
Age-Dependent Effects of Yolkin on Contact Sensitivity and Immune Phenotypes in Juvenile Mice

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Results
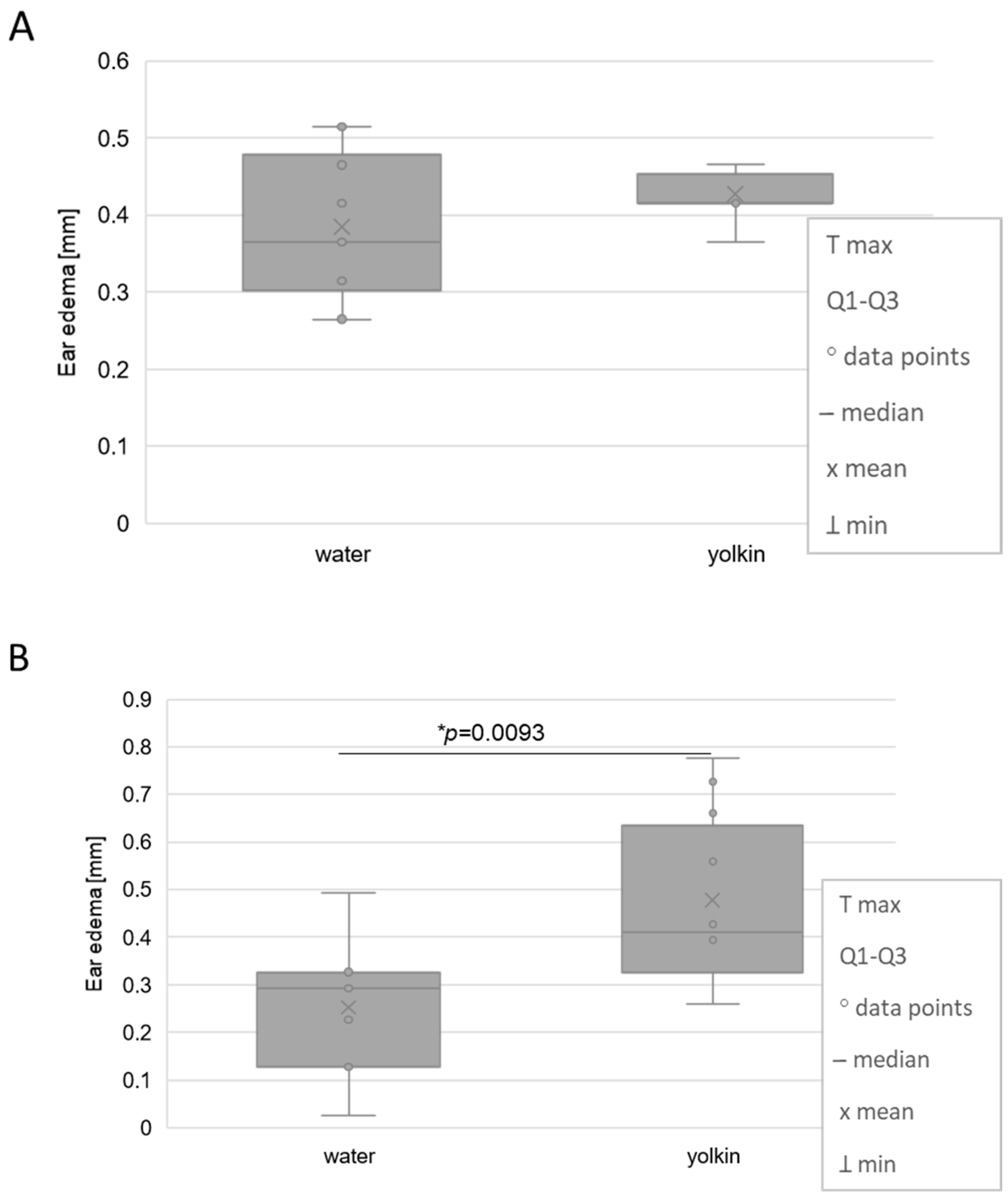
2.1. Effects of Yolkin on Contact Sensitivity to Oxazolone in Adolescent Mice
2.2. Effects of Yolkin on Blood Cell Type Composition in Contact Sensitivity to Oxazolone in Adolescent Mice
2.3. Effects of Yolkin on the Phenotype of Cells in the Lymphoid Organs
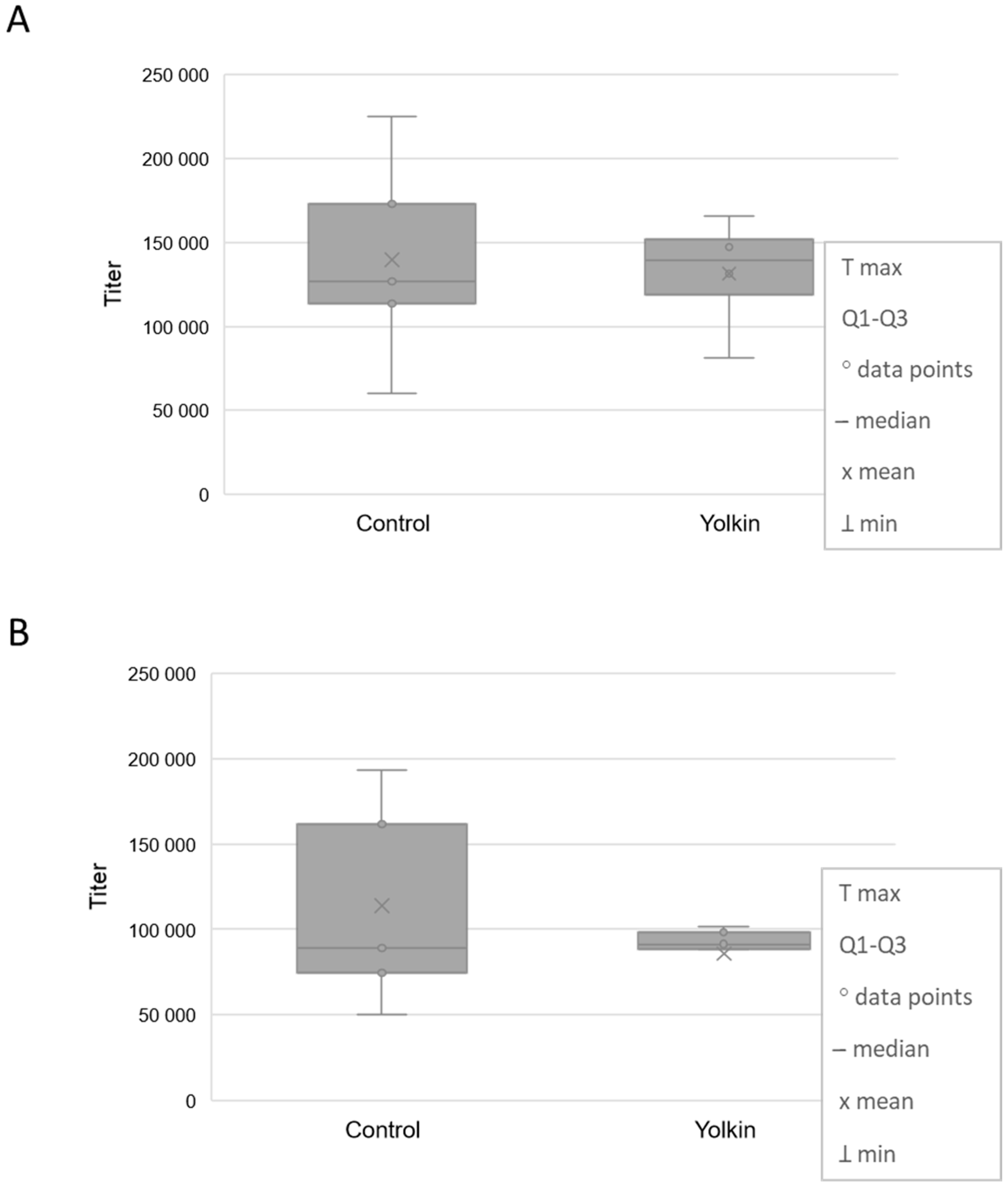
2.4. Effects of Yolkin on the Antibody Production to Ovalbumin
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Reagents
4.3. Yolkin Preparation
4.4. Experimental Design
4.5. Phenotypic Determinations of Splenocytes, Mesenteric Lymph Node Lymphocytes, and Bone Marrow Cells
4.6. Contact Sensitivity to Oxazolone
4.7. Analysis of the Peripheral Blood Cell Composition
4.8. Immunization of Mice with OVA
4.9. End-Point Titrations of Anti-OVA IgG and IgG Antibodies of Each Animal Serum
4.10. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adkins, B.; Bu, Y.; Guevara, P. The generation of Th memory in neonates versus adults: Prolonged primary Th2 effector function and impaired development of Th1 memory effector function in murine neonates. J. Immunol. 2001, 166, 918925. [Google Scholar] [CrossRef]
- Adkins, B.; Bu, Y.; Cepero, E.; Perez, R. Exclusive Th2 primary effector function in spleens but mixed Th1/Th2 function in lymph nodes of murine neonates. J. Immunol. 2000, 164, 2347–2353. [Google Scholar] [CrossRef]
- Woodward, B.D.; Woods, J.W.; Crouch, D.A. Direct evidence that primary acquired cell-mediated immunity is less resistant than is primary thymus-dependent humoral immunity to the depressive influence of wasting protein-energy malnutrition in weanling mice. Am. J. Clin. Nutr. 1992, 55, 1180–1185. [Google Scholar] [CrossRef]
- Steevels, T.A.M.; Hillyer, L.M.; Monk, J.M.; Fisher, M.E.; Woodward, B.D. Effector/memory T cells of the weanling mouse exhibit Type 2 cytokine polarization in vitro and in vivo in the advanced stages of acute energy deficit. J. Nutr. Biochem. 2010, 21, 504–511. [Google Scholar] [CrossRef]
- Zimecki, M.; Mazurier, J.; Spik, G.; Kapp, J.A. Human lactoferrin induces phenotypic and functional changes in murine splenic B cells. Immunology 1995, 86, 122–127. [Google Scholar]
- Zimecki, M.; Whiteley, P.J.; Pierce, C.W.; Kapp, J.A. Presentation of antigen by B cells subsets. I. Lyb-5+ and Lyb-5- B cells differ in ability to stimulate antigen specific T cells. Arch. Immunol. Ther. Exp. 1994, 42, 115–123. [Google Scholar]
- Itakura, A.; Szczepanik, M.; Campos, R.A.; Paliwal, V.; Majewska, M.; Matsuda, H.; Takatsu, K.; Askenase, P.W. An hour after immunization peritoneal B-1 cells are activated to migrate to lymphoid organs where within 1 day they produce IgM antibodies that initiate elicitation of contact sensitivity. J. Immunol. 2005, 175, 7170–7178. [Google Scholar] [CrossRef] [PubMed]
- Kerfoot, S.M.; Szczepanik, M.; Tung, J.W.; Askenase, P.W. Identification of initiator B cells, a novel subset of activation-induced deaminase-dependent B-1-like cells that mediate initiation of contact sensitivity. J. Immunol. 2008, 181, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, J.; Li, K.; Li, M.; Xu, S.; Liu, X.; Zhang, Z.; Xu, X.; Zhang, Y.; Pan, J.; et al. Single-cell genomics identifies distinct B1 cell developmental pathways and reveals aging-related changes in the B-cell receptor repertoire. Cell Biosci. 2022, 12, 57. [Google Scholar] [CrossRef]
- Dewar, A.L.; Doherty, K.V.; Woods, G.M.; Lyons, A.B.; Muller, H.K. Acquisition of immune function during the development of the Langerhans cell network in neonatal mice. Immunology 2001, 103, 61–69. [Google Scholar] [CrossRef]
- Muthukkumar, S.; Goldstein, J.; Stein, K.E. The ability of B cells and dendritic cells to present antigen increases during ontogeny. J. Immunol. 2000, 165, 4803–4813. [Google Scholar] [CrossRef] [PubMed]
- Funch, A.B.; Mraz, V.; Gadsbøll, A.-S.Ø.; Jee, M.H.; Weber, J.F.; Ødum, N.; Woetmann, A.; Johansen, J.D.; Geisler, C.; Bonefeld, C.M. CD8+ tissue-resident memory T cells recruit neutrophils that are essential for flare-ups in contact dermatitis. Allergy 2022, 77, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Fyhrquist, N.; Wolff, H.; Lauerma, A.; Alenius, H. CD8+ T cell migration to the skin requires CD4+ help in a murine model of contact hypersensitivity. PLoS ONE 2012, 7, e41038. [Google Scholar] [CrossRef] [PubMed]
- Castillo-González, R.; Cibrian, D.; Fernández-Gallego, N.; Ramírez-Huesca, M.; Saiz, L.M.; Navarro, M.N.; Fresno, M.; de la Fuente, H.; Sánchez-Madrid, F. Galectin-1 Expression in CD8+ T Lymphocytes Controls Inflammation in Contact Hypersensitivity. J. Investig. Dermatol. 2021, 141, 1522–1532.e3. [Google Scholar] [CrossRef] [PubMed]
- Ring, S.; Schäfer, S.C.; Mahnke, K.; Lehr, H.-A.; Enk, A.H. CD4+ CD25+ regulatory T cells suppress contact hypersensitivity reactions by blocking influx of effector T cells into inflamed tissue. Eur. J. Immunol. 2006, 36, 2981–2992. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.; Saio, M.; Yamashita, H.; Tanaka, H.; Takami, T.; Ezaki, T.; Inagaki, N. Lactobacillus acidophilus strain L-92 induces CD4(+)CD25(+)Foxp3(+) regulatory T cells and suppresses allergic contact dermatitis. Biol. Pharm. Bull. 2012, 35, 612–616. [Google Scholar] [CrossRef]
- Majewska-Szczepanik, M.; Zemelka-Wiącek, M.; Ptak, W.; Wen, L.; Szczepanik, M. Epicutaneous immunization with DNP-BSA induces CD4+ CD25+ Treg cells that inhibit Tc1-mediated CS. Immunol. Cell Biol. 2012, 90, 784–795. [Google Scholar] [CrossRef]
- Artym, J.; Zimecki, M. An overview on immunological activity of calf thymus extract (TFX®) and its therapeutic benefits. J. Biomed. Res. Ther. 2021, 1, 1–22. [Google Scholar]
- Zimecki, M.; Kochanowska, I.; Zaczyńska, E.; Kocięba, M.; Artym, J.; Zambrowicz, A.; Matwiejczyk, M.; Besman, M.; Kuchar, K.; Skotnicki, A. Immunoregulatory actions of calf thymus extract (TFX®) in vitro in relation to its effect on expression of mitogen activated protein kinases. Int. Immunopharmacol. 2023, 118, 109995. [Google Scholar] [CrossRef]
- Zimecki, M.; Mazurier, J.; Machnicki, M.; Wieczorek, Z.; Montreuil, J.; Spik, G. Immunostimulatory activity of lactotransferrin and maturation of CD4- CD8- murine thymocytes. Immunol. Lett. 1991, 30, 119–123. [Google Scholar] [CrossRef]
- Zimecki, M. A proline-rich polypeptide from ovine colostrum: Colostrinin with immunomodulatory activity. Adv. Exp. Med. Biol. 2008, 606, 241–250. [Google Scholar] [PubMed]
- Janusz, A.; Zabłocka, A. Colostrinin: A proline-rich polypeptide complex of potential therapeutic interest. Cell. Mol. Biol. 2013, 58, 4–11. [Google Scholar]
- Hillyer, L.M.; Woodward, B. A comparison of the capacity of six cold-pressed plant oils to support development of acquired immune competence in the weanling mouse: Superiority of low-linoleic-acid oils. Br. J. Nutr. 2002, 88, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Polanowski, A.; Sosnowska, A.; Zabłocka, A.; Janusz, M.; Trziszka, T. Immunologically active peptides that accompany hen egg yolk immunoglobulin Y: Separation and identification. Biol. Chem. 2013, 394, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, S.; Li, H.; Li, L. Vitellogenin, a multivalent sensor and an antimicrobial effector. Int. J. Biochem. Cell Biol. 2011, 43, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Kazana, W.; Mitkiewicz, M.; Ochnik, M.; Sochocka, M.; Zambrowicz, A.; Piechowiak, G.; Macała, J.; Miernikiewicz, P.; Zabłocka, A. Yolkin Isolated from Hen Egg Yolk as a Natural Immunoregulator, Activating Innate Immune Response in BMDM Macrophages. Oxidat. Med. Cell. Longev. 2020, 2020, 5731021. [Google Scholar] [CrossRef] [PubMed]
- Zabłocka, A.; Sosnowska, A.; Urbaniak, A.; Janusz, M.; Polanowski, A. Peptides accompanying chicken egg yolk IgY—Alternative methods of isolation and immunoregulatory activity. Food Funct. 2014, 5, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Zambrowicz, A.; Zabłocka, A.; Bobak, Ł.; Macała, J.; Janusz, M.; Polanowski, A.; Trziszka, T. A simple and rapid method of isolation of active polypeptide complex, yolkin, from chicken egg yolk. Food Chem. 2017, 230, 705–711. [Google Scholar] [CrossRef]
- Sugawara, Y.; Uehara, A.; Fujimoto, Y.; Kusumoto, S.; Fukase, K.; Shibata, K.; Sugawara, S.; Sasano, T.; Takada, H. Toll-like receptors, NOD1, and NOD2 in oral epithelial cells. J. Dent. Res. 2006, 85, 524–529. [Google Scholar] [CrossRef]
- Ortega-Cava, C.F.; Ishihara, S.; Rumi, M.A.K.; Kawashima, K.; Ishimura, N.; Kazumori, H.; Udagawa, J.; Kadowaki, Y.; Kinoshita, Y. Strategic compartmentalization of Toll-like receptor 4 in the mouse gut. J. Immunol. 2003, 170, 3977–3985. [Google Scholar] [CrossRef]
- Lemieszewska, M.; Jakubik-Witkowska, M.; Stańczykiewicz, B.; Zambrowicz, A.; Zabłocka, A.; Polanowski, A.; Trziszka, T.; Rymaszewska, J. Pro-cognitive properties of the immunomodulatory polypeptide complex, yolkin, from chicken egg yolk and colostrum-derived substances: Analyses based on animal model of age-related cognitive deficits. Arch. Immunol. Ther. Exp. 2016, 64, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Obmińska-Mrukowicz, B.; Szczypka, M.; Lis, M.; Pawlak, A.; Suszko-Pawłowska, A.; Sysak, A.; Zambrowicz, A.; Burster, T.; Kocięba, M.; Artym, J.; et al. Effects of yolkin on the immune response of mice and its plausible mechanism of action. Immunol. Lett. 2020, 220, 21–31. [Google Scholar] [CrossRef]
- Szczypka, M.; Lis, M.; Kuczkowski, M.; Bobrek, K.; Pawlak, A.; Zambrowicz, A.; Gaweł, A.; Obminska-Mrukowicz, B. Yolkin, a Polypeptide Complex from Egg Yolk, Affects Cytokine Levels and Leukocyte Populations in Broiler Chicken Blood and Lymphoid Organs after In Ovo Administration. Int. J. Mol. Sci. 2023, 24, 17494. [Google Scholar] [CrossRef]
- Menager-Marcq, I.; Pomie, C.; Romagnoli, P.; van Meerwijk, J.P. CD8+CD28- regulatory T lymphocytes prevent experimental inflammatory bowel disease in mice. Gastroenterology 2006, 131, 1775–1785. [Google Scholar] [CrossRef]
- Pomié, C.; Ménager-Marcq, I.; van Meerwijk, J.P.M. Murine CD8+ regulatory T lymphocytes: The new era. Hum. Immunol. 2008, 69, 708–714. [Google Scholar] [CrossRef]
- Gunduz, O.; Sapmaz-Metin, M.; Topuz, R.D.; Kaya, O.; Karadag, C.H.; Ulugol, A. Anti-Inflammatory and Antipruritic Effects of Remote Ischaemic Postconditioning in a Mouse Model of Experimental Allergic Contact Dermatitis. Medicina 2023, 59, 1816. [Google Scholar] [CrossRef] [PubMed]
- Kazana, W.; Jakubczyk, D.; Pacyga-Prus, K.; Leszczyńska, K.; Górska, S.; Siednienko, J.; Macała, J.; Piechowiak, G.; Zabłocka, A. A Novel Mechanism of Macrophage Activation by the Natural Yolkin Polypeptide Complex from Egg Yolk. Int. J. Mol. Sci. 2022, 23, 3125. [Google Scholar] [CrossRef] [PubMed]
- McGrath-Morrow, S.A.; Lee, S.; Gibbs, K.; Lopez, A.; Collaco, J.M.; Neptune, E.; Soloski, M.J.; Scott, A.; D’Alessio, F. Immune Response to Intrapharyngeal LPS in Neonatal and Juvenile Mice. Am. J. Respir. Cell Mol. Biol. 2015, 52, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Boehmer, E.D.; Meehan, M.J.; Cutro, B.T.; Kovacs, E.J. Aging negatively skews macrophage TLR2- and TLR4-mediated pro-inflammatory responses without affecting the IL-2-stimulated pathway. Mech. Ageing Dev. 2005, 126, 1305–1313. [Google Scholar] [CrossRef]
- Noonan, F.P.; Halliday, W.J. Studies on contact hypersensitivity and tolerance in vivo and in vitro. I. Basic characteristics of the reactions and confirmation of an immune response in tolerant mice. Int. Arch. Allergy Appl. Immunol. 1978, 56, 523–532. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Experimental Group | % of Cell Types in the Circulating Blood | |||||
|---|---|---|---|---|---|---|
| B | S | E | L | M | B | |
| water/BG | 1.5 ± 0.0 | 32.1 ± 1.3 | 1.5 ± 0.2 | 62.2 ± 0.4 | 2.0 ± 0.4 | 1.0 ± 0.2 |
| water/OXA | 3.4 ± 0.9 | 48.5 ± 2.8 * | 1.5 ± 0.2 | 41.2 ± 2.6 * | 3.8 ± 1.1 | 1.5 ± 0.2 |
| yolkin/OXA | 2.7 ± 0.6 | 52.9 ± 0.8 * | 1.0 ± 0.2 | 39.3 ± 1.2 * | 1.9 ± 0.5 | 1.4 ± 0.2 |
| Splenocytes | ||||||
| CD8+ | CD4+ | CD3+ | CD19+ | CD14+CD169+ | CD25+Foxp3+ | |
| Control | 1.69 ± 0.22 | 9.56 ± 0.94 | 18.32 ± 3.11 | 69.29 ± 6.03 | 2.53 ± 0.12 | 10.48 ± 0.36 |
| Yolkin | 1.42 ± 0.13 | 11.23 ± 0.45 | 15.31 ± 0.97 | 75.94 ± 0.88 | 1.91 ± 0.34 | 8.48 ± 0.20 * |
| Mesenteric lymph node cells | ||||||
| CD8+ | CD4+ | CD3+ | CD19+ | |||
| Control | 5.39 ± 0.35 | 42.10 ± 1.35 | 48.61 ± 1.37 | 49.19 ± 1.28 | ||
| Yolkin | 8.59 ± 0.52 * | 50.22 ± 2.01 * | 55.97 ± 1.43 * | 42.62 ± 1.43 * | ||
| Bone marrow cells | ||||||
| CD19+ | CD11c+ | |||||
| Gated for lymphocytes only | Gated for all cells | |||||
| Control | 82.62 ± 0.82 | 36.83 ± 1.76 | 2.83 ± 0.22 | |||
| Yolkin | 84.95 ± 1.93 | 40.70 ± 2.32 | 2.25 ± 0.35 | |||
| Splenocytes | ||||||
| CD8+ | CD4+ | CD3+ | CD19+ | CD14+CD169+ | CD25+Foxp3+ | |
| Control | 2.81 ± 0.22 | 17.55 ± 1.52 | 22.62 ± 2.49 | 68.99 ± 2.33 | 3.30 ± 0.35 | 9.85 ± 0.40 |
| Yolkin | 2.99 ± 0.14 | 16.50 ± 1.67 | 20.44 ± 2.31 | 66.43 ± 2.11 | 3.58 ± 0.13 | 8.12 ± 0.21 * |
| Mesenteric lymph node cells | ||||||
| CD8+ | CD4+ | CD3+ | CD19+ | |||
| Control | 7.91 ± 0.66 | 45.81 ± 2.67 | 49.86 ± 3.22 | 47.78 ± 3.24 | ||
| Yolkin | 4.69 ± 0.17 * | 42.47 ± 1.08 | 44.40 ± 2.06 | 52.97 ± 1.73 | ||
| Bone marrow cells | ||||||
| CD19+ | CD11c+ | |||||
| Gated for lymphocytes only | Gated for all cells | |||||
| Control | 73.09 ± 1.65 | 30.91 ± 0.89 | 4.62 ± 0.65 | |||
| Yolkin | 77.63 ± 1.40 | 32.55 ± 1.51 | 5.45 ± 0.24 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimecki, M.; Artym, J.; Kocięba, M.; Zaczyńska, E.; Sysak, A.; Szczypka, M.; Lis, M.; Pawlak, A.; Obmińska-Mrukowicz, B.; Kaleta-Kuratewicz, K.; et al. Age-Dependent Effects of Yolkin on Contact Sensitivity and Immune Phenotypes in Juvenile Mice. Molecules 2024, 29, 3254. https://doi.org/10.3390/molecules29143254
Zimecki M, Artym J, Kocięba M, Zaczyńska E, Sysak A, Szczypka M, Lis M, Pawlak A, Obmińska-Mrukowicz B, Kaleta-Kuratewicz K, et al. Age-Dependent Effects of Yolkin on Contact Sensitivity and Immune Phenotypes in Juvenile Mice. Molecules. 2024; 29(14):3254. https://doi.org/10.3390/molecules29143254
Chicago/Turabian StyleZimecki, Michał, Jolanta Artym, Maja Kocięba, Ewa Zaczyńska, Angelika Sysak, Marianna Szczypka, Magdalena Lis, Aleksandra Pawlak, Bożena Obmińska-Mrukowicz, Katarzyna Kaleta-Kuratewicz, and et al. 2024. "Age-Dependent Effects of Yolkin on Contact Sensitivity and Immune Phenotypes in Juvenile Mice" Molecules 29, no. 14: 3254. https://doi.org/10.3390/molecules29143254
APA StyleZimecki, M., Artym, J., Kocięba, M., Zaczyńska, E., Sysak, A., Szczypka, M., Lis, M., Pawlak, A., Obmińska-Mrukowicz, B., Kaleta-Kuratewicz, K., Zambrowicz, A., & Bobak, Ł. (2024). Age-Dependent Effects of Yolkin on Contact Sensitivity and Immune Phenotypes in Juvenile Mice. Molecules, 29(14), 3254. https://doi.org/10.3390/molecules29143254