
Ameliorative Effects of Zingiber officinale Rosc on Antibiotic-Associated Diarrhea and Improvement in Intestinal Function



Abstract
1. Introduction
2. Results and Discussion
2.1. Chemical Properties of ZO Extract
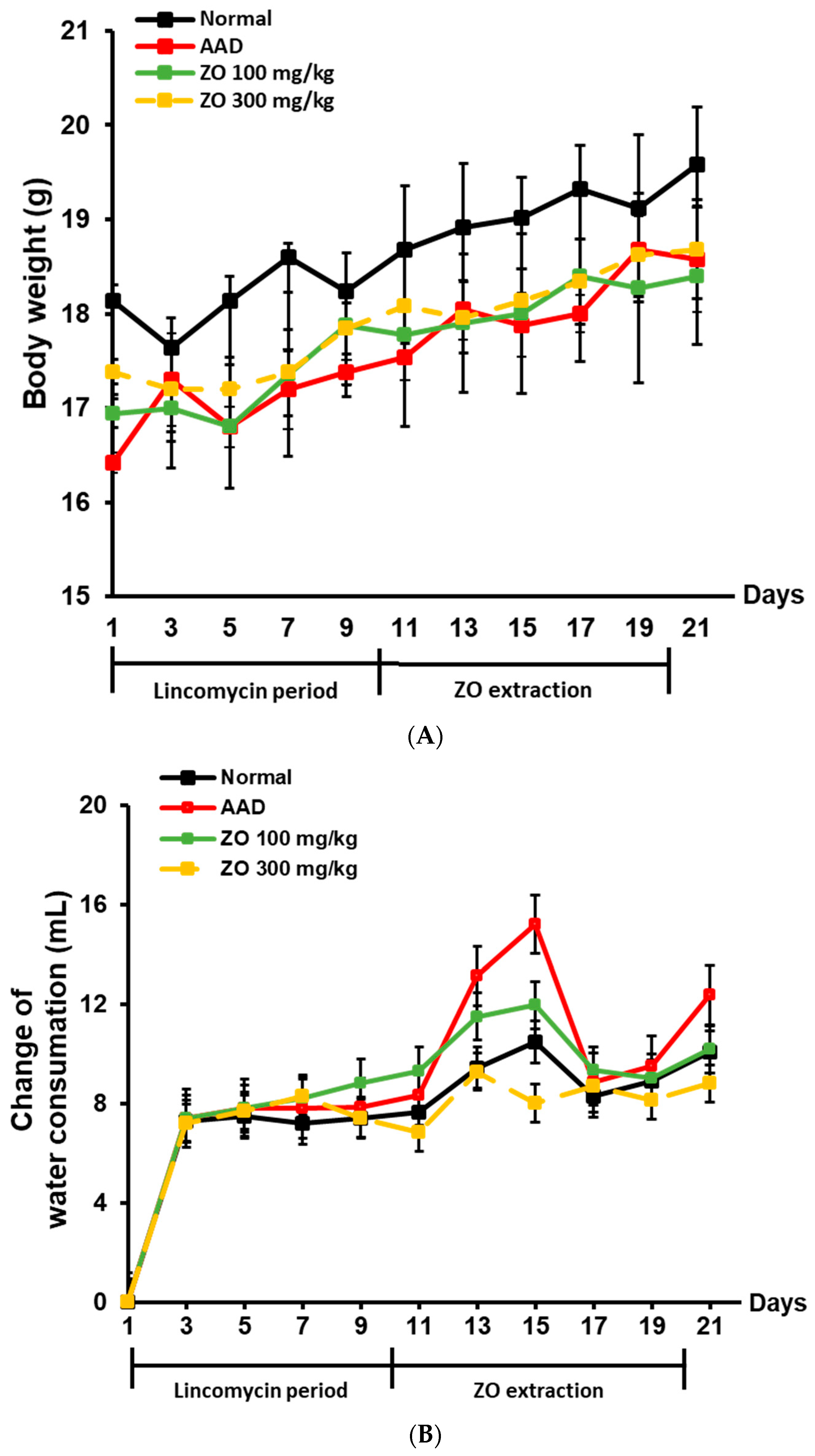
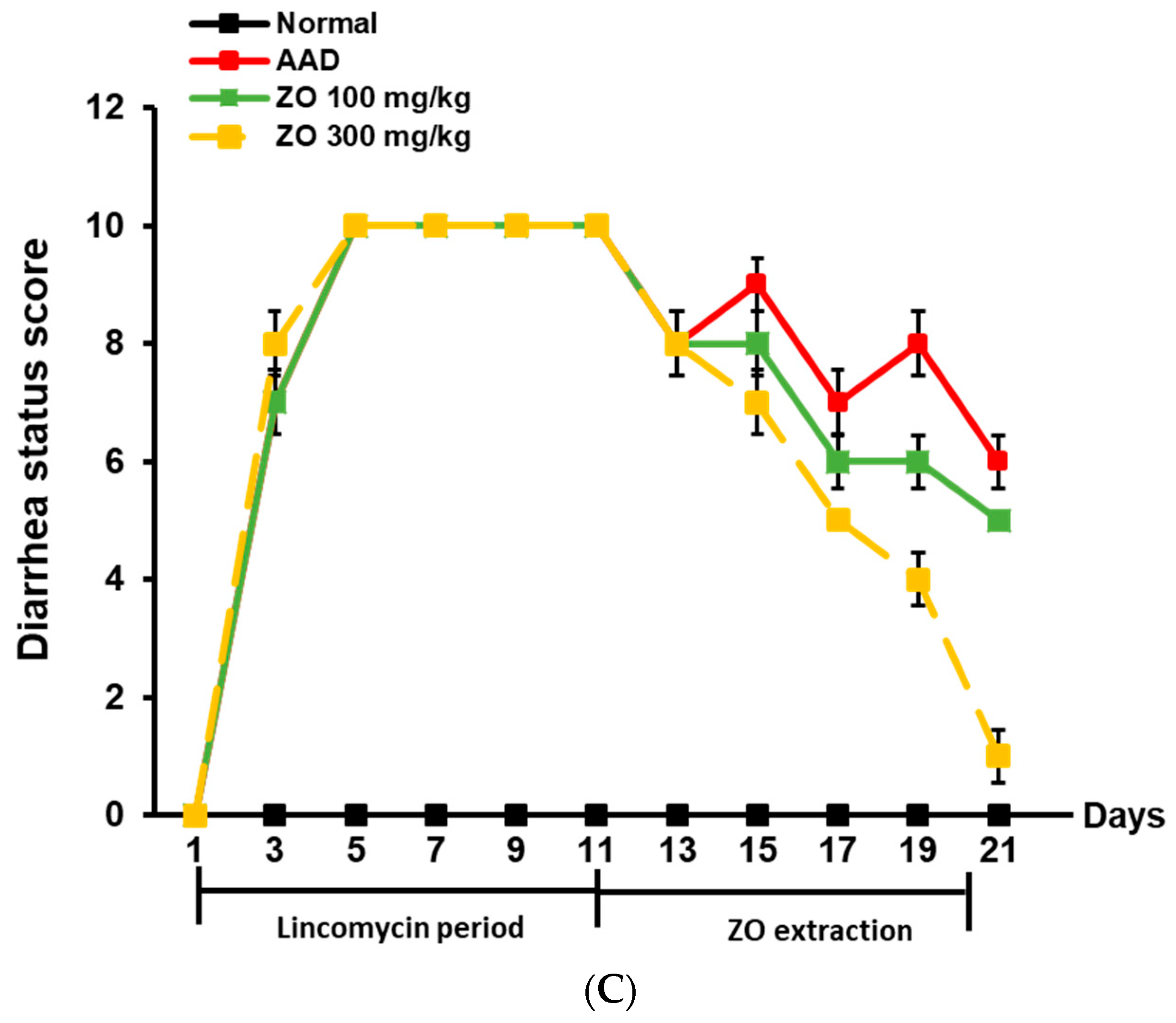
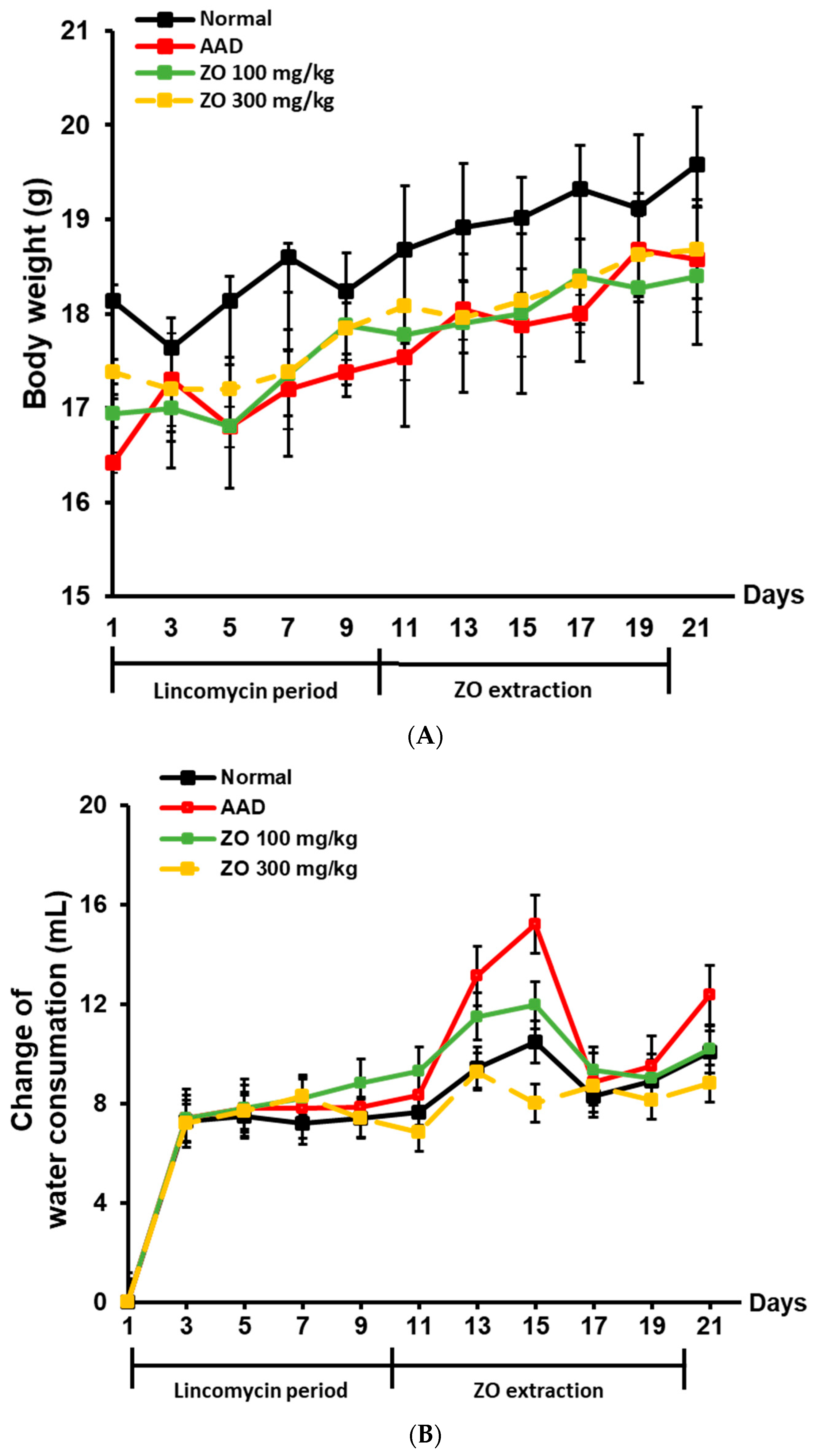
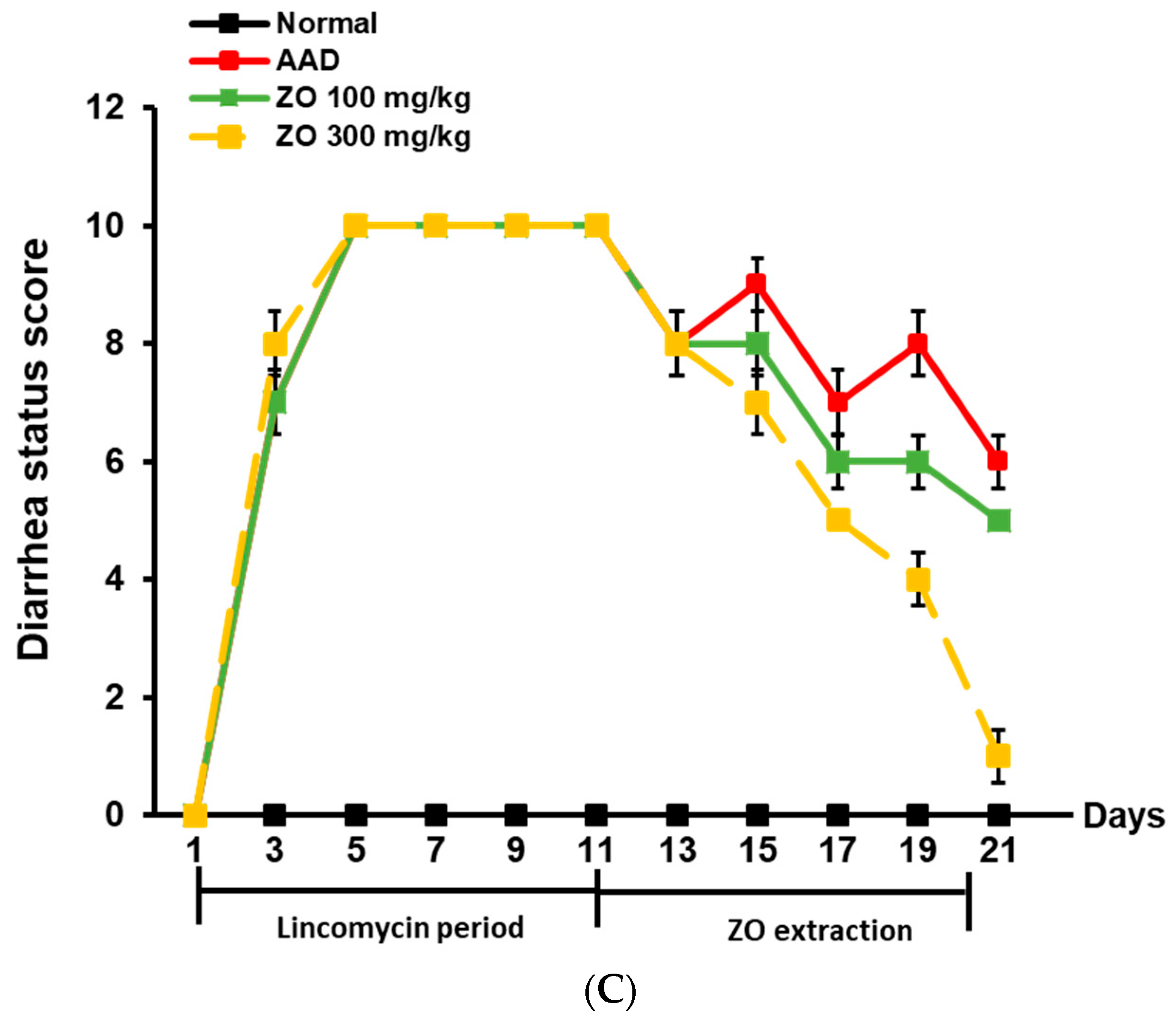
2.2. Ameliorating Effect of ZO Extract on AAD in a Lincomycin-Induced-Diarrhea Mouse Model
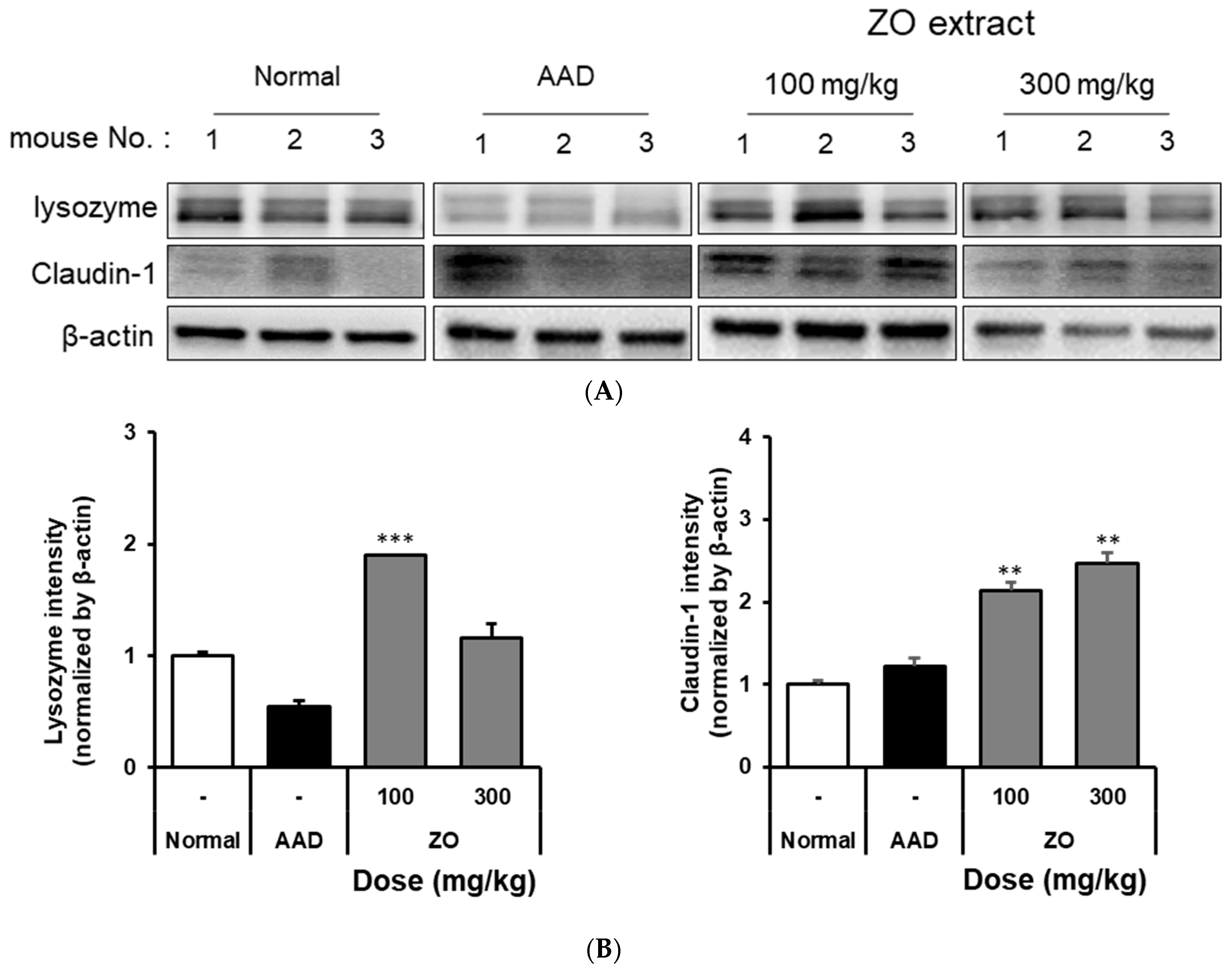
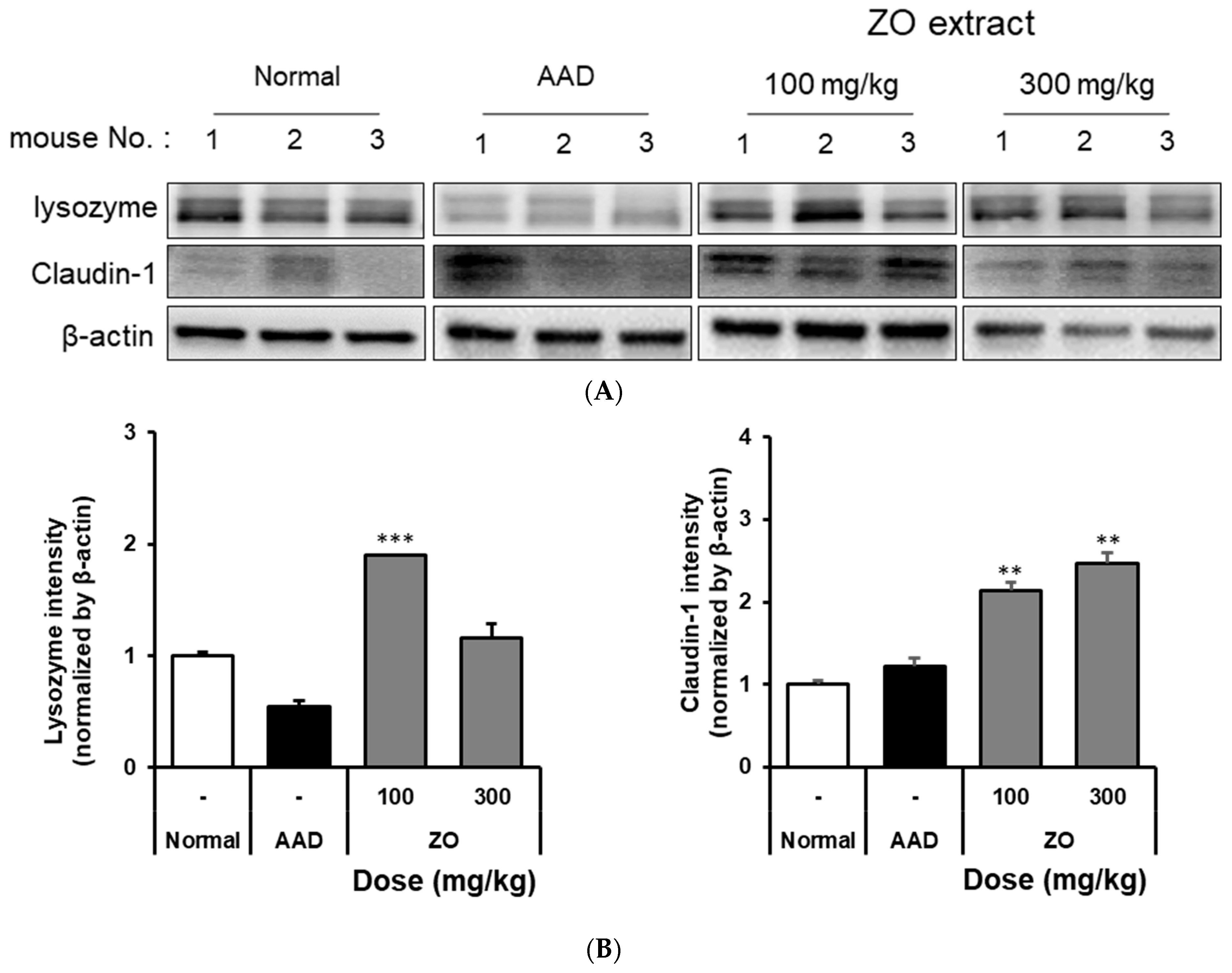
2.3. Analysis of the Effects of ZO Extract on Changes in the Expression of Intestinal Lysozyme and Claudin-1 in the Lincomycin-Induced-Diarrhea Mouse Model
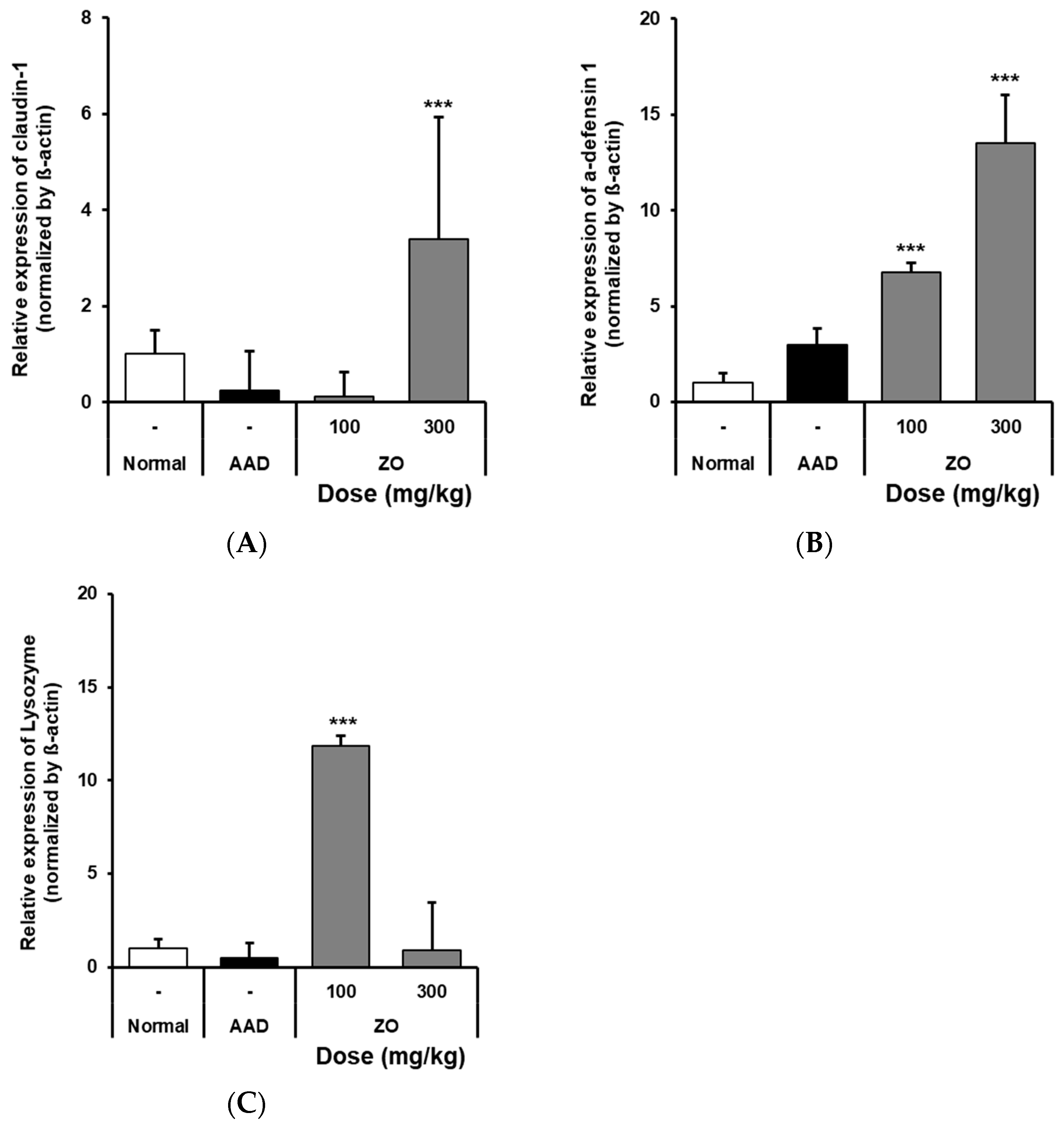
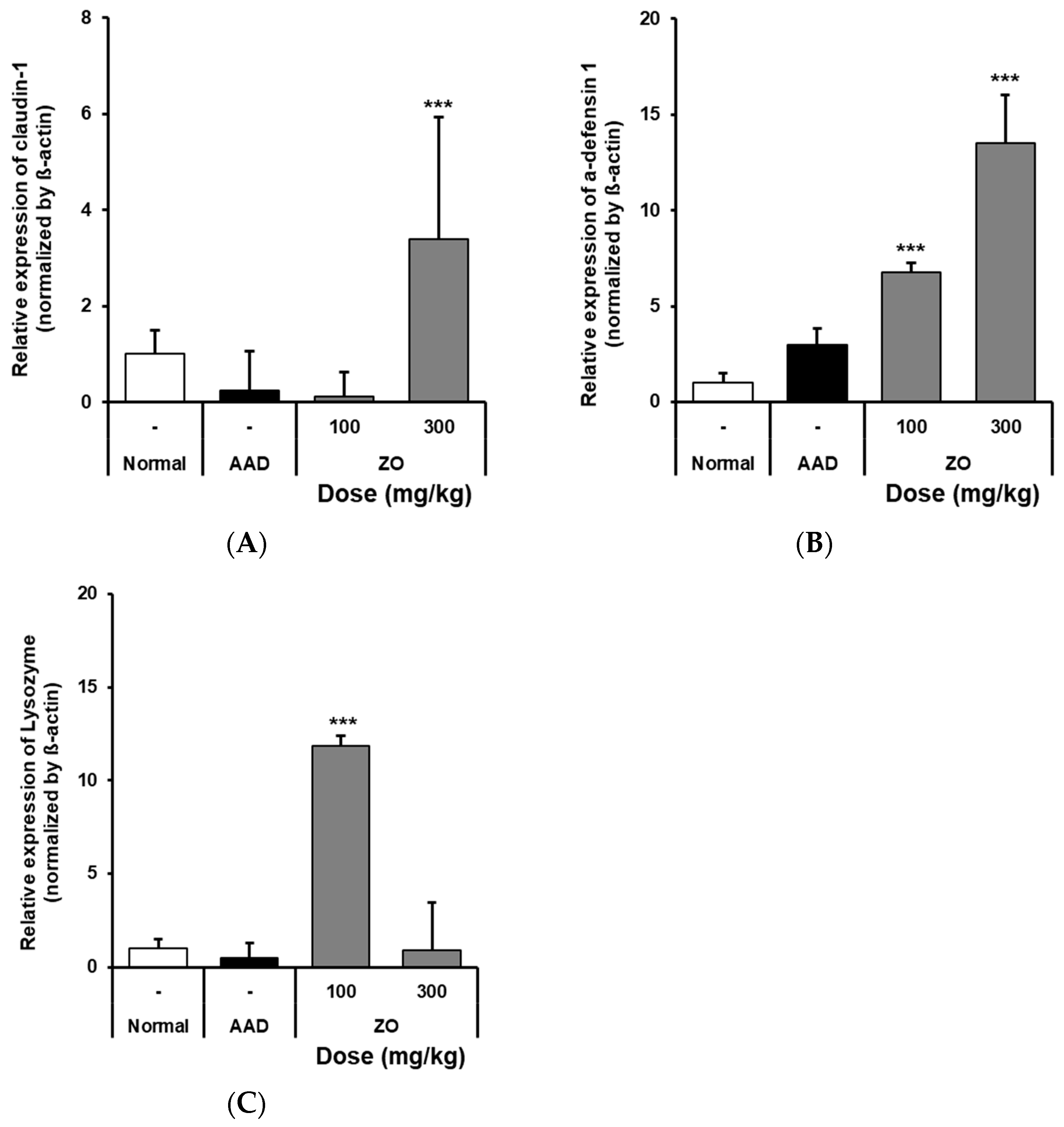
2.4. Analysis of the Effects of ZO Extract on the mRNA Expression of Intestinal Claudin-1, α-Defensin-1, and Lysozyme in the Lincomycin-Induced-Diarrhea Mouse Model
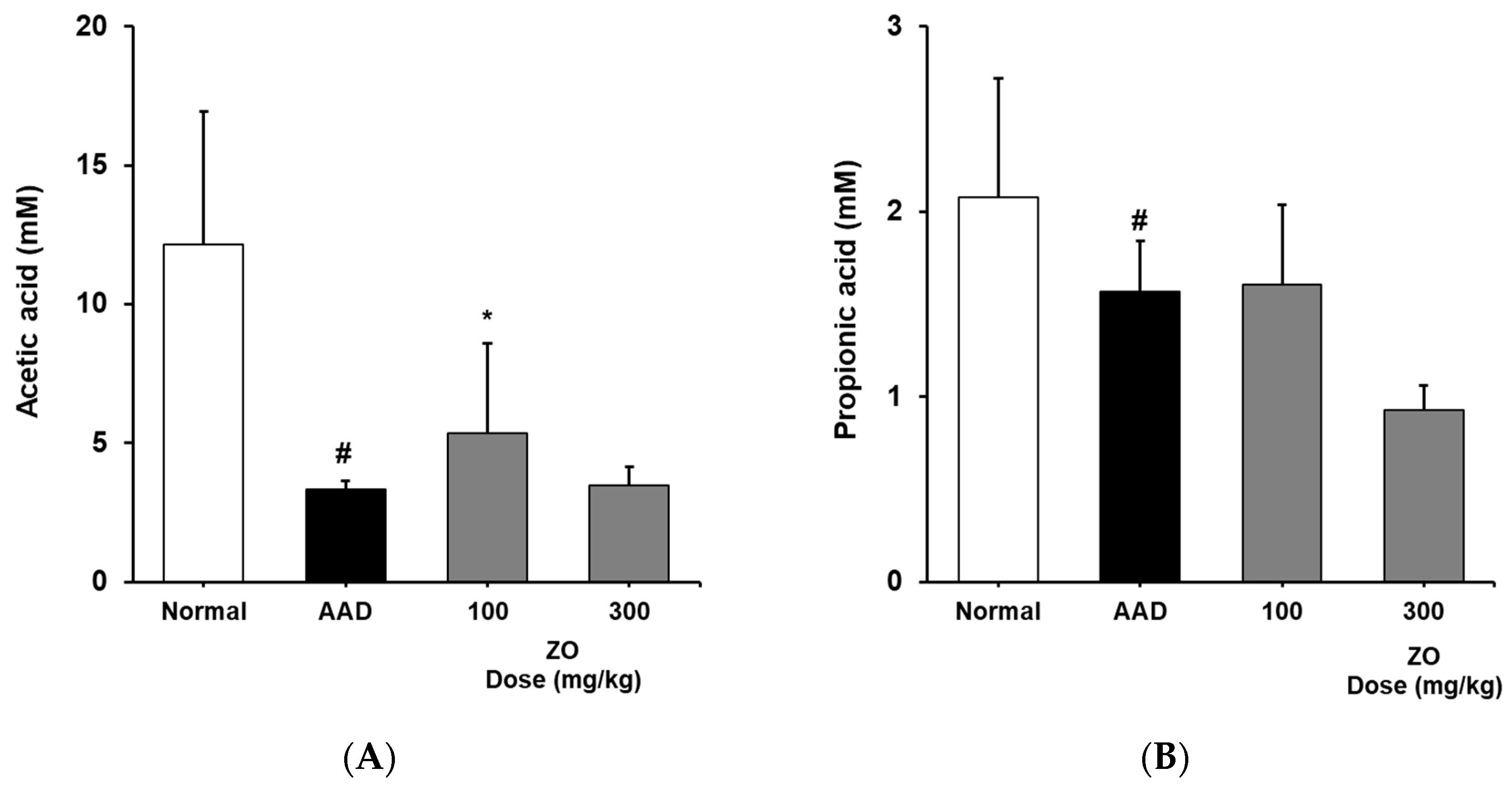
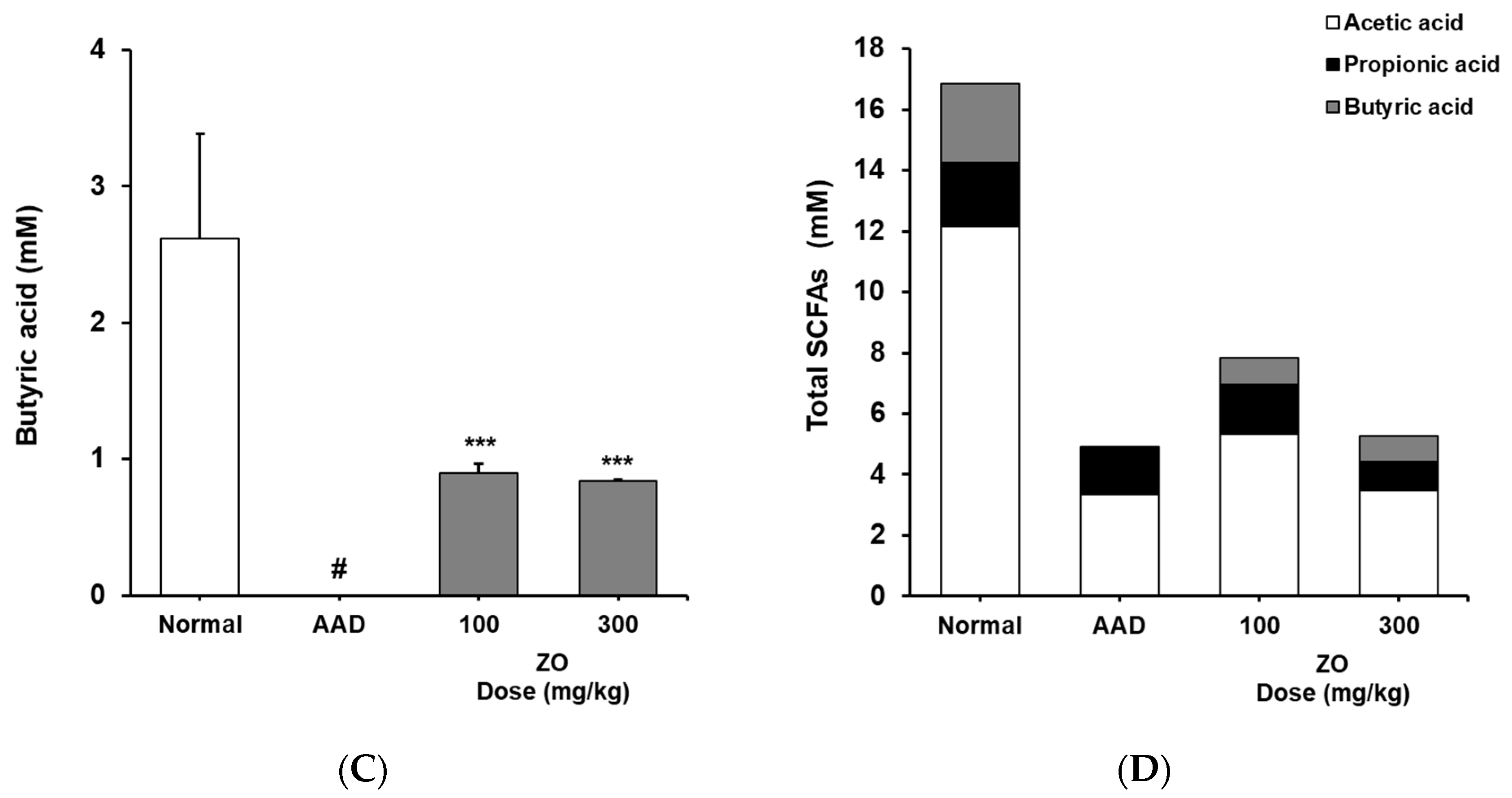
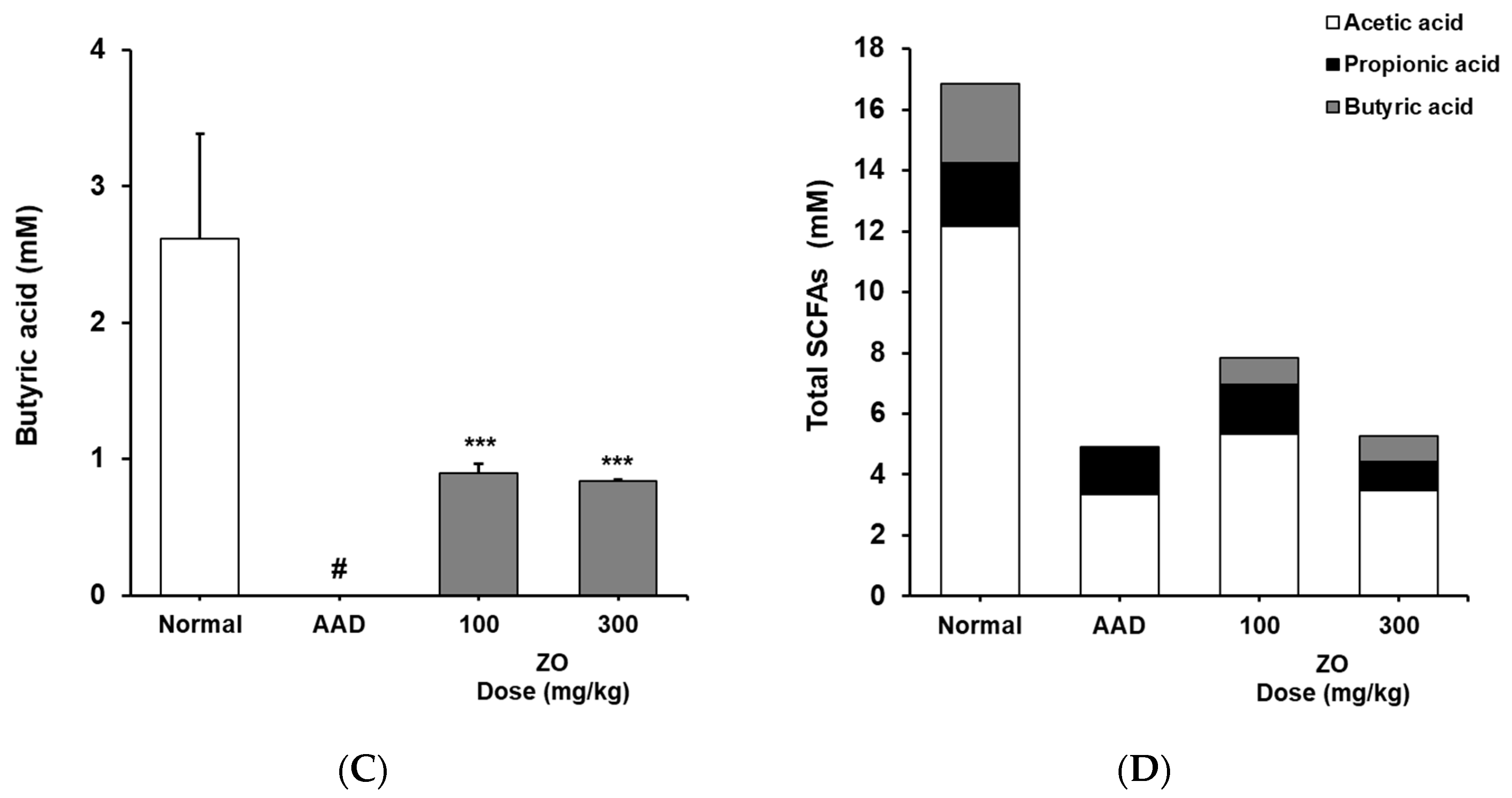
2.5. Analysis of the Effects of ZO Extract on the Changes in the Levels of SCFAs in the Lincomycin-Induced-Diarrhea Model
3. Materials and Methods
3.1. Preparation of ZO Extract
3.2. Chemicals and Antibodies
3.3. Monosaccharide Composition Analysis of ZO Extract
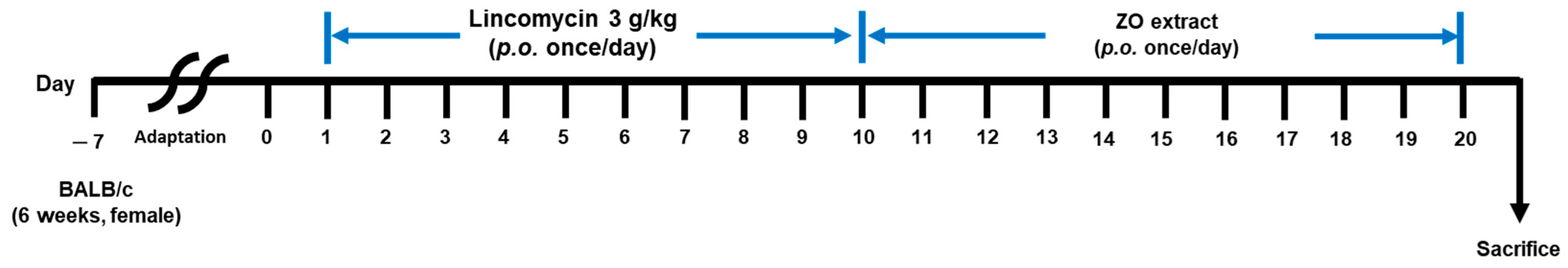
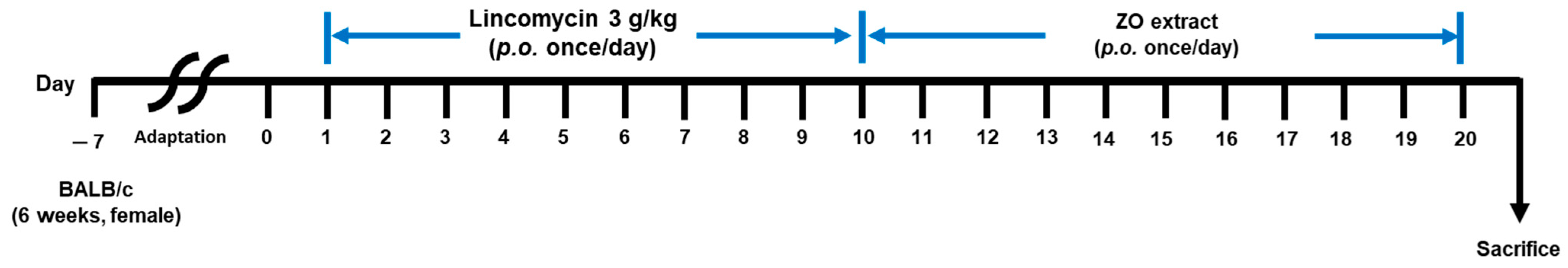
3.4. Animal and Experimental Design
3.5. Immunoblotting
3.6. RT-qPCR
3.7. Determination of Short-Chain Fatty Acids
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cohen, M.L. Changing patterns of infectious disease. Nature 2000, 406, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Aminov, R. History of antimicrobial drug discovery: Major classes and health impact. Biochem. Pharmacol. 2017, 133, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Nam, J.P.; Kim, J.H.; Kim, Y.M.; Nah, J.W.; Jang, M.K. Antimicrobial action of water-soluble β-chitosan against clinical multi-drug resistant bacteria. Int. J. Mol. Sci. 2015, 16, 7995–8007. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, K.; Xiao, N.; Peng, M.; Tan, Z. The effect of qiweibaizhu powder crude polysaccharide on antibiotic-associated diarrhea mice is associated with restoring intestinal mucosal bacteria. Front. Nutr. 2022, 9, 952647. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.; Liu, C.; Ye, C.; Sun, J.; Tan, X.; Zhang, C.; Qu, Q.; Shi, D.; Guo, S. Structural modulation of gut microbiota during alleviation of antibiotic-associated diarrhea with herbal formula. Int. J. Biol. Macromol. 2017, 105, 1622–1629. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Qin, P.; Wang, J. High-fat diet alters the intestinal microbiota in streptozotocin-induced type 2 diabetic mice. Microorganisms 2019, 7, 176. [Google Scholar] [CrossRef]
- La Fata, G.; Weber, P.; Mohajeri, M.H. Probiotics and the gut immune system: Indirect regulation. Probiotics Antimicrob. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, B.; Xu, J.; Liu, Y.; Qiu, E.; Li, Z.; Li, Z.; He, Y.; Zhou, H.; Bai, Y.; et al. Bacteroides fragilis protects against antibiotic-associated diarrhea in rats by modulating intestinal defenses. Front. Immunol. 2018, 9, 1040. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Q.; Huang, J.; Liu, D.; Lan, Y.; Yuan, L.; Chen, Q. Xianglian Pill ameliorates antibiotic-associated diarrhea by restoring intestinal microbiota and attenuating mucosal damage. J. Ethnopharmacol. 2021, 264, 113377. [Google Scholar] [CrossRef]
- Ramirez, J.; Guarner, F.; Bustos Fernandez, L.; Maruy, A.; Sdepanian, V.L.; Cohen, H. Antibiotics as major disruptors of gut microbiota. Front. Cell. Infect. Microbiol. 2020, 10, 572912. [Google Scholar] [CrossRef] [PubMed]
- Willing, B.P.; Russell, S.L.; Finlay, B.B. Shifting the balance: Antibiotic effects on host–microbiota mutualism. Nat. Rev. Microbiol. 2011, 9, 233–243. [Google Scholar] [CrossRef]
- Park, D.H.; Han, B.; Shin, M.S.; Hwang, G.S. Enhanced intestinal immune response in mice after oral administration of Korea red ginseng-derived polysaccharide. Polymers 2020, 12, 2186. [Google Scholar] [CrossRef]
- Ahn, H.-R.; Park, D.H.; Shin, M.-S.; Nguyen, Q.N.; Park, J.Y.; Kim, D.-W.; Kang, K.S.; Lee, H.L. The ameliorating effect of Lizhong-Tang on antibiotic-associated imbalance in the gut microbiota in mouse. Appl. Sci. 2022, 12, 6943. [Google Scholar] [CrossRef]
- Ma, Z.J.; Wang, H.J.; Ma, X.J.; Li, Y.; Yang, H.J.; Li, H.; Su, J.R.; Zhang, C.E.; Huang, L.Q. Modulation of gut microbiota and intestinal barrier function during alleviation of antibiotic-associated diarrhea with rhizoma Zingiber officinale (Ginger) extract. Food Funct. 2020, 11, 10839–10851. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.-S.; Yu, K.-W.; Shin, K.-S.; Lee, H. Enhancement of immunological activity in mice with oral administration of cell wall components of Bifidobacterium bifidum. Food Sci. Biotechnol. 2004, 13, 85–89. [Google Scholar]
- Min, S.J.; Kim, S.J.; Park, J.Y.; Seo, C.S.; Choi, Y.K. Preparation of herbal extracts for intestinal immune modulation activity based on in vitro screening and in vivo evaluation of Zingiber officinale Rosc. Extracts. Molecules 2023, 28, 6743. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.H.; Ni, Z.J.; Zhu, Y.Y.; Thakur, K.; Zhang, F.; Zhang, Y.Y.; Hu, F.; Zhang, J.G.; Wei, Z.J. A recent update on the multifaceted health benefits associated with ginger and its bioactive components. Food Funct. 2021, 12, 519–542. [Google Scholar] [CrossRef]
- Grzanna, R.; Lindmark, L.; Frondoza, C.G. Ginger—An herbal medicinal product with broad anti-inflammatory actions. J. Med. Food 2005, 8, 125–132. [Google Scholar] [CrossRef]
- Nikkhah Bodagh, M.; Maleki, I.; Hekmatdoost, A. Ginger in gastrointestinal disorders: A systematic review of clinical trials. Food Sci. Nutr. 2019, 7, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Blunden, G.; Tanira, M.O.; Nemmar, A. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol. 2008, 46, 409–420. [Google Scholar] [CrossRef]
- Connell DW, Sutherland MD A re-examination of gingerol, shogaol, and zingerone the pungent principles of ginger (Zingiber officinale Roscoe). Aust. J. Chem. 1969, 22, 1033–1043. [CrossRef]
- Lee BS, Ko MS, Kim HJ, Kwak IS, Kim DH, Chung BW Separation of 6-gingerol from ginger (Zingiber officinale Roscoe) and antioxidative activity. Korean J. Biotechnol. Bioeng. 2006, 21, 484–488.
- Moon, Y.S.; Lee, H.S.; Lee, S.E. Inhibitory effects of three monoterpenes from ginger essential oil on growth and aflatoxin production of Aspergillus flavus and their gene regulation in aflatoxin biosynthesis. Appl. Biol. Chem. 2018, 61, 243–250. [Google Scholar] [CrossRef]
- Shimoda, H.; Shan, S.J.; Tanaka, J.; Seki, A.; Seo, J.W.; Kasajima, N.; Murakami, N. Anti-inflammatory properties of red ginger (Zingiber officinale var. Rubra) extract and suppression of nitric oxide production by its constituents. J. Med. Food 2010, 13, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Samad, M.B.; Mohsin, M.N.A.B.; Razu, B.A.; Hossain, M.T.; Mahzabeen, S.; Unnoor, N.; Muna, I.A.; Akhter, F.; Kabir, A.U.; Hannan, J.M.A. [6]-Gingerol, from Zingiber officinale, potentiates GLP-1 mediated glucose-stimulated insulin secretion pathway in pancreatic β-cells and increases RAB8/RAB10-regulated membrane presentation of GLUT4 transporters in skeletal muscle to improve hyperglycemia in Leprdb/db type 2 diabetic mice. BMC Complement. Altern. Med. 2017, 17, 395. [Google Scholar] [CrossRef]
- Zakaria-Runkat, F.; Prangdimurti, E. Antioxidant and immunoenhancement activities of ginger (Zingiber officinale Roscoe) extracts and compounds in vitro and in vivo mouse and human system. Nutraceuticals Foods 2003, 8, 96–104. [Google Scholar] [CrossRef]
- McCartney, F.N.; Allen, J.B.; Mizel, D.E.; Albina, J.E.; Xie, Q.W.; Nathan, C.F.; Wahl, S.M. Suppression of arthritis by an inhibitor of nitric oxide synthase. J. Exp. Med. 1993, 178, 749–754. [Google Scholar] [CrossRef]
- Shin, H.-Y.; Kim, H.; Shin, J.-Y.; Lee, S.J.; Yu, K.-W. The physiological activity of crude polysaccharide solvent extracted from herbal medicine mixture. Korean J. Food Nutr. 2021, 34, 36–46. [Google Scholar]
- Kim, H.W.; Shin, M.S.; Lee, S.J.; Park, H.R.; Jee, H.S.; Yoon, T.J.; Shin, K.S. Signaling pathways associated with macrophage-activating polysaccharides purified from fermented barley. Int. J. Biol. Macromol. 2019, 131, 1084–1091. [Google Scholar] [CrossRef]
- Kim, S.J.; Baek, S.H.; Kang, K.S.; Shin, M.S. Characterization of macrophage activation after treatment with polysaccharides from ginseng according to heat processing. Appl. Biol. Chem. 2023, 66, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Guo, C.; Li, X. Chitosan ameliorates DSS-induced ulcerative colitis mice by enhancing intestinal barrier function and improving microflora. Int. J. Mol. Sci. 2019, 20, 5751. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Connell, D. The chemistry of the essential oil and oleoresin of ginger (Zingiber officinale Roscoe). Flavour Ind. 1970, 1, 677–693. [Google Scholar]
- Okumura, R.; Takeda, K. Roles of intestinal epithelial cells in the maintenance of gut homeostasis. Exp. Mol. Med. 2017, 49, e338. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Deng, Z.; Hou, Y.; Zhang, G. Regulation of the intestinal barrier function by host defense peptides. Front. Vet. Sci. 2015, 2, 57. [Google Scholar] [CrossRef]
- El Kaoutari, A.E.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Microbiol. 2013, 11, 497–504. [Google Scholar] [CrossRef]
- Iraha, A.; Chinen, H.; Hokama, A.; Yonashiro, T.; Kinjo, T.; Kishimoto, K.; Nakamoto, M.; Hirata, T.; Kinjo, N.; Higa, F.; et al. Fucoidan enhances intestinal barrier function by upregulating the expression of claudin-1. World J. Gastroenterol. 2013, 19, 5500–5507. [Google Scholar] [CrossRef]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.; De Boeck, G.; Becker, K. Dietary roles of non-starch polysaccharides in human nutrition: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 899–935. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef]
- Pope, J.L.; Bhat, A.A.; Sharma, A.; Ahmad, R.; Krishnan, M.; Washington, M.K.; Beauchamp, R.D.; Singh, A.B.; Dhawan, P. Claudin-1 regulates intestinal epithelial homeostasis through the modulation of Notch-signalling. Gut 2014, 63, 622–634. [Google Scholar] [CrossRef]
- Hijova, E.; Chmelarova, A. Short chain fatty acids and colonic health. Bratisl. Lek. Listy 2007, 108, 354–358. [Google Scholar]
- Wong, J.M.; De Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Folin, O.; Ciocalteu, V. On tyrosine and tryptophane determinations in proteins. J. biol. Chem. 1927, 73, 627–650. [Google Scholar] [CrossRef]
- Kelly, P. Infectious diarrhoea. Medicine 2011, 39, 201–206. [Google Scholar] [CrossRef]
- Hanauer, S.B. The role of loperamide in gastrointestinal disorders. Rev. Gastroenterol. Disord. 2008, 8, 15–20. [Google Scholar] [PubMed]
- Cheung, R.P.; DiPiro, J.T. Vancomycin: An update. Pharmacotherapy 1986, 6, 153–169. [Google Scholar] [CrossRef]
- Hernández Ceruelos, A.; Romero-Quezada, L.C.; Ruvalcaba Ledezma, J.C.; López Contreras, L. Therapeutic uses of metronidazole and its side effects: An update. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Surawicz, C.M. Role of probiotics in antibiotic-associated diarrhea, Clostridium difficile-associated diarrhea, and recurrent Clostridium difficile-associated diarrhea. J. Clin. Gastroenterol. 2008, 42 (Suppl. S2), S64–S70. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Ishihara, K.; Morota, T.; Takeda, S.; Aburada, M. Permeability of the flavonoids liquiritigenin and its glycosides in licorice roots and davidigenin, a hydrogenated metabolite of liquiritigenin, using human intestinal cell line Caco-2. J. Ethnopharmacol. 2003, 89, 285–289. [Google Scholar] [CrossRef]
- Olivas-Méndez, P.; Chávez-Martínez, A.; Santellano-Estrada, E.; Guerrero Asorey, L.; Sánchez-Vega, R.; Rentería-Monterrubio, A.L.; Chávez-Flores, D.; Tirado-Gallegos, J.M.; Méndez-Zamora, G. Antioxidant and antimicrobial Activity of Rosemary (Rosmarinus officinalis) and Garlic (Allium sativum) Essential Oils and Chipotle Pepper Oleoresin (Capsicum annum) on Beef Hamburgers. Foods 2022, 11, 2018. [Google Scholar] [CrossRef]
- Xu, H.; Wang, S.; Jiang, Y.; Wu, J.; Chen, L.; Ding, Y.; Zhou, Y.; Deng, L.; Chen, X. Poria cocos polysaccharide ameliorated antibiotic-associated diarrhea in mice via regulating the homeostasis of the gut microbiota and intestinal mucosal barrier. Int. J. Mol. Sci. 2023, 24, 1423. [Google Scholar] [CrossRef]
- Han, K.; Kim, K.; Wang, J.; Kim, H. Effect of unfermented and fermented Atractylodes macrocephalae on gut permeability and lipopolysaccharide-induced inflammation. J. Soc. Korean Med. Obes. Res. 2013, 13, 24–32. [Google Scholar]
- Ridley, B.L.; O’Neill, M.A.; Mohnen, D. Pectins: Structure, biosynthesis, and oligogalacturonide-related signaling. Phytochemistry 2001, 57, 929–967. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, M.; Albersheim, P.; Darvill, A. The pectic polysaccharides of primary cell walls. In Carbohydrates; Elsevier: Amsterdam, The Netherlands, 1990; Volume 2, pp. 415–441. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Properties | Mole (%) |
| Neutral sugar | 86.4 ± 5.1 |
| Uronic acid | 8.8 ± 0.8 |
| Protein | 4.0 ± 0.1 |
| Polyphenol | 0.8 ± 0.2 |
| KDO-like material | not detected |
| Component Sugar | Mole (%) |
| Mannose | - |
| Rhamnose | - |
| Glucuronic acid | - |
| Galacturonic acid | - |
| Glucose | 97.3 ± 0.6 |
| Galactose | 2.7 ± 0.9 |
| Xylose | - |
| Arabinose | - |
| Fucose | - |
| Scores | Diarrhea Status |
|---|---|
| 0 | Normal (no diarrhea) |
| 1 | Loose, light colored and non-sticky perianal stool status; general mental state |
| 2 | Adhesion of stool at the anus; mental depression; no appetite for food; weight loss |
| Sample Name | Origin | Freeze-Dried Extract (g) | Yield (%) |
|---|---|---|---|
| Zingiber officinale Roscoe | Peru | 10.3 | 34.3 |
| Pump | Liquid chromatography LC-20AD (Shimadzu Corporation, Kyoto, Japan) |
| Detector | UV/VIS detector (Shimadzu Corporation, Kyoto, Japan) |
| Column | Acclaim TM 120 C18 (Thermo Scientific, Sunnyvale, CA, USA) |
| Column size | 4.6 × 250 mm |
| Column temp. | 30 °C |
| Flow rate | 1 mL/min |
| Eluent | 0.1 M sodium phosphate buffer (pH 6.7): acetonitrile (82:18) |
| Injection vol. | 10 μL |
| Integrator | Shimadzu data module (Shimadzu Corporation, Kyoto, Japan) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.J.; Shin, M.-S.; Choi, Y.-K. Ameliorative Effects of Zingiber officinale Rosc on Antibiotic-Associated Diarrhea and Improvement in Intestinal Function. Molecules 2024, 29, 732. https://doi.org/10.3390/molecules29030732
Kim SJ, Shin M-S, Choi Y-K. Ameliorative Effects of Zingiber officinale Rosc on Antibiotic-Associated Diarrhea and Improvement in Intestinal Function. Molecules. 2024; 29(3):732. https://doi.org/10.3390/molecules29030732
Chicago/Turabian StyleKim, Sung Jin, Myoung-Sook Shin, and You-Kyung Choi. 2024. "Ameliorative Effects of Zingiber officinale Rosc on Antibiotic-Associated Diarrhea and Improvement in Intestinal Function" Molecules 29, no. 3: 732. https://doi.org/10.3390/molecules29030732
APA StyleKim, S. J., Shin, M.-S., & Choi, Y.-K. (2024). Ameliorative Effects of Zingiber officinale Rosc on Antibiotic-Associated Diarrhea and Improvement in Intestinal Function. Molecules, 29(3), 732. https://doi.org/10.3390/molecules29030732