Acylhydrazone Derivative A5 Promotes Neurogenesis by Up-Regulating Neurogenesis-Related Genes and Inhibiting Cell-Cycle Progression in Neural Stem/Progenitor Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Compound A5 Promotes Neuronal Differentiation and Neurite Outgrowth of Neuro-2a Cells
2.2. Compound A5 Enhances Neurogenesis in Cultured NSPCs
2.3. Compound A5 Promotes Morphological Maturation of Newborn Neurons Derived from NSPCs
2.4. Global Analyses of Transcriptome Changes after Treatment with Compound A5 in NSPCs
2.5. Compound A5 up-Regulates Genes Related to Neurogenesis
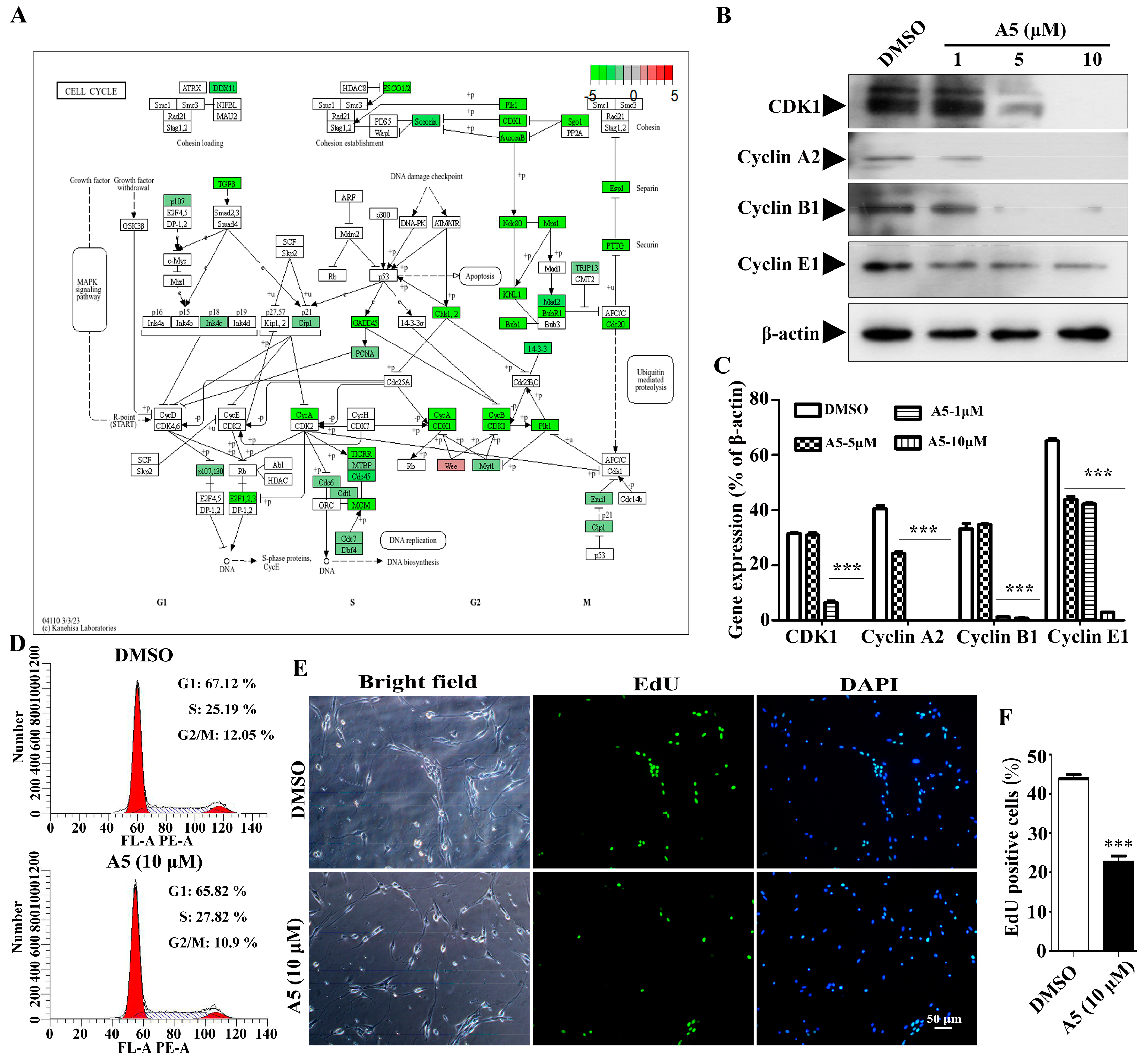
2.6. Compound A5 Inhibits Cell Cycle Progression in NPSCs
3. Materials and Methods
3.1. Reagents
3.2. Cell Culture
3.3. MTT Analysis
3.4. Western Blotting
3.5. Immunostaining
3.6. Differential Expression Analysis by RNA-Seq
3.7. Cell Cycle Assay by Flow Cytometry
3.8. EdU Assay
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gage, F.H. Mammalian Neural Stem Cells. Science 2000, 287, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Ho, N.F.; Hooker, J.M.; Sahay, A.; Holt, D.J.; Roffman, J.L. In vivo imaging of adult human hippocampal neurogenesis: Progress, pitfalls and promise. Mol. Psychiatry 2013, 18, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Aimone, J.B.; Gage, F.H. New neurons and new memories: How does adult hippocampal neurogenesis affect learning and memory? Nat. Rev. Neurosci. 2010, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Azari, M.F.; Profyris, C.; Zang, D.W.; Petratos, S.; Cheema, S.S. Induction of endogenous neural precursors in mouse models of spinal cord injury and disease. Eur. J. Neurol. Off. J. Eur. Fed. Neurol. Soc. 2015, 12, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Horgusluoglu, E.; Nudelman, K.; Nho, K.; Saykin, A.J. Adult neurogenesis and neurodegenerative diseases: A systems biology perspective. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2016, 174, 93–112. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Gage, F.H. Adult hippocampal neurogenesis and its role in Alzheimer′s disease. Mol. Neurodegener. 2011, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Mattova, S.; Simko, P.; Urbanska, N.; Kiskova, T. Bioactive Compounds and Their Influence on Postnatal Neurogenesis. Int. J. Mol. Sci. 2023, 24, 16614. [Google Scholar] [CrossRef]
- Rahman, A.A.; Amruta, N.; Pinteaux, E.; Bix, G.J. Neurogenesis after Stroke: A Therapeutic Perspective. Transl. Stroke Res. 2020, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Berger, T.; Lee, H.; Young, A.H.; Aarsland, D.; Thuret, S. Adult Hippocampal Neurogenesis in Major Depressive Disorder and Alzheimer’s Disease. Trends Mol. Med. 2020, 26, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Potts, M.B.; Lim, D.A. An Old Drug for New Ideas: Metformin Promotes Adult Neurogenesis and Spatial Memory Formation. Cell Stem Cell 2012, 11, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Akbari-Fakhrabadi, M.; Najafi, M.; Mortazavian, S.; Memari, A.; Shidfar, F.; Shahbazi, A.; Heshmati, J. Saffron (Crocus Sativus L.), Combined with Endurance Exercise, Synergistically Enhances BDNF, Serotonin, and NT-3 in Wistar Rats. Rep. Biochem. Mol. Biol. 2021, 9, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vidaa, D.I.; Po, K.T.; Fung, K.H.; Chow, K.W.; Tsang, W.H. Lavender essential oil ameliorates depression-like behavior and increases neurogenesis and dendritic complexity in rats. Neurosci. Lett. 2019, 701, 426–434. [Google Scholar]
- Ayuob, N.N.; Balgoon, M.J.; Ali, S.; Alnoury, I.S.; Abdelfattah, A.A. Ocimum basilicum (Basil) Modulates Apoptosis and Neurogenesis in Olfactory Pulp of Mice Exposed to Chronic Unpredictable Mild Stress. Front. Psychiatry 2020, 11, 569711–569720. [Google Scholar] [CrossRef] [PubMed]
- Esfandiary, E.; Karimipour, M.; Mardani, M.; Alaei, H.; Esmaeili, A. Novel effects of Rosa damascena extract on memory and neurogenesis in a rat model of Alzheimer′s disease. J. Neurosci. Res. 2014, 92, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Tang, G.; Yang, J.; Ding, J.; Lin, H.; Xiang, X. Synthesis of some new acylhydrazone compounds containing the 1,2,4-triazole structure and their neuritogenic activities in Neuro-2a cells. RSC Adv. 2020, 10, 18927–18935. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, A.; Ye, B.; Dotti, C.G. Neuronal polarity: Demarcation, growth and commitment. Curr. Opin. Cell Biol. 2012, 24, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Pauklin, S.; Vallier, L. The Cell-Cycle State of Stem Cells Determines Cell Fate Propensity. Cell 2013, 155, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Michowski, W.; Kolodziejczyk, A.; Sicinski, P. The cell cycle in stem cell proliferation, pluripotency and differentiation. Nat. Cell Biol. 2019, 21, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Cheffer, A.; Tárnok, A.; Ulrich, H. Cell Cycle Regulation During Neurogenesis in the Embryonic and Adult Brain. Stem Cell Rev. Rep. 2013, 9, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Zhuang, X.; Li, S.; Shi, L. Arhgef1 is expressed in cortical neural progenitor cells and regulates neurite outgrowth of newly differentiated neurons. Neurosci. Lett. 2016, 638, 27–34. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, X.; Jiang, X.; Lin, H.; Yu, M.; Wu, L.; Zhou, R. Acylhydrazone Derivative A5 Promotes Neurogenesis by Up-Regulating Neurogenesis-Related Genes and Inhibiting Cell-Cycle Progression in Neural Stem/Progenitor Cells. Molecules 2024, 29, 3330. https://doi.org/10.3390/molecules29143330
Xiang X, Jiang X, Lin H, Yu M, Wu L, Zhou R. Acylhydrazone Derivative A5 Promotes Neurogenesis by Up-Regulating Neurogenesis-Related Genes and Inhibiting Cell-Cycle Progression in Neural Stem/Progenitor Cells. Molecules. 2024; 29(14):3330. https://doi.org/10.3390/molecules29143330
Chicago/Turabian StyleXiang, Xiaoliang, Xia Jiang, Hongwei Lin, Meixing Yu, Liming Wu, and Rong Zhou. 2024. "Acylhydrazone Derivative A5 Promotes Neurogenesis by Up-Regulating Neurogenesis-Related Genes and Inhibiting Cell-Cycle Progression in Neural Stem/Progenitor Cells" Molecules 29, no. 14: 3330. https://doi.org/10.3390/molecules29143330
APA StyleXiang, X., Jiang, X., Lin, H., Yu, M., Wu, L., & Zhou, R. (2024). Acylhydrazone Derivative A5 Promotes Neurogenesis by Up-Regulating Neurogenesis-Related Genes and Inhibiting Cell-Cycle Progression in Neural Stem/Progenitor Cells. Molecules, 29(14), 3330. https://doi.org/10.3390/molecules29143330