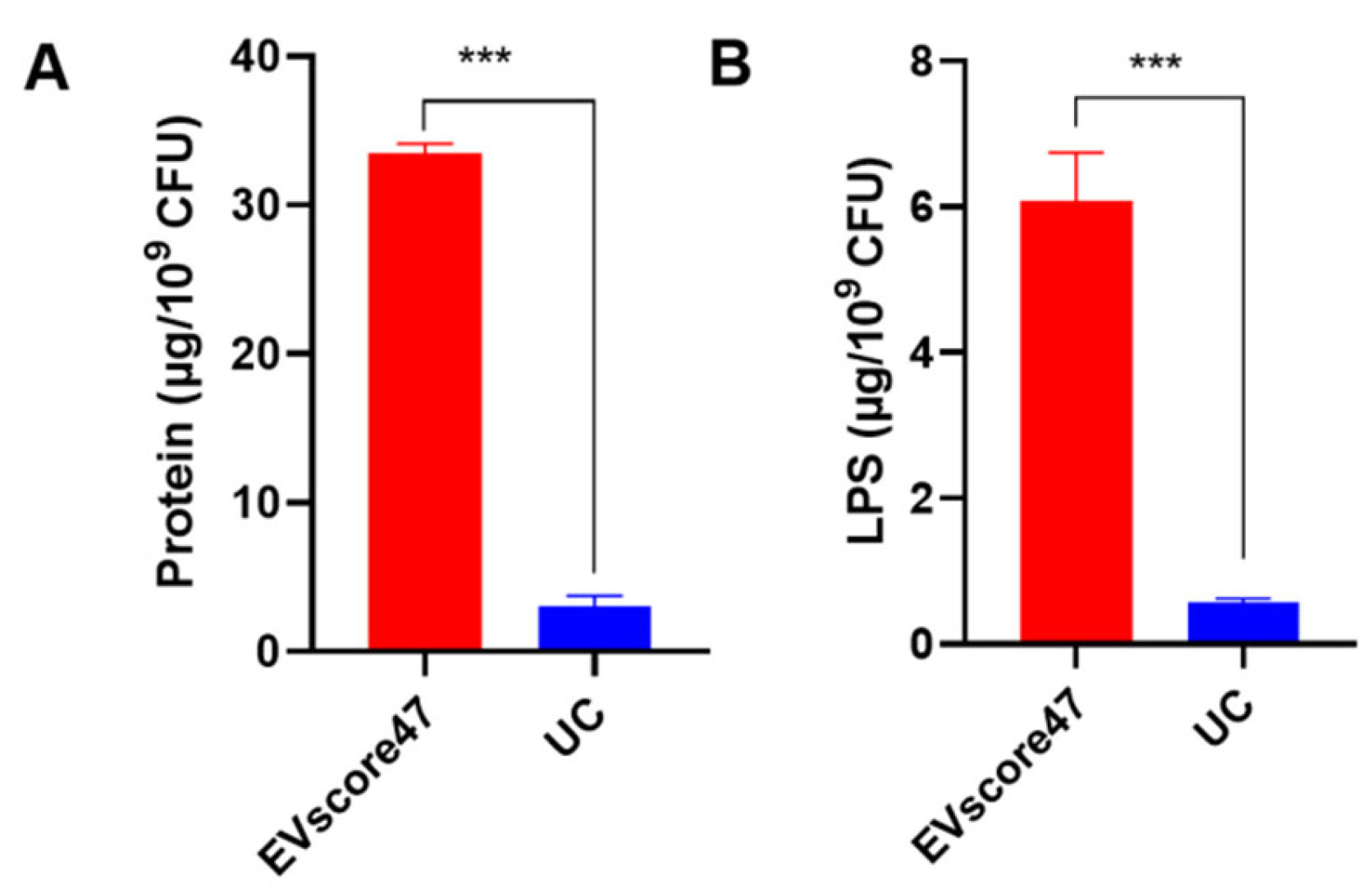
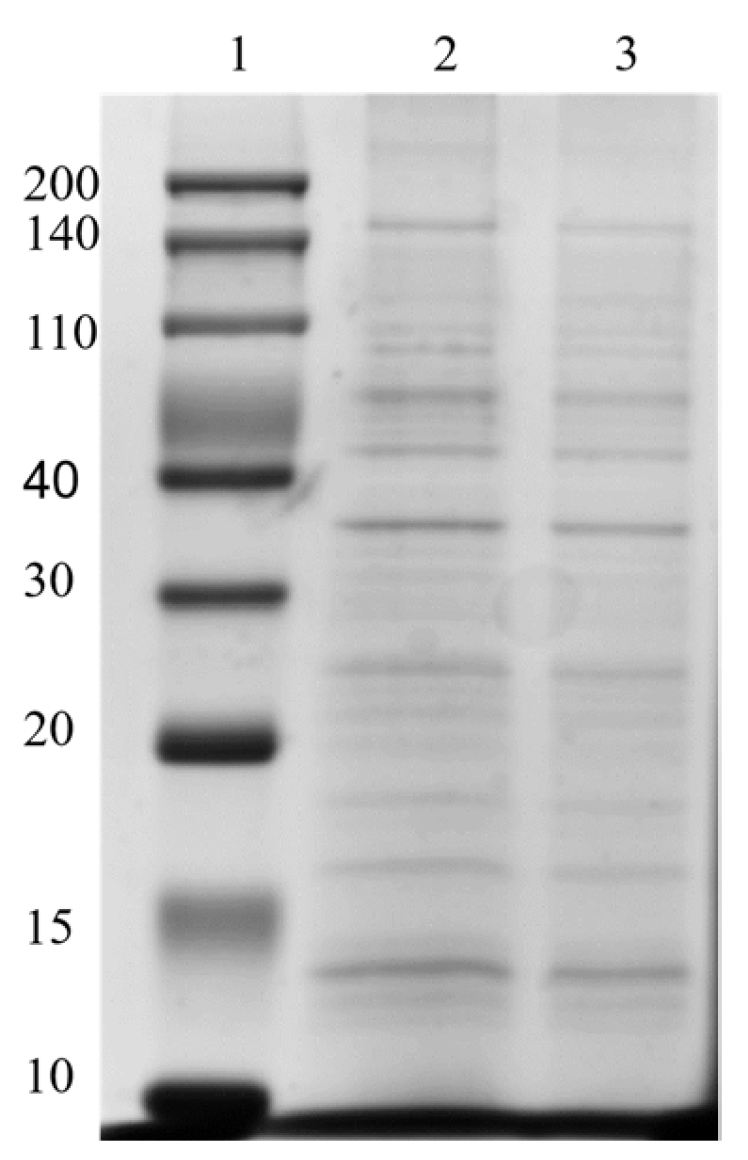
2.6. Protein Composition of OMVs
2.7. Purity of OMVs
3. Materials and Methods
3.1. Materials
3.2. Preparation of EVscore47 Microspheres
3.3. Structural Properties
3.4. Adsorption Isotherms
3.5. Confocal Microscopy
3.6. Isolation and Characterization of OMVs from E. coli
3.6.1. Bacterial Culture
3.6.2. Sample Processing
3.6.3. Purification of OMVs Using EVscore47 Beads
3.6.4. Isolation of OMVs via Ultracentrifugation
3.6.5. Characterization of OMVs
3.6.6. Protein and Lipopolysaccharide Contents in OMVs
3.6.7. Protein Composition of OMVs
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Furuyama, N.; Sircili, M.P. Outer membrane vesicles (OMVs) produced by gram-negative bacteria: Structure, functions, biogenesis, and vaccine application. BioMed Res. Int. 2021, 2021, 1490732. [Google Scholar] [CrossRef] [PubMed]
- Juodeikis, R.; Carding, S.R. Outer membrane vesicles: Biogenesis, functions, and issues. Microbiol. Mol. Biol. Rev. 2022, 86, e00032-22. [Google Scholar] [CrossRef] [PubMed]
- Kaparakis-Liaskos, M.; Ferrero, R.L. Immune modulation by bacterial outer membrane vesicles. Nat. Rev. Immunol. 2015, 15, 375–387. [Google Scholar] [CrossRef]
- Micoli, F.; Adamo, R.; Nakakana, U. Outer membrane vesicle vaccine platforms. BioDrugs 2024, 38, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Prior, J.T.; Davitt, C.; Kurtz, J.; Gellings, P.; McLachlan, J.B.; Morici, L.A. Bacterial-derived outer membrane vesicles are potent adjuvants that drive humoral and cellular immune responses. Pharmaceutics 2021, 13, 131. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.-W.; Wang, M.-Z.; Niu, J.; Chu, Y.; Guo, K.-R.; Peng, L.-H. Outer membrane vesicles derived from E. coli as novel vehicles for transdermal and tumor targeting delivery. Nanoscale 2020, 12, 18965–18977. [Google Scholar] [CrossRef] [PubMed]
- Dell’Annunziata, F.; Dell’Aversana, C.; Doti, N.; Donadio, G.; Dal Piaz, F.; Izzo, V.; De Filippis, A.; Galdiero, M.; Altucci, L.; Boccia, G. Outer membrane vesicles derived from Klebsiella pneumoniae are a driving force for horizontal gene transfer. Int. J. Mol. Sci. 2021, 22, 8732. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.Q.; Besio, R.; Xiao, L.; Forlino, A. Outer membrane vesicles (OMVs) as biomedical tools and their relevance as immune-modulating agents against H. pylori infections: Current status and future prospects. Int. J. Mol. Sci. 2023, 24, 8542. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Azze, R.F. Cross-protection induced by VA-MENGOC-BC® vaccine. Hum. Vaccines Immunother. 2018, 14, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Vipond, C.; Findlay, L.; Feavers, I.; Care, R. Limitations of the rabbit pyrogen test for assessing meningococcal OMV based vaccines. ALTEX-Altern. Anim. Exp. 2016, 33, 47–53. [Google Scholar] [CrossRef][Green Version]
- Oster, P.; Lennon, D.; O’Hallahan, J.; Mulholland, K.; Reid, S.; Martin, D. MeNZB™: A safe and highly immunogenic tailor-made vaccine against the New Zealand Neisseria meningitidis serogroup B disease epidemic strain. Vaccine 2005, 23, 2191–2196. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.S.; Turner, D.P. Clinical experience with the meningococcal B vaccine, Bexsero®: Prospects for reducing the burden of meningococcal serogroup B disease. Vaccine 2016, 34, 875–880. [Google Scholar] [CrossRef]
- Ahonkhai, V.; Lukacs, L.; Jonas, L.; Calandra, G. Clinical experience with PedvaxHIB, a conjugate vaccine of Haemophilus influenzae type b polysaccharide-Neisseria meningitidis outer membrane protein. Vaccine 1991, 9, S38–S41. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Xu, J.; Li, Y.; Cheng, K.; Feng, Q.; Ma, X.; Ma, N.; Zhang, T.; Wang, X.; Zhao, X. Antigen-bearing outer membrane vesicles as tumour vaccines produced in situ by ingested genetically engineered bacteria. Nat. Biomed. Eng. 2022, 6, 898–909. [Google Scholar] [CrossRef] [PubMed]
- Won, S.; Lee, C.; Bae, S.; Lee, J.; Choi, D.; Kim, M.G.; Song, S.; Lee, J.; Kim, E.; Shin, H. Mass-produced gram-negative bacterial outer membrane vesicles activate cancer antigen-specific stem-like CD8+ T cells which enables an effective combination immunotherapy with anti-PD-1. J. Extracell. Vesicles 2023, 12, 12357. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Feng, Q.; Wang, J.; Zhao, X. Bacterial outer membrane vesicle-based cancer nanovaccines. Cancer Biol. Med. 2022, 19, 1290. [Google Scholar] [CrossRef]
- Sarra, A.; Celluzzi, A.; Bruno, S.P.; Ricci, C.; Sennato, S.; Ortore, M.G.; Casciardi, S.; Del Chierico, F.; Postorino, P.; Bordi, F. Biophysical characterization of membrane phase transition profiles for the discrimination of outer membrane vesicles (OMVs) from Escherichia coli grown at different temperatures. Front. Microbiol. 2020, 11, 290. [Google Scholar] [CrossRef] [PubMed]
- Benedikter, B.J.; Bouwman, F.G.; Vajen, T.; Heinzmann, A.C.; Grauls, G.; Mariman, E.C.; Wouters, E.F.; Savelkoul, P.H.; Lopez-Iglesias, C.; Koenen, R.R. Ultrafiltration combined with size exclusion chromatography efficiently isolates extracellular vesicles from cell culture media for compositional and functional studies. Sci. Rep. 2017, 7, 15297. [Google Scholar] [CrossRef]
- Sharif, E.; Eftekhari, Z.; Mohit, E. The effect of growth stage and isolation method on properties of ClearColi™ outer membrane vesicles (OMVs). Curr. Microbiol. 2021, 78, 1602–1614. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.J.; Turner, K.B.; DiVito, K.A.; Daniele, M.A.; Walper, S.A. Affinity purification of bacterial outer membrane vesicles (OMVs) utilizing a His-tag mutant. Res. Microbiol. 2017, 168, 139–146. [Google Scholar] [CrossRef]
- Collins, S.M.; Nice, J.B.; Chang, E.H.; Brown, A.C. Size exclusion chromatography to analyze bacterial outer membrane vesicle heterogeneity. J. Vis. Exp. 2021, 169, e62429. [Google Scholar]
- Castillo-Romero, K.F.; Santacruz, A.; González-Valdez, J. Production and purification of bacterial membrane vesicles for biotechnology applications: Challenges and opportunities. Electrophoresis 2023, 44, 107–124. [Google Scholar] [CrossRef]
- Sadeghi, L.; Mohit, E.; Moallemi, S.; Ahmadi, F.M.; Bolhassani, A. Recent advances in various bio-applications of bacteria-derived outer membrane vesicles. Microb. Pathog. 2023, 185, 106440. [Google Scholar] [CrossRef]
- Hong, J.; Dauros-Singorenko, P.; Whitcombe, A.; Payne, L.; Blenkiron, C.; Phillips, A.; Swift, S. Analysis of the Escherichia coli extracellular vesicle proteome identifies markers of purity and culture conditions. J. Extracell. Vesicles 2019, 8, 1632099. [Google Scholar] [CrossRef]
- Corso, G.; Mäger, I.; Lee, Y.; Görgens, A.; Bultema, J.; Giebel, B.; Wood, M.J.; Nordin, J.Z.; Andaloussi, S.E. Reproducible and scalable purification of extracellular vesicles using combined bind-elute and size exclusion chromatography. Sci. Rep. 2017, 7, 11561. [Google Scholar] [CrossRef]
- Onódi, Z.; Pelyhe, C.; Terézia Nagy, C.; Brenner, G.B.; Almási, L.; Kittel, Á.; Manček-Keber, M.; Ferdinandy, P.; Buzás, E.I.; Giricz, Z. Isolation of high-purity extracellular vesicles by the combination of iodixanol density gradient ultracentrifugation and bind-elute chromatography from blood plasma. Front. Physiol. 2018, 9, 1479. [Google Scholar] [CrossRef]
- McNamara, R.P.; Caro-Vegas, C.P.; Costantini, L.M.; Landis, J.T.; Griffith, J.D.; Damania, B.A.; Dittmer, D.P. Large-scale, cross-flow based isolation of highly pure and endocytosis-competent extracellular vesicles. J. Extracell. Vesicles 2018, 7, 1541396. [Google Scholar] [CrossRef]
- Sánchez-Trasviña, C.; Fuks, P.; Mushagasha, C.; Kimerer, L.; Mayolo-Deloisa, K.; Rito-Palomares, M.; Carta, G. Structure and functional properties of Capto™ Core 700 core-shell particles. J. Chromatogr. A 2020, 1621, 461079. [Google Scholar] [CrossRef]
- Matlschweiger, A.; Engelmaier, H.; Himmler, G.; Hahn, R. Secretory immunoglobulin purification from whey by chromatographic techniques. J. Chromatogr. B 2017, 1060, 53–62. [Google Scholar] [CrossRef]
- Tseng, Y.-F.; Weng, T.-C.; Lai, C.-C.; Chen, P.-L.; Lee, M.-S.; Hu, A.Y.-C. A fast and efficient purification platform for cell-based influenza viruses by flow-through chromatography. Vaccine 2018, 36, 3146–3152. [Google Scholar] [CrossRef]
- Cunha, B.; Silva, R.J.; Aguiar, T.; Serra, M.; Daicic, J.; Maloisel, J.-L.; Clachan, J.; Åkerblom, A.; Carrondo, M.J.; Peixoto, C. Improving washing strategies of human mesenchymal stem cells using negative mode expanded bed chromatography. J. Chromatogr. A 2016, 1429, 292–303. [Google Scholar] [CrossRef]
- Shen, Z.; Lin, D.Q.; Yao, S. Determination of pore size distribution of porous media by inverse size-exclusion chromatography. CIESC J. 2011, 61, 867–874. [Google Scholar]
- DePhillips, P.; Lenhoff, A.M. Pore size distributions of cation-exchange adsorbents determined by inverse size-exclusion chromatography. J. Chromatogr. A 2000, 883, 39–54. [Google Scholar] [CrossRef]
- Tulkens, J.; Wever, O.D.; Hendrix, A. Increased levels of systemic LPS-positive bacterial extracellular vesicles in cancer patients with chemotherapy-induced intestinal barrier dysfunction. Cancer Res. 2019, 79 (Suppl. S13), 1489. [Google Scholar] [CrossRef]
- Welsh, J.A.; Goberdhan, D.C.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.; Erdbrügger, U. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef]
- Shi, R.; Dong, Z.; Ma, C.; Wu, R.; Lv, R.; Liu, S.; Ren, Y.; Liu, Z.; van der Mei, H.C.; Busscher, H.J. High-yield, magnetic harvesting of extracellular outer-membrane vesicles from Escherichia coli. Small 2022, 18, 2204350. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | 0 mM NaCl | 250 mM NaCl | 500 mM NaCl | |||
|---|---|---|---|---|---|---|
| qm | KL | qm | KL | qm | KL | |
| BSA | 20.45 | 9.54 | 20.13 | 1.81 | 15.58 | 1.94 |
| Tg | 84.04 | 2.61 | 100.15 | 2.77 | 143.44 | 2.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, G.; Yang, X.; Wang, J.; Wei, W.; Hu, K.; Huang, X.; Qiu, Y.; He, Y. Isolation of Extracellular Outer Membrane Vesicles (OMVs) from Escherichia coli Using EVscore47 Beads. Molecules 2024, 29, 1831. https://doi.org/10.3390/molecules29081831
Shi G, Yang X, Wang J, Wei W, Hu K, Huang X, Qiu Y, He Y. Isolation of Extracellular Outer Membrane Vesicles (OMVs) from Escherichia coli Using EVscore47 Beads. Molecules. 2024; 29(8):1831. https://doi.org/10.3390/molecules29081831
Chicago/Turabian StyleShi, Gongming, Xiaohong Yang, Jikai Wang, Wenjing Wei, Kecui Hu, Xingyue Huang, Yanfei Qiu, and Yun He. 2024. "Isolation of Extracellular Outer Membrane Vesicles (OMVs) from Escherichia coli Using EVscore47 Beads" Molecules 29, no. 8: 1831. https://doi.org/10.3390/molecules29081831
APA StyleShi, G., Yang, X., Wang, J., Wei, W., Hu, K., Huang, X., Qiu, Y., & He, Y. (2024). Isolation of Extracellular Outer Membrane Vesicles (OMVs) from Escherichia coli Using EVscore47 Beads. Molecules, 29(8), 1831. https://doi.org/10.3390/molecules29081831