
Comparative Analysis of the Biochemical Composition, Amino Acid, and Fatty Acid Contents of Diploid, Triploid, and Tetraploid Crassostrea gigas
 ,
,
Abstract
1. Introduction
2. Results
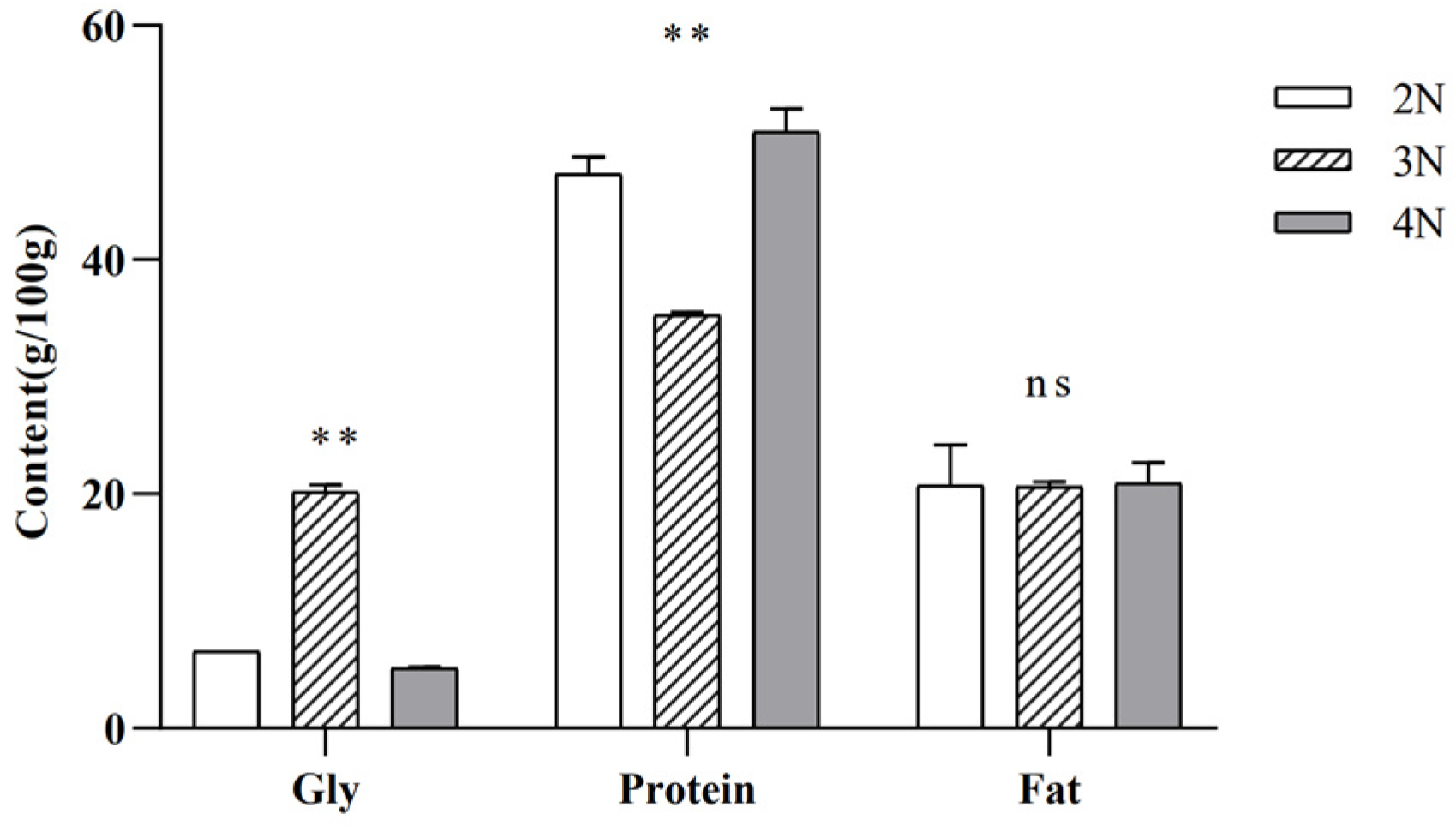
2.1. Differences in Glycogen, Protein, and Fat Contents among Diploid, Triploid, and Tetraploid Oysters
2.2. Differences in Glycogen, Protein, and Fat Contents between Males and Females
2.3. Differences in Amino Acid Contents in C. gigas with Different Ploidies and Sexes
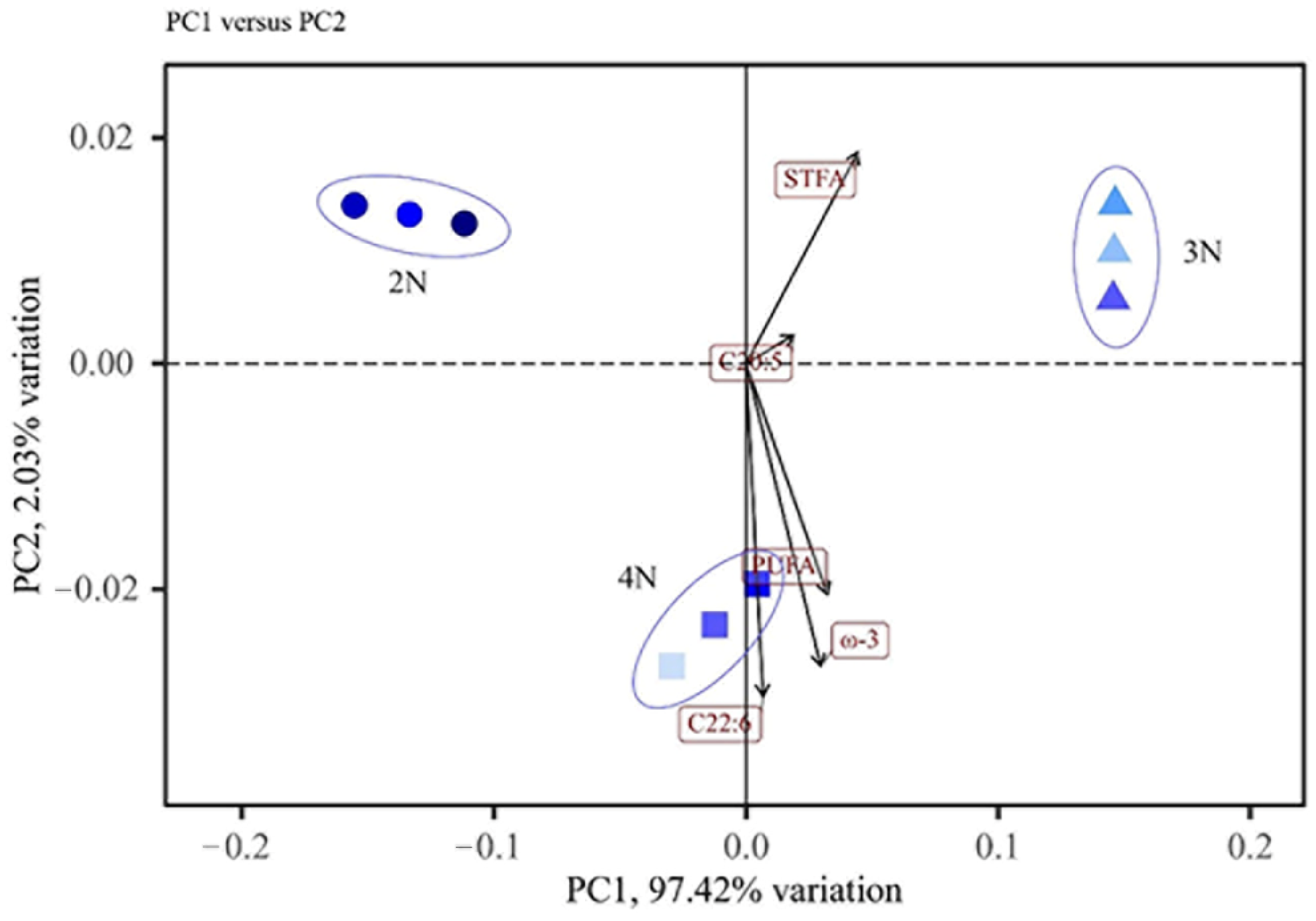
2.4. Differences in Fatty Acid Contents in C. gigas with Different Ploidies and Sexes
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Sample Preparation
4.3. Determination of Protein, Glycogen, and Fat Contents
4.4. Determination of Fatty Acid and Amino Acid Contents
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Allen, S.K.; Downing, S.L. Performance of triploid Pacific oysters, Crassostrea gigas (Thunberg). I. Survival, growth, glycogen content, and sexual maturation in yearlings. J. Exp. Mar. Biol. Ecol. 1986, 102, 197–208. [Google Scholar] [CrossRef]
- Maguire, G.B.; Gardner, N.C.; Nell, J.A.; Kent, G.N.; Kent, A.S. Studies on triploid oysters in Australia. 2. Growth, condition index gonad area, and glycogen content of triploid and diploid Pacific oysters, Crassostrea gigas, from oyster leases in Tasmania, Australia. Aquaculture 1995, 137, 357. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Wang, Z.; Yan, X.; Yu, Z. Phenotypic trait analysis of diploid and triploid hybrids from female Crassostrea hongkongensis × male C. gigas. Aquaculture 2014, 434, 307–314. [Google Scholar] [CrossRef]
- Yue, C.; Qin, Y.; Wan, W.; Shi, G.; Li, S.; Li, J.; Wang, Z.; Ma, H.; Li, J.; Yu, Z.; et al. Phenotypic traits of re-ciprocal tetraploid hybrids derived from tetraploid Crassostrea gigas and tetraploid Crassostrea angulata. Aquaculture 2024, 582, 740495. [Google Scholar] [CrossRef]
- Li, H.; Yu, R.; Li, Q. Effects of inbreeding on fertilization, growth and survival in the tetraploid Pacific oyster Crassostrea gigas. In Aquaculture International; SpringerLink: Berlin/Heidelberg, Germany, 2024; pp. 1–14. [Google Scholar] [CrossRef]
- Roac, P.J.; Depaoli-Roach, A.A.; Hurley, T.D.; Tagliabracci, V.S. Glycogen and its metabolism: Some new developments and old themes. Biochem. J. 2012, 441, 763–787. [Google Scholar] [CrossRef] [PubMed]
- Tabarini, C.L. Induced triploidy in the bay scallop, Argopecten irradians, and its effects on growth and gametogenesis. Aquaculture 1984, 42, 151–160. [Google Scholar] [CrossRef]
- Li, B.; Meng, J.; Li, L.; Liu, S.; Wang, T.; Zhang, G. Identification and Functional Characterization of the Glycogen Synthesis Related Gene Glycogenin in Pacific Oysters (Crassostrea gigas). J. Agric. Food Chem. 2017, 65, 7764–7773. [Google Scholar] [CrossRef] [PubMed]
- Manninen, H.; Rotola-Pukkila, M.; Aisala, H.; Hopia, A.; Laaksonen, T. Free amino acids and 5′-nucleotides in Finnish forest mushrooms. Food Chem. 2018, 247, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Qin, L.; Xue, J. Simultaneous quantification of free amino acids and 5′-nucleotides in shiitake mushrooms by stable isotope labeling-LC-MS/MS analysis. Food Chem. 2018, 268, 57–65. [Google Scholar] [CrossRef]
- Ojea, J.; Pazos, A.J.; Mart, D.; Novoa, S.; Sánchez, J.L.; Abad, M. Seasonal variation in weight and biochemical composition of the tissues of Ruditapes decussatus in relation to the gametogenic cycle. Aquaculture 2004, 238, 451–468. [Google Scholar] [CrossRef]
- Fuke, S. Taste-Active Components of Seafoods with Special Reference to Umami Substances; Blackie Academic: Glasgow, UK, 1994; pp. 115–139. [Google Scholar]
- Gao, J.X.; Zhang, Y.Y.; Huang, X.H.; Liu, R.; Dong, X.P.; Zhu, B.W.; Qin, L. Comparison of amino acid, 5′-nucleotide and lipid metabolism of oysters (Crassostrea gigas Thunberg) captured in different seasons. Food Res. Int. 2021, 147, 110560. [Google Scholar] [CrossRef] [PubMed]
- Appleton, K.M.; Rogers, P.J.; Ness, A.R. Updated systematic review and meta-analysis of the effects of n-3 long-chain polyunsaturated fatty acids on depressed mood. Am. J. Clin. Nutr. 2010, 91, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, K.M.; Wheeler, D.C.; Mucci, L.A. Fish consumption and prostate cancer risk: A review and meta-analysis. Am. J. Clin. Nutr. 2010, 92, 1223. [Google Scholar] [CrossRef]
- Li, H.; Song, Y.; Zhang, H.; Wang, X.; Xue, C. Comparative lipid profile of four edible shellfishes by UPLC-Triple TOF-MS/MS. Food Chem. 2019, 310, 125947. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zhang, Y.; Ma, H.; Wu, X.; Xiao, S.; Li, J.; Mo, R.; Yu, Z. Comparison of the biochemical composition and nutritional quality between diploid and triploid Hong Kong oysters, Crassostrea hongkongensis. Front. Physiol. 2018, 9, 1674. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Li, R.; Liao, Q.; Shi, G.; Zhou, Y.; Wan, W.; Li, J.; Ma, H.; Zhang, Y.; Yu, Z. Comparison of biochemical composition, nutritional quality, and metals concentrations between males and females of three different Crassostrea sp. Food Chem. 2022, 398, 133868. [Google Scholar] [CrossRef] [PubMed]
- Karsoon, T.; Helu, L.; Hongkuan, Z.; Shengkang, L.; Huaiping, Z. Carotenoids content and lipid nutritional quality variation in tissues of male and female polymorphic (golden and brown) noble scallops Chlamys nobilis. Aquaculture 2021, 536, 736483. [Google Scholar] [CrossRef]
- Randle, P.J.; Garland, P.B.; Hales, C.N.; Newsholme, E.A. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1963, 1, 785–789. [Google Scholar] [CrossRef]
- Pogoda, B.; Buck, B.H.; Saborowski, R.; Hagen, W. Biochemical and elemental composition of the offshore-cultivated oysters Ostrea edulis and Crassostrea gigas. Aquaculture 2013, 400, 53–60. [Google Scholar] [CrossRef]
- Sheng, L.; Wei, W.; Busu, L.; Guofan, Z. Characterization, fluctuation and tissue differences in nutrient content in the Pacific oyster (Crassostrea gigas) in Qingdao, northern China. Aquac. Res. 2020, 51, 14463. [Google Scholar]
- Dridi, S.; Romdhane, M.S.; Elcafsi, M.H. Seasonal variation in weight and biochemical composition of the Pacific oyster, Crassostrea gigas in relation to the gametogenic cycle and environmental conditions of the Bizert lagoon, Tunisia. Aquaculture 2007, 263, 238–248. [Google Scholar] [CrossRef]
- Barber, B.J.; Blake, N.J. Energy storage and utilization in relation to gametogenesis in Argopecten irradians concentricus (say). J. Exp. Mar. Biol. Ecol. 1981, 52, 121–134. [Google Scholar] [CrossRef]
- Epp, J.; Bricelj, V.M.; Malouf, R.E. Seasonal partitioning and utilisation of energy reserves in two age classes of the bay scallop Argopecten irradians irradians (Lamarck). J. Exp. Mar. Biol. Ecol. 1988, 121, 113–136. [Google Scholar] [CrossRef]
- Akashige, S.; Fushimi, T. Growth, Survival, and Glycogen Content of Triploid Pacific Oyster Crassostrea gigas in the Waters of Hiroshima, Japan. Nihon-Suisan-Gakkai-Shi 1992, 58, 1063–1071. [Google Scholar] [CrossRef]
- Ruiz-Verdugo, C.A.; Racotta, I.S.; Ibarra, A.M. Comparative biochemical composition in gonad and adductor muscle of triploid and diploid catarina scallop (Argopecten ventricosus Sowerby II, 1842). J. Exp. Mar. Biol. Ecol. 2001, 259, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-C.; Liu, H.-H.; Su, X.-R.; Li, T.-W. Fatty Acid Levels in 7 Economic Shellfishes. Fish. Sci. 2005, 24, 20–22. [Google Scholar]
- Gabbott, P.A. Storage cycles in marine bivalve molluscs: A hypothesis concerning the relationship between glycogen metabolism and gametogenesis. In Ninth European Marine Biology; Aberdeen University Press: Aberdeen, UK, 1975; pp. 191–211. [Google Scholar]
- Guo, X.; Allen, S.K. Sex and meiosis in autotetraploid Pacific oyster, Crassostrea gigas (Thunberg). Genome 1997, 40, 397. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; Qin, Y.; Shi, G.; Li, S.; Liao, Q.; Ma, H.; Li, J.; Suo, A.; Ding, D.; Yu, Z.; et al. Genetic improvement of aquaculture performance for tetraploid Pacific oysters, Crassostrea gigas: A case study of four consecutive generations of selective breeding. Aquaculture 2023, 563, 738910. [Google Scholar] [CrossRef]
- Beninger, P.G.; Pennec, P. Demonstration of nutrient pathway from the digestive system to oocytes in the gonad intestinal loop of the scallop Pecten maximus L. Biol Bull. 2003, 205, 83–92. [Google Scholar] [CrossRef]
- Gallager, S.M.; Mann, R. Growth and survival of larvae of Mercenaria mercenaria (L.) and Crassostrea virginica (Gmelin) relative to broodstock conditioning and lipid content of eggs. Aquaculture 1986, 56, 105–121. [Google Scholar] [CrossRef]
- Li, Q.; Osada, M.; Mori, K. Seasonal biochemical variations in Pacific oyster gonadal tissue during sexual maturation. Fish. Sci. 2000, 66, 502–508. [Google Scholar] [CrossRef]
- Pollero, R.J.; Ré, M.E.; Brenner, R.R. Seasonal changes of the lipids of the mollusc Chlamys tehuelcha. Comp. Biochem. Physiol. Part A Physiol. 1979, 64, 257–263. [Google Scholar] [CrossRef]
- Berthelin, C.; Kellner, K.; Mathieu, M. Storage metabolism in the Pacific oyster (Crassostrea gigas) in relation to summer mortalities and reproductive cycle (West Coast of France). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2000, 125, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Unuma, T.; Suzuki, T.; Kurokawa, T.; Akiyama, Y.T. A Protein Identical to the Yolk Protein Is Stored in the Testis in Male Red Sea Urchin, Pseudocentrotus depressus. Biol. Bull. 1998, 194, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Spindler, M.; Stadler, R.; Tanner, H. Amino acid analysis of feedstuffs: Determination of methionine and cystine after oxidation with performic acid and hydrolysis. J. Agric. Food Chem. 1984, 32, 1366–1371. [Google Scholar] [CrossRef]
- Wang, L.; Folsom, A.R.; Zheng, Z.J.; Pankow, J.S.; Eckfeldt, J.H. Plasma fatty acid composition and 6-year incidence of hypertension in middle-aged adults: The Atherosclerosis Risk in Communities (ARIC) Study. Am. J. Clin. Nutr. 2003, 78, 91–98. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, J.; Pei, Z.; Wei, P.; Xiang, D.; Cao, X.; Shen, X.; Li, C. Volatile flavour components and the mechanisms underlying their production in golden pompano (Trachinotus blochii) fillets subjected to different drying methods: A comparative study using an electronic nose, an electronic tongue and SDE-GC-MS. Food Res. 2019, 123, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.W.; Zhang, M.; Shrestha, S. Compositional characteristics and nutritional quality of Chinese mitten crab (Eriocheir sinensis). Food Chem. 2007, 103, 1343–1349. [Google Scholar] [CrossRef]
- GB5009.124-2016; National Health and Family Planning Commission of the People’s Republic of China. National food safety standard—Determination of Amino Acids in Foods. National Health and Family Planning Commission of the People’s Republic of China: Beijing, China, 2016.
- GB5009.168-2016; National Health and Family Planning Commission of the People’s Republic of China. National Food Safety standard—Determination of Fatty Acids in Foods. National Health and Family Planning Commission of the People’s Republic of China: Beijing, China, 2016.
- World Health Organization; Food and Agriculture Organization; United Nations University. Energy and protein requirements Report of a Joint FAO/WHO/UNU Expert Consultation. WHO Tech. Rep. Ser. 1985, 724, 1–206. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Amino Acid | 2N | 3N | 4N |
|---|---|---|---|
| Aspartic acid + | 0.85 ± 0.05 a | 0.74 ± 0.01 b | 0.88 ± 0.01 a |
| Threonine * | 0.44 ± 0.01 a | 0.38 ± 0.01 b | 0.47 ± 0.01 a |
| Serine | 0.47 ± 0.01 a | 0.36 ± 0.02 b | 0.47 ± 0.02 a |
| Glutamic acid + | 1.21 ± 0.01 b | 1.15 ± 0.04 b | 1.29 ± 0.01 a |
| Glycine + | 0.61 ± 0.01 a | 0.49 ± 0.02 b | 0.68 ± 0.03 a |
| Alanine + | 0.54 ± 0.01 ab | 0.49 ± 0.03 b | 0.61 ± 0.02 a |
| Valine * | 0.46 ± 0.00 a | 0.40 ± 0.01 b | 0.50 ± 0.02 a |
| Methionine * | 0.26 ± 0.01 a | 0.23 ± 0.01 a | 0.26 ± 0.01 a |
| Isoleucine * | 0.44 ± 0.01 a | 0.38 ± 0.01 b | 0.46 ± 0.01 a |
| Leucine * | 0.68 ± 0.01 a | 0.54 ± 0.00 b | 0.70 ± 0.01 a |
| Tyrosine + | 0.34 ± 0.01 a | 0.28 ± 0.01 b | 0.33 ± 0.01 a |
| Phenylalanine *+ | 0.42 ± 0.01 a | 0.34 ± 0.00 b | 0.42 ± 0.01 a |
| Lysine * | 0.74 ± 0.02 a | 0.56 ± 0.01 b | 0.78 ± 0.02 a |
| Histidine * | 0.23 ± 0.00 a | 0.21 ± 0.01 b | 0.24 ± 0.01 a |
| Arginine | 0.61 ± 0.04 a | 0.44 ± 0.02 b | 0.65 ± 0.04 a |
| Proline | 0.44 ± 0.01 b | 0.50 ± 0.01 a | 0.49 ± 0.00 a |
| TAA | 8.67 ± 0.11 b | 7.45 ± 0.01 c | 9.16 ± 0.01 a |
| EAA | 3.64 ± 0.02 b | 3.03 ± 0.06 c | 3.79 ± 0.03 a |
| DAA | 3.95 ± 0.02 b | 3.48 ± 0.01 c | 4.19 ± 0.00 a |
| Amino Acid | 2N | 4N | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Aspartic acid + | 0.74 ± 0.04 a | 0.95 ± 0.06 a | 0.82 ± 0.01 b | 0.95 ± 0.01 a |
| Threonine * | 0.40 ± 0.01 b | 0.48 ± 0.01 a | 0.46 ± 0.01 a | 0.49 ± 0.01 a |
| Serine | 0.40 ± 0.01 b | 0.54 ± 0.01 a | 0.44 ± 0.01 a | 0.49 ± 0.03 a |
| Glutamic acid + | 1.08 ± 0.01 b | 1.35 ± 0.03 a | 1.21 ± 0.01 b | 1.38 ± 0.04 a |
| Glycine + | 0.71 ± 0.04 a | 0.51 ± 0.01 b | 0.79 ± 0.04 a | 0.57 ± 0.02 b |
| Alanine + | 0.58 ± 0.01 a | 0.50 ± 0.01 b | 0.67 ± 0.02 a | 0.54 ± 0.02 b |
| Valine * | 0.42 ± 0.00 b | 0.49 ± 0.00 a | 0.49 ± 0.02 a | 0.50 ± 0.01 a |
| Methionine * | 0.22 ± 0.01 b | 0.29 ± 0.01 a | 0.22 ± 0.01 b | 0.28 ± 0.00 a |
| Isoleucine * | 0.40 ± 0.01 b | 0.48 ± 0.01 a | 0.45 ± 0.02 a | 0.48 ± 0.01 a |
| Leucine * | 0.62 ± 0.01 b | 0.72 ± 0.03 a | 0.68 ± 0.01 a | 0.72 ± 0.01 a |
| Tyrosine + | 0.30 ± 0.01 b | 0.38 ± 0.01 a | 0.31 ± 0.01 a | 0.35 ± 0.01 a |
| Phenylalanine *+ | 0.36 ± 0.01 b | 0.47 ± 0.00 a | 0.38 ± 0.00 b | 0.45 ± 0.01 a |
| Lysine * | 0.72 ± 0.01 a | 0.75 ± 0.03 a | 0.79 ± 0.04 a | 0.75 ± 0.00 a |
| Histidine * | 0.22 ± 0.01 b | 0.24 ± 0.00 a | 0.23 ± 0.00 a | 0.24 ± 0.01 a |
| Arginine | 0.62 ± 0.03 a | 0.58 ± 0.04 a | 0.70 ± 0.04 a | 0.59 ± 0.02 a |
| Proline | 0.42 ± 0.01 a | 0.46 ± 0.02 a | 0.47 ± 0.00 a | 0.51 ± 0.00 a |
| TAA | 8.17 ± 0.00 b | 9.16 ± 0.22 a | 9.08 ± 0.02 a | 9.24 ± 0.06 a |
| EAA | 3.35 ± 0.02 b | 3.92 ± 0.06 a | 3.69 ± 0.04 b | 3.88 ± 0.02 a |
| DAA | 3.75 ± 0.04 a | 4.14 ± 0.08 a | 4.16 ± 0.04 a | 4.22 ± 0.04 a |
| Amino Acid | 2N | 3N | 4N |
|---|---|---|---|
| Threonine | 126.87 | 127.52 | 128.28 |
| Valine | 106.11 | 94.67 | 109.17 |
| Methionine | 85.68 | 88.21 | 81.10 |
| Isoleucine | 126.87 | 127.52 | 125.55 |
| Leucine | 112.04 | 103.55 | 109.17 |
| Phenylalanine + Tyrosine | 146.10 | 138.70 | 136.46 |
| Lysine | 155.19 | 136.67 | 154.82 |
| Amino Acid | 2N | 4N | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Threonine | 122.40 | 131.00 | 126.65 | 132.58 |
| Valine | 102.82 | 106.99 | 107.93 | 108.23 |
| Methionine | 76.94 | 90.46 | 69.23 | 86.58 |
| Isoleucine | 122.40 | 131.00 | 123.90 | 129.87 |
| Leucine | 108.41 | 112.29 | 106.99 | 111.32 |
| Phenylalanine + Tyrosine | 134.64 | 154.66 | 126.65 | 144.30 |
| Lysine | 160.23 | 148.87 | 158.19 | 147.58 |
| Fatty Acid | 2N | 3N | 4N |
|---|---|---|---|
| C14:0 | 0.117 ± 0.00 b | 0.110 ± 0.00 c | 0.123 ± 0.00 a |
| C16:0 | 0.534 ± 0.00 a | 0.510 ± 0.00 b | 0.512 ± 0.00 b |
| C16:1 | 0.130 ± 0.00 a | 0.107 ± 0.00 c | 0.114 ± 0.00 b |
| C17:0 | 0.024 ± 0.00 c | 0.030 ± 0.00 a | 0.025 ± 0.00 b |
| C18:0 | 0.079 ± 0.00 c | 0.090 ± 0.00 a | 0.083 ± 0.00 b |
| C18:1 | 0.211 ± 0.01 c | 0.290 ± 0.00 a | 0.232 ± 0.00 b |
| C18:2 | 0.045 ± 0.00 b | 0.050 ± 0.00 a | 0.046 ± 0.00 b |
| C18:3 | 0.076 ± 0.00 c | 0.090 ± 0.00 a | 0.083 ± 0.01 b |
| C20:4 (ARA) | 0.025 ± 0.00 b | 0.030 ± 0.00 a | 0.026 ± 0.00 b |
| C20:5 (EPA) | 0.296 ± 0.01 c | 0.370 ± 0.00 a | 0.327 ± 0.00 b |
| C22:6 (DHA) | 0.262 ± 0.00 b | 0.290 ± 0.00 a | 0.289 ± 0.00 a |
| SFA | 0.759 ± 0.01 a | 0.750 ± 0.00 a | 0.755 ± 0.01 a |
| MUFA | 0.325 ± 0.00 b | 0.357 ± 0.00 a | 0.326 ± 0.00 b |
| PUFA | 0.705 ± 0.01 c | 0.832 ± 0.00 a | 0.770 ± 0.01 b |
| Σω-3 | 0.635 ± 0.01 c | 0.752 ± 0.00 a | 0.699 ± 0.01 b |
| Σω-6 | 0.070 ± 0.00 b | 0.081 ± 0.00 a | 0.071 ± 0.00 b |
| STFA | 1.804 ± 0.02 c | 1.98 ± 0.00 a | 1.870 ± 0.02 b |
| Fatty Acid | 2N | 4N | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| C14:0 | 0.05 ± 0.01 b | 0.19 ± 0.00 a | 0.06 ± 0.00 b | 0.19 ± 0.01 a |
| C16:0 | 0.31 ± 0.01 b | 0.76 ± 0.00 a | 0.36 ± 0.00 b | 0.66 ± 0.01 a |
| C17:0 | 0.02 ± 0.00 b | 0.03 ± 0.00 a | 0.02 ± 0.00 a | 0.03 ± 0.00 a |
| C18:0 | 0.05 ± 0.01 b | 0.10 ± 0.00 a | 0.07 ± 0.01 b | 0.10 ± 0.00 a |
| C16:1 | 0.04 ± 0.01 b | 0.22 ± 0.00 a | 0.06 ± 0.00 b | 0.17 ± 0.00 a |
| C18:1 | 0.13 ± 0.00 b | 0.29 ± 0.00 a | 0.18 ± 0.00 b | 0.28 ± 0.01 a |
| C18:2 | 0.03 ± 0.00 b | 0.06 ± 0.00 a | 0.03 ± 0.00 b | 0.06 ± 0.00 a |
| C18:3 | 0.04 ± 0.00 b | 0.11 ± 0.00 a | 0.06 ± 0.00 b | 0.11 ± 0.00 a |
| C20:4 (ARA) | 0.02 ± 0.00 b | 0.03 ± 0.00 a | 0.02 ± 0.00 a | 0.03 ± 0.00 a |
| C20:5 (EPA) | 0.19 ± 0.01 b | 0.40 ± 0.00 a | 0.25 ± 0.00 b | 0.40 ± 0.01 a |
| C22:6 (DHA) | 0.22 ± 0.01 b | 0.31 ± 0.00 a | 0.26 ± 0.00 b | 0.31 ± 0.01 a |
| SFA | 0.42 ± 0.01 b | 1.10 ± 0.01 a | 0.52 ± 0.01 b | 0.99 ± 0.02 a |
| MUFA | 0.16 ± 0.00 b | 0.49 ± 0.00 a | 0.22 ± 0.00 b | 0.43 ± 0.01 a |
| PUFA | 0.49 ± 0.02 b | 0.92 ± 0.01 a | 0.63 ± 0.01 b | 0.91 ± 0.03 a |
| Σω-3 | 0.45 ± 0.02 b | 0.82 ± 0.00 a | 0.58 ± 0.01 b | 0.82 ± 0.03 a |
| Σω-6 | 0.05 ± 0.00 b | 0.09 ± 0.00 a | 0.05 ± 0.01 b | 0.09 ± 0.00 a |
| STFA | 1.09 ± 0.04 b | 2.52 ± 0.01 a | 1.39 ± 0.02 b | 2.35 ± 0.06 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, J.; Zhang, E.; Yu, W.; Wang, W.; Sun, Y.; Dong, L.; Zhang, Y.; Sun, G.; Li, Z.; Luo, Q.; et al. Comparative Analysis of the Biochemical Composition, Amino Acid, and Fatty Acid Contents of Diploid, Triploid, and Tetraploid Crassostrea gigas. Molecules 2024, 29, 2671. https://doi.org/10.3390/molecules29112671
Fu J, Zhang E, Yu W, Wang W, Sun Y, Dong L, Zhang Y, Sun G, Li Z, Luo Q, et al. Comparative Analysis of the Biochemical Composition, Amino Acid, and Fatty Acid Contents of Diploid, Triploid, and Tetraploid Crassostrea gigas. Molecules. 2024; 29(11):2671. https://doi.org/10.3390/molecules29112671
Chicago/Turabian StyleFu, Jingjing, Enshuo Zhang, Wensong Yu, Weijun Wang, Youmei Sun, Luyao Dong, Yousen Zhang, Guohua Sun, Zan Li, Qihao Luo, and et al. 2024. "Comparative Analysis of the Biochemical Composition, Amino Acid, and Fatty Acid Contents of Diploid, Triploid, and Tetraploid Crassostrea gigas" Molecules 29, no. 11: 2671. https://doi.org/10.3390/molecules29112671
APA StyleFu, J., Zhang, E., Yu, W., Wang, W., Sun, Y., Dong, L., Zhang, Y., Sun, G., Li, Z., Luo, Q., & Yang, J. (2024). Comparative Analysis of the Biochemical Composition, Amino Acid, and Fatty Acid Contents of Diploid, Triploid, and Tetraploid Crassostrea gigas. Molecules, 29(11), 2671. https://doi.org/10.3390/molecules29112671