1. Introduction
The meadow moth is a major migratory pest in China and even the whole northern hemisphere. Diapause is a defensive measure evolved by the meadow moth to adapt to the cold climate in northern China during its migration [
1]. It is of great significance to study the diapause mechanism of the meadow moth for scientific prevention and control of the occurrence and damage of meadow moth. Previous studies have shown that the entry of insects into diapause is not only the closure of gene expression [
2], but also involves the complex regulation of metabolites and proteins in insect cells. The rapid and comprehensive progress of proteomics and metabonomics sequencing technology has been widely used to study various biological functions and processes of insects. In this study, we sequenced the non-diapause period, pre-diapause period, diapause period, cold treatment period, and post-diapause period of meadow moth by proteomics and metabonomics, in order to further understand the regulation mechanism of meadow moth diapause.
Diapause is an adaptive strategy employed by insects to enhance survival capabilities in response to adverse environmental conditions and seasonal changes. The meadow moth (
Loxostege sticticalis) over-winters through diapause, with larvae entering diapause under short-day conditions in autumn. They survive the winter at temperatures as low as −20 °C and terminate diapause in the following spring, with the diapause period lasting up to six months. The meadow moth is a long-day developmental-type insect, where under long-day conditions (≥16 h), larvae almost completely avoid diapause; however, under short-day conditions of 15 h, some larvae enter diapause, and the number of diapausing larvae increases as the photoperiod shortens [
3]. Since the diapause stage of the meadow moth is spent encased in a cocoon and buried in the soil, it is widely believed that the photoperiod experienced during the late developmental stages is the primary light factor influencing diapause in this species [
4].
Histone acetylation is a type of histone modification that can function independently of genomic changes at any stage of insect development, and may therefore play a role in regulating changes in gene expression necessary for diapause initiation, maintenance, and termination [
5]. In the cell, the helical chromatin structure is formed by packaging DNA into nucleosomes composed of histones H2a, H2b, H3, and H4. Histones H3 and H4 possess a tail that can be covalently modified to increase or decrease interactions between the DNA chain and nucleosomes, thereby altering transcriptional activity at specific DNA loci [
6]. Increasing the interaction between DNA and histones typically results in a compact chromatin structure, while reducing the interaction between histones and DNA creates regions of relaxed chromatin with helicase activity. Evidence that histone acetylation affects diapause transcription comes from studies on the southern ground cricket Allonemobius socius [
7], the northern house mosquito Culex pipiens [
8], and the flesh fly Sarcophaga bullata [
9]. Cui et al. conducted transcriptome analysis of diapause-regulating genes in the fall armyworm, finding that histone deacetylases play a significant role in regulating diapause [
10].
Ribosomes, fundamental sites of protein synthesis in eukaryotes, comprise 60s and 40s subunits, which are assembled in the nucleus and independently transported out of the nucleus. The intricate biogenesis of ribosomes plays a critical role in regulating biological development at multiple levels, including translational regulation in multicellular organisms. Xu discovered that histone deacetylases in rice are involved in the deacetylation of ribosomal proteins, potentially affecting ribosomal stability and translational efficiency [
11]. Urwanisch found that histone deacetylases can down-regulate ribosomal proteins [
12].
Previous studies have investigated the metabolomics of diapause in various insects: in diapausing wheat blossom midge (
Sitodiplosis mosellana), significant increases were observed in trehalose and glycerol compared to non-diapausing individuals [
13]. In diapausing Asian tiger mosquito Aedes albopictus, 84.7% of diapause-specific metabolites were categorized as “lipids and lipid-like molecules” [
14]. These metabolites, showing significant differences between diapause and non-diapause stages in insects, likely play roles in energy provision and cold resistance during diapause.
This study utilized proteomics and metabolomics techniques to identify and investigate changes in protein and metabolite levels during different diapause stages of the oriental armyworm, providing foundational research data on proteins and metabolites associated with diapause. This aims to better our understanding of the molecular regulatory mechanisms underlying diapause formation in the oriental armyworm.
3. Discussion
As an important agricultural pest in China, studying its diapause is of great significance for scientific prevention and control of meadow moth outbreaks. During diapause, various changes occur in neuroendocrine, molecular, cellular, enzymatic, metabolic, endocrine, and behavioral aspects [
18]. Diapause typically occurs at specific developmental stages of insects, namely embryo, larva/nymph, pupa, and adult [
19]. It is divided into three phases: pre-diapause, diapause, and post-diapause. The pre-diapause phase involves the induction and preparation for diapause, encompassing the initiation, maintenance, and termination stages, while the post-diapause phase is characterized by a state of quiescence and rapid development [
20,
21]. Typically, since diapause occurs at life cycle stages that resist unfavorable developmental environments, the intensity of diapause varies significantly among different insects; the duration of diapause can serve as a measure of its intensity. During diapause, most insects do not feed or, in some cases of larvae and adults, feed minimally. This indicates that insects must have sufficient food reserves before diapause to meet their metabolic needs during diapause, and still have enough reserves to complete development and resume activities post-diapause [
21,
22].
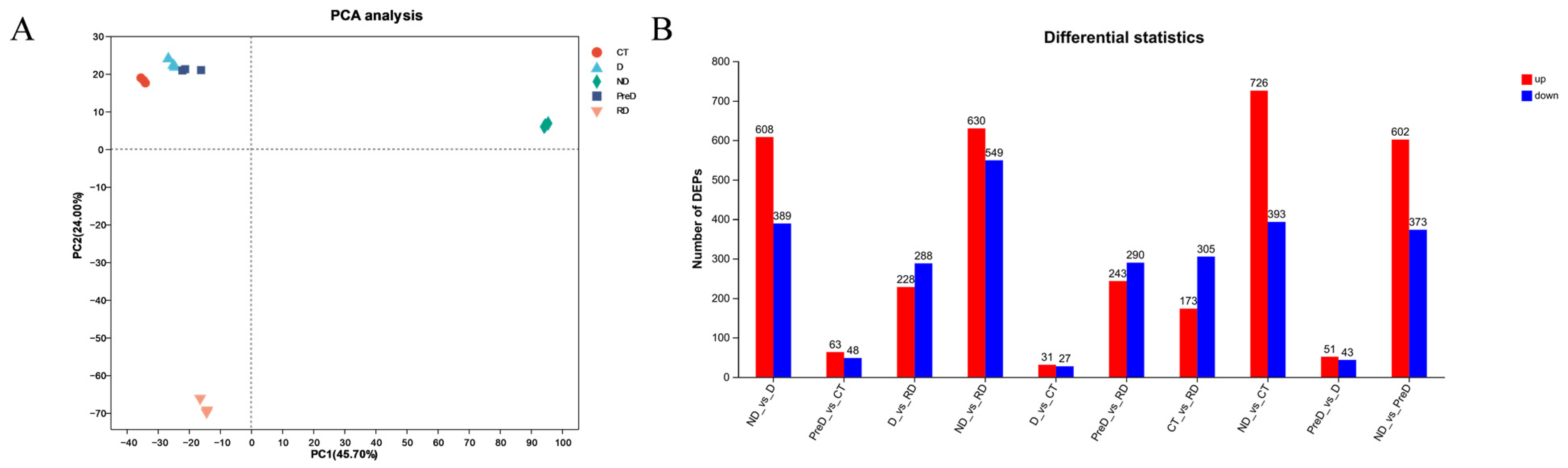
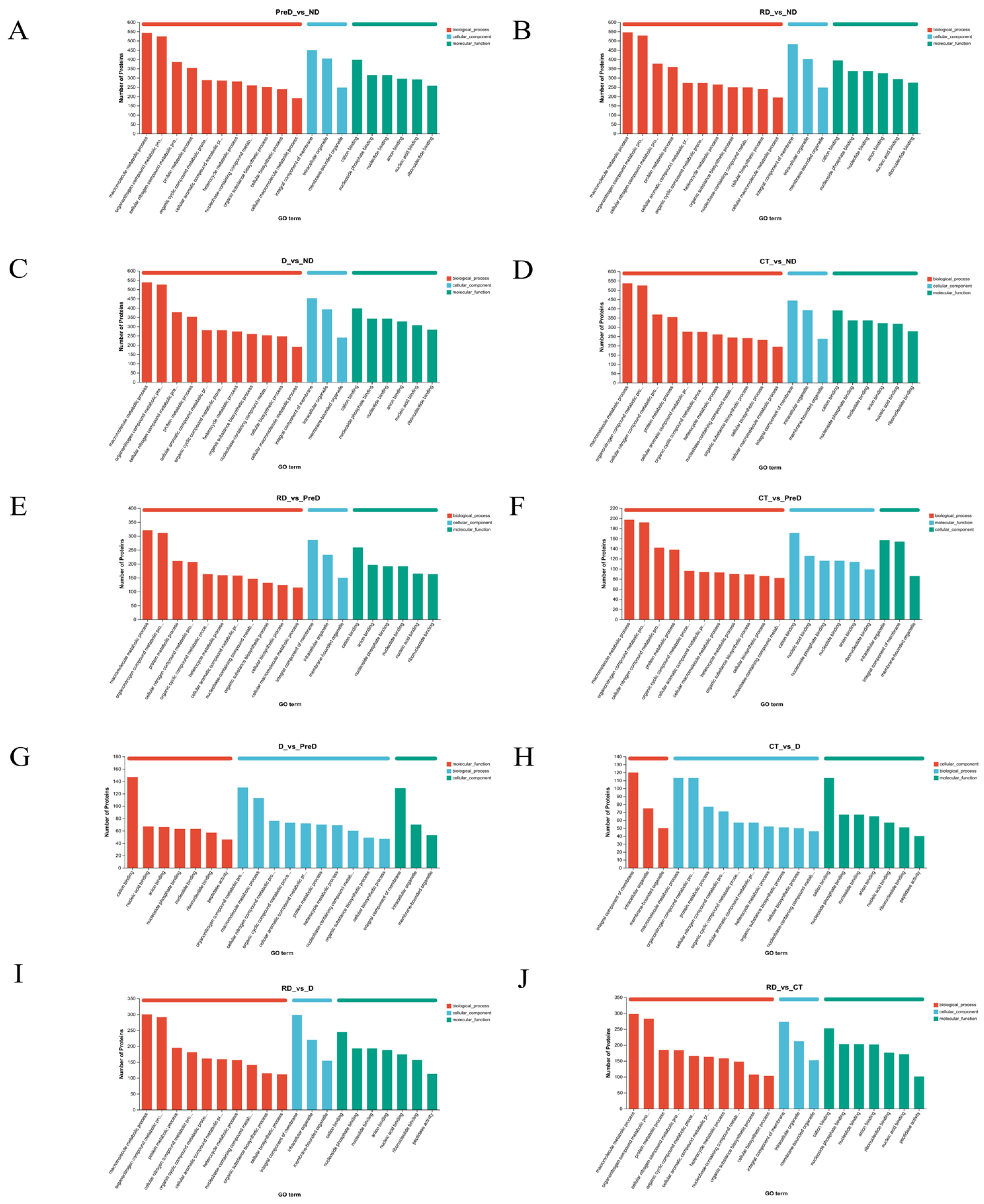
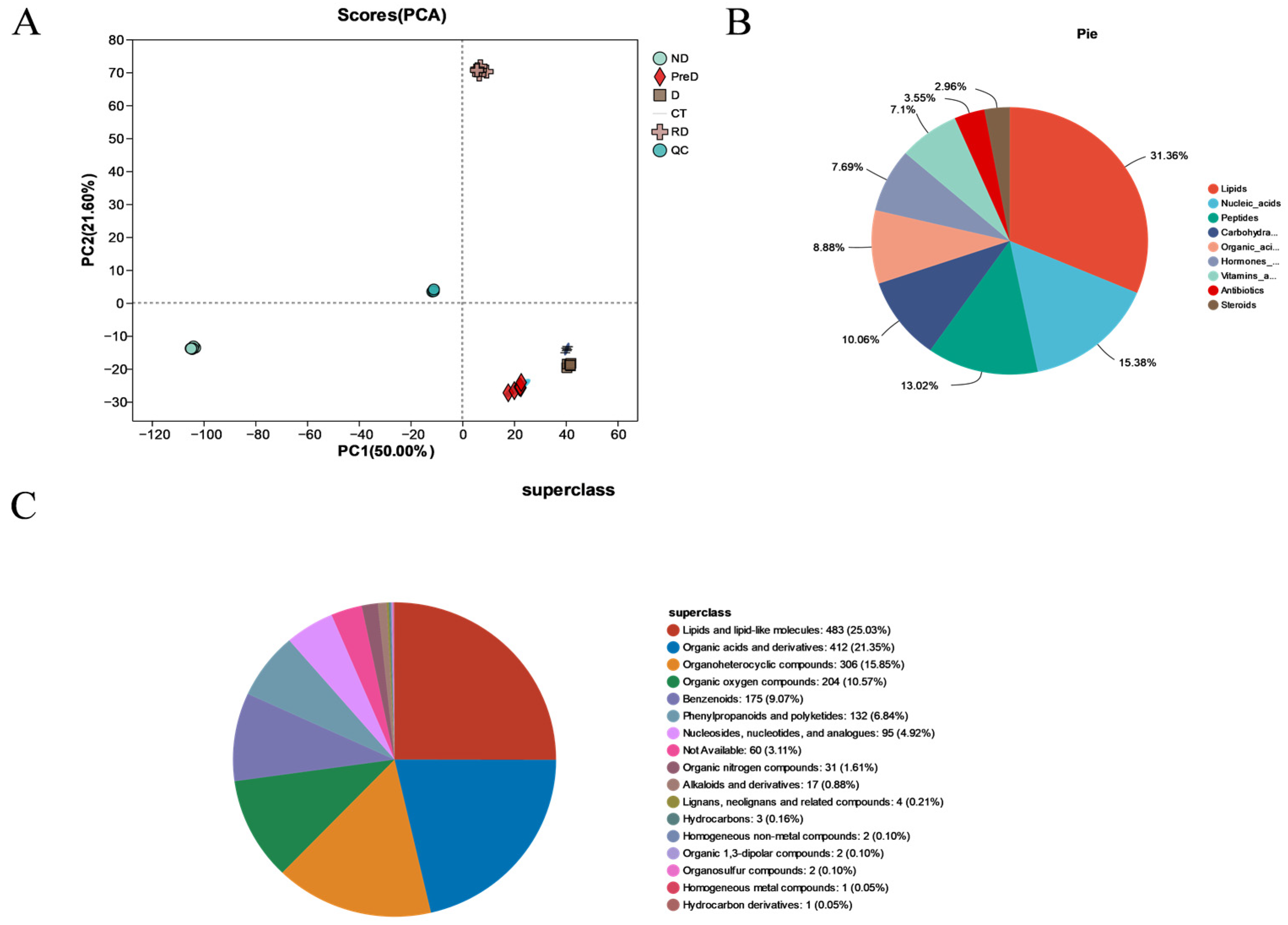
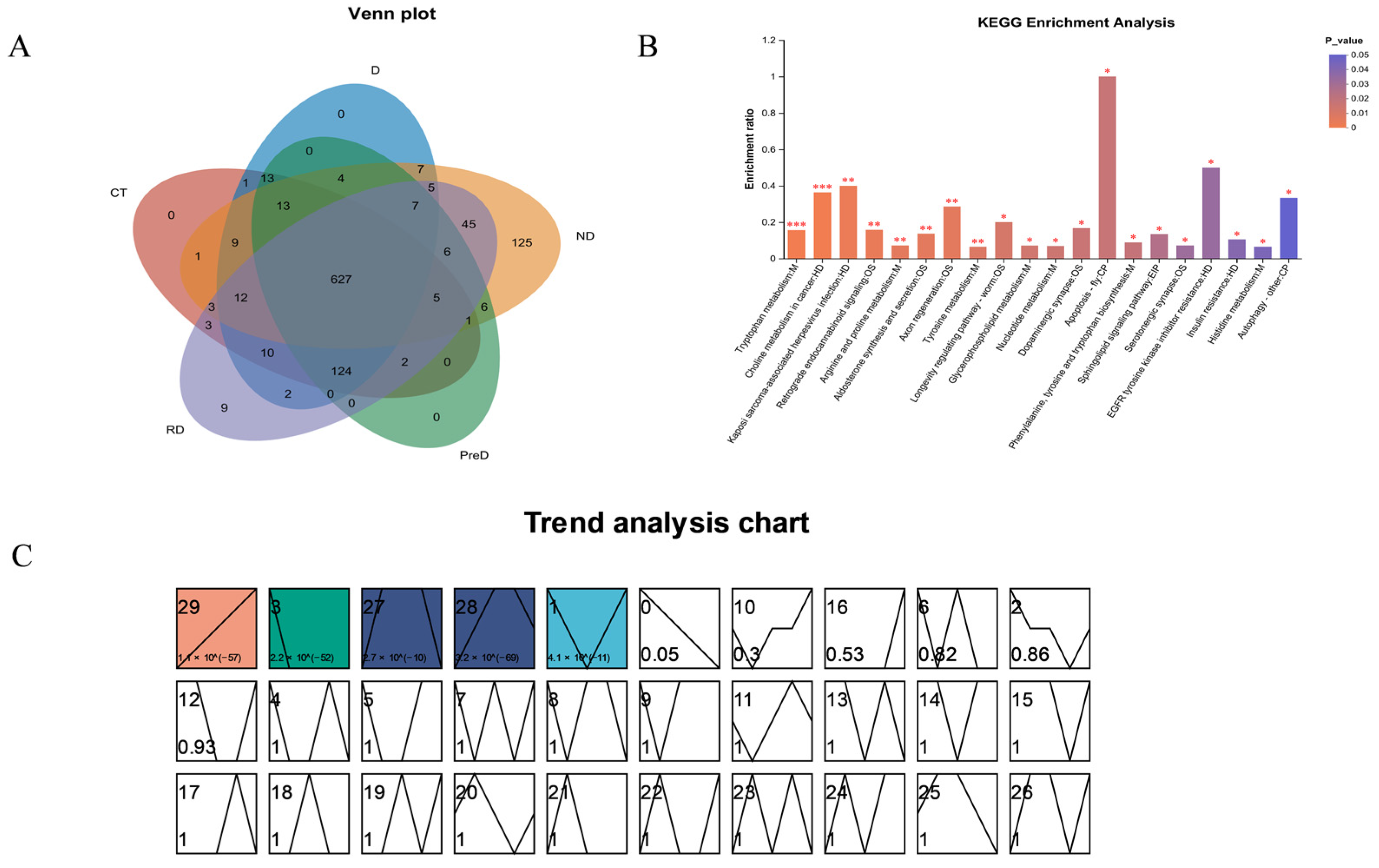
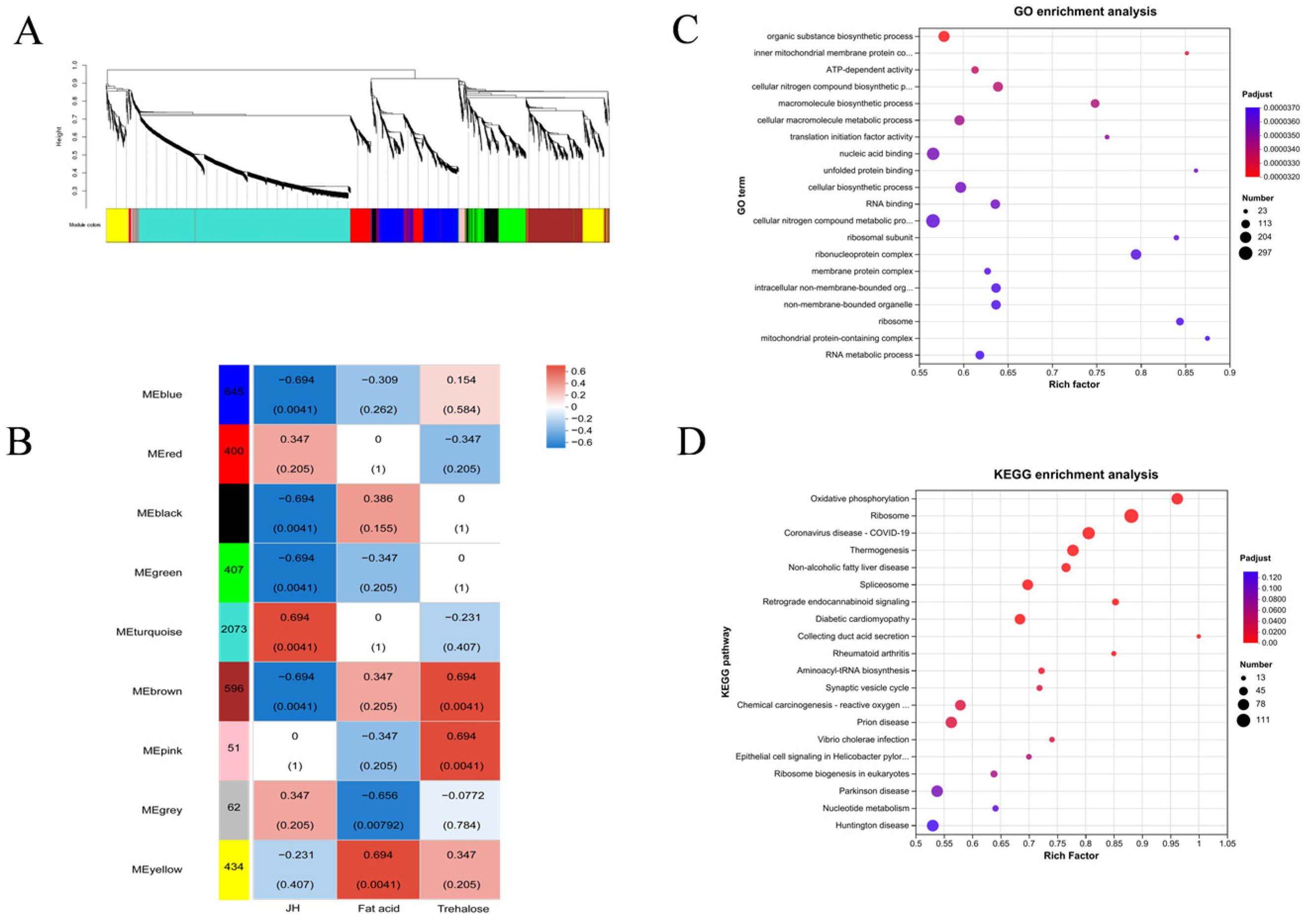
In this study, a total of 5367 DEP and 2020 DAM species were identified, which may include the proteins and metabolites most closely related to diapause of meadow moth. These proteins and metabolites include not only DEP and DAM in the prophase of diapause, diapause, and diapause termination, but also DEP and DAM, which play a key role in the diapause process of meadow moth. We further analyzed the differentially expressed proteins in proteomics and screened 1179 differentially accumulated proteins at different diapause stages. Through comparative analysis, cluster analysis, and weighted gene co-expression network analysis, the important proteins related to carbohydrate metabolism, stress and defense response, lipid metabolism, and insect development were screened out. Based on the metabonomic time series and KEGG enrichment analysis of five groups of diapause larvae of meadow moth, 1255 different metabolites were identified, including amino acid metabolism, lipid metabolism, nucleotide metabolism, etc.
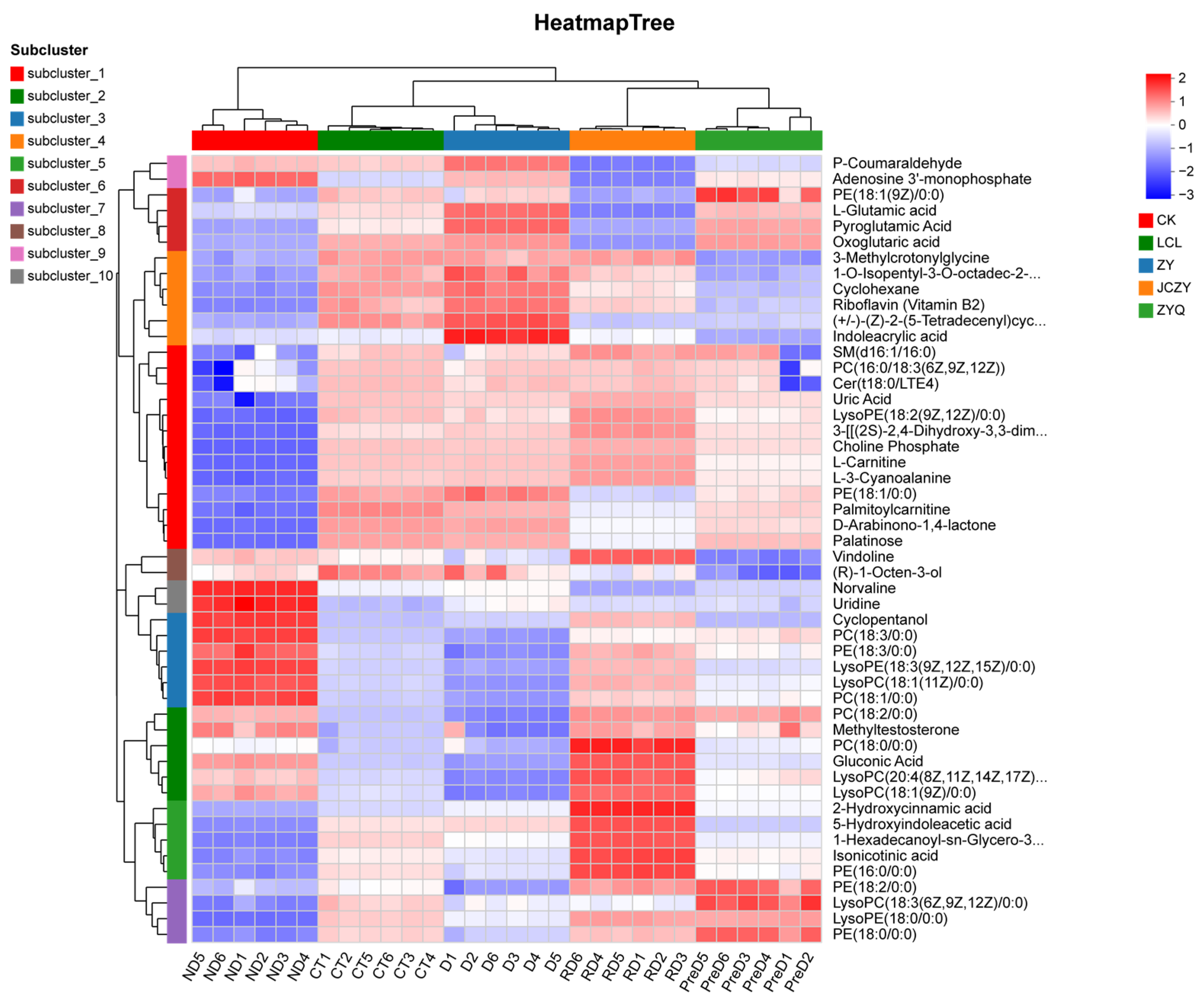
KEGG metabolite analysis shows that among 1255 metabolites, phospholipids are closely related to diapause. Most of the secondary metabolic pathways are important regulators in the diapause process of meadow moth. Phospholipids can promote lipid metabolism within insects and are integral components of cell membranes, capable of binding with membrane proteins to regulate cellular metabolism, energy conversion, cell repair, and signal transduction [
23]. In this study, certain phospholipids such as PI (Phosphatidyl inositol), PC (Phosphatidyl choline, also known as lecithin), PE (Phosphatidyl ethanolamine, also known as cephalin), PS (Phosphatidyl serine), and PA (Phosphatidic acid) were significantly higher in the diapause-affected Ostrinia furnacalis compared to the non-diapause group. Research on Bombyx mori has also shown that phospholipid levels positively correlate with cold resistance [
24]. Increased phospholipid content may influence lipid metabolism, transformation, and intercellular energy transfer during diapause in Ostrinia furnacalis, thereby enhancing cold tolerance. Lysophospholipids, derivatives of phospholipid hydrolysis, are more polar than phospholipids and can enhance membrane fluidity upon extensive contact with cell membranes [
25].
To investigate whether circadian rhythm gene-related proteins and thermogenesis gene-related proteins play a role in protein variations during different diapause stages of the fall armyworm, we conducted Gene Set Enrichment Analysis (GSEA) on various treatment groups. The results showed that the role of circadian rhythm gene-related proteins in diapause and diapause release of meadow moth was not obvious. Heat-generating genes are the main reason for the differences in each stage. This may be because the regulation network of circadian rhythm signals is extremely complex, and the analysis result is not significant [
26].
WGCNA analysis of differentially expressed proteins revealed that 2073 proteins were highly correlated with JH (r = 0.694,
p = 4.1 × 10
−3). We believe that the 2073 proteins most related to JH in the light blue module are the key protein set for regulating diapause of meadow moth. We analyzed the protein interaction network of the light blue module and found that the 40s and 60s subunits of the ribosome played an important role in the whole network. Previous studies have shown that HDAC may play an important role in the diapause of meadow moth [
10]. At the same time, it was reported that lysine acetylation may increase the combination diversity of ribosomal activity, and then affect the histone modification in plant cells [
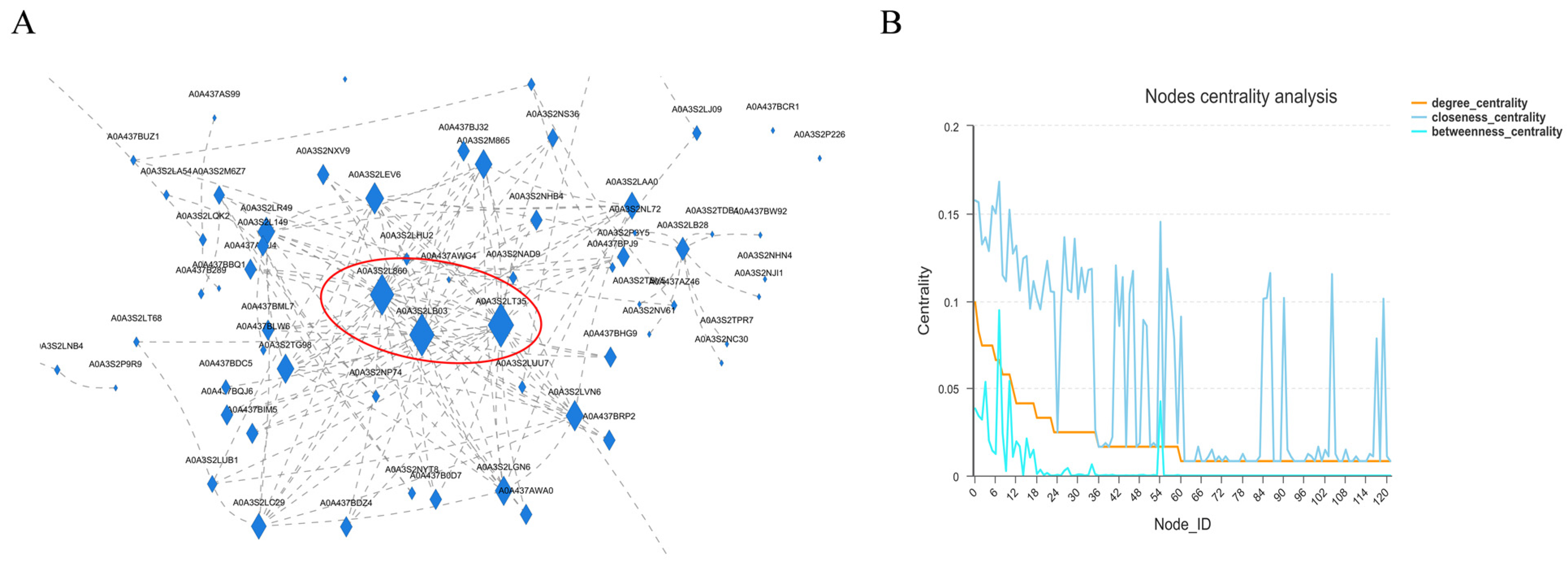
11]. Therefore, we speculate that the 40s and 60s subunits of ribosomes are regulated by histone deacetylase to compress the chromatin structure during diapause of meadow moth, which makes it difficult for RNA polymerase to enter, thus reducing protein-related expression. That is to say, there may be an interaction between HDAC and 40s and 60s subunits of ribosomes. These results expand the relevant regulatory network of meadow moth diapause and provide new insights.
However, the regulation of thermogenic pathways directly involved in energy metabolism is more direct than that of circadian rhythm genes. In the next step, we will further verify the functions of ribosomal subunits and HDAC. At the same time, we studied the influence mechanism of histone acetylase-related genes on diapause in meadow moth, and then determined the molecular mechanism of acetylation and endocrine regulation coupling. In addition, in order to mediate acetylation regulation, we will study the cellular signal transduction and co-regulation of acetylation in insect diapause. Acetylation may be a molecular marker for diapause regulation or developmental decision in insects. It is of great significance to analyze the specific role of epigenetic code in diapause regulation, and then to understand the diapause mechanism of insects from a broader perspective.
4. Materials and Methods
4.1. Insect Collection
The caterpillars used in this experiment were collected from the agricultural fields of Dalad Banner and Otog Front Banner in Ordos City, Inner Mongolia Autonomous Region. To ensure that the experimental insects maintained consistent vitality, we reared the collected over-wintering generation of caterpillars in an insect-controlled climate chamber at 21 °C, with a humidity of 60–70% and an L:D ratio of 16:8 for three generations, after which they underwent diapause treatment.
4.2. Acquisition of Diapause and Diapause-Lifting Larvae
The diapause of the meadow moth is induced under the conditions of 21 °C and a 12:12 L:D cycle. In this laboratory, the diapause induction training of the meadow moth was conducted according to previous methods [
4]. The 1st to 4th instar larvae of the meadow moth were reared under 21 °C and a 16:8 L:D cycle, while the 5th instar larvae were induced under 21 °C and a 12:12 L:D cycle (ND). The feeding conditions for the control group and experimental group were identical, but the control group maintained a 16L:8D light condition after entering the 5th instar. After the control group’s meadow moths burrowed into the soil to make cocoons and emerged, we collected the non-burrowed portion of the diapause-treated group as pre-diapause (PreD), while collecting the cocoons from the control group, cutting off one end of its cocoon, and observing its growth. After the control group pupated for over a week, the diapause-treated group remained in a larval state, and was named the diapause group (D). The meadow moths after two weeks of diapause were transferred to diapause-release conditions (0–5 °C, 0L:24D) for 20 days, serving as the cold treatment group (CT). After being placed under normal cultivation conditions (21 ± 1 °C, 16L:8D) for 7–10 days, some of the moths in the bottle began to pupate, and those that remained in larval form were considered diapause-released larvae (RD). All tested meadow moths were placed in a rearing box measuring 28 × 18 × 13.5 cm, with 50 individuals per box. Five states of meadow moths (ND, PreD, D, CT, RD) were subjected to proteomic sequencing (
Figure 13A). Twenty samples were collected from each treatment and replicated three times, with each sample collected near the end of the photoperiod.
4.3. Sample Preparation for Proteomics Analysis
An appropriate amount of sample in the frozen state is taken out and transferred to the vibrating tube. An appropriate amount of protein lysate (8 M urea + 1% SDS, containing protease inhibitor) is added. A high-flux tissue grinder was used to vibrate the sample 3 times, for 40 s each time. Pyrolysis was performed on ice for 30 min, and vortex mixing was carried out for 5–10 s every 5 min. The supernatant was collected after centrifugation at 4 °C for 30 min at 12,000 G. Subsequently, the protein content was determined by the BCA method, and the operation was carried out in strict accordance with the instructions of the BCA reagent [
26].
The final concentration of teab was made to be 100 mm by adding teab (triethylammonium bicarbonate buffer) to 100 µg of protein; TCEP (tris (2-carboxyethyl) phosphine) was added so that the final concentration of TCEP was 10 mm, and the reaction was carried out at 37 °C for 60 min; IAM (iodoacetamide) was added to make the final concentration of IAM 40 mm, and the mixture was kept in the dark for 40 min at room temperature. Each tube was added with pre-cooled acetone (acetone/sample
v/
v = 6:1) and was precipitated at −20 °C for 4 h. Centrifugation was performed at 10,000×
g for 20 min, and the sediment was collected. The sample was fully dissolved with 100 µL of 100 mm teab. Trypsin was added according to an enzyme/protein ratio (m/m) of 1:50, and enzymolysis was performed overnight at 37 °C. The TMT reagent was taken out at 20 °C and allowed to return to room temperature; acetonitrile was added, vortexed, and centrifuged. Each tube was added with 100 μg of polypeptide into a TMT reagent and incubated at room temperature for 2 h. Hydroxylamine was added, reacted at room temperature for 30 min, mixed with the same amount of labeled product in a tube, and drained with a vacuum concentrator [
27,
28].
The peptide samples were redissolved with UPLC loading buffer (2% acetonitrile (adjusted to pH 10 with ammonia water)), and high-pH liquid-phase separation was performed on an acquity UPLC beh C18 column of 1.7 µm, 2.1 mm × 150 mm (Waters, Milford, MA, USA). Phase A involved 2% acetonitrile (adjusted to pH 10 with ammonia); phase B involved 80% acetonitrile (adjusted to pH 10 with ammonia), 0–1.9 min, 0–0% B; 9~2 min, 0~5% B; 2–17 min, 5–5% B; 17~18 min, 5~10% B; 5 min, 10−30% B; 5~38 min, 30~36% B; 38~39 min, 36~42% B; 39~40 min, 42~100% B; 40–44 min, 100% B; 44~45 min, 100~0% B; 45~48 min, 0~0% B. The UV detection wavelength was 214 nm, and the volume flow rate was 200 μL/min. The elution time was 48 min. According to the peak shape and time, 28 fractions were collected and combined into 14 fractions, which were concentrated by vacuum centrifugation.
4.4. Sample Preparation for Metabonomic Analysis
The solid sample was taken into a 2 mL centrifuge tube, and a grinding ball with a diameter of 6 mm was added; 400 μL of extract (methanol/water = 4:1 (
v:
v)) containing 0.02 mg/mL of internal standard (l-2-chlorophenylalanine) was used for the extraction of metabolites. The sample solution was ground for 6 min (−10 °C, 50 Hz) with a frozen tissue grinder, and then subjected to low-temperature ultrasound for 30 min (5 °C, 40 kHz). The sample was then placed at −20 °C for 30 min and centrifuged for 15 min (4 °C, 13,000×
g), and the supernatant was transferred to the injection vial with an internal cannula for analysis [
29].
4.5. LC-MS/MS Analysis
Proteomics in the second dimension was analyzed by liquid chromatography tandem mass spectrometry (evosep one combined with obitrap exploris 480 mass spectrometer) (Shanghai Meiji Biomedical Technology Co., Ltd, Shanghai, China). The peptide was dissolved in a mass spectrometry loading buffer and then loaded on a C18 column (150 mm). The volume flow rate was 300 NL/min; 1% formic acid aqueous solution was in phase a, with 100% acetonitrile plus 0. 1% formic acid in phase B, 0~2 min, 5~5% B; 2–30 min, 5–38% B; 30–40 min, 38–90% B; 40–44 min, 90–90% B. The MS scanning range (M/z) was 350–1500, the acquisition mode was DDA, and the fragmentation mode was HCD. The resolution of primary mass spectrometry was 60,000, the resolution of secondary mass spectrometry was 15,000, and the dynamic exclusion time was 30 s. Turbotmt intelligent acquisition improves the resolution of reported ion isotopes.
The instrument platform for metabonomic LC-MS analysis is the uhplc-q exactive system (Shanghai Meiji Biomedical Technology Co., Ltd.) of semefier. Chromatographic conditions: a 3 μL sample was separated by an HSS T3 chromatographic column (100 mm × 2.1 mm I.D., 1.8 µm) and detected by mass spectrometry. Mass spectrum conditions: the sample mass spectrum signal was collected in the positive and negative ion scanning mode, and the mass scanning range was m/z: 70–1050. The ion spray voltage and positive ion voltage were 3500 V, the negative ion voltage was 2800 V, the sheath gas was 40 psi, the auxiliary heating gas was 10 psi, and the ion source heating temperature was 400 °C, with 20–40–60 V cyclic collision energy, an MS1 resolution of 70,000, and an MS2 resolution of 17,500.
4.6. Bioinformatics Analysis
The original files of mass spectrometry off the machine were analyzed by proteomediscoverertm software 2.4, and the database used for searching the database was the Meiji biological self-developed database. The false discovery rate (FDR) of peptide identification was set to FDR ≤ 0.01, the mass tolerance of peptides was set to 20 ppm, and the fragment mass tolerance was set to 0.02 da, containing at least one specific peptide [
30].
The original data of LC-MS were imported into the metabonomics processing software progenesis Qi (Waters Corporation, Milford, MA, USA) for baseline filtering, peak identification, integration, retention time correction, and peak alignment. Finally, a data matrix of retention time, mass charge ratio, and peak intensity was obtained. At the same time, MS and MSMS mass spectrometry information was linked to the metabolism public databases HMDB (
http://www.hmdb.ca/, accessed on 12 December 2023) and metlin (
https://metlin.scripps.edu/, accessed on 17 December 2023) and meggith built its own database to match and obtain the metabolite information. We uploaded the database-searched data matrix to the meggitt cloud platform (cloud.majorbio.com, accessed on 17 December 2023) for analysis. Firstly, the data matrix was preprocessed as follows: the data matrix used an 80% rule to remove missing values, that is, to retain at least 80% of non-zero variables in a group of samples, and then fill the gap value (the minimum value in the original matrix fills the gap value). In order to reduce the error caused by sample preparation and instrument instability, the total normalization method was used to normalize the response intensity of the sample mass spectrum peak to obtain the normalized data matrix. At the same time, the variables with a relative standard deviation (RSD) > 30% of QC samples were deleted, and log10 logarithm processing was performed to obtain the data matrix for subsequent analysis. Secondly, the ropls package (version1.6.2) in R language was used to perform principal component analysis (PCA) and orthogonal least partial binary discriminant analysis (OPLS-DA) on the preprocessed data matrix, and the stability of the model was evaluated by seven cycles of interactive validation. The selection of significantly different metabolites was determined based on the variable weight value (VIP) obtained by the opls-da model and the
p value of Student’s
t test. The metabolites with vip > 1 and
p < 0.05 were significantly different metabolites.
Differential metabolites were annotated by the KEGG database (
https://www.kegg.jp/kegg/pathway.html, accessed on 17 December 2023) to obtain the pathways involved by differential metabolites. Python software (Version 3.10) package scipy.stats was used for pathway enrichment analysis, and a Fisher exact test was used to obtain the biological pathway that was most relevant to the experimental treatment.
5. Conclusions
In summary, through TMT proteomic and metabolomic analysis on non-diapause (ND), pre-diapause (PreD), diapause (D), cold treatment (CT), and post-diapause (RD) stages of the meadow moth, we found significant differences in protein and metabolic profiles between diapause and non diapause states. Then, we screened for differential proteins and metabolites among the different treatment groups of grassland moths mentioned above, and annotated a total of 1179 differential proteins and 1255 co expressed differential metabolites. We conducted KEGG analysis and time series analysis on the co expressed metabolites, and found that phospholipids were annotated in significantly different modules, indicating their important role in the diapause process of grassland moths. Using phospholipids as a weighted gene co expression network analysis indicator to analyze differentially expressed proteins, it was found that ribosomal 40s and 60s subunits were the most relevant proteins for diapause. Based on previous research in our laboratory, it has been found that the histone deacetylase of the meadow moth is related to its diapause. The results demonstrate that histone deacetylase regulates the 40s and 60s subunits of ribosomes, thereby affecting the diapause of the meadow moth.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}