

Myricetin Acts as an Inhibitor of Type II NADH Dehydrogenase from Staphylococcus aureus

 , ,
, ,
Abstract
1. Introduction
2. Results
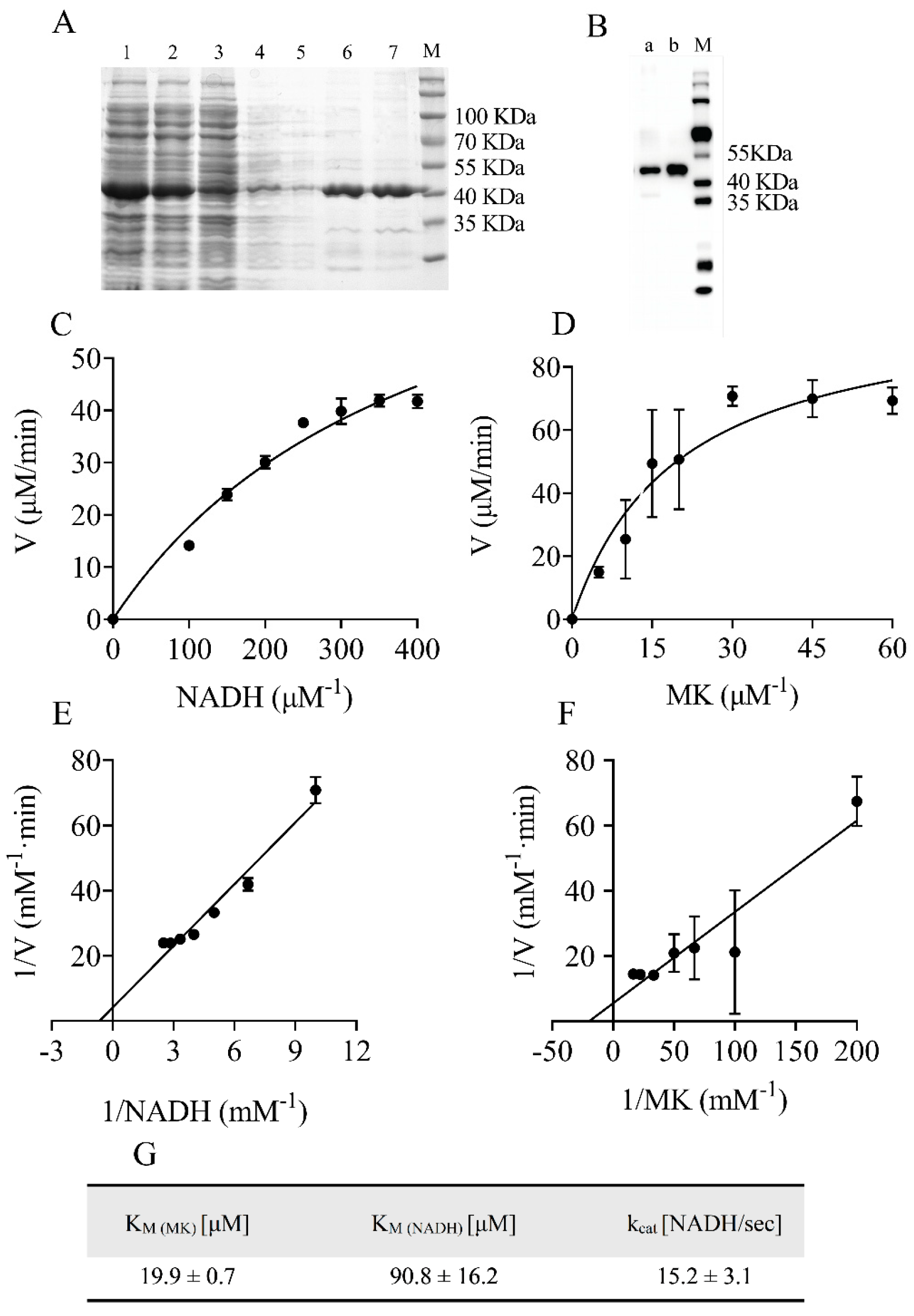
2.1. Protein Purification and Enzymatic Characterization
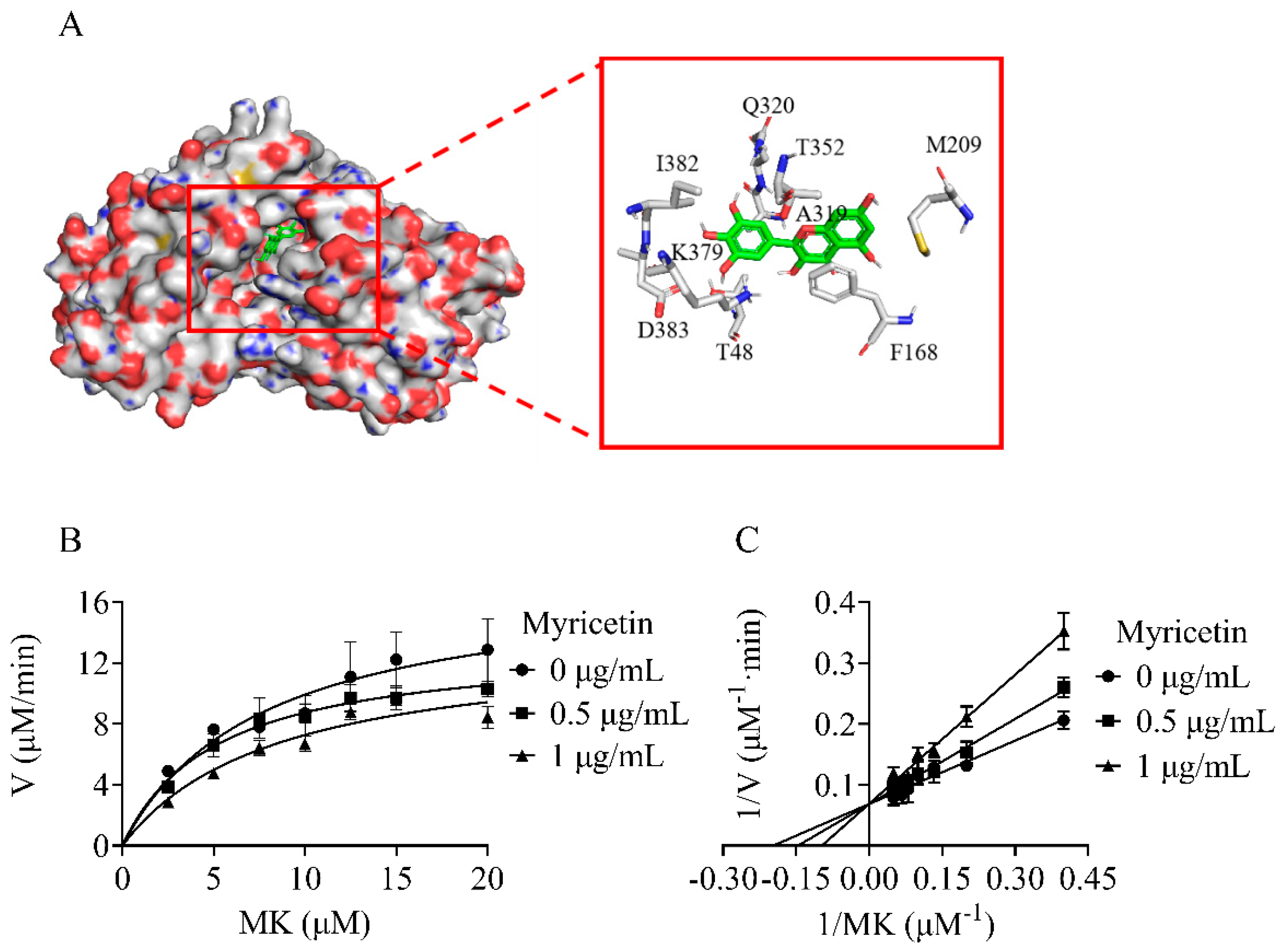
2.2. Myricetin Acted as a Competitive Inhibitor of NDH-2 Substrate Menadione
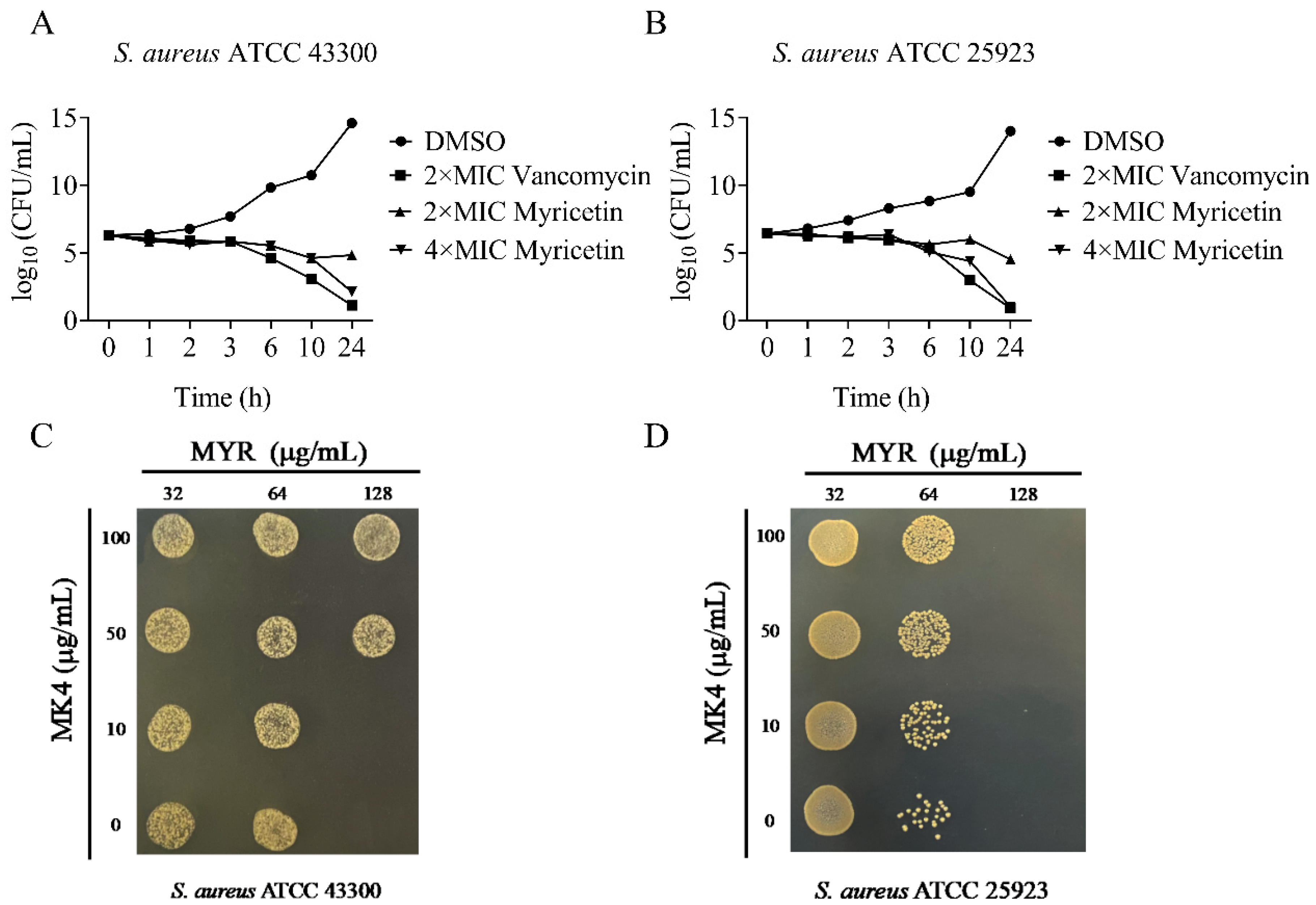
2.3. The Anti-Staphylococcal Activity of Myricetin Was Antagonized by MK
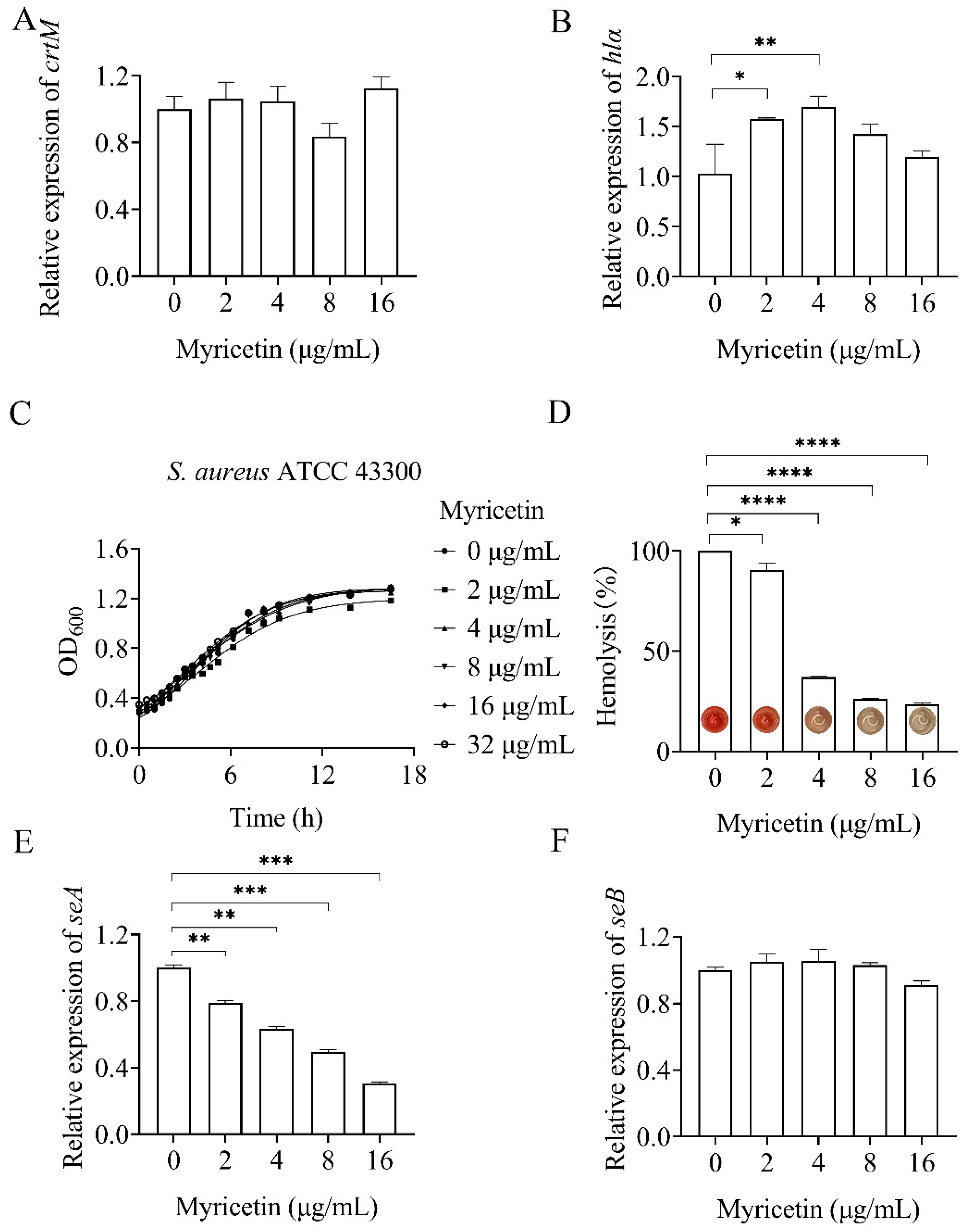
2.4. Myricetin Exhibited a Complex Impact on the Virulence Factors of S. aureus
3. Discussion
4. Methods
4.1. Strains and Chemicals
4.2. Expression Plasmid Construction
4.3. Protein Preparation
4.4. Enzyme Activity Assay
4.5. Molecular Docking Simulation
4.6. Antimicrobial Activity Assays
4.7. Quantitative Polymerase Chain Reaction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wertheim, H.F.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Howden, B.P.; Giulieri, S.G.; Wong Fok Lung, T.; Baines, S.L.; Sharkey, L.K.; Lee, J.Y.H.; Hachani, A.; Monk, I.R.; Stinear, T.P. Staphylococcus aureus host interactions and adaptation. Nat. Rev. Microbiol. 2023, 21, 380–395. [Google Scholar] [CrossRef] [PubMed]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Hennekinne, J.A.; De Buyser, M.L.; Dragacci, S. Staphylococcus aureus and its food poisoning toxins: Characterization and outbreak investigation. FEMS Microbiol. Rev. 2012, 36, 815–836. [Google Scholar] [CrossRef] [PubMed]
- Sellamuthu, S.; Singh, M.; Kumar, A.; Singh, S.K. Type-II NADH Dehydrogenase (NDH-2): A promising therapeutic target for antitubercular and antibacterial drug discovery. Expert. Opin. Ther. Targets 2017, 21, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Schurig-Briccio, L.A.; Yano, T.; Rubin, H.; Gennis, R.B. Characterization of the type 2 NADH: Menaquinone oxidoreductases from Staphylococcus aureus and the bactericidal action of phenothiazines. Biochim. Biophys. Acta 2014, 1837, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Schurig-Briccio, L.A.; Parraga Solorzano, P.K.; Lencina, A.M.; Radin, J.N.; Chen, G.Y.; Sauer, J.D.; Kehl-Fie, T.E.; Gennis, R.B. Role of respiratory NADH oxidation in the regulation of Staphylococcus aureus virulence. EMBO Rep. 2020, 21, e45832. [Google Scholar] [CrossRef]
- Weinstein, E.A.; Yano, T.; Li, L.S.; Avarbock, D.; Avarbock, A.; Helm, D.; McColm, A.A.; Duncan, K.; Lonsdale, J.T.; Rubin, H. Inhibitors of type II NADH: Menaquinone oxidoreductase represent a class of antitubercular drugs. Proc. Natl. Acad. Sci. USA 2005, 102, 4548–4553. [Google Scholar] [CrossRef]
- Dunn, E.A.; Roxburgh, M.; Larsen, L.; Smith, R.A.; McLellan, A.D.; Heikal, A.; Murphy, M.P.; Cook, G.M. Incorporation of triphenylphosphonium functionality improves the inhibitory properties of phenothiazine derivatives in Mycobacterium tuberculosis. Bioorg Med. Chem. 2014, 22, 5320–5328. [Google Scholar] [CrossRef] [PubMed]
- Nizi, M.G.; Desantis, J.; Nakatani, Y.; Massari, S.; Mazzarella, M.A.; Shetye, G.; Sabatini, S.; Barreca, M.L.; Manfroni, G.; Felicetti, T.; et al. Antitubercular polyhalogenated phenothiazines and phenoselenazine with reduced binding to CNS receptors. Eur. J. Med. Chem. 2020, 201, 112420. [Google Scholar] [CrossRef]
- Heikal, A.; Hards, K.; Cheung, C.Y.; Menorca, A.; Timmer, M.S.; Stocker, B.L.; Cook, G.M. Activation of type II NADH dehydrogenase by quinolinequinones mediates antitubercular cell death. J. Antimicrob. Chemother. 2016, 71, 2840–2847. [Google Scholar] [CrossRef]
- Hong, W.D.; Gibbons, P.D.; Leung, S.C.; Amewu, R.; Stocks, P.A.; Stachulski, A.; Horta, P.; Cristiano, M.L.S.; Shone, A.E.; Moss, D.; et al. Rational Design, Synthesis, and Biological Evaluation of Heterocyclic Quinolones Targeting the Respiratory Chain of Mycobacterium tuberculosis. J. Med. Chem. 2017, 60, 3703–3726. [Google Scholar] [CrossRef]
- Shirude, P.S.; Paul, B.; Roy Choudhury, N.; Kedari, C.; Bandodkar, B.; Ugarkar, B.G. Quinolinyl Pyrimidines: Potent Inhibitors of NDH-2 as a Novel Class of Anti-TB Agents. ACS Med. Chem. Lett. 2012, 3, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Åkerbladh, L.; Ahmad, S.; Konda, V.; Cao, S.; Vocat, A.; Maes, L.; Cole, S.T.; Hughes, D.; Larhed, M.; et al. Synthesis and In Vitro Biological Evaluation of Quinolinyl Pyrimidines Targeting Type II NADH-Dehydrogenase (NDH-2). ACS Infect. Dis. 2022, 8, 482–498. [Google Scholar] [CrossRef]
- Nguyen, N.; Wilson, D.W.; Nagalingam, G.; Triccas, J.A.; Schneider, E.K.; Li, J.; Velkov, T.; Baell, J. Broad activity of diphenyleneiodonium analogues against Mycobacterium tuberculosis, malaria parasites and bacterial pathogens. Eur. J. Med. Chem. 2018, 148, 507–518. [Google Scholar] [CrossRef]
- Dam, S.; Tangara, S.; Hamela, C.; Hattabi, T.; Faïon, L.; Carre, P.; Antoine, R.; Herledan, A.; Leroux, F.; Piveteau, C.; et al. Tricyclic SpiroLactams Kill Mycobacteria in vitro and in vivo by Inhibiting Type II NADH Dehydrogenases. J. Med. Chem. 2022, 65, 16651–16664. [Google Scholar] [CrossRef] [PubMed]
- Sena, F.V.; Batista, A.P.; Catarino, T.; Brito, J.A.; Archer, M.; Viertler, M.; Madl, T.; Cabrita, E.J.; Pereira, M.M. Type-II NADH:quinone oxidoreductase from Staphylococcus aureus has two distinct binding sites and is rate limited by quinone reduction. Mol. Microbiol. 2015, 98, 272–288. [Google Scholar] [CrossRef]
- Vamshi Krishna, K.; Venkata Mohan, S. Purification and Characterization of NDH-2 Protein and Elucidating Its Role in Extracellular Electron Transport and Bioelectrogenic Activity. Front. Microbiol. 2019, 10, 880. [Google Scholar] [CrossRef]
- Lencina, A.M.; Franza, T.; Sullivan, M.J.; Ulett, G.C.; Ipe, D.S.; Gaudu, P.; Gennis, R.B.; Schurig-Briccio, L.A. Type 2 NADH Dehydrogenase Is the Only Point of Entry for Electrons into the Streptococcus agalactiae Respiratory Chain and Is a Potential Drug Target. mBio 2018, 9, e01034-18. [Google Scholar] [CrossRef] [PubMed]
- Petri, J.; Shimaki, Y.; Jiao, W.; Bridges, H.R.; Russell, E.R.; Parker, E.J.; Aragão, D.; Cook, G.M.; Nakatani, Y. Structure of the NDH-2—HQNO inhibited complex provides molecular insight into quinone-binding site inhibitors. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.N.; Da Hora, G.C.A.; Soares, T.A.; Bojer, M.S.; Ingmer, H.; Macedo, A.J.; Trentin, D.S. Myricetin protects Galleria mellonella against Staphylococcus aureus infection and inhibits multiple virulence factors. Sci. Rep. 2017, 7, 2823. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Saeed, F.; Hussain, G.; Imran, A.; Mehmood, Z.; Gondal, T.A.; El-Ghorab, A.; Ahmad, I.; Pezzani, R.; Arshad, M.U.; et al. Myricetin: A comprehensive review on its biological potentials. Food Sci. Nutr. 2021, 9, 5854–5868. [Google Scholar] [CrossRef] [PubMed]
- Taheri, Y.; Suleria, H.A.R.; Martins, N.; Sytar, O.; Beyatli, A.; Yeskaliyeva, B.; Seitimova, G.; Salehi, B.; Semwal, P.; Painuli, S.; et al. Myricetin bioactive effects: Moving from preclinical evidence to potential clinical applications. BMC Complement. Med. Ther. 2020, 20, 241. [Google Scholar] [CrossRef]
- Yadav, A.K.; Thakur, J.; Prakash, O.; Khan, F.; Saikia, D.; Gupta, M.M. Screening of flavonoids for antitubercular activity and their structure–activity relationships. Med. Chem. Res. 2013, 22, 2706–2716. [Google Scholar] [CrossRef]
- Hu, P.; Lv, B.; Yang, K.; Lu, Z.; Ma, J. Discovery of myricetin as an inhibitor against Streptococcus mutans and an anti-adhesion approach to biofilm formation. Int. J. Med. Microbiol. 2021, 311, 151512. [Google Scholar] [CrossRef]
- Xu, H.; Ziegelin, G.; Schröder, W.; Frank, J.; Ayora, S.; Alonso, J.C.; Lanka, E.; Saenger, W. Flavones inhibit the hexameric replicative helicase RepA. Nucleic Acids Res. 2001, 29, 5058–5066. [Google Scholar] [CrossRef]
- Jayaraman, P.; Sakharkar, M.K.; Lim, C.S.; Tang, T.H.; Sakharkar, K.R. Activity and interactions of antibiotic and phytochemical combinations against Pseudomonas aeruginosa in vitro. Int. J. Biol. Sci. 2010, 6, 556–568. [Google Scholar] [CrossRef]
- Krzyżek, P.; Migdał, P.; Paluch, E.; Karwańska, M.; Wieliczko, A.; Gościniak, G. Myricetin as an Antivirulence Compound Interfering with a Morphological Transformation into Coccoid Forms and Potentiating Activity of Antibiotics against Helicobacter pylori. Int. J. Mol. Sci. 2021, 22, 2695. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, P.; Lv, H.; Deng, X.; Wang, J. A Natural Dietary Flavone Myricetin as an α-Hemolysin Inhibitor for Controlling Staphylococcus aureus Infection. Front. Cell Infect. Microbiol. 2020, 10, 330. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, G.; Si, X.; Zhang, X.; Liu, W.; Li, L.; Wang, J. Inhibition of suilysin activity and inflammation by myricetin attenuates Streptococcus suis virulence. Life Sci. 2019, 223, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Nakane, H.; Fukushima, M.; Chermann, J.C.; Barré-Sinoussi, F. Differential inhibitory effects of various flavonoids on the activities of reverse transcriptase and cellular DNA and RNA polymerases. Eur. J. Biochem. 1990, 190, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.C.; Hsieh, Y.S.; Lin, J.Y. Inhibitory effects of flavonoids on Moloney murine leukemia virus reverse transcriptase activity. J. Nat. Prod. 1992, 55, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Griep, M.A.; Blood, S.; Larson, M.A.; Koepsell, S.A.; Hinrichs, S.H. Myricetin inhibits Escherichia coli DnaB helicase but not primase. Bioorg. Med. Chem. 2007, 15, 7203–7208. [Google Scholar] [CrossRef] [PubMed]
- Nitulescu, G.; Nicorescu, I.M.; Olaru, O.T.; Ungurianu, A.; Mihai, D.P.; Zanfirescu, A.; Nitulescu, G.M.; Margina, D. Molecular docking and screening studies of new natural Sortase a inhibitors. Int. J. Mol. Sci. 2017, 18, 2217. [Google Scholar] [CrossRef]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus Virulence. Microbiol. Spectr. 2019, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhou, H.; Huang, J.; Zhang, R.; Rao, X. Virulence alterations in Staphylococcus aureus upon treatment with the sub-inhibitory concentrations of antibiotics. J. Adv. Res. 2021, 31, 165–175. [Google Scholar] [CrossRef]
- Hodille, E.; Rose, W.; Diep, B.A.; Goutelle, S.; Lina, G.; Dumitrescu, O. The Role of Antibiotics in Modulating Virulence in Staphylococcus aureus. Clin. Microbiol. Rev. 2017, 30, 887–917. [Google Scholar] [CrossRef]
- Kong, C.; Neoh, H.M.; Nathan, S. Targeting Staphylococcus aureus Toxins: A Potential form of Anti-Virulence Therapy. Toxins 2016, 8, 72. [Google Scholar] [CrossRef]
- Sabino, Y.N.V.; Cotter, P.D.; Mantovani, H.C. Anti-virulence compounds against Staphylococcus aureus associated with bovine mastitis: A new therapeutic option? Microbiol. Res. 2023, 271, 127345. [Google Scholar] [CrossRef] [PubMed]
- Pecoraro, C.; Carbone, D.; Parrino, B.; Cascioferro, S.; Diana, P. Recent Developments in the Inhibition of Bacterial Adhesion as Promising Anti-Virulence Strategy. Int. J. Mol. Sci. 2023, 24, 4872. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Le, C.Y.; Ye, Y.J.; Xu, J.; Li, L.; Feng, X.Q.; Chen, N.P.; Zhu, B.Q.; Ding, Z.S.; Qian, C.D. Hinokitiol Selectively Enhances the Antibacterial Activity of Tetracyclines against Staphylococcus aureus. Microbiol. Spectr. 2023, 11, e0320522. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (μg/mL) | ||
|---|---|---|---|
| Myricetin | HQNO | Vancomycin | |
| S. aureus ATCC 43300 | 128 | 8 | 2 |
| S. aureus ATCC 25923 | 128 | 8 | 1 |
| S. aureus RN4220 | 64 | 4 | 1 |
| S. aureus Newman | 64 | 4 | 1 |
| Name | Primer Sequence (5′ to 3′) |
|---|---|
| NdhC-F | CTTGCAGACGAATGTCGGCATCACAATCTGACTGAATCTTGCTTG |
| NdhC-R | CGGCTTAATAGCTCACGCTATGTACAACAATAAAGCCCTTCAGTG |
| YZ-1-F | TAGGTTGAGGCCGTTGA |
| YX-1-R | CTTCAATGCCAAAGGT |
| YZ-2-F | GCGTGGCCAAAAATAT |
| YZ-2-R | GTTCCTCCTTTCAGC |
| hlα-F | ACAATTTTAGAGAGCCCAACTGAT |
| hlα-R | TCCCCAATTTTGATTCACCAT |
| crtm-F | ATCCAGAACCACCCGTTTTT |
| crtm-R | GCGATGAAGGTATTGGCATT |
| sea-F | ACGATCAATTTTTACAGC |
| sea-R | TGCATGTTTTCAGAGTTAATC |
| seb-F | ATTCTATTAAGGACACTAAGTTAGGGGA |
| seb-R | ATCCCGTTTCATAAGGCGAGT |
| gyrb-F | CAAATGATCACAGCATTTGGTACAG |
| gyrb-R | CGGCATCAGTCATAATGACGAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.-L.; Chen, H.-H.; Xu, J.; Huang, M.-Y.; Wang, J.-F.; Shen, H.-J.; Shen, S.-X.; Gao, C.-X.; Qian, C.-D. Myricetin Acts as an Inhibitor of Type II NADH Dehydrogenase from Staphylococcus aureus. Molecules 2024, 29, 2354. https://doi.org/10.3390/molecules29102354
Zhou J-L, Chen H-H, Xu J, Huang M-Y, Wang J-F, Shen H-J, Shen S-X, Gao C-X, Qian C-D. Myricetin Acts as an Inhibitor of Type II NADH Dehydrogenase from Staphylococcus aureus. Molecules. 2024; 29(10):2354. https://doi.org/10.3390/molecules29102354
Chicago/Turabian StyleZhou, Jia-Le, Hao-Han Chen, Jian Xu, Ming-Yu Huang, Jun-Feng Wang, Hao-Jie Shen, Sheng-Xiang Shen, Cheng-Xian Gao, and Chao-Dong Qian. 2024. "Myricetin Acts as an Inhibitor of Type II NADH Dehydrogenase from Staphylococcus aureus" Molecules 29, no. 10: 2354. https://doi.org/10.3390/molecules29102354
APA StyleZhou, J.-L., Chen, H.-H., Xu, J., Huang, M.-Y., Wang, J.-F., Shen, H.-J., Shen, S.-X., Gao, C.-X., & Qian, C.-D. (2024). Myricetin Acts as an Inhibitor of Type II NADH Dehydrogenase from Staphylococcus aureus. Molecules, 29(10), 2354. https://doi.org/10.3390/molecules29102354