


Dynamic Changes and Potential Correlations between Microbial Diversity and Volatile Flavor Compounds in Chinese Medium-Temperature Daqu during Manufacturing
 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
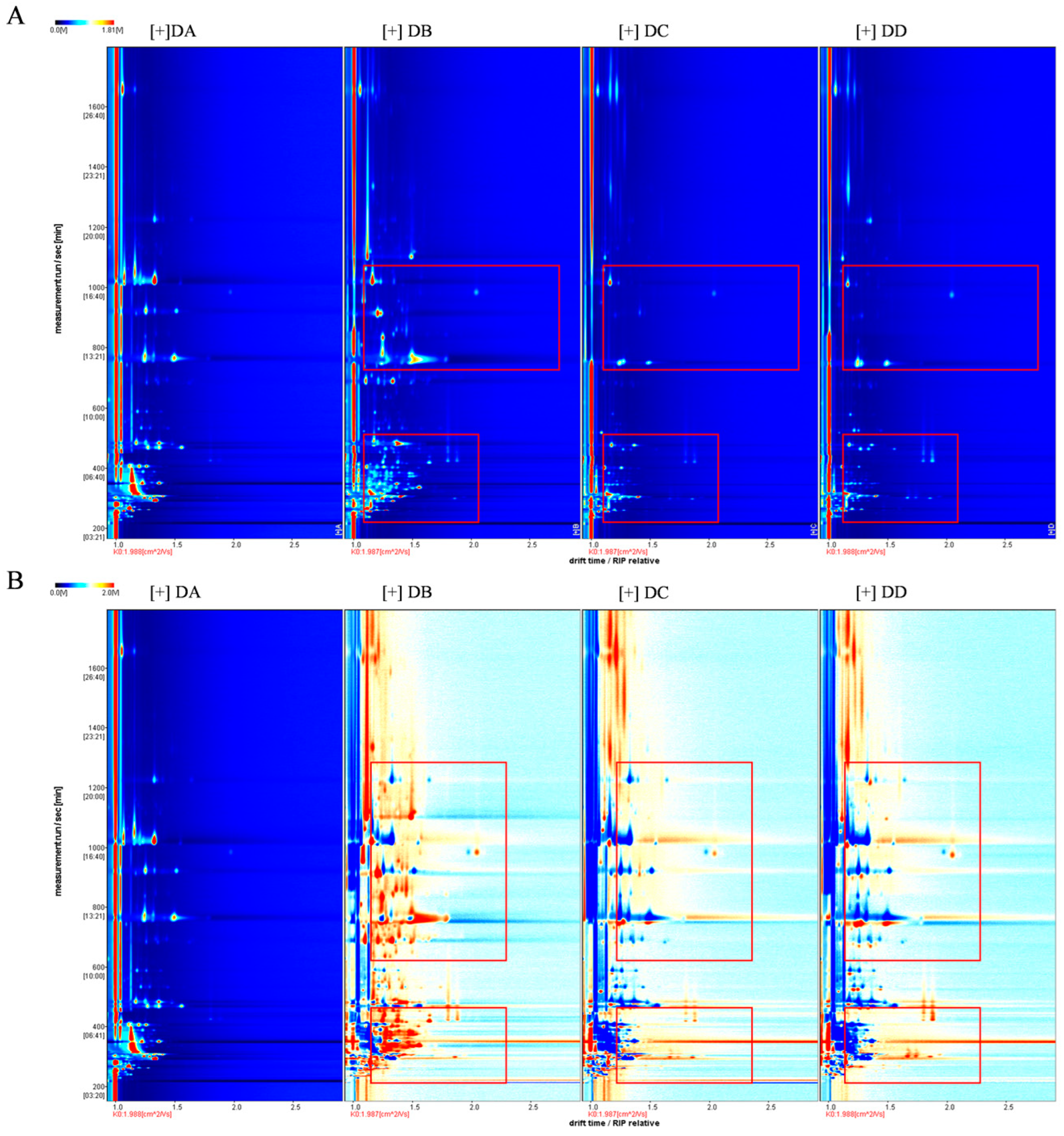
2.1. Qualitative Analysis of the VOCs in MTD
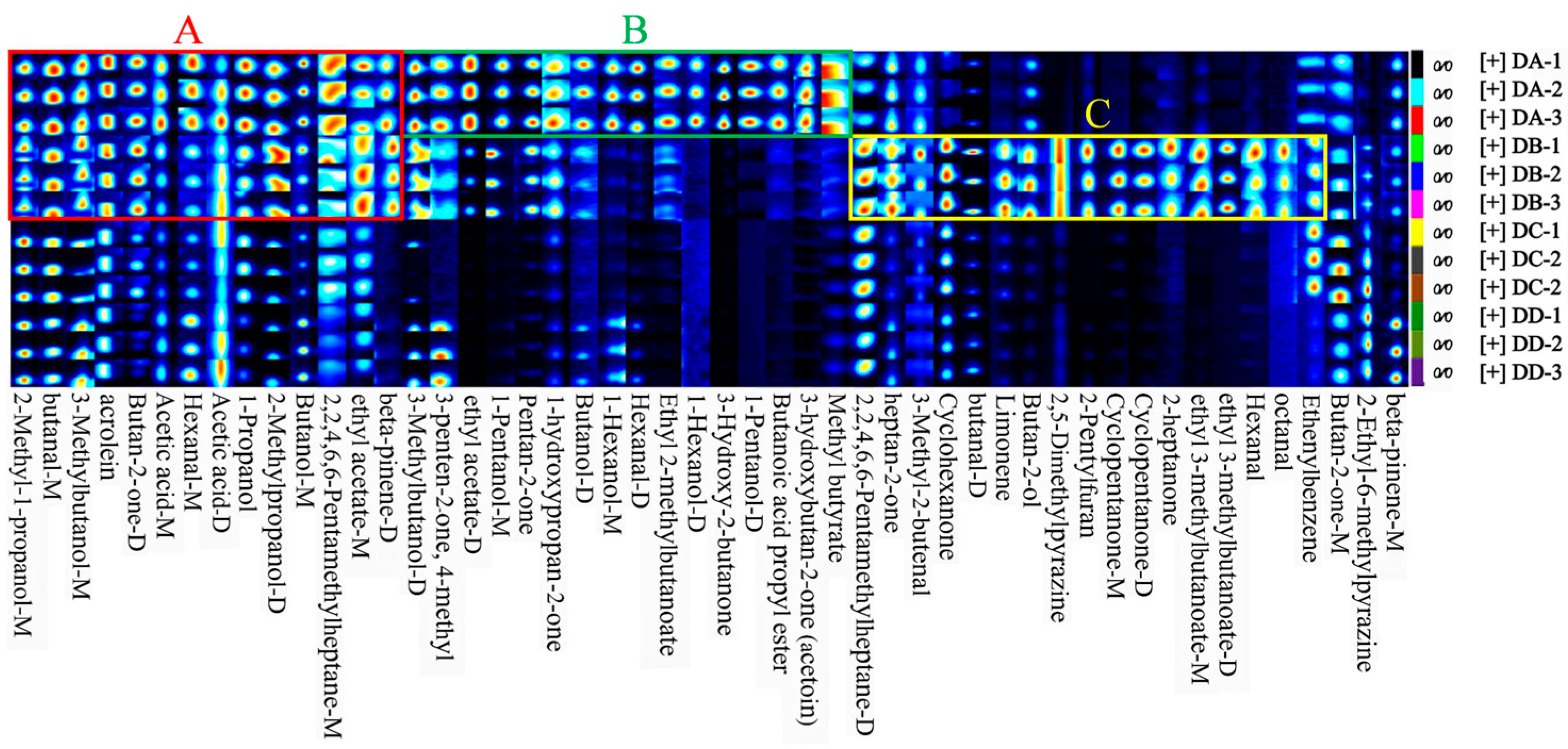
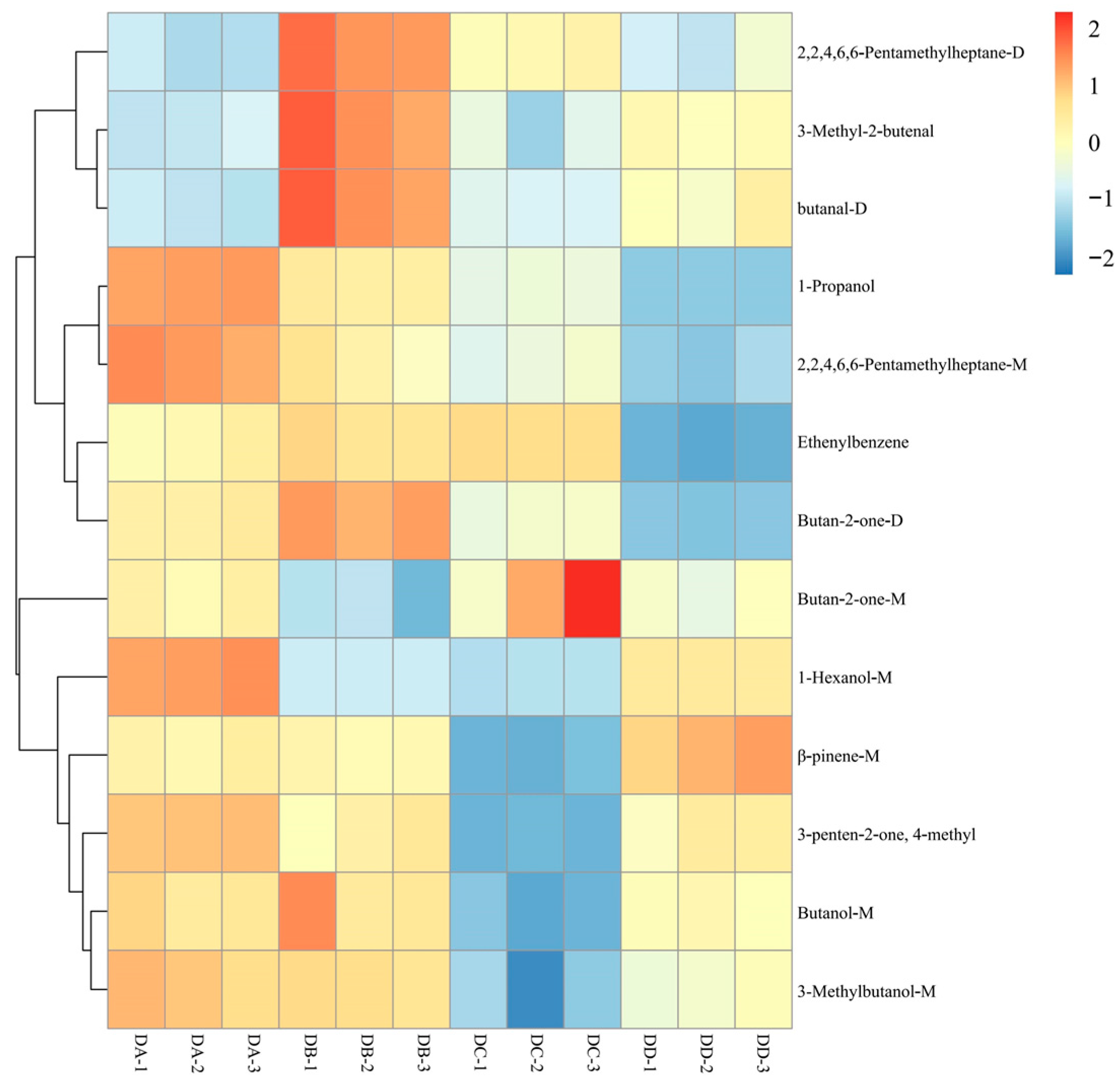
2.2. Fingerprint Analysis of VOCs
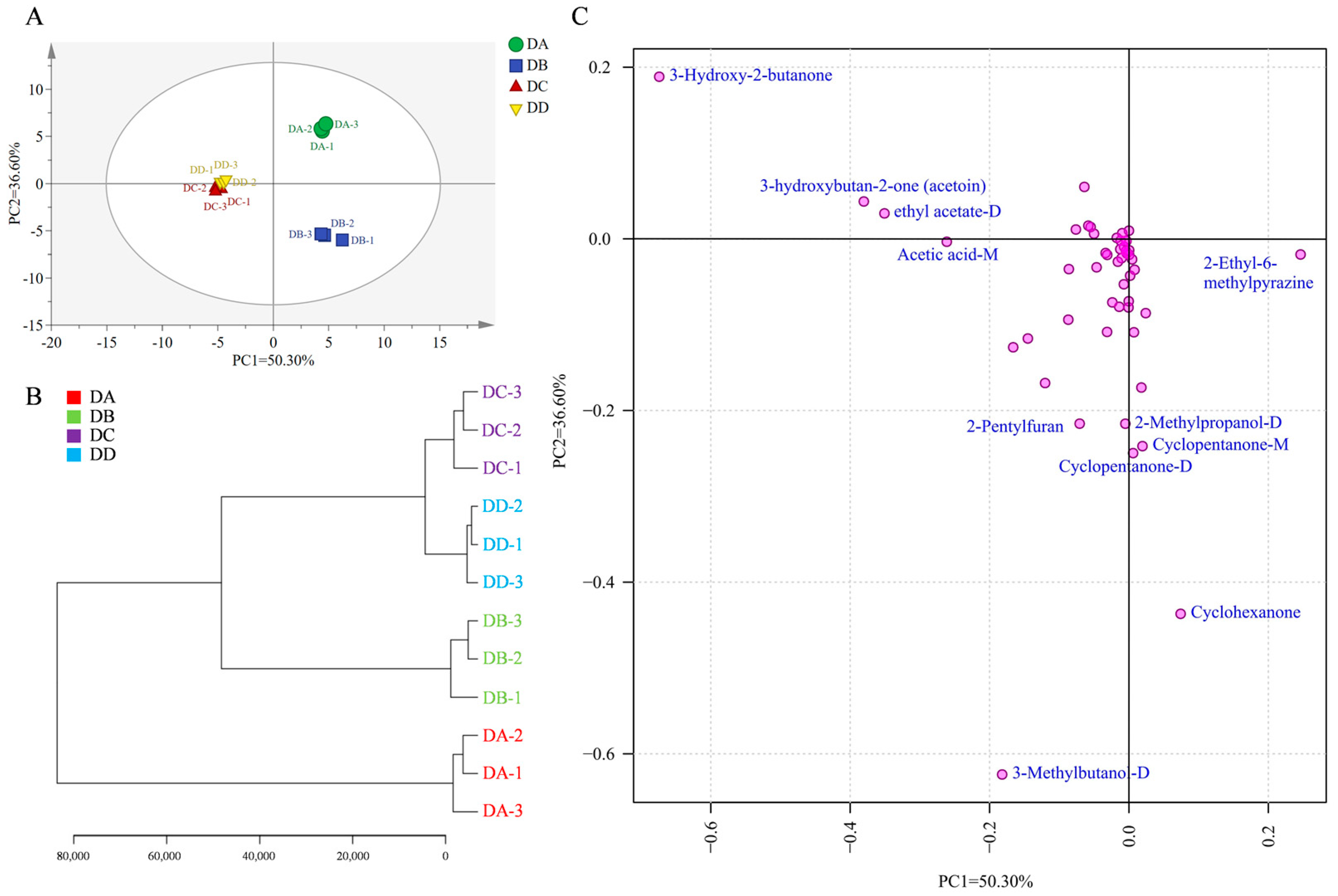
2.3. Multivariate Statistical Analysis of the VOCs
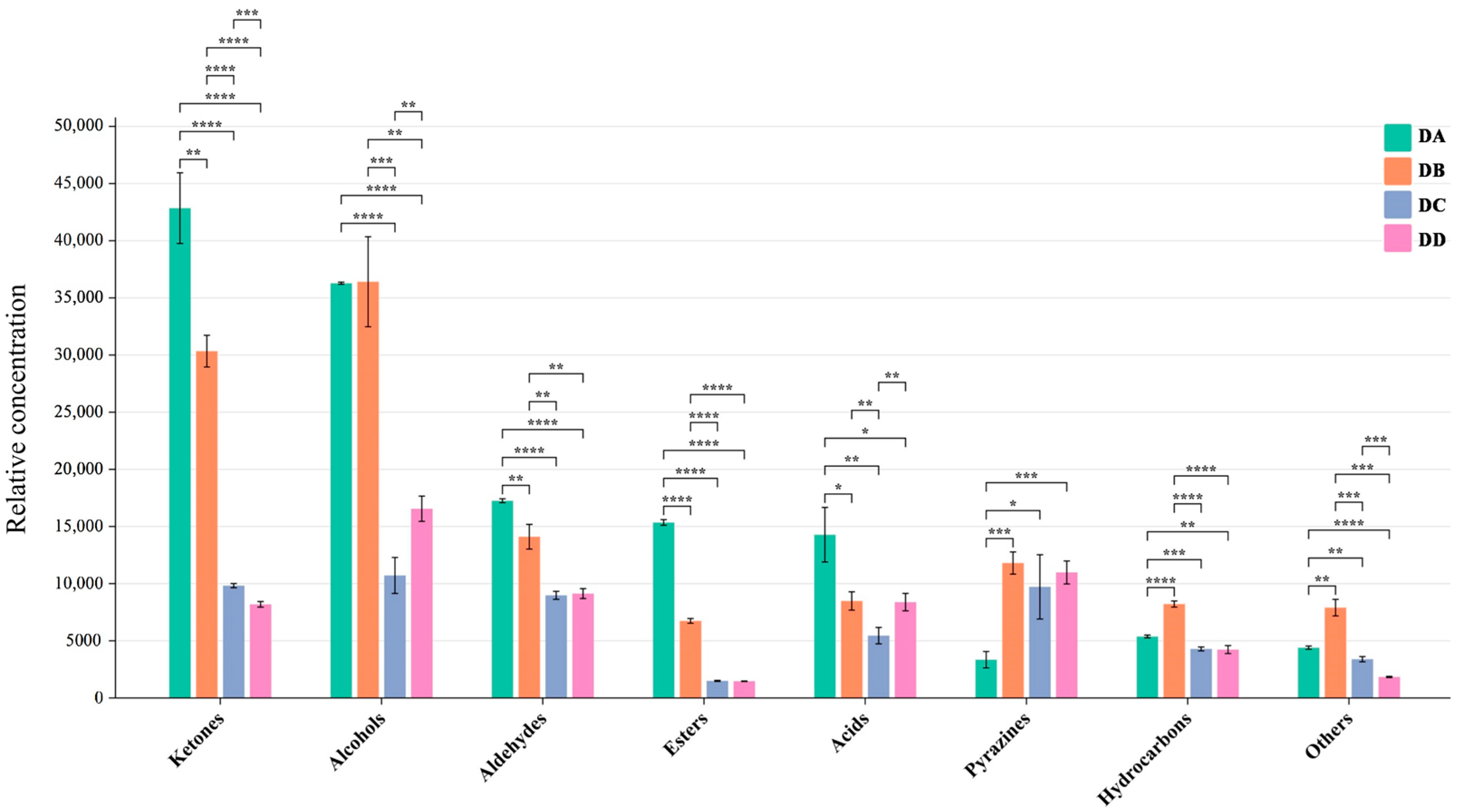
2.4. Analysis of the Differential VOCs in MTD
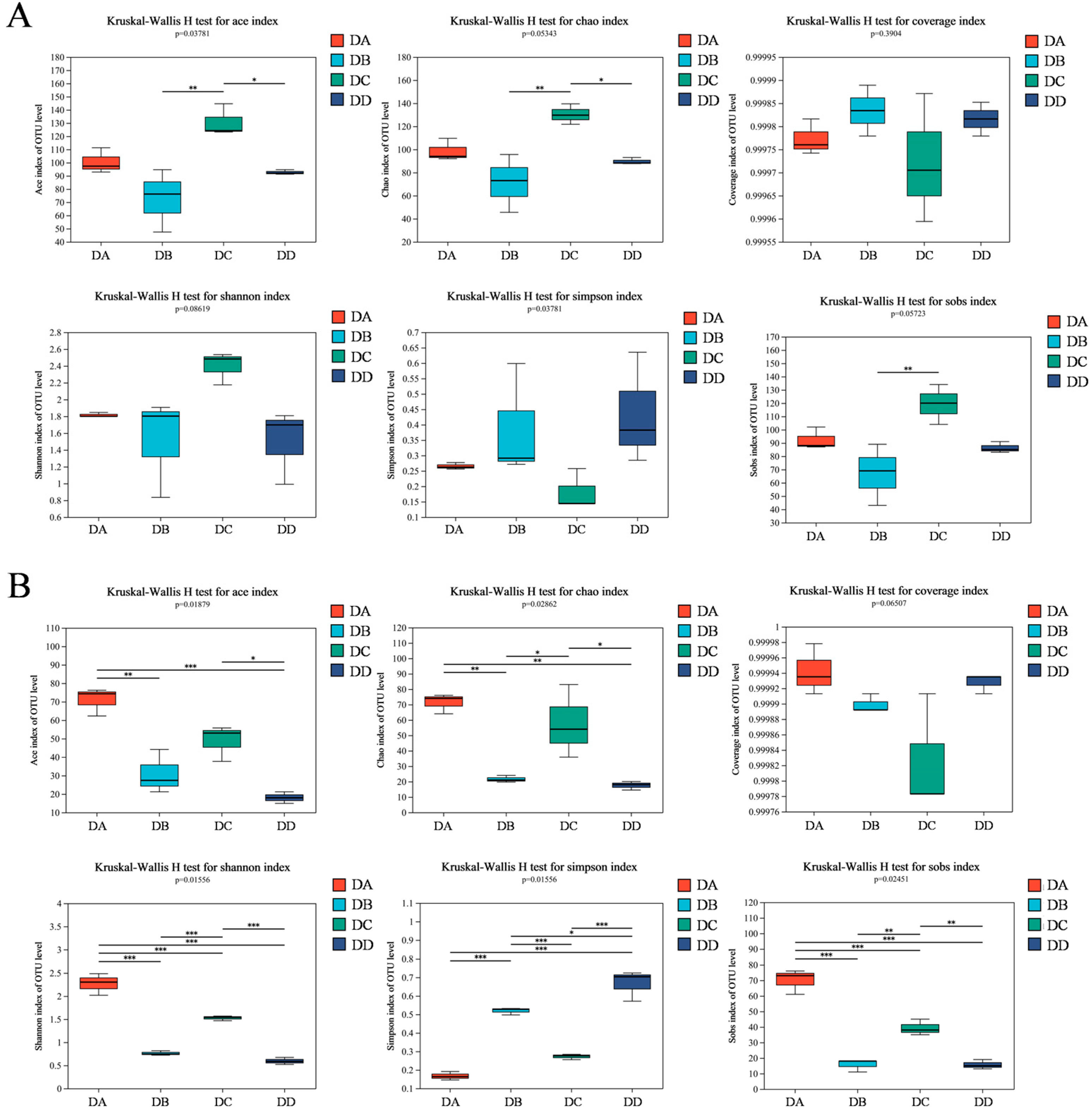
2.5. α-Diversity Analysis of MTD Based on HTS
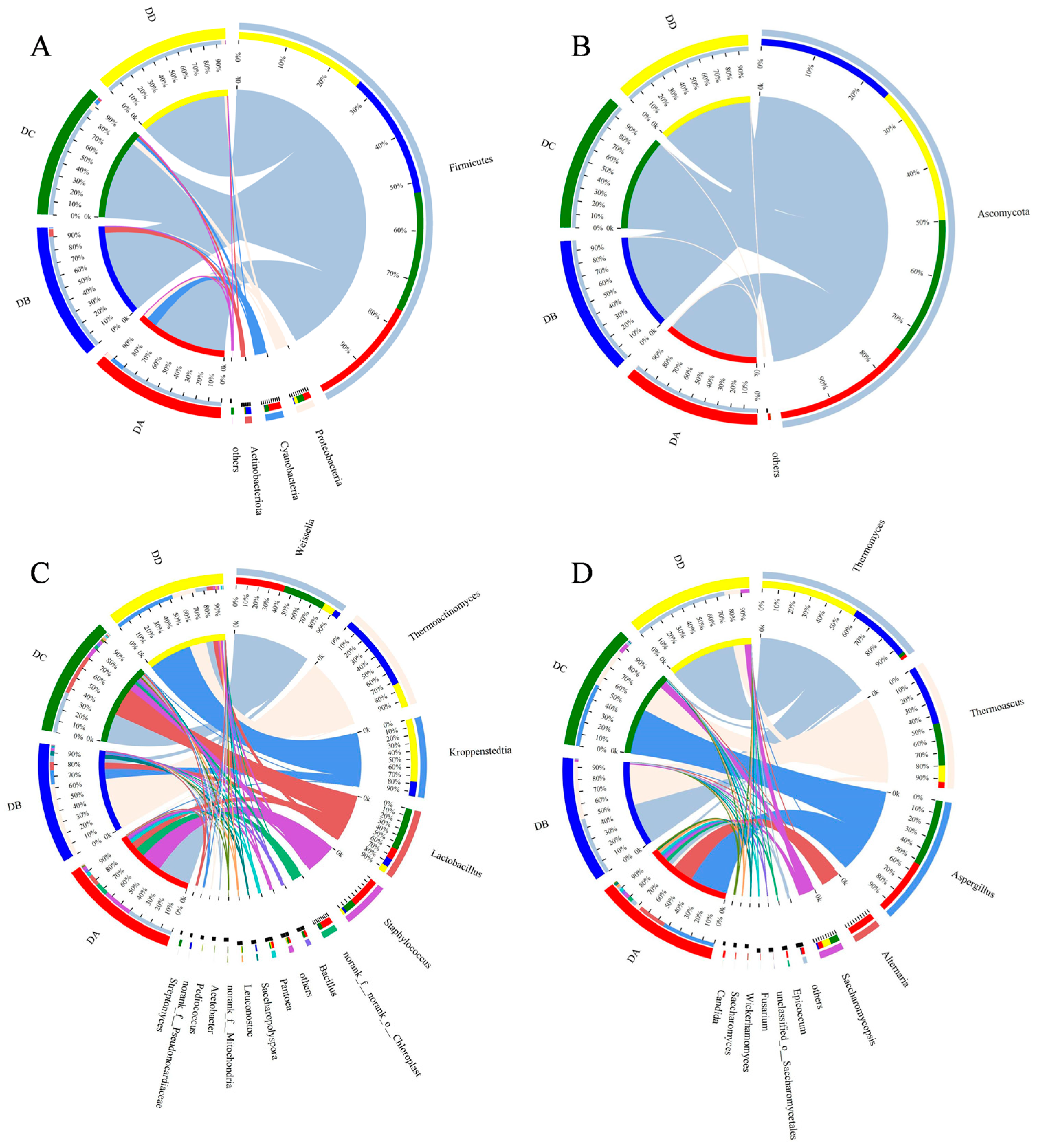
2.6. Analysis of Microbial Community Structure in MTD
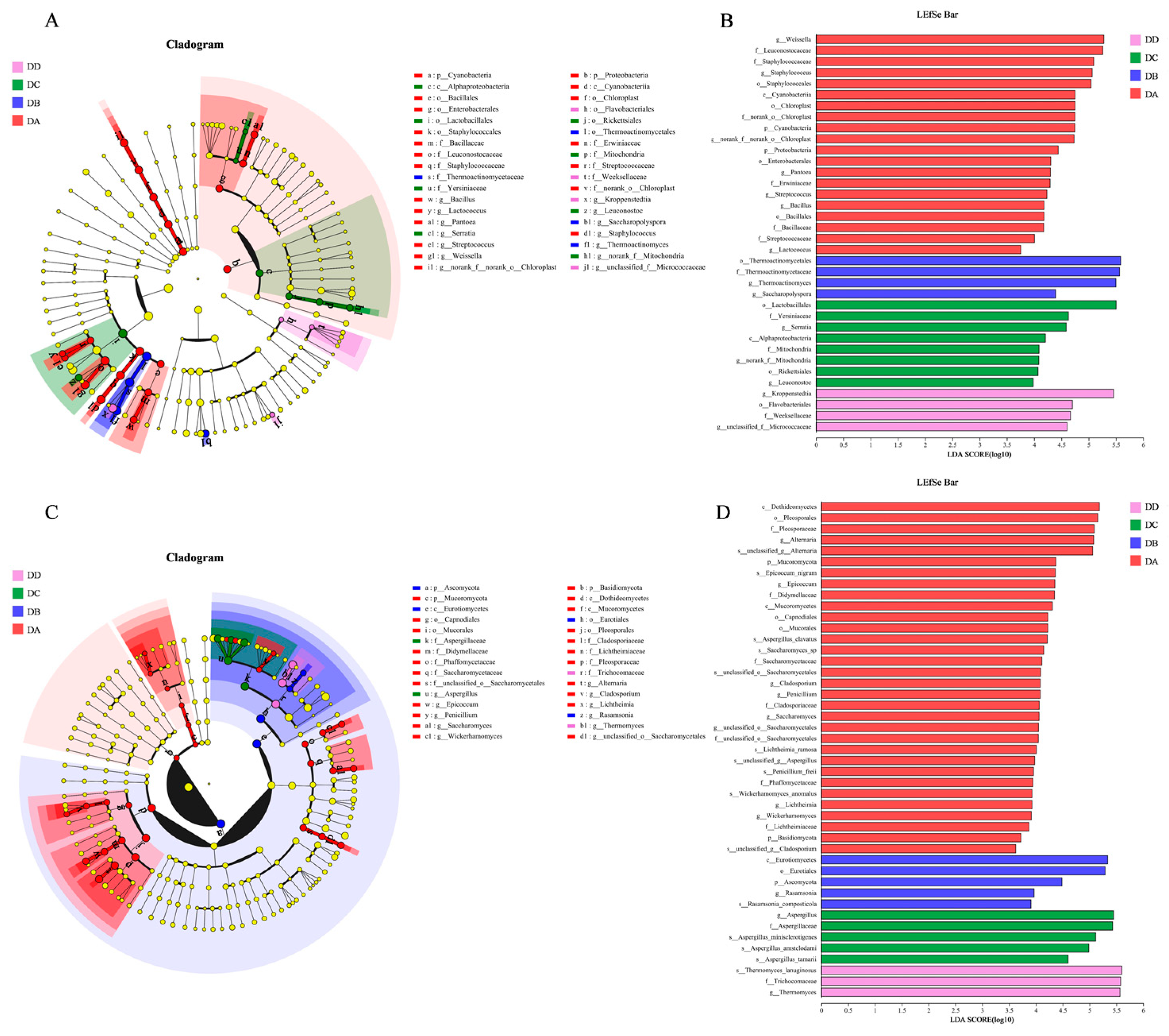
2.7. Analysis of LEfSe Differences in Microorganisms
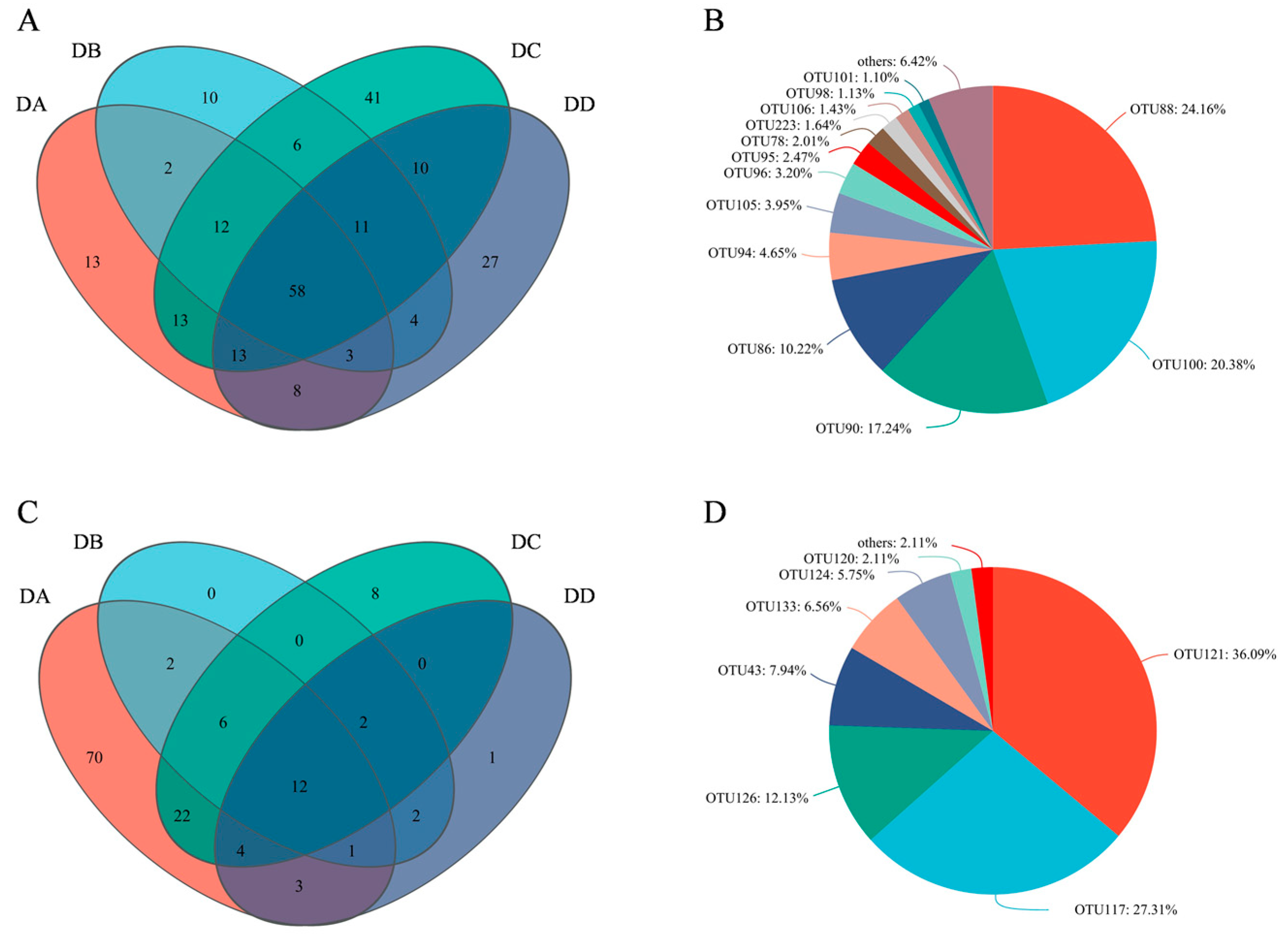
2.8. Core Flora Analysis Based on OTU Levels
2.9. Potential Correlations between Microbial Community and VOCs
3. Materials and Methods
3.1. Collection of MTD
3.2. Detection of VOCs in MTD by GCIMS
3.3. DNA Extraction and Sequencing
3.4. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Cai, W.; Wang, W.; Shu, N.; Zhang, Z.; Hou, Q.; Shan, C.; Guo, Z. Analysis of microbial diversity and functional differences in different types of high-temperature Daqu. Food Sci. Nutr. 2021, 9, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Jing, R.; Chen, W.; Geng, X.; Li, M.; Yang, F.; Yan, Y.; Liu, Y. High-throughput sequencing of the microbial diversity of roasted-sesame-like flavored Daqu with different characteristics. 3 Biotech 2020, 10, 502. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, H.; Dong, S.; Zhou, Z.; Zhang, Z.; Huang, R.; Han, S.; Hou, J.; Pan, C. Dynamic changes and correlations of microbial communities, physicochemical properties, and volatile metabolites during Daqu fermentation of Taorong-type Baijiu. LWT 2023, 173, 114290. [Google Scholar] [CrossRef]
- Deng, J.; Zheng, J.; Huang, D.; Huang, Z.; Ye, G.; Luo, H. Characterization of physicochemical properties, volatile compounds and microbial community structure in four types of Daqu. LWT 2023, 184, 115064. [Google Scholar] [CrossRef]
- Yan, S.; Tong, Q.; Guang, J. Yeast dynamics and changes in volatile compounds during the fermentation of the traditional Chinese strong-flavor Daqu. LWT 2019, 106, 57–63. [Google Scholar] [CrossRef]
- Zhu, C.; Cheng, Y.; Shi, Q.; Ge, X.; Yang, Y.; Huang, Y. Metagenomic analyses reveal microbial communities and functional differences between Daqu from seven provinces. Food Res. Int. 2023, 172, 113076. [Google Scholar] [CrossRef]
- Kang, J.; Chen, X.; Han, B.-Z.; Xue, Y. Insights into the bacterial, fungal, and phage communities and volatile profiles in different types of Daqu. Food Res. Int. 2022, 158, 111488. [Google Scholar] [CrossRef]
- Ma, S.; Shang, Z.; Chen, J.; Shen, Y.; Li, Z.; Huang, D.; Luo, H. Differences in structure, volatile metabolites, and functions of microbial communities in Nongxiangxing daqu from different production areas. LWT 2022, 166, 113784. [Google Scholar] [CrossRef]
- Sidira, M.; Kandylis, P.; Kanellaki, M.; Kourkoutas, Y. Effect of immobilized Lactobacillus casei on the evolution of flavor compounds in probiotic dry-fermented sausages during ripening. Meat Sci. 2015, 100, 41–51. [Google Scholar] [CrossRef]
- Zhu, S.; Lu, X.; Ji, K.; Guo, K.; Li, Y.; Wu, C.; Xu, G. Characterization of flavor compounds in Chinese liquor Moutai by comprehensive two-dimensional gas chromatography/time-of-flight mass spectrometry. Anal. Chim. Acta 2007, 597, 340–348. [Google Scholar] [CrossRef]
- Yang, L.; Fan, W.; Xu, Y. GC × GC-TOF/MS and UPLC-Q-TOF/MS based untargeted metabolomics coupled with physicochemical properties to reveal the characteristics of different type daqus for making soy sauce aroma and flavor type baijiu. LWT 2021, 146, 111416. [Google Scholar] [CrossRef]
- Liu, W.-H.; Chai, L.-J.; Wang, H.-M.; Lu, Z.-M.; Zhang, X.-J.; Xiao, C.; Wang, S.-T.; Shen, C.-H.; Shi, J.-S.; Xu, Z.-H. Bacteria and filamentous fungi running a relay race in Daqu fermentation enable macromolecular degradation and flavor substance formation. Int. J. Food Microbiol. 2023, 390, 110118. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, Z.; Qian, M.; Yu, X.; Xu, Y.; Chen, S. Unraveling the chemosensory characteristics of strong-aroma type Baijiu from different regions using comprehensive two-dimensional gas chromatography–time-of-flight mass spectrometry and descriptive sensory analysis. Food Chem. 2020, 331, 127335. [Google Scholar] [CrossRef]
- Song, X.; Jing, S.; Zhu, L.; Ma, C.; Song, T.; Wu, J.; Zhao, Q.; Zheng, F.; Zhao, M.; Chen, F. Untargeted and targeted metabolomics strategy for the classification of strong aroma-type baijiu (liquor) according to geographical origin using comprehensive two-dimensional gas chromatography-time-of-flight mass spectrometry. Food Chem. 2020, 314, 126098. [Google Scholar] [CrossRef]
- Wu, S.; Du, H.; Xu, Y. Daqu microbiota adaptability to altered temperature determines the formation of characteristic compounds. Int. J. Food Microbiol. 2023, 385, 109995. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Deng, M.; Chen, K.; Chen, Y.; Cai, W.; Wu, C.; Liu, C.; Wu, S.; Wan, Y. Peak-temperature effects of starter culture (Daqu) on microbial community succession and volatile substances in solid-state fermentation (Jiupei) during traditional Chinese special-flavour Baijiu production. LWT 2021, 152, 112132. [Google Scholar] [CrossRef]
- Fan, W.; Qian, M. Identification of aroma compounds in Chinese ‘Yanghe Daqu’ liquor by normal phase chromatography fractionation followed by gas chromatography olfactometry. Flavour Fragr. J. 2006, 21, 333–342. [Google Scholar] [CrossRef]
- Killian, E.; Ough, C.S. Fermentation Esters—Formation and Retention as Affected by Fermentation Temperature. Am. J. Enol. Vitic. 1979, 30, 301–305. [Google Scholar] [CrossRef]
- Wang, P.; Wu, Q.; Jiang, X.; Wang, Z.; Tang, J.; Xu, Y. Bacillus licheniformis affects the microbial community and metabolic profile in the spontaneous fermentation of Daqu starter for Chinese liquor making. Int. J. Food Microbiol. 2017, 250, 59–67. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, R.; Yang, F.; Xie, Y.; Guo, Y.; Yao, W.; Zhou, W. Control strategies of pyrazines generation from Maillard reaction. Trends Food Sci. Technol. 2021, 112, 795–807. [Google Scholar] [CrossRef]
- Wang, Y.; Quan, S.; Zhao, Y.; Xia, Y.; Zhang, R.; Ran, M.; Wu, Z.; Zhang, W. The active synergetic microbiota with Aspergillus as the core dominates the metabolic network of ester synthesis in medium-high temperature Daqu. Food Microbiol. 2023, 115, 104336. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhao, J.; Liu, X.; Zhang, C.; Zhao, Z.; Li, X.; Sun, B. Flavor mystery of Chinese traditional fermented baijiu: The great contribution of ester compounds. Food Chem. 2022, 369, 130920. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Peng, Z.; Zhu, Q.; Zheng, T.; Liu, X.; Yang, J.; Zhang, J.; Li, J. Exploration of seasonal fermentation differences and the possibility of flavor substances as regulatory factors in Daqu. Food Res. Int. 2023, 168, 112686. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, G.; Yao, Y.; Zhu, W.; Xu, S.; Li, H. Research on the aroma properties and microbial succession patterns in the processing of Chinese yellow sticky rice jiuqu steamed bread. LWT 2023, 180, 114704. [Google Scholar] [CrossRef]
- Qiao, L.; Wang, J.; Wang, R.; Zhang, N.; Zheng, F. A review on flavor of Baijiu and other world-renowned distilled liquors. Food Chem. X 2023, 20, 100870. [Google Scholar] [CrossRef]
- Starowicz, M.; Zieliński, H. How Maillard Reaction Influences Sensorial Properties (Color, Flavor and Texture) of Food Products? Food Rev. Int. 2019, 35, 707–725. [Google Scholar] [CrossRef]
- Zhang, Y.; Tong, X.; Chen, B.; Wu, S.; Wang, X.; Zheng, Q.; Jiang, F.; Qiao, Y. Novel application of HS-GC-IMS for characteristic fingerprints and flavor compound variations in citrus reticulatae pericarpium during storage with different Aspergillus niger fermentation. Food Chem. X 2023, 18, 100653. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Gao, Y.; Xu, Y.; Zhou, H.; Zhou, K.; Li, C.; Xu, B. Microbiota dynamics and volatile metabolite generation during sausage fermentation. Food Chem. 2023, 423, 136297. [Google Scholar] [CrossRef]
- Wang, Z.; Shi, Y.; Zhou, K.; Zhou, H.; Li, X.; Li, C.; Wang, Z.; Xu, B. Effects of different thermal temperatures on the shelf life and microbial diversity of Dezhou-braised chicken. Food Res. Int. 2020, 136, 109471. [Google Scholar] [CrossRef]
- Tang, J.; Rao, J.; Zou, Y.; Liao, L.; Huang, D.; Luo, H. The community assembly patterns determined differences between the surface and the core microbial communities of Nongxiangxing Daqu. LWT 2023, 183, 114936. [Google Scholar] [CrossRef]
- Zhu, Q.; Chen, L.; Peng, Z.; Zhang, Q.; Huang, W.; Yang, F.; Du, G.; Zhang, J.; Wang, L. Analysis of environmental driving factors on Core Functional Community during Daqu fermentation. Food Res. Int. 2022, 157, 111286. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Mao, X.; Liu, D.; Ning, X.Q.; Shen, Y.; Chen, B.; Nie, H.F.; Huang, D.; Luo, H.B. Comparative Analysis of Physicochemical Properties and Microbial Composition in High-Temperature Daqu With Different Colors. Front. Microbiol. 2020, 11, 588117. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Zhong, H.; Zhao, D.; Du, H.; Xu, Y. Succession rate of microbial community causes flavor difference in strong-aroma Baijiu making process. Int. J. Food Microbiol. 2019, 311, 108350. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Yang, Y.; Xu, Y.; Zhang, J.; Lu, Y. Balance between community assembly processes mediates species coexistence in agricultural soil microbiomes across eastern China. ISME J. 2020, 14, 202–216. [Google Scholar] [CrossRef]
- Quan, S.; Wang, Y.; Ran, M.; Zhang, R.; Luo, X.; Wang, W.; Wu, Z.; Gomi, K.; Zhang, W. Contrasting the microbial community and non-volatile metabolites involved in ester synthesis between Qing-flavor Daqu and Nong-flavor Daqu. J. Biosci. Bioeng. 2023, 136, 213–222. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, Q.; Xu, Y. Isolation and identification of a black Aspergillus strain and the effect of its novel protease on the aroma of Moutai-flavoured liquor. J. Inst. Brew. 2014, 120. [Google Scholar] [CrossRef]
- Hou, X.; Hui, M.; Sun, Z.; Li, X.; Shi, X.; Xiao, R.; Wang, J.; Pan, L.; Pan, C.; Li, R. Associations between wheat varieties and the functional microbial community of Nongxiangxing daqu from northern China. LWT 2023, 185, 115119. [Google Scholar] [CrossRef]
- Hou, Q.; Wang, Y.; Cai, W.; Ni, H.; Zhao, H.; Zhang, Z.; Liu, Z.; Liu, J.; Zhong, J.a.; Guo, Z. Metagenomic and physicochemical analyses reveal microbial community and functional differences between three types of low-temperature Daqu. Food Res. Int. 2022, 156, 111167. [Google Scholar] [CrossRef]
- Zheng, Y.; Liang, F.; Wu, Y.; Ban, S.; Huang, H.; Xu, Y.; Wang, X.; Wu, Q. Unraveling multifunction of low-temperature Daqu in simultaneous saccharification and fermentation of Chinese light aroma type liquor. Int. J. Food Microbiol. 2023, 397, 110202. [Google Scholar] [CrossRef]
- Wu, Q.; Zhu, Y.; Fang, C.; Wijffels, R.H.; Xu, Y. Can we control microbiota in spontaneous food fermentation?—Chinese liquor as a case example. Trends Food Sci. Technol. 2021, 110, 321–331. [Google Scholar] [CrossRef]
- Zhu, C.; Cheng, Y.; Zuo, Q.; Huang, Y.; Wang, L. Exploring the impacts of traditional crafts on microbial community succession in Jiang-flavored Daqu. Food Res. Int. 2022, 158, 111568. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zou, Y.; Zeng, K.; Chen, D.; Li, Z.; Guo, H.; Huang, D.; Wang, X.; Luo, H. Effect of Bacillus subtilis fortified inoculation on the microbial communities in different niches of Daqu. J. Biosci. Bioeng. 2022, 134, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Oleinikova, Y.; Alybayeva, A.; Daugaliyeva, S.; Alimzhanova, M.; Ashimuly, K.; Yermekbay, Z.; Khadzhibayeva, I.; Saubenova, M. Development of an antagonistic active beverage based on a starter including Acetobacter and assessment of its volatile profile. Int. Dairy J. 2024, 148, 105789. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Ding, F.; Deng, W.; Wang, X.; Xue, Y.; Chen, X.; Han, B.-Z. Multidimensional profiling indicates the shifts and functionality of wheat-origin microbiota during high-temperature Daqu incubation. Food Res. Int. 2022, 156, 111191. [Google Scholar] [CrossRef]
- Nie, Z.; Zheng, Y.; Du, H.; Xie, S.; Wang, M. Dynamics and diversity of microbial community succession in traditional fermentation of Shanxi aged vinegar. Food Microbiol. 2015, 47, 62–68. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Wang, H.; Yang, F.; Chen, L.; Hao, F.; Lv, X.; Du, H.; Xu, Y. Effects of initial temperature on microbial community succession rate and volatile flavors during Baijiu fermentation process. Food Res. Int. 2021, 141, 109887. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, Z.; Gu, Z.; Deng, X.; Liu, J.; Luo, X.; Song, C.; Jiang, X. Fermentation-promoting effect of three salt-tolerant Staphylococcus and their co-fermentation flavor characteristics with Zygosaccharomyces rouxii in soy sauce brewing. Food Chem. 2024, 432, 137245. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAS | Flavor Substance | VIP | Aroma Descriptions | |
|---|---|---|---|---|
| 1 | 127-91-3 | β-pinene-M | 1.46287 | Pine, Wood |
| 2 | 287399-41-1 | Ethenylbenzene | 1.38991 | -- |
| 3 | 141-79-7 | 3-Penten-2-one, 4-methyl | 1.19342 | Honey |
| 4 | 71-36-3 | Butanol-M | 1.12958 | Fruit |
| 5 | 111-27-3 | 1-Hexanol-M | 1.1228 | Banana, Flower, Grass |
| 6 | 78-93-3 | Butan-2-one-M | 1.06204 | Fragrant, Fruit, Pleasant |
| 7 | 78-93-3 | Butan-2-one-D | 1.05515 | Fragrant, Fruit, Pleasant |
| 8 | 123-51-3 | 3-Methylbutanol-M | 1.05106 | Burnt, Brandy |
| 9 | 107-86-8 | 3-Methyl-2-butenal | 1.04748 | Almond, Roasted |
| 10 | 71-23-8 | 1-Propanol | 1.04289 | Alcohol, Candy, Pungent |
| 11 | 123-72-8 | Butanal-D | 1.04221 | Banana, Green, Pungent |
| 12 | 13475-82-6 | 2,2,4,6,6-Pentamethylheptane-M | 1.03767 | -- |
| 13 | 13475-82-6 | 2,2,4,6,6-Pentamethylheptane-D | 1.01351 | -- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, X.; Jia, X.; Zhu, K.; Ling, Z.; Chen, H.; Xie, J.; Ao, Z.; Song, C.; Shen, C.; Zhu, C.; et al. Dynamic Changes and Potential Correlations between Microbial Diversity and Volatile Flavor Compounds in Chinese Medium-Temperature Daqu during Manufacturing. Molecules 2024, 29, 4851. https://doi.org/10.3390/molecules29204851
Nie X, Jia X, Zhu K, Ling Z, Chen H, Xie J, Ao Z, Song C, Shen C, Zhu C, et al. Dynamic Changes and Potential Correlations between Microbial Diversity and Volatile Flavor Compounds in Chinese Medium-Temperature Daqu during Manufacturing. Molecules. 2024; 29(20):4851. https://doi.org/10.3390/molecules29204851
Chicago/Turabian StyleNie, Xin, Xiaohan Jia, Kaixian Zhu, Ziqing Ling, Hongfan Chen, Jing Xie, Zonghua Ao, Chuan Song, Caihong Shen, Chenglin Zhu, and et al. 2024. "Dynamic Changes and Potential Correlations between Microbial Diversity and Volatile Flavor Compounds in Chinese Medium-Temperature Daqu during Manufacturing" Molecules 29, no. 20: 4851. https://doi.org/10.3390/molecules29204851
APA StyleNie, X., Jia, X., Zhu, K., Ling, Z., Chen, H., Xie, J., Ao, Z., Song, C., Shen, C., Zhu, C., Yan, W., Wang, J., Wang, Y., & Zhao, Z. (2024). Dynamic Changes and Potential Correlations between Microbial Diversity and Volatile Flavor Compounds in Chinese Medium-Temperature Daqu during Manufacturing. Molecules, 29(20), 4851. https://doi.org/10.3390/molecules29204851