
Synthesis and Isolation of Phenol- and Thiol-Derived Epicatechin Adducts Prepared from Avocado Peel Procyanidins Using Centrifugal Partition Chromatography and the Evaluation of Their Antimicrobial and Antioxidant Activity

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results and Discussion
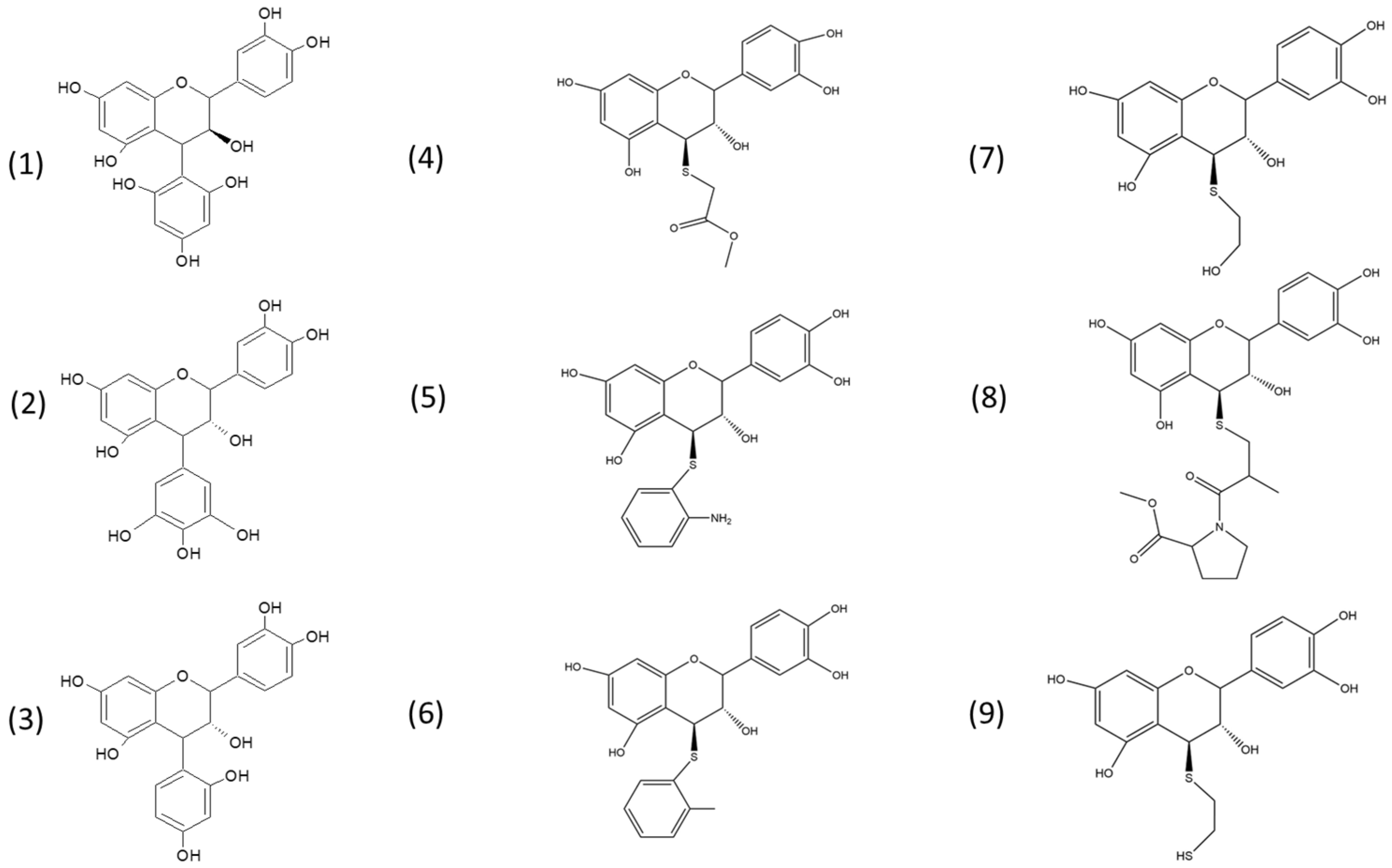
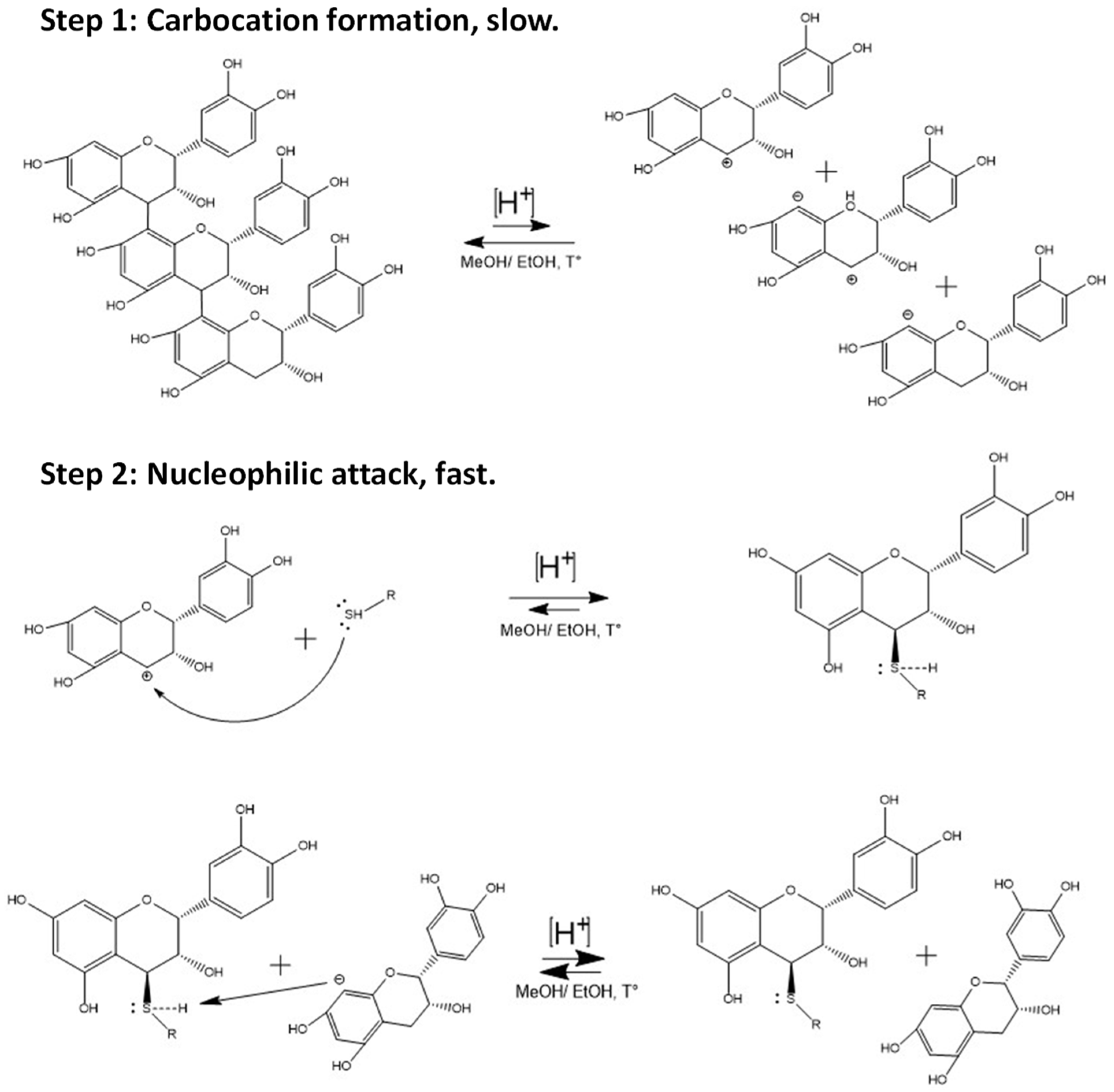
2.1. Extraction of Avocado Peel Procyanidins and Semi-Synthesis of Phenol- and Thiol-Derivatives
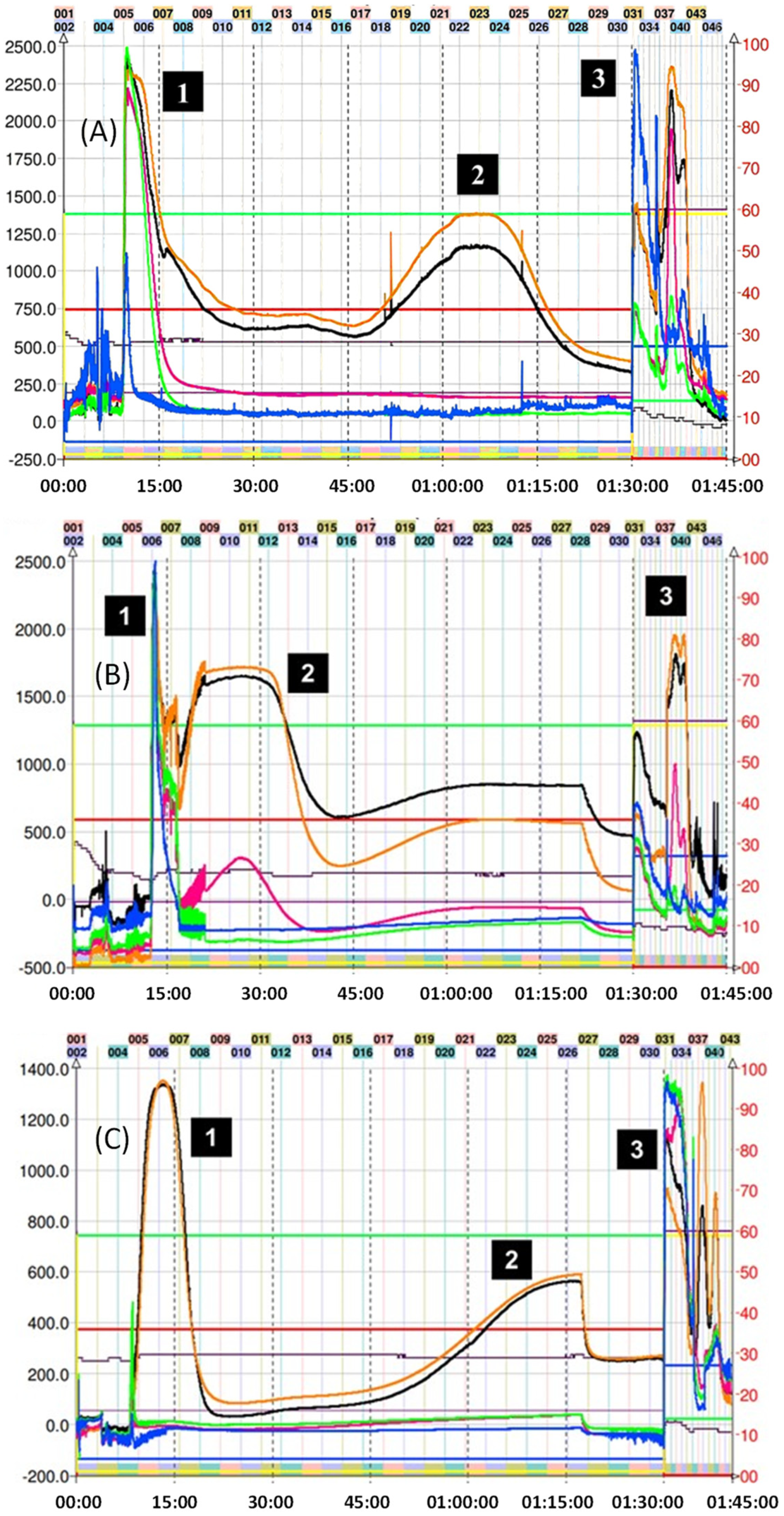
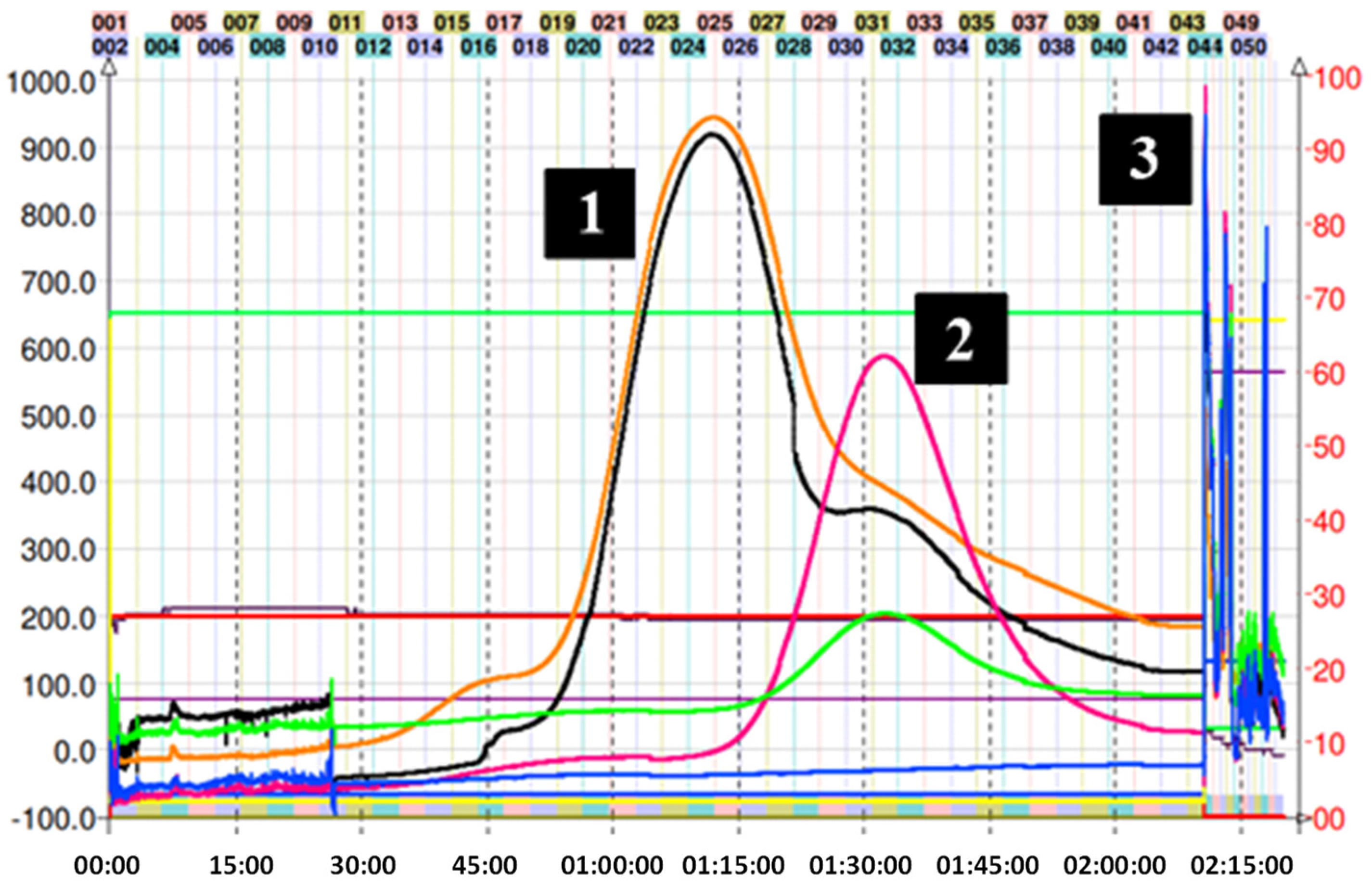
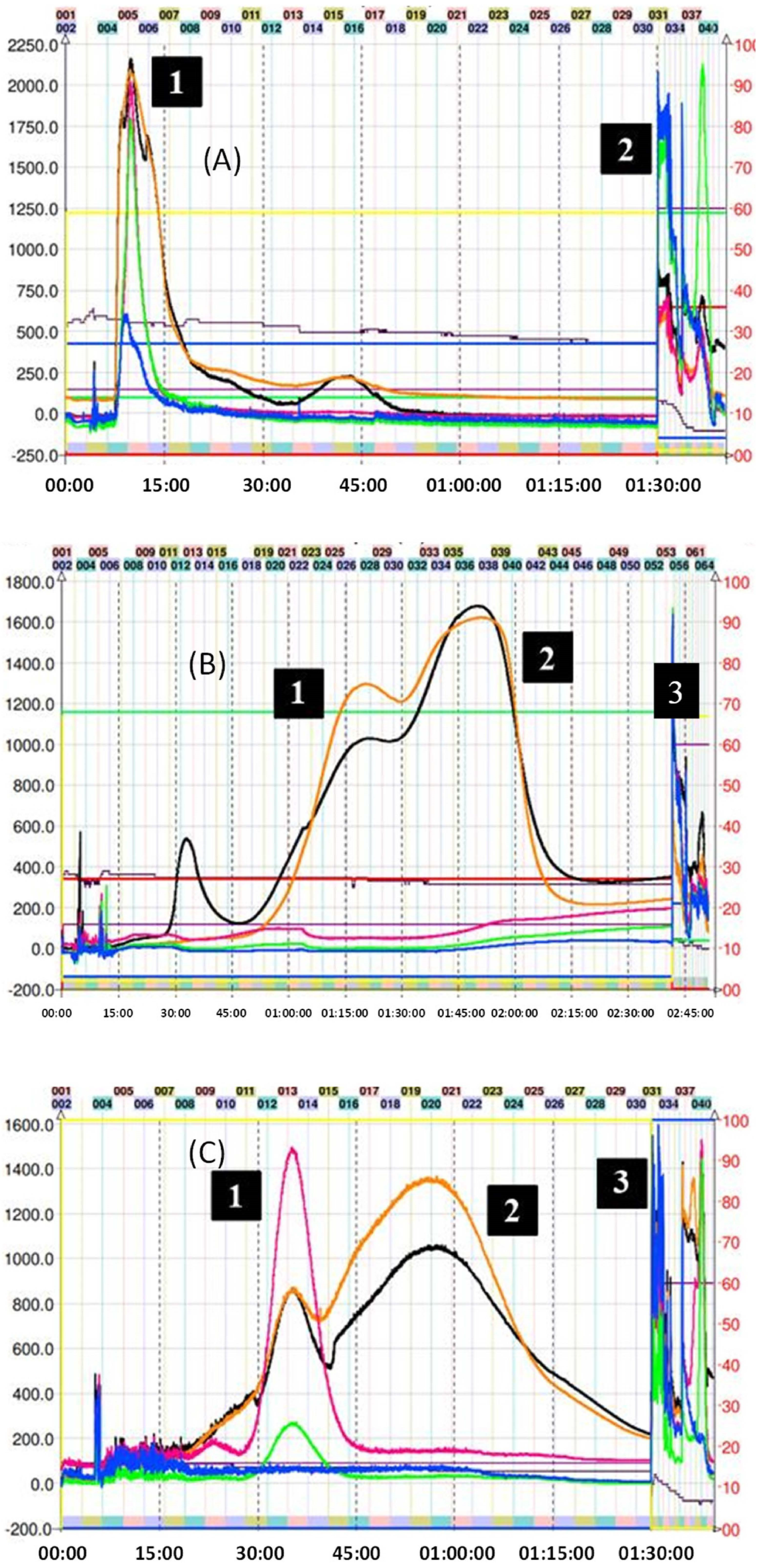
2.2. Isolation of Epicatechin Phenol and Thiol Adducts by Preparative Centrifugal Partition Chromatography (CPC)
2.3. HPLC-ESI-QTOF-MS-MS Analysis of Thiol Adducts
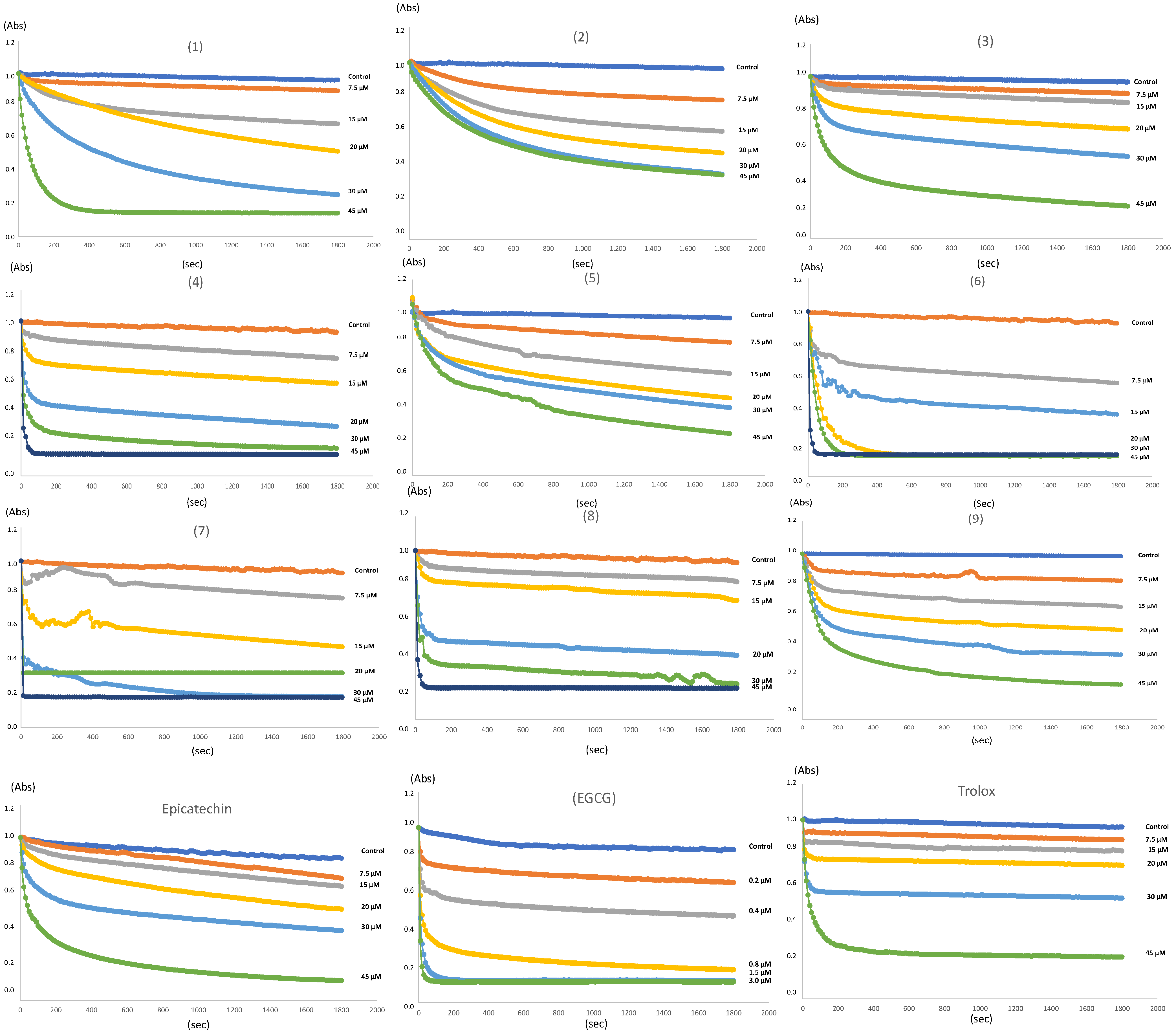
2.4. Antioxidant Activity
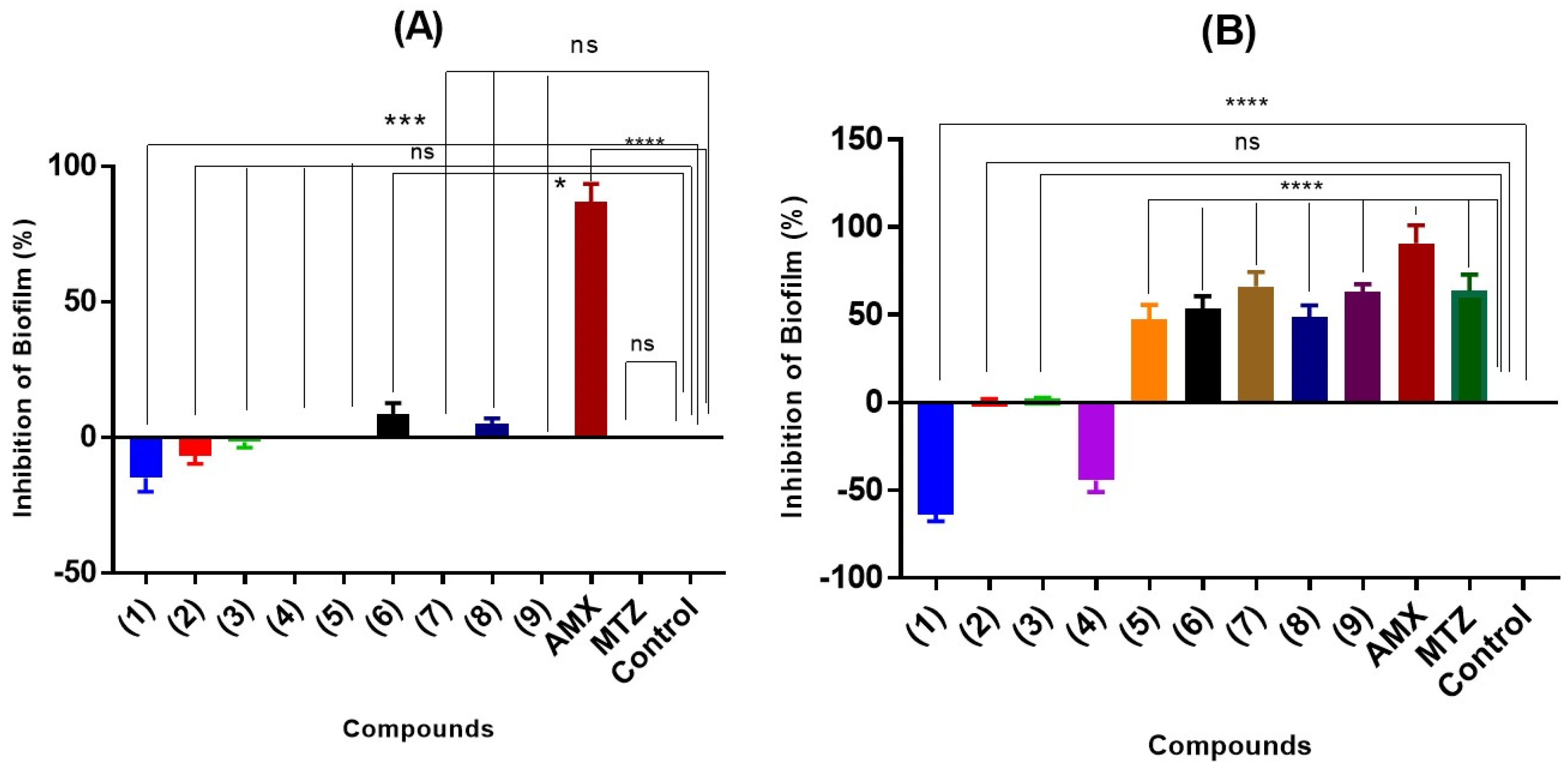
2.5. Antimicrobial and Anti-Biofilm Properties
3. Materials and Methods
3.1. Chemicals
3.2. Avocado Peel Extraction
3.3. HPLC and TLC Analysis
3.4. Semi-Synthesis of Flavan-3-ol Adducts with Phenol and Thiol Nucleophiles
3.5. HPLC-ESI-QTOF-MS-MS
3.6. CPC Separation Procedure
KD Calculations
3.7. DPPH Free Radical Scavenging Activity
3.8. Microbial Culture and Maintenance
3.8.1. Screening Disk and Well-Diffusion Test Susceptibility
3.8.2. Biofilm Test on Biofilm Formation
4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valencia-Hernandez, L.J.; Wong-Paz, J.E.; Ascacio-Valdés, J.A.; Chávez-González, M.L.; Contreras-Esquivel, J.C.; Aguilar, C.N. Procyanidins: From Agro-Industrial Waste to Food as Bioactive Molecules. Foods 2021, 10, 3152. [Google Scholar] [CrossRef]
- Trujillo-Mayol, I.; Viegas, O.; Sobral, M.M.C.; Casas-Forero, N.; Fiallos, N.; Pastene-Navarrete, E.; Faria, M.A.; Alarcón-Enos, J.; Pinho, O.; Ferreira, I.M.P.L.V.O. In Vitro Gastric Bioaccessibility of Avocado Peel Extract in Beef and Soy-Based Burgers and Its Impact on Helicobacter Pylori Risk Factors. Food Chem. 2022, 373, 131505. [Google Scholar] [CrossRef]
- Trujillo-Mayol, I.; Casas-Forero, N.; Pastene-Navarrete, E.; Lima Silva, F.; Alarcón-Enos, J. Fractionation and Hydrolyzation of Avocado Peel Extract: Improvement of Antibacterial Activity. Antibiotics 2020, 10, 23. [Google Scholar] [CrossRef]
- Pastene, E.; Speisky, H.; Troncoso, M.; Alarcón, J.; Figueroa, G. In Vitro Inhibitory Effect of Apple Peel Extract on the Growth of Helicobacter Pylori and Respiratory Burst Induced on Human Neutrophils. J. Agric. Food Chem. 2009, 57, 7743–7749. [Google Scholar] [CrossRef]
- Pastene, E.; Parada, V.; Avello, M.; Ruiz, A.; García, A. Catechin-Based Procyanidins from Peumus Boldus Mol. Aqueous Extract Inhibit Helicobacter Pylori Urease and Adherence to Adenocarcinoma Gastric Cells. Phytother. Res. 2014, 28, 1637–1645. [Google Scholar] [CrossRef]
- Bhagwat, S.; Haytowitz, D. USDA Database for the Proanthocyanidin Content of Selected Foods Release 2; US Department of Agriculture, Agricultural Service, Nutrient Data Laboratory: Beltsville, MD, USA, 2015.
- Araújo, R.G.; Rodriguez-Jasso, R.M.; Ruiz, H.A.; Pintado, M.M.E.; Aguilar, C.N. Avocado By-Products: Nutritional and Functional Properties. Trends Food Sci. Technol. 2018, 80, 51–60. [Google Scholar] [CrossRef]
- Arapitsas, P.; Perenzoni, D.; Guella, G.; Mattivi, F. Improving the Phloroglucinolysis Protocol and Characterization of Sagrantino Wines Proanthocyanidins. Molecules 2021, 26, 1087. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhang, H.; Awika, J.M.; Wang, L.; Qian, H.; Gu, L. Depolymerization of Sorghum Procyanidin Polymers into Oligomers Using HCl and Epicatechin: Reaction Kinetics and Optimization. J. Cereal Sci. 2016, 70, 170–176. [Google Scholar] [CrossRef]
- Bordiga, M.; Coïsson, J.D.; Locatelli, M.; Arlorio, M.; Travaglia, F. Pyrogallol: An Alternative Trapping Agent in Proanthocyanidins Analysis. Food Anal. Methods 2013, 6, 148–156. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Jones, G.P. Analysis of Proanthocyanidin Cleavage Products Following Acid-Catalysis in the Presence of Excess Phloroglucinol. J. Agric. Food Chem. 2001, 49, 1740–1746. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.; García, A.; Aranda, M.; Saéz, V.; Zúñiga, F.; Alarcón, J.; Avello, M.; Pastene, E. One-step purification of two semi-synthetic epicatechin adducts prepared from avocado peels procyanidins by centrifugal partition chromatography and evaluation of their anti-inflammatory effects on adenocarcinoma gastric cells infected with helicobacter pylori. J. Chil. Chem. Soc. 2018, 63, 4222–4228. [Google Scholar] [CrossRef]
- Andrade, V.; Cortés, N.; Pastor, G.; Gonzalez, A.; Ramos-Escobar, N.; Pastene, E.; Rojo, L.E.; Maccioni, R.B. N-Acetyl Cysteine and Catechin-Derived Polyphenols: A Path Toward Multi-Target Compounds Against Alzheimer’s Disease. J. Alzheimer’s Dis. 2020, 75, 1219–1227. [Google Scholar] [CrossRef]
- Carreras, A.; Mesa, J.A.; Cascante, M.; Torres, J.L.; Juliá, L. High Electron Transfer Capacity of Thio-Derivatives of Tea Catechins Measured Using a Water Soluble Stable Free Radical and Their Effects on Colon Cancer Cells. New J. Chem. 2013, 37, 2043–2050. [Google Scholar] [CrossRef]
- Ugartondo, V.; Mitjans, M.; Lozano, C.; Torres, J.L.; Vinardell, M.P. Comparative Study of the Cytotoxicity Induced by Antioxidant Epicatechin Conjugates Obtained from Grape. J. Agric. Food Chem. 2006, 54, 6945–6950. [Google Scholar] [CrossRef]
- García, A. Biofilm and Helicobacter pylori: From Environment to Human Host. World J. Gastroenterol. 2014, 20, 5632. [Google Scholar] [CrossRef]
- Sharma, S.; Mohler, J.; Mahajan, S.D.; Schwartz, S.A.; Bruggemann, L.; Aalinkeel, R. Microbial Biofilm: A Review on Formation, Infection, Antibiotic Resistance, Control Measures, and Innovative Treatment. Microorganisms 2023, 11, 1614. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.P.; DiTullio, P.; Parkinson, S. Bisphosphocins: Novel Antimicrobials for Enhanced Killing of Drug-Resistant and Biofilm-Forming Bacteria. Future Microbiol. 2015, 10, 1751–1758. [Google Scholar] [CrossRef] [PubMed]
- Masák, J.; Čejková, A.; Schreiberová, O.; Řezanka, T. Pseudomonas Biofilms: Possibilities of Their Control. FEMS Microbiol. Ecol. 2014, 89, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Fu, C.; Qin, Y.; Dejian, H. One-pot depolymerizative extraction of proanthocyanidins from mangosteen pericarps. Food Chem. 2009, 114, 874–880. [Google Scholar] [CrossRef]
- Liang, N.; He, F.; Pan, Q.; Wang, J.; Reeves, M.J.; Duan, C. Optimization of Sample Preparation and Phloroglucinol Analysis of Marselan Grape Skin Proanthocyanidins using HPLC-DADESI- MS/MS. S. Afr. J. Enol. Vitic. 2016, 33, 122–131. [Google Scholar] [CrossRef]
- Ito, Y. Golden Rules and Pitfalls in Selecting Optimum Conditions for High-Speed Counter-Current Chromatography. J. Chromatogr. A 2005, 1065, 145–168. [Google Scholar] [CrossRef]
- Köhler, N.; Winterhalter, P. Large-Scale Isolation of Flavan-3-Ol Phloroglucinol Adducts by High-Speed Counter-Current Chromatography. J. Chromatogr. A 2005, 1072, 217–222. [Google Scholar] [CrossRef]
- Cui, C.; Shi, A.; Bai, S.; Yan, P.; Li, Q.; Bi, K. Novel Antihypertensive Prodrug from Grape Seed Proanthocyanidin Extract via Acid-Mediated Depolymerization in the Presence of Captopril: Synthesis, Process Optimization, and Metabolism in Rats. J. Agric. Food Chem. 2018, 66, 3700–3707. [Google Scholar] [CrossRef]
- Sei-ichi, K.; Toda, K.; Matsumoto, K.; Ishihara, C.; Nonobe, S.; Matsunaga, C.; Gomi, Y.K.; Senga, S.; Kawaguchi, K.; Yamamoto, A.; et al. Isolation and Characterization of a Novel Oligomeric Proanthocyanidin with Significant Anti-Cancer Activities from Grape Stems (Vitis vinifera). Sci. Rep. 2019, 9, 12046. [Google Scholar] [CrossRef]
- Selga, A.; Sort, X.; Bobet, R.; Torres, J.L. Efficient One Pot Extraction and Depolymerization of Grape (Vitis vinifera) Pomace Procyanidins for the Preparation of Antioxidant Thio-Conjugates. J. Agric. Food Chem. 2004, 52, 467–473. [Google Scholar] [CrossRef]
- Torres, J.L.; Bobet, R. New Flavanol Derivatives from Grape (Vitis vinifera) Byproducts. Antioxidant Aminoethylthio−Flavan-3-Ol Conjugates from a Polymeric Waste Fraction Used as a Source of Flavanols. J. Agric. Food Chem. 2001, 49, 4627–4634. [Google Scholar] [CrossRef]
- Torres, J.L.; Lozano, C.; Julià, L.; Sánchez-Baeza, F.J.; Anglada, J.M.; Centelles, J.J.; Cascante, M. Cysteinyl-Flavan-3-Ol Conjugates from Grape Procyanidins. Antioxidant and Antiproliferative Properties. Bioorg. Med. Chem. 2002, 10, 2497–2509. [Google Scholar] [CrossRef]
- Selga, A.; Torres, J.L. Efficient Preparation of Catechin Thio Conjugates by One Step Extraction/Depolymerization of Pine (Pinus pinaster) Bark Procyanidins. J. Agric. Food Chem. 2005, 53, 7760–7765. [Google Scholar] [CrossRef]
- Suo, H.; Tian, R.; Xu, W.; Li, L.; Cui, Y.; Zhang, S.; Sun, B. Novel Catechin–Tiopronin Conjugates Derived from Grape Seed Proanthocyanidin Degradation: Process Optimization, High-Speed Counter-Current Chromatography Preparation, as Well as Antibacterial Activity. J. Agric. Food Chem. 2019, 67, 11508–11517. [Google Scholar] [CrossRef]
- Tian, R.; Suo, H.; Zhang, S.; Sun, B. Separation of a Family of Antioxidants Flavan-3-Ol Thio-Conjugates from Procyanidins by High-Speed Counter-Current Chromatography. Eur. Food Res. Technol. 2020, 246, 1017–1029. [Google Scholar] [CrossRef]
- Marchal, L.; Legrand, J.; Foucault, A. Centrifugal Partition Chromatography: A Survey of Its History, and Our Recent Advances in the Field. Chem. Rec. 2003, 3, 133–143. [Google Scholar] [CrossRef]
- Berthod, A.; Brown, L.; Leitão, G.G.; Leitão, S.G. Operating a Countercurrent Chromatography Machine; Elsevier Science: Amsterdam, The Netherlands, 2002; Chapter 2; pp. 21–47. [Google Scholar]
- Roehrer, S.; Minceva, M. Evaluation of Inter-Apparatus Separation Method Transferability in Countercurrent Chromatography and Centrifugal Partition Chromatography. Separations 2019, 6, 36. [Google Scholar] [CrossRef]
- Gao, C.; Cunningham, D.G.; Liu, H.; Khoo, C.; Gu, L. Development of a Thiolysis HPLC Method for the Analysis of Procyanidins in Cranberry Products. J. Agric. Food Chem. 2018, 66, 2159–2167. [Google Scholar] [CrossRef]
- Shi, L.; Wang, J.; Lv, Y. Research Progress on Analysis Methods of Procyanidins. In Proceedings of the IOP Conference Series: Earth and Environmental Science; IOP Publishing Ltd.: Bristol, UK, 2021; Volume 705. [Google Scholar]
- He, F.; Pan, Q.H.; Shi, Y.; Duan, C.Q. Chemical Synthesis of Proanthocyanidins in Vitro and Their Reactions in Aging Wines. Molecules 2008, 13, 3007–3032. [Google Scholar] [CrossRef]
- Mouls, L.; Fulcrand, H. UPLC-ESI-MS Study of the Oxidation Markers Released from Tannin Depolymerization: Toward a Better Characterization of the Tannin Evolution over Food and Beverage Processing. J. Mass Spectrom. 2012, 47, 1450–1457. [Google Scholar] [CrossRef]
- Mateos-Martín, M.L.; Fuguet, E.; Quero, C.; Pérez-Jiménez, J.; Torres, J.L. New Identification of Proanthocyanidins in Cinnamon (Cinnamomum zeylanicum L.) Using MALDI-TOF/TOF Mass Spectrometry. Anal. Bioanal. Chem. 2012, 402, 1327–1336. [Google Scholar] [CrossRef]
- Wang, X.; Liu, C.; Ma, Q.-Y.; Tian, Z.-H.; Jiang, H.-Q.; Lv, Q.-T.; Rong, R. A Rapid and Practical Prediction Method for the Arizona Solvent System Family Used in High Speed Countercurrent Chromatography. J. Chromatogr. A 2020, 1629, 461426. [Google Scholar] [CrossRef]
- Sang, S.; Lambert, J.D.; Hong, J.; Tian, S.; Lee, M.J.; Stark, R.E.; Ho, C.T.; Yang, C.S. Synthesis and structure identification of thiol conjugates of (-)-epigallocatechin gallate and their urinary levels in mice. Chem. Res. Toxicol. 2005, 18, 1762–1769. [Google Scholar] [CrossRef]
- Symma, N.; Hensel, A. Advanced Analysis of Oligomeric Proanthocyanidins: Latest Approaches in Liquid Chromatography and Mass Spectrometry Based Analysis. Phytochem. Rev. 2022, 21, 809–833. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, S.; Kumar, B. LC-MS Identification of Proanthocyanidins in Bark and Fruit of Six Terminalia Species. Nat. Prod. Commun. 2018, 13, 1934578X1801300. [Google Scholar] [CrossRef]
- Kondo, K.; Kurihara, M.; Fukuhara, K.; Tanaka, T.; Suzuki, T.; Miyata, N.; Toyoda, M. Conversion of Procyanidin B-Type (Catechin Dimer) to A-Type: Evidence for Abstraction of C-2 Hydrogen in Catechin during Radical Oxidation. Tetrahedron Lett. 2000, 41, 485–488. [Google Scholar] [CrossRef]
- Karonen, M.; Imran, I.B.; Engström, M.T.; Salminen, J.P. Characterization of Natural and Alkaline-Oxidized Proanthocyanidins in Plant Extracts by Ultrahigh-Resolution UHPLC-MS/MS. Molecules 2021, 26, 1873. [Google Scholar] [CrossRef]
- de Menezes, B.B.; Frescura, L.M.; Duarte, R.; Villetti, M.A.; da Rosa, M.B. A Critical Examination of the DPPH Method: Mistakes and Inconsistencies in Stoichiometry and IC50 Determination by UV–Vis Spectroscopy. Anal. Chim. Acta 2021, 1157, 338398. [Google Scholar] [CrossRef] [PubMed]
- Angeli, L.; Morozova, K.; Scampicchio, M. A Kinetic-Based Stopped-Flow DPPH• Method. Sci. Rep. 2023, 13, 7621. [Google Scholar] [CrossRef]
- Foti, M.C.; Daquino, C.; Geraci, C. Electron-Transfer Reaction of Cinnamic Acids and Their Methyl Esters with the DPPH• Radical in Alcoholic Solutions. J. Org. Chem. 2004, 69, 2309–2314. [Google Scholar] [CrossRef] [PubMed]
- Valachová, K.; Baňasová, M.; Topoľská, D.; Sasinková, V.; Juránek, I.; Collins, M.N.; Šoltés, L. Influence of Tiopronin, Captopril and Levamisole Therapeutics on the Oxidative Degradation of Hyaluronan. Carbohydr. Polym. 2015, 134, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-X.; Liu, Z.-Q. Captopril and 6-Mercaptopurine: Whose SH Possesses Higher Antioxidant Ability? Eur. J. Med. Chem. 2009, 44, 4841–4847. [Google Scholar] [CrossRef]
- Hunsaker, D. The Determination of Thiols with Diphenylpicrylhydrazyl as a Spectrophotometric Reagent. Talanta 1983, 30, 475–480. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, Y.; Luo, J.; Jin, Z.; Liu, Y.; Tang, Y. Synthesis and Antibacterial Activities of Novel Pleuromutilin Derivatives Bearing an Aminothiophenol Moiety. Chem. Biol. Drug Des. 2018, 92, 1627–1637. [Google Scholar] [CrossRef]
- Chai, F.; Wang, J.; Zhou, K.-X.; Wang, S.-K.; Liu, Y.-H.; Jin, Z.; Tang, Y.-Z. Design, Synthesis and Biological Evaluation of Novel Pleuromutilin Derivatives Possessing 4-Aminothiophenol Linker as Promising Antibacterial Agents. Bioorg. Chem. 2022, 126, 105859. [Google Scholar] [CrossRef]
- Garcia-Castillo, V.; Marín-Vega, A.M.; Ilabaca, A.; Albarracín, L.; Marcial, G.; Kitazawa, H.; Garcia-Cancino, A.; Villena, J. Characterization of the Immunomodulatory and Anti- Helicobacter pylori Properties of the Human Gastric Isolate Lactobacillus rhamnosus UCO-25A. Biofouling 2019, 35, 922–937. [Google Scholar] [CrossRef] [PubMed]
- Santativongchai, P.; Tulayakul, P.; Jeon, B. Enhancement of the Antibiofilm Activity of Nisin against Listeria Monocytogenes Using Food Plant Extracts. Pathogens 2023, 12, 444. [Google Scholar] [CrossRef] [PubMed]
- Salas-Jara, M.; Ilabaca, A.; Vega, M.; García, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Wei, X.; Song, J.; Xu, X.; Huang, H.; Fan, S.; Zhang, D.; Han, L.; Lin, J. Interactions between Gut Microbiota and Polyphenols: New Insights into the Treatment of Fatigue. Molecules 2022, 27, 7377. [Google Scholar] [CrossRef]
- Cires, M.J.; Wong, X.; Carrasco-Pozo, C.; Gotteland, M. The Gastrointestinal Tract as a Key Target Organ for the Health-Promoting Effects of Dietary Proanthocyanidins. Front. Nutr. 2017, 3, 1. [Google Scholar] [CrossRef]
- Andriamihaja, M.; Lan, A.; Beaumont, M.; Grauso, M.; Gotteland, M.; Pastene, E.; Cires, M.J.; Carrasco-Pozo, C.; Tomé, D.; Blachier, F. Proanthocyanidin-Containing Polyphenol Extracts from Fruits Prevent the Inhibitory Effect of Hydrogen Sulfide on Human Colonocyte Oxygen Consumption. Amino Acids 2018, 50, 755–763. [Google Scholar] [CrossRef]
- Liu, Z.; Vincken, J.-P.; de Bruijn, W.J.C. Tea Phenolics as Prebiotics. Trends Food Sci. Technol. 2022, 127, 156–168. [Google Scholar] [CrossRef]
- Mukhi, M.; Vishwanathan, A.S. Beneficial Biofilms: A Minireview of Strategies To Enhance Biofilm Formation for Biotechnological Applications. Appl. Environ. Microbiol. 2022, 88, e01994-21. [Google Scholar] [CrossRef]
- Wu, Y.; Luo, X.; Qin, B.; Li, F.; Häggblom, M.M.; Liu, T. Enhanced Current Production by Exogenous Electron Mediators via Synergy of Promoting Biofilm Formation and the Electron Shuttling Process. Environ. Sci. Technol. 2020, 54, 7217–7225. [Google Scholar] [CrossRef]
- Zhao, A.; Sun, J.; Liu, Y. Understanding Bacterial Biofilms: From Definition to Treatment Strategies. Front. Cell Infect. Microbiol. 2023, 13, 1137947. [Google Scholar] [CrossRef]
- Li, Y.; Feng, T.; Wang, Y. The Role of Bacterial Signaling Networks in Antibiotics Response and Resistance Regulation. Mar. Life Sci. Technol. 2022, 4, 163–178. [Google Scholar] [CrossRef]
- Paula, A.J.; Hwang, G.; Koo, H. Dynamics of Bacterial Population Growth in Biofilms Resemble Spatial and Structural Aspects of Urbanization. Nat. Commun. 2020, 11, 1354. [Google Scholar] [CrossRef]
- Kye, H.; Kim, J.; Ju, S.; Lee, J.; Lim, C.; Yoon, Y. Microplastics in Water Systems: A Review of Their Impacts on the Environment and Their Potential Hazards. Heliyon 2023, 9, e14359. [Google Scholar] [CrossRef]
- Djaoudi, K.; Onrubia, J.; Angel, T.; Boukra, A.; Guesnay, L.; Portas, A.; Barry-Martinet, R.; Angeletti, B.; Mounier, S.; Lenoble, V.; et al. Seawater Copper Content Controls Biofilm Bioaccumulation and Microbial Community on Microplastics. Sci. Total Environ. 2022, 814, 152278. [Google Scholar] [CrossRef]
- Liu, S.Y.; Leung, M.M.-L.; Fang, J.K.-H.; Chua, S.L. Engineering a Microbial ‘Trap and Release’ Mechanism for Microplastics Removal. Chem. Eng. J. 2021, 404, 127079. [Google Scholar] [CrossRef]
- Okeke, E.S.; Ezeorba, T.P.C.; Chen, Y.; Mao, G.; Feng, W.; Wu, X. Ecotoxicological and Health Implications of Microplastic-Associated Biofilms: A Recent Review and Prospect for Turning the Hazards into Benefits. Environ. Sci. Pollut. Res. 2022, 29, 70611–70634. [Google Scholar] [CrossRef]
- Kucia, M.; Wietrak, E.; Szymczak, M.; Kowalczyk, P. Effect of Ligilactobacillus Salivarius and Other Natural Components against Anaerobic Periodontal Bacteria. Molecules 2020, 25, 4519. [Google Scholar] [CrossRef]
- Zotta, T.; Ricciardi, A.; Ianniello, R.G.; Storti, L.V.; Glibota, N.A.; Parente, E. Aerobic and Respirative Growth of Heterofermentative Lactic Acid Bacteria: A Screening Study. Food Microbiol. 2018, 76, 117–127. [Google Scholar] [CrossRef]
- Nag, A.; Dhull, N.; Gupta, A. Evaluation of Tea (Camellia sinensis L.) Phytochemicals as Multi-Disease Modulators, a Multidimensional in Silico Strategy with the Combinations of Network Pharmacology, Pharmacophore Analysis, Statistics and Molecular Docking. Mol. Divers. 2023, 27, 487–509. [Google Scholar] [CrossRef]
- Chávez, F.; Aranda, M.; García, A.; Pastene, E. Antioxidant Polyphenols Extracted from Avocado Epicarp (Persea americana var. Hass) Inhibit Helicobacter Pylori Urease. Bol. Latinoam. Caribe Plantas Med. Aromat. 2011, 10, 265–280. [Google Scholar]
- Wong, X.; Madrid, A.M.; Tralma, K.; Castillo, R.; Carrasco-Pozo, C.; Navarrete, P.; Beltrán, C.; Pastene, E.; Gotteland, M. Polyphenol Extracts Interfere with Bacterial Lipopolysaccharide in Vitro and Decrease Postprandial Endotoxemia in Human Volunteers. J. Funct. Foods 2016, 26, 406–417. [Google Scholar] [CrossRef]
- Glavnik, V.; Simonovska, B.; Vovk, I. Densitometric Determination of (+)-Catechin and (−)-Epicatechin by 4-Dimethylaminocinnamaldehyde Reagent. J. Chromatogr. A 2009, 1216, 4485–4491. [Google Scholar] [CrossRef]
- Lu, Y.; Demleitner, M.F.; Song, L.; Rychlik, M.; Huang, D. Oligomeric Proanthocyanidins Are the Active Compounds in Abelmoschus Esculentus Moench for Its α-Amylase and α-Glucosidase Inhibition Activity. J. Funct. Foods 2016, 20, 463–471. [Google Scholar] [CrossRef]
- Fiehn, O.; Robertson, D.; Griffin, J.; vab der Werf, M.; Nikolau, B.; Morrison, N.; Sumner, L.W.; Goodacre, R.; Hardy, N.W.; Taylor, C.; et al. The Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 175–178. [Google Scholar] [CrossRef]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of Antiradical Properties of Antioxidants Using DPPH Assay: A Critical Review and Results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Castro-Seriche, S.; Jerez-Morales, A.; Smith, C.T.; Sánchez-Alonzo, K.; García-Cancino, A. Candida Albicans, a Reservoir of Listeria Monocytogenes? Infect. Genet. Evol. 2021, 90, 104779. [Google Scholar] [CrossRef]
- Adhikari, B.; Shah, P.K.; Karki, R. Antibiogram and Phytochemical Analysis of Cinnamon, Clove, and Sichuan Pepper Extracts. Nepal J. Biotechnol. 2021, 9, 1–7. [Google Scholar] [CrossRef]
- Hebel-Gerber, S.; García-Cancino, A.; Urbina, A.; Simirgiotis, M.J.; Echeverría, J.; Bustamante-Salazar, L.; Sáez-Carrillo, K.; Alarcón, J.; Pastene-Navarrete, E. Chilean Rhubarb, Gunnera Tinctoria (Molina) Mirb. (Gunneraceae): UHPLC-ESI-Orbitrap-MS Profiling of Aqueous Extract and Its Anti-Helicobacter Pylori Activity. Front. Pharmacol. 2021, 11, 583961. [Google Scholar] [CrossRef]
- Sierra-Quitian, A.G.; Hernandez-Moreno, L.V.; Pabon-Baquero, L.C.; Prieto-Rodriguez, J.A.; Patiño-Ladino, O.J. Antiquorum and Antibiofilm Activities of Piper bogotense C. DC. against Pseudomonas aeruginosa and Identification of Bioactive Compounds. Plants 2023, 12, 1901. [Google Scholar] [CrossRef]
- Shinde, S.; Lee, L.H.; Chu, T. Inhibition of Biofilm Formation by the Synergistic Action of EGCG-S and Antibiotics. Antibiotics 2021, 10, 102. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adduct | System | n-Hexane | EtOAc | MeOH | H2O | Mode | Partition Coefficient (KD) 1 |
|---|---|---|---|---|---|---|---|
| 1 | C | 1 | 9 | 1 | 9 | Ascending | 1.25 |
| 2 | C | 1 | 9 | 1 | 9 | Ascending | 1.11 |
| 3 | K | 1 | 2 | 1 | 1 | Ascending | 0.88 |
| 4 | K | 1 | 2 | 1 | 1 | Ascending | 2.61 |
| 5 | K | 1 | 2 | 1 | 2 | Ascending | 3.01 |
| 6 | K | 1 | 2 | 1 | 2 | Ascending | 4.06 |
| 7 | H | 1 | 3 | 1 | 3 | Ascending | 2.30 |
| 8 | K 2 | 0.5 | 1.5 | 0.5 | 1.2 | Descending | 1.92 |
| 9 | K | 1 | 2 | 1 | 2 | Ascending | 1.01 |
| Adduct | Avocado PACs (g) | Adduct Formed (mg) | Yield * (%) | Purity (%) |
|---|---|---|---|---|
| 1 | 1 | 218 | 21.8 | 98.7 |
| 2 | 1 | 222 | 22.2 | 98.2 |
| 3 | 1 | 180 | 18.0 | 96.1 |
| 4 | 3 | 450 | 15.0 | 95.4 |
| 5 | 2 | 440 | 22.0 | 94.0 |
| 6 | 1 | 221 | 22.1 | 97.3 |
| 7 | 1 | 179 | 17.9 | 93.5 |
| 8 | 2 | 313 | 15.7 | 97.2 |
| 9 | 0.7 | 99 | 14.1 | 98.1 |
| Compound | IC50 μM |
|---|---|
| 1 | 14.01 ± 2.27 |
| 2 | 22.12 ± 1.18 |
| 3 | 30.08 ± 3.01 |
| 4 | 25.32 ± 1.16 |
| 5 | 30.16 ± 2.02 |
| 6 | 13.11 ± 1.27 |
| 7 | 13.41 ± 1.14 |
| 8 | 25.51 ± 2.01 |
| 9 | 30.13 ± 2.17 |
| Epicatechin | 24.01 ± 2.02 |
| EGCG | 0.40 ± 1.01 |
| Trolox | 18.08 ± 3.52 |
| Strain | Inhibition Zone (Mean ± SD, mm) | ||||
|---|---|---|---|---|---|
| (1) 2 | (2) 2 | (3) 2 | AMX 1 | MTZ 1 | |
| L. fermentum UCO 979C | (-) | (-) | (-) | 23.7 ± 4.7 | 7.3 ± 1.2 |
| L. rhamnosus UCO 25A | (-) | (-) | (-) | 20.7 ± 0.6 | 7.3 ± 1.2 |
| L. monocytogenes 19115 | (-) | (-) | (-) | 26.3 ± 1.5 | 8.3 ± 1.0 |
| L. monocytogenes 7644 | (-) | (-) | (-) | 15.0 ± 1.0 | 7.7 ± 0.8 |
| E. coli 25922 | (-) | (-) | (-) | (-) | (-) |
| S. aureus 9144 | (-) | (-) | (-) | (-) | (-) |
| Strain | Inhibition Zone (Mean ± SD, mm) 1,2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (4) | (5) | (6) | (7) | (8) | (9) | AMX | MTZ | GEN | CFN | |
| L. fermentum UCO 979C | (-) | (-) | (-) | (-) | (-) | (-) | 11.3 ± 0.6 | 7.3 ±1.2 | 7.3 ± 1.0 | Nd |
| L. rhamnosus UCO 25A | (-) | (-) | (-) | (-) | (-) | (-) | 9.7 ± 0.6 | 7.3 ±1.2 | 7.3 ± 1.2 | Nd |
| L. monocytogenes 19115 | 15.7 ± 1.0 | 29.1 ± 0.8 | 16.0 ± 0.2 | 8.6 ± 0.4 | (-) | 22.1 ± 0.7 | 18.6 ± 0.8 | 7.7 ± 0.4 | 31 ± 0.6 | 30 ± 0.6 |
| L. monocytogenes 7644 | 15.0 ± 0.5 | 30.1 ± 0.8 | 16.1 ± 0.6 | 7.2 ± 0.4 | (-) | 19.2 ± 0.8 | 15.1 ± 0.8 | 8.3 ± 0.8 | 35 ± 0.6 | 34 ± 0.6 |
| S. aureus 9144 | 18.0 ± 0.6 | 17.1 ± 0.6 | 21.2 ± 0.4 | (-) | 14.1 ± 0.6 | 19.2 ± 1 | 27.8 ± 0.6 | Nd | 28 ± 0.6 | 35 ± 0.7 |
| E. coli 25922 | 10.2 ± 0.6 | 15.8 ± 0.6 | 14.7 ± 0.6 | 9.9 ± 0.6 | 8.6 ± 0.6 | 12.0 ± 0.6 | (-) | Nd | 22 ± 0.5 | 32 ± 0.6 |
| E. coli 11775 | 9.1 ± 0.6 | 17.3 ± 0.3 | 16.0 ± 0.6 | 11.1 ± 0.2 | 9.3 ± 0.6 | 11.4 ± 0.7 | 27.03 ± 0.6 | Nd | 25 ± 0.6 | 30 ± 0.6 |
| S. enterica 13076 | 14.4 ± 0.4 | 16.4 ± 0.4 | 17.2 ± 0.2 | 12.2 ± 0.2 | 11.0 ± 0.8 | 12.0 ± 0.6 | Nd | Nd | 25 ± 0.6 | 30 ± 0.6 |
| Strain | Viability 1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) | AMX 2 | MTZ 2 | |
| L. fermentum UCO 979C | (++) | (++) | (++) | (++) | (++) | (++) | (++) | (++) | (++) | (++) | (+) |
| L. monocytogenes 19115 | (++) | (++) | (++) | (++) | (+) | (−) | (++) | (++) | (+) | (++) | (+) |
| L. fermentum UCO 979C | (++) | (++) | (++) | (++) | (++) | (+) | (++) | (++) | (++) | (+) | (+) |
| L. monocytogenes 19115 | (++) | (++) | (++) | (+) | (−) | (−) | (++) | (++) | (−) | (+) | (+) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berrios-Henríquez, B.; Venegas-Toloza, M.; Reyes-Fuentes, M.; Zúñiga-Arbalti, F.; Bustamante, L.; García-Cancino, A.; Alarcón-Enos, J.; Pastene-Navarrete, E. Synthesis and Isolation of Phenol- and Thiol-Derived Epicatechin Adducts Prepared from Avocado Peel Procyanidins Using Centrifugal Partition Chromatography and the Evaluation of Their Antimicrobial and Antioxidant Activity. Molecules 2024, 29, 2872. https://doi.org/10.3390/molecules29122872
Berrios-Henríquez B, Venegas-Toloza M, Reyes-Fuentes M, Zúñiga-Arbalti F, Bustamante L, García-Cancino A, Alarcón-Enos J, Pastene-Navarrete E. Synthesis and Isolation of Phenol- and Thiol-Derived Epicatechin Adducts Prepared from Avocado Peel Procyanidins Using Centrifugal Partition Chromatography and the Evaluation of Their Antimicrobial and Antioxidant Activity. Molecules. 2024; 29(12):2872. https://doi.org/10.3390/molecules29122872
Chicago/Turabian StyleBerrios-Henríquez, Barbara, Matías Venegas-Toloza, María Reyes-Fuentes, Felipe Zúñiga-Arbalti, Luis Bustamante, Apolinaria García-Cancino, Julio Alarcón-Enos, and Edgar Pastene-Navarrete. 2024. "Synthesis and Isolation of Phenol- and Thiol-Derived Epicatechin Adducts Prepared from Avocado Peel Procyanidins Using Centrifugal Partition Chromatography and the Evaluation of Their Antimicrobial and Antioxidant Activity" Molecules 29, no. 12: 2872. https://doi.org/10.3390/molecules29122872
APA StyleBerrios-Henríquez, B., Venegas-Toloza, M., Reyes-Fuentes, M., Zúñiga-Arbalti, F., Bustamante, L., García-Cancino, A., Alarcón-Enos, J., & Pastene-Navarrete, E. (2024). Synthesis and Isolation of Phenol- and Thiol-Derived Epicatechin Adducts Prepared from Avocado Peel Procyanidins Using Centrifugal Partition Chromatography and the Evaluation of Their Antimicrobial and Antioxidant Activity. Molecules, 29(12), 2872. https://doi.org/10.3390/molecules29122872