
Natural Polyhydroxyalkanoates—An Overview of Bacterial Production Methods


Abstract
1. Introduction
2. Metabolism of PHAs
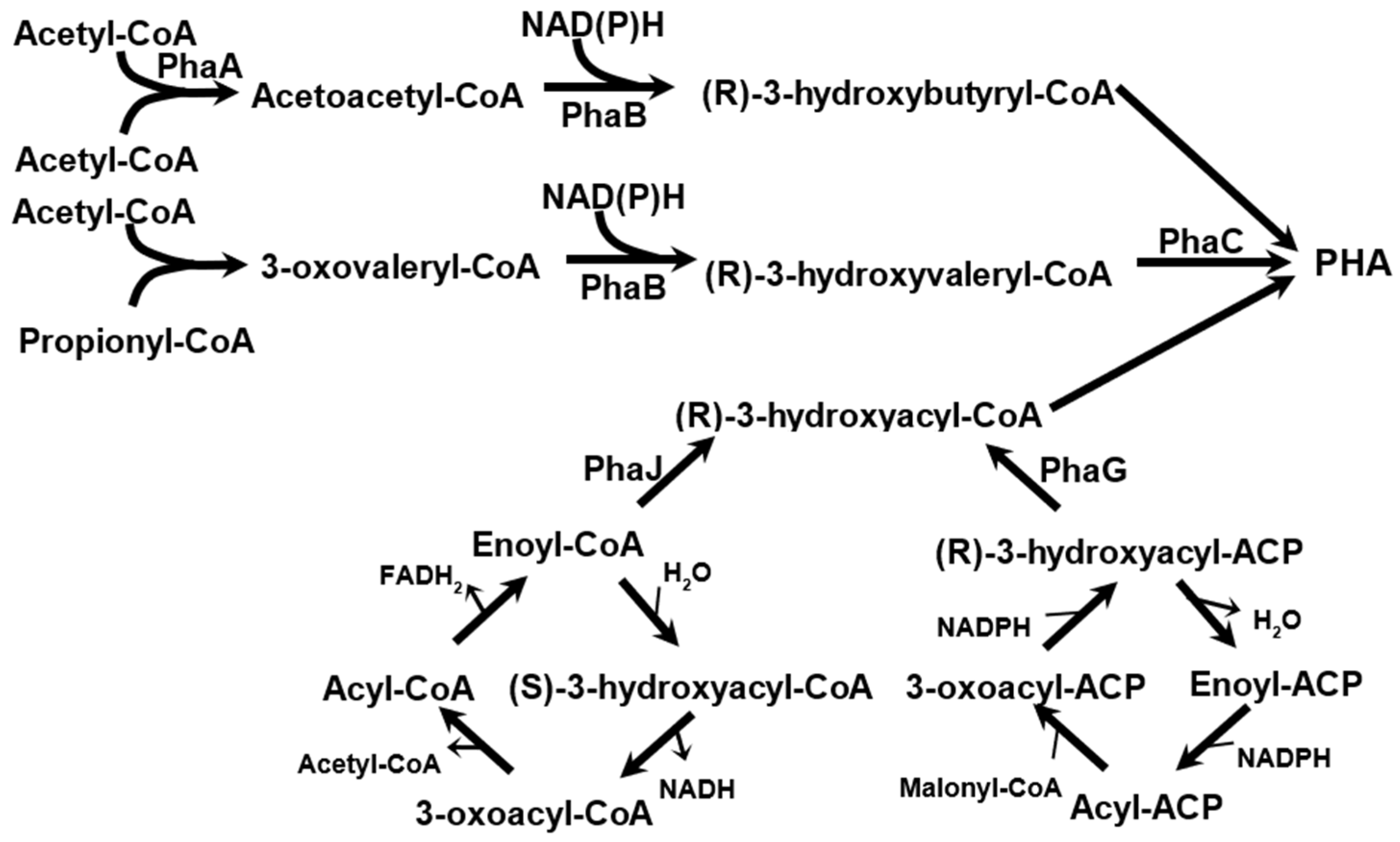
2.1. Biosynthesis
2.2. PHA Granules
2.3. Mobilization of PHAs
2.4. Role of PHAs in Cell Physiology
3. Determination of PHA Content in Cells
4. Extraction Methods
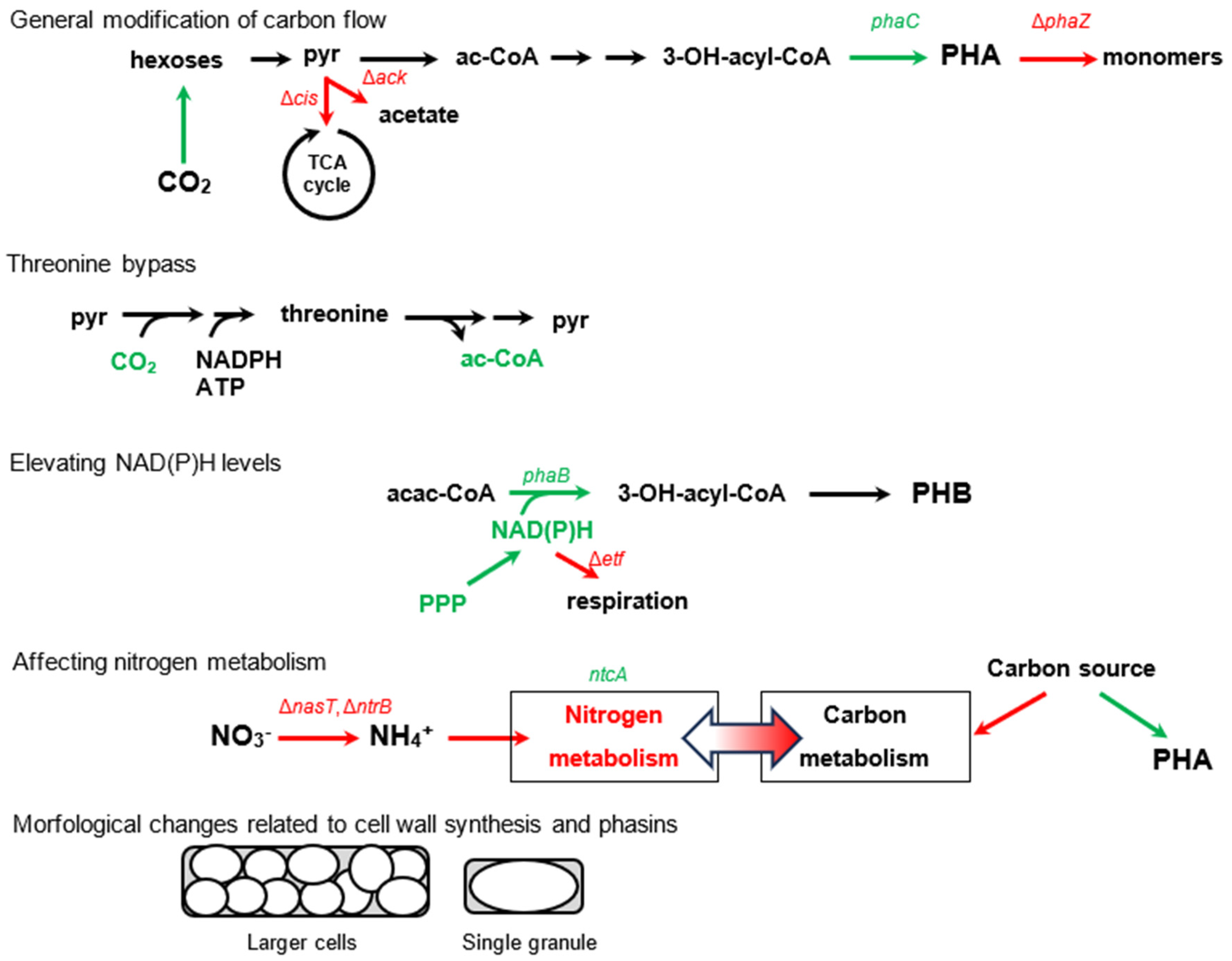
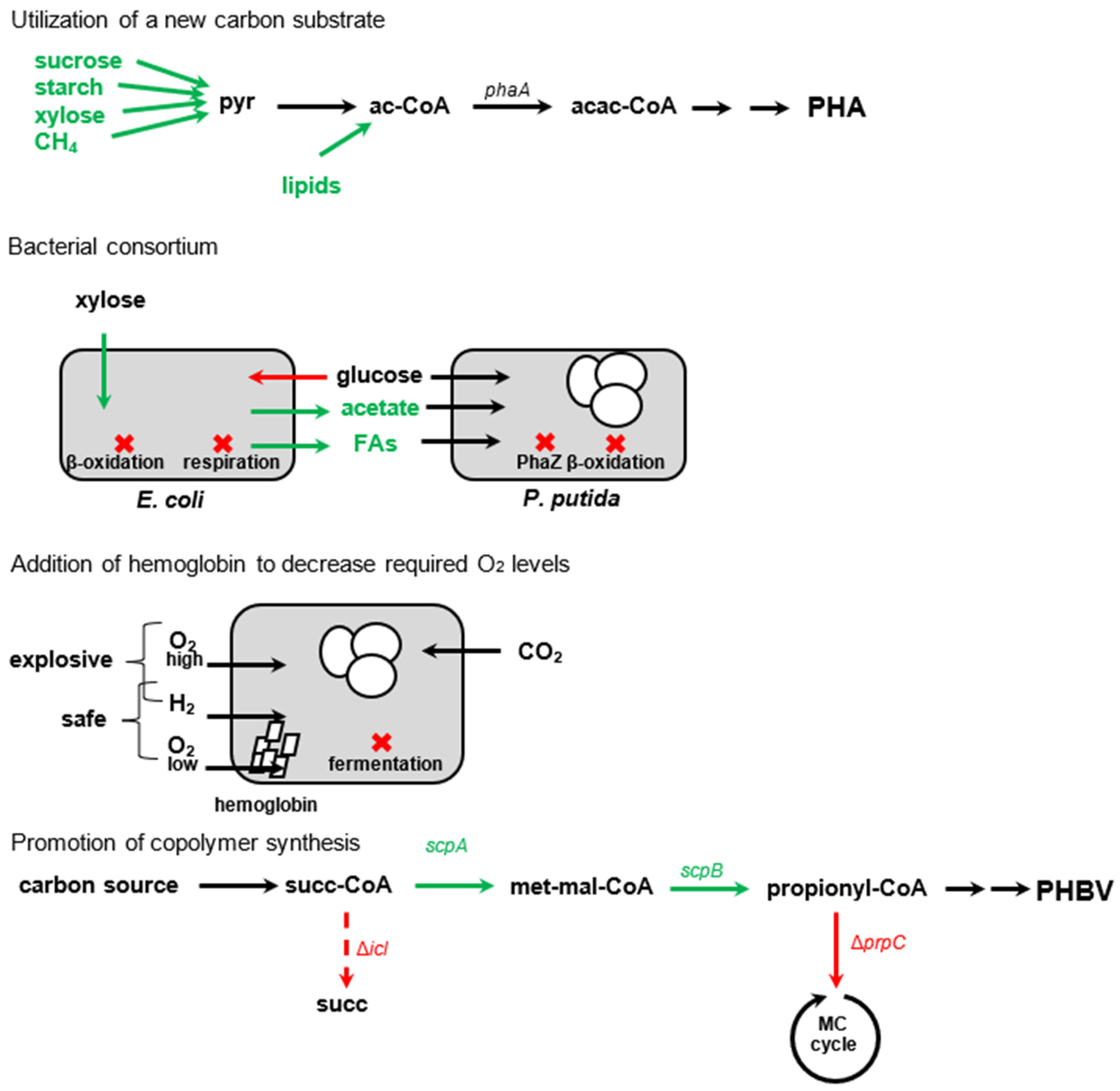
5. Overview of Artificial Modifications to Enhance PHA Production
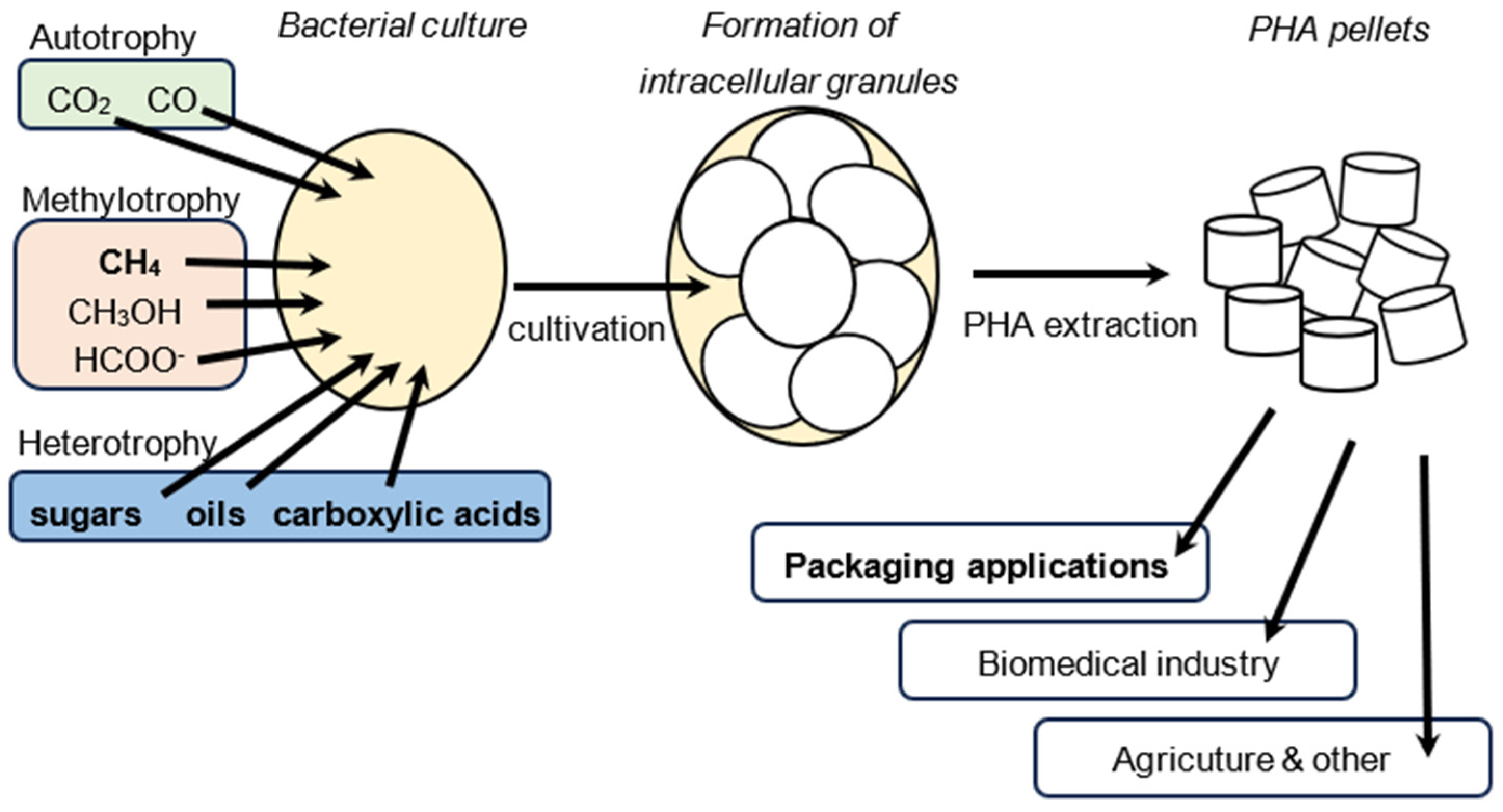
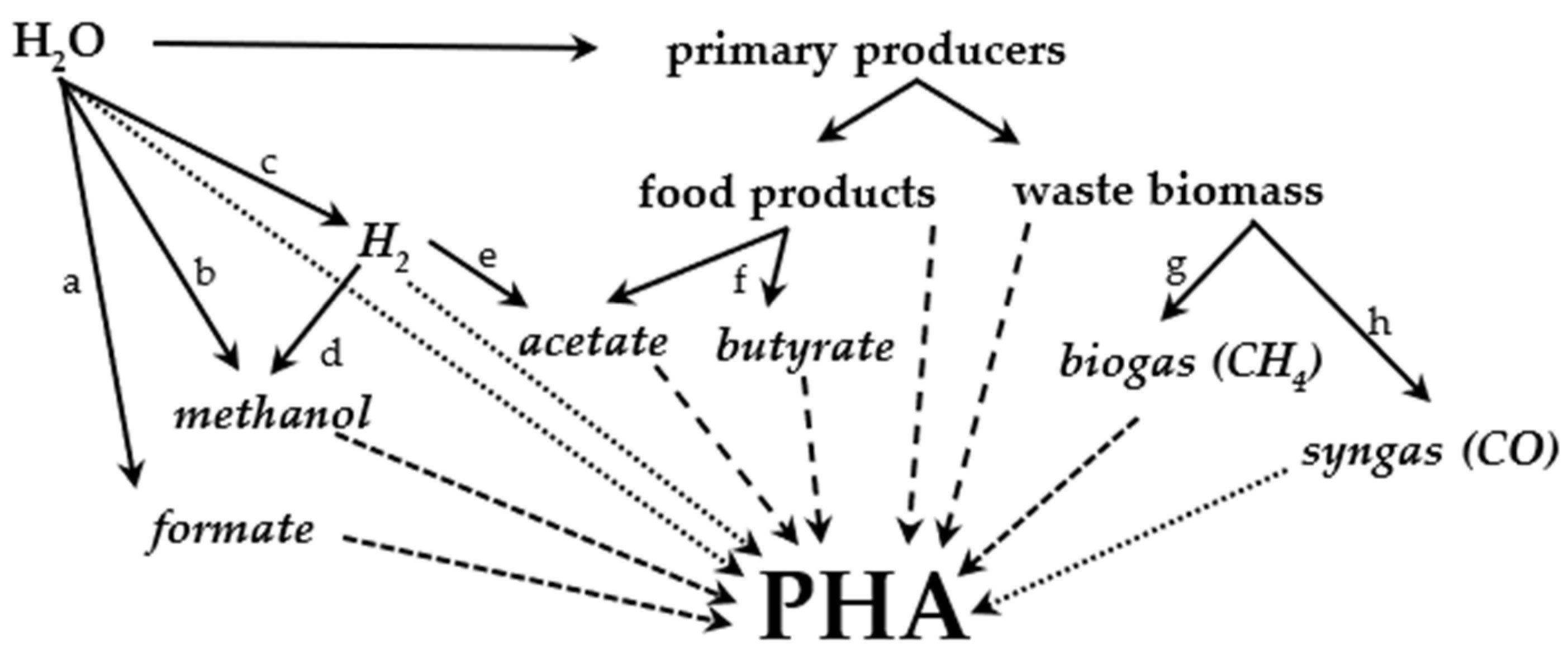
6. Application of PHA-Producing Strains
6.1. Autotrophy
6.1.1. Photoautotrophy (Sunlight)
6.1.2. Hydrogenotrophy (H2)
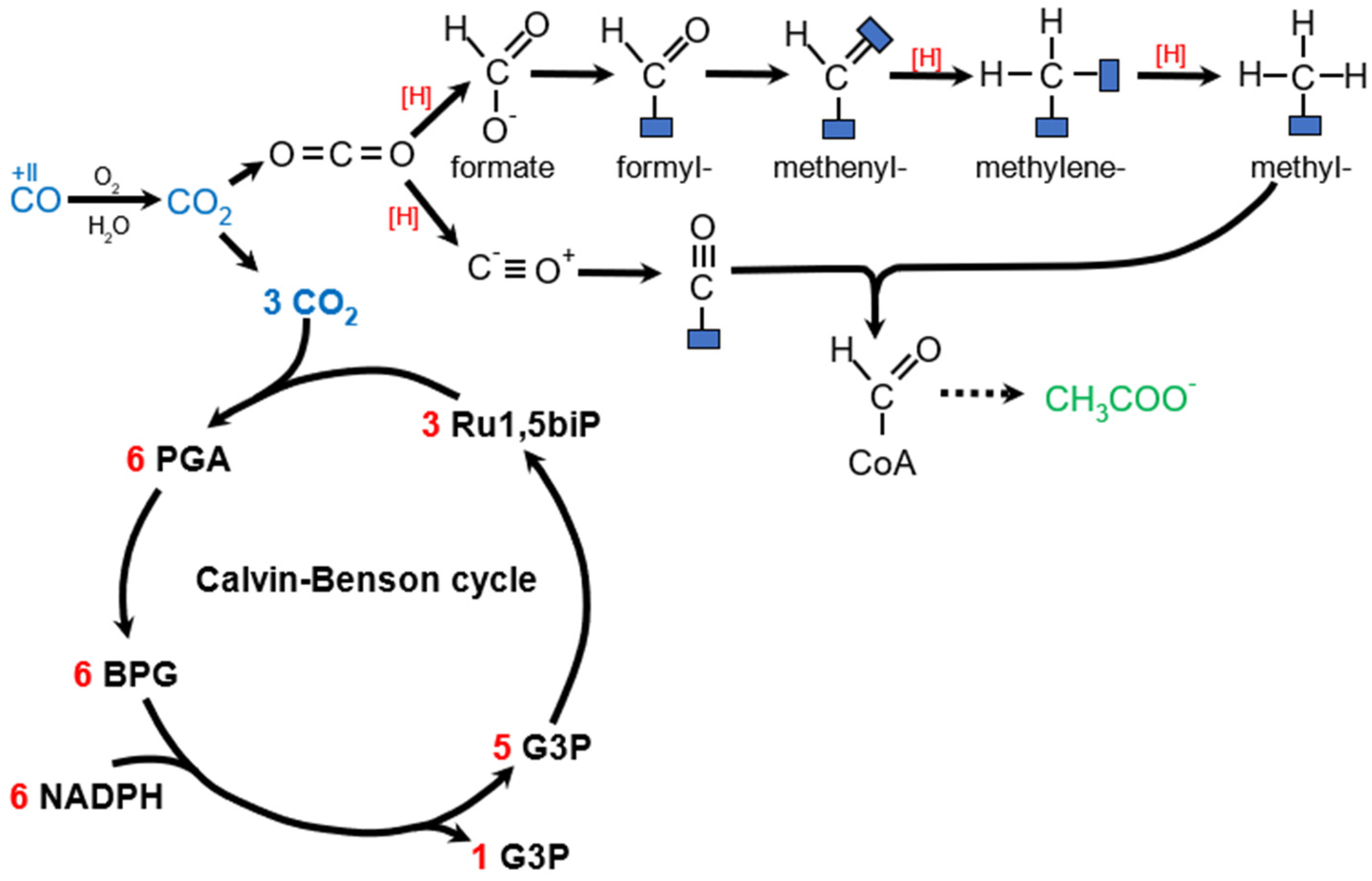
6.1.3. Carboxydotrophy (CO)
6.2. Methylotrophy
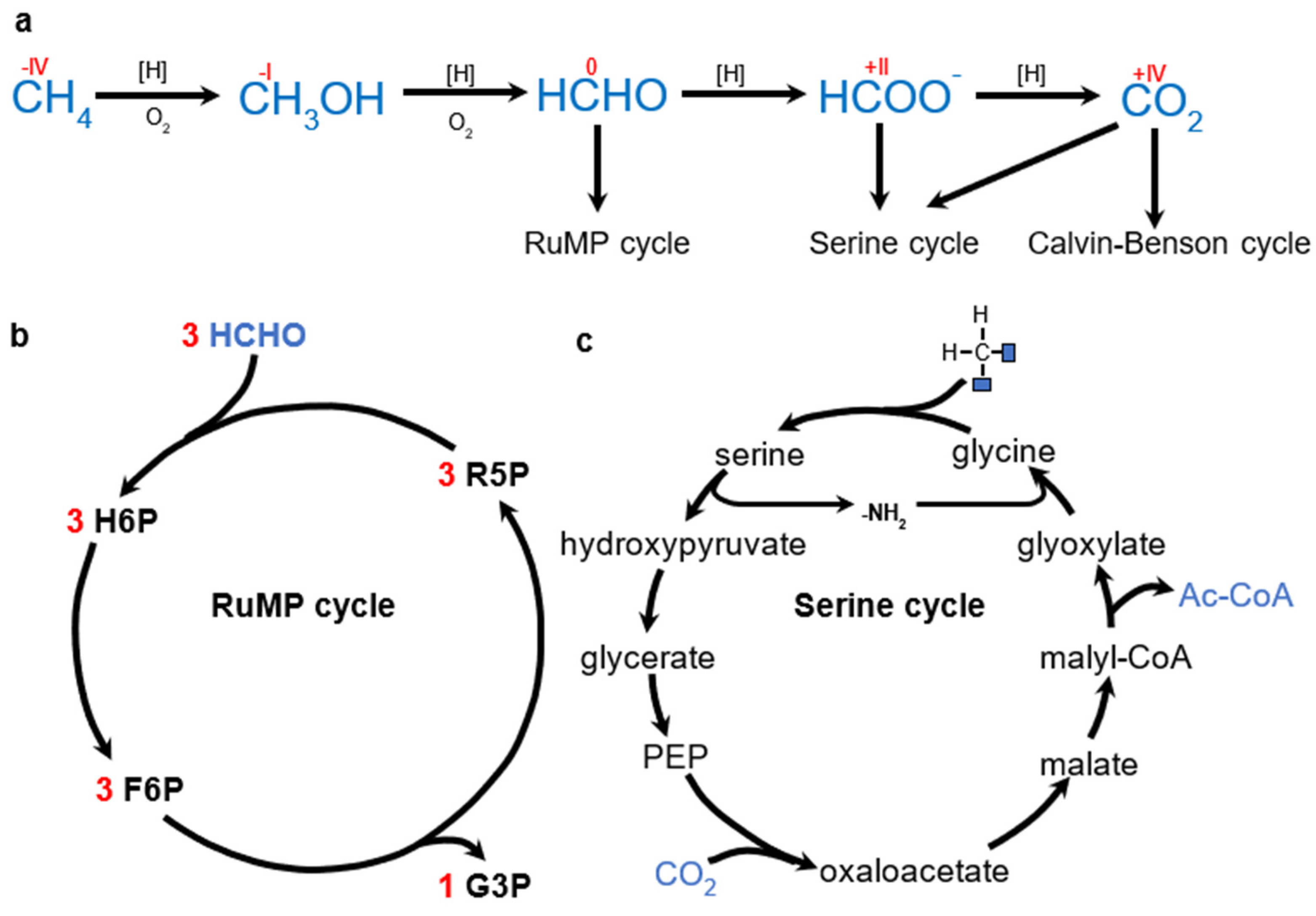
6.2.1. Methane (CH4)
6.2.2. Methanol (CH3OH)
6.2.3. Formate (HCOO−)
6.3. Non-Methylotrophic Heterotrophy
6.3.1. Food Products
6.3.2. Food and Other Industry Waste
6.3.3. Hydrocarbons
6.3.4. Short-Chain Carboxylic Acids
6.3.5. Photo-Organoheterotrophy
7. Discussion and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
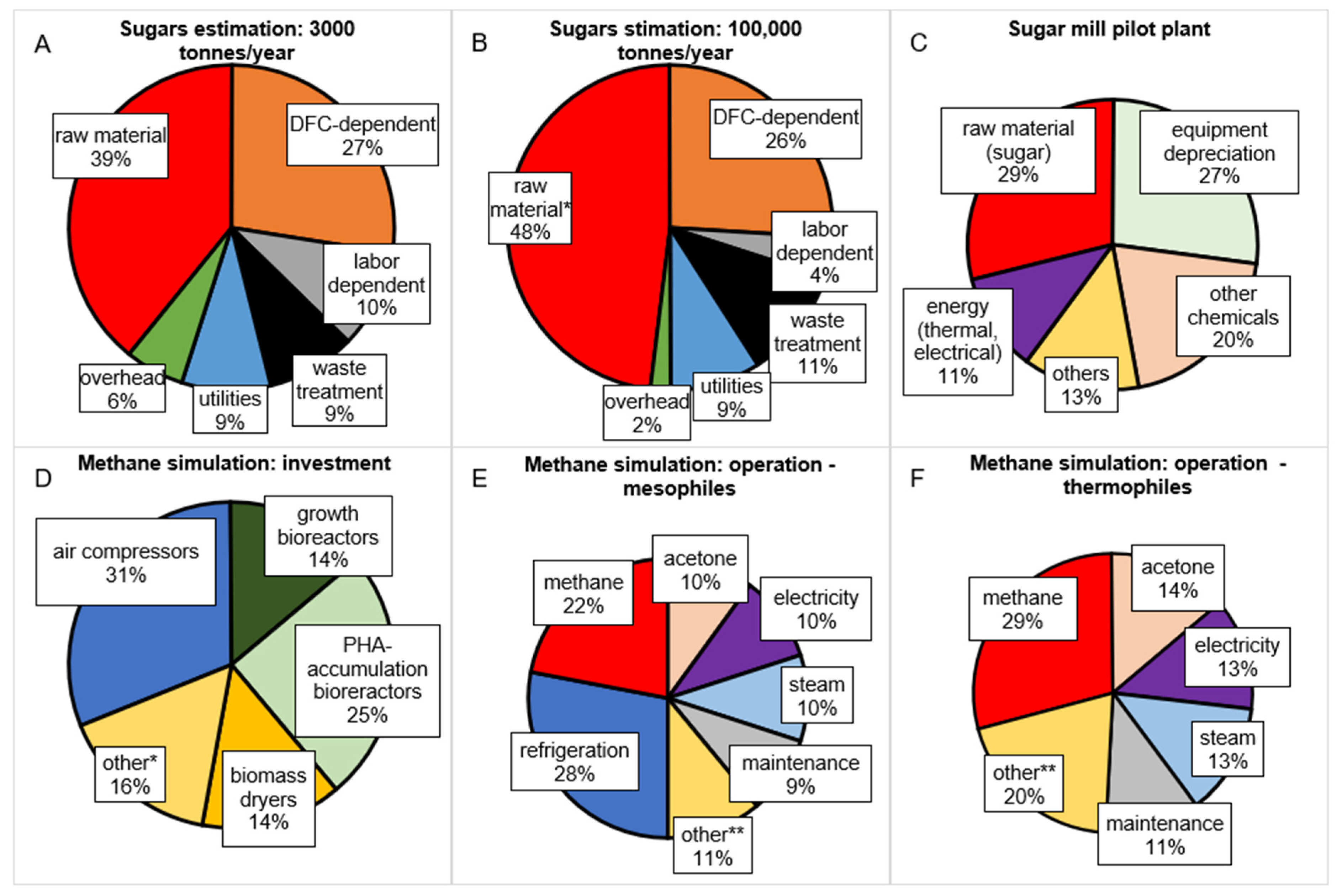

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ΔfH°comb (kJ/mol) | ρ (g/cm3) | M (g/mol) |
|---|---|---|---|
| MeOH (l) | −238.4 | 0.792 | 32.042 |
| HCOOH (l) | −425.09 | 1.22 | 46.025 |
| CO2 (g) | −393.51 | ||
| H2O (l) | −285.83 | ||
| H2 (l, −253 °C) | 0.071 | 2.016 | |
| H2 (g, 70 MPa) | 37 | 2.016 |
References
- Ainali, N.M.; Kalaronis, D.; Evgenidou, E.; Kyzas, G.Z.; Bobori, D.C.; Kaloyianni, M.; Yang, X.; Bikiaris, D.N.; Lambropoulou, D.A. Do poly(lactic acid) microplastics instigate a threat? A perception for their dynamic towards environmental pollution and toxicity. Sci. Total. Environ. 2022, 832, 155014. [Google Scholar] [CrossRef]
- Atarés, L.; Chiralt, A.; González-Martínez, C.; Vargas, M. Production of Polyhydroxyalkanoates for Biodegradable Food Packaging Applications Using Haloferax mediterranei and Agrifood Wastes. Foods 2024, 13, 950. [Google Scholar] [CrossRef]
- Bulantekin, Ö.; Alp, D. Perspective Chapter: Development of Food Packaging Films from Microorganism-Generated Polyhydroxyalkanoates. In Food Processing and Packaging Technologies—Recent Advances; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Bonartsev, A.P.; Bonartseva, G.A.; Reshetov, I.V.; Kirpichnikov, M.P.; Shaitan, K.V. Application of Polyhydroxyalkanoates in Medicine and the Biological Activity of Natural Poly(3-Hydroxybutyrate). Acta Nat. 2019, 11, 4–16. [Google Scholar] [CrossRef]
- Levett, I.; Pratt, S.; Donose, B.C.; Brackin, R.; Pratt, C.; Redding, M.; Laycock, B. Understanding the Mobilization of a Nitrification Inhibitor from Novel Slow Release Pellets, Fabricated through Extrusion Processing with PHBV Biopolymer. J. Agric. Food Chem. 2019, 67, 2449–2458. [Google Scholar] [CrossRef]
- Doi, Y.; Kitamura, S.; Abe, H. Microbial Synthesis and Characterization of Poly(3-Hydroxybutyrate-Co-3-Hydroxyhexanoate). Macromolecules 1995, 28, 4822–4828. [Google Scholar] [CrossRef]
- Bhubalan, K.; Rathi, D.-N.; Abe, H.; Iwata, T.; Sudesh, K. Improved synthesis of P(3HB-co-3HV-co-3HHx) terpolymers by mutant Cupriavidus necator using the PHA synthase gene of Chromobacterium sp. USM2 with high affinity towards 3HV. Polym. Degrad. Stabil. 2010, 95, 1436–1442. [Google Scholar] [CrossRef]
- Bhubalan, K.; Lee, W.-H.; Loo, C.-Y.; Yamamoto, T.; Tsuge, T.; Doi, Y.; Sudesh, K. Controlled biosynthesis and characterization of poly(3-hydroxybutyrate-co-3-hydroxyvalerate-co-3-hydroxyhexanoate) from mixtures of palm kernel oil and 3HV-precursors. Polym. Degrad. Stabil. 2008, 93, 17–23. [Google Scholar] [CrossRef]
- Yamane, T.; Chen, X.F.; Ueda, S. Growth-associated production of poly(3-hydroxyvalerate) from n-pentanol by a methylotrophic bacterium, Paracoccus denitrificans. Appl. Environ. Microbiol. 1996, 62, 380–384. [Google Scholar] [CrossRef]
- Saito, Y.; Doi, Y. Microbial Synthesis and Properties of Poly(3-Hydroxybutyrate-Co-4-Hydroxybutyrate) in Comamonas acidovorans. Int. J. Biol. Macromol. 1994, 16, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Shi, Z.-Y.; Meng, D.-C.; Wu, Q.; Chen, J.-C.; Chen, G.-Q. Production of 3-hydroxypropionate homopolymer and poly(3-hydroxypropionate-co-4-hydroxybutyrate) copolymer by recombinant Escherichia coli. Metab. Eng. 2011, 13, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Wang, Z.H.; Luo, H.N.; Xu, M.; Ren, X.Y.; Zheng, G.X.; Wu, B.J.; Zhang, X.H.; Lu, X.Y.; Chen, F.; et al. Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate)-based scaffolds for tissue engineering. Braz. J. Med. Biol. Res. 2014, 47, 533–539. [Google Scholar] [CrossRef]
- Choi, J.I.; Lee, S.Y. Process analysis and economic evaluation for poly(3-hydroxybutyrate) production by fermentation. Bioprocess. Eng. 1997, 17, 335–342. [Google Scholar] [CrossRef]
- Nonato, R.V.; Mantelatto, P.E.; Rossell, C.E.V. Integrated production of biodegradable plastic, sugar and ethanol. Appl. Microbiol. Biotechnol. 2001, 57, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Levett, I.; Birkett, G.; Davies, N.; Bell, A.; Langford, A.; Laycock, B.; Lant, P.; Pratt, S. Techno-economic assessment of poly-3-hydroxybutyrate (PHB) production from methane—The case for thermophilic bioprocessing. J. Environ. Chem. Eng. 2016, 4, 3724–3733. [Google Scholar] [CrossRef]
- Jaeger, K.-E.; Steinbüchel, A.; Jendrossek, D. Substrate Specificities of Bacterial Polyhydroxyalkanoate Depolymerases and Lipases: Bacterial Lipases Hydrolyze Poly(ω-Hydroxyalkanoates). Appl. Environ. Microbiol. 1995, 8, 3113–3118. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.; Salvador, A.; Alves, M.M.; Vicente, A.A. Factors affecting polyhydroxyalkanoates biodegradation in soil. Polym. Degrad. Stab. 2020, 182, 109408. [Google Scholar] [CrossRef]
- Kim, J.; Gupta, N.S.; Bezek, L.B.; Linn, J.; Bejagam, K.K.; Banerjee, S.; Dumont, J.H.; Nam, S.Y.; Kang, H.W.; Park, C.H.; et al. Biodegradation Studies of Polyhydroxybutyrate and Polyhydroxybutyrate-co-Polyhydroxyvalerate Films in Soil. Int. J. Mol. Sci. 2023, 24, 7638. [Google Scholar] [CrossRef]
- Davis, R.; Chandrashekar, A.; Shamala, T.R. Role of (R)-specific enoyl coenzyme A hydratases of Pseudomonas sp. in the production of polyhydroxyalkanoates. Antonie Van. Leeuwenhoek 2008, 93, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Rehm, B.H.A.; Krüger, N.; Steinbüchel, A. A new metabolic link between fatty acid de novo synthesis and polyhydroxyalkanoic acid synthesis: The phaG gene from Pseudomonas putida KT2440 encodes a 3-hydroxyacyl-acyl carrier protein coenzyme A transferase. J. Biol. Chem. 1998, 273, 24044–24051. [Google Scholar] [CrossRef]
- Maehara, A.; Ueda, S.; Nakano, H.; Yamane, T. Analyses of a polyhydroxyalkanoic acid granule-associated 16-kilodalton protein and its putative regulator in the pha locus of Paracoccus denitrificans. J. Bacteriol. 1999, 181, 2914–2921. [Google Scholar] [CrossRef]
- Pfeiffer, D.; Wahl, A.; Jendrossek, D. Identification of a multifunctional protein, PhaM, that determines number, surface to volume ratio, subcellular localization and distribution to daughter cells of poly(3-hydroxybutyrate), PHB, granules in Ralstonia eutropha H16. Mol. Microbiol. 2011, 82, 936–951. [Google Scholar] [CrossRef] [PubMed]
- Brigham, C.J.; Sinskey, A.J. Polyhydroxybutyrate Production Enzymes: A Survey and Biological perspective. J. Sib. Fed. Univ. Biol. 2012, 3, 220–242. [Google Scholar]
- Lu, J.; Takahashi, A.; Ueda, S. 3-Hydroxybutyrate Oligomer Hydrolase and 3-Hydroxybutyrate Dehydrogenase Participate in Intracellular Polyhydroxybutyrate and Polyhydroxyvalerate Degradation in Paracoccus denitrificans. Appl. Environ. Microbiol. 2014, 80, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Koskimäki, J.J.; Kajula, M.; Hokkanen, J.; Ihantola, E.-L.; Kim, J.H.; Hautajärvi, H.; Hankala, E.; Suokas, M.; Pohjanen, J.; Podolich, O.; et al. Methyl-esterified 3-hydroxybutyrate oligomers protect bacteria from hydroxyl radicals. Nat. Chem. Biol. 2016, 12, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Seebach, D.; Brunner, A.; Bürger, H.M.; Schneider, J.; Reusch, R.N. Isolation and 1H-NMR spectroscopic identification of poly(3-hydroxybutanoate) from prokaryotic and eukaryotic organisms—Determination of the absolute-configuration (R) of the monomeric unit 3-hydroxybutanoic acid from Escherichia coli and spinach. Eur. J. Biochem. 1994, 224, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Reusch, R.N.; Sparrow, A.W.; Gardiner, J. Transport of poly-β-hydroxybutyrate in human plasma. Biochim. Biophys. Acta 1992, 1123, 33–40. [Google Scholar] [CrossRef]
- Reusch, R.M.; Sadoff, H.L. Putative structure and functions of a poly-β-hydroxybutyrate/calcium polyphosphate channel in bacterial plasma membranes. Proc. Natl. Acad. Sci. USA 1988, 85, 4176–4180. [Google Scholar] [CrossRef]
- Elustondo, P.A.; Nichols, M.; Negoda, A.; Thirumaran, A.; Zakharian, E.; Robertson, G.S.; Pavlov, E.V. Mitochondrial permeability transition pore induction is linked to formation of the complex of ATPase C-Subunit, polyhydroxybutyrate and inorganic polyphosphate. Cell Death Discov. 2016, 2, 16070. [Google Scholar] [CrossRef] [PubMed]
- Xian, M.; Fuerst, M.M.; Shabalin, Y.; Rosetta, R.N. Sorting Signal of Escherichia coli OmpA is Modified by Oligo-(R)-3-Hydroxybutyrate. Biochim. Biophys. Acta Biomemb. 2007, 1768, 2660–2666. [Google Scholar] [CrossRef] [PubMed]
- Reusch, R.N.; Shabalin, O.; Crumbaugh, A.; Wagner, R.; Schröder, O.; Wurm, R. Posttranslational modification of E. coli histone-like protein H-NS and bovine histones by short-chain poly-(R)-3-hydroxybutyrate (cPHB). FEBS Lett. 2002, 527, 319–322. [Google Scholar] [CrossRef]
- Dai, D.; Reusch, R.N. Poly-3-hydroxybutyrate Synthase from the Periplasm of Escherichia coli. Biochem. Biophys. Res. Commun. 2008, 374, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Reusch, R.N. Physiological Importance of Poly-(R)-3-Hydroxybutyrates. Chem. Biodivers. 2009, 9, 2343–2366. [Google Scholar] [CrossRef] [PubMed]
- Zuriani, R.; Vigneswari, S.; Azizan, M.N.M.; Majid, M.I.A.; Amirul, A.A. A high throughput Nile red fluorescence method for rapid quantification of intracellular bacterial polyhydroxyalkanoates. Biotechnol. Bioprocess Eng. 2013, 18, 472–478. [Google Scholar] [CrossRef]
- Lakshman, K.; Shamala, T.R. Extraction of polyhydroxyalkanoate from Sinorhizobium meliloti cells using Microbispora sp. culture and its enzymes. Enzyme Microb. Technol. 2006, 39, 1471–1475. [Google Scholar] [CrossRef]
- Randriamahefa, S.; Renard, E.; Guerin, P.; Langlois, V. Fourier transform infrared spectroscopy for screening and quantifying production of PHAs by Pseudomonas grown on sodium octanoate. Biomacromolecules 2003, 4, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Isak, I.; Patel, M.; Riddell, M.; West, M.; Bowers, T.; Wijeyekoon, S.; Lloyd, J. Quantification of polyhydroxyalkanoates in mixed and pure cultures biomass by Fourier transform infrared spectroscopy: Comparison of different approaches. Lett. Appl. Microbiol. 2016, 63, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Censi, V.; Saiano, F.; Bongiorno, D.; Indelicato, S.; Napoli, A.; Piazzese, D. Bioplastics: A new analytical challenge. Front. Chem. 2022, 10, 987669. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, P.; Vasheghani-Farahani, E.; Yamini, Y. Supercritical fluid disruption of Ralstonia eutropha for poly(beta-hydroxybutyrate) recovery. Biotechnol. Prog. 2003, 19, 1519–1523. [Google Scholar] [CrossRef] [PubMed]
- Aramvash, A.; Gholami-Banadkuki, N.; Moazzeni-Zavareh, F.; Hajizadeh-Turchi, S. An Environmentally Friendly and Efficient Method for Extraction of PHB Biopolymer with Non-Halogenated Solvents. J. Microbiol. Biotechnol. 2015, 25, 1936–1943. [Google Scholar] [CrossRef]
- Riedel, S.L.; Brigham, C.J.; Budde, C.F.; Bader, J.; Rha, C.; Stah, U.; Sinskey, A.J. Recovery of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from Ralstonia eutropha cultures with non-halogenated solvents. Biotechnol. Bioeng. 2013, 110, 461–470. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Brigham, C.; Willis, L.; Rha, C.; Sinskey, A. Improved detergent-based recovery of polyhydroxyalkanoates (PHAs). Biotechnol. Lett. 2011, 33, 937–942. [Google Scholar] [CrossRef]
- Xiong, B.; Fang, Q.; Wei, T.; Wang, Z.; Shen, R.; Cheng, M.; Zhou, W. Chemical digestion method to promote activated sludge cell wall breaking and optimize the polyhydroxyalkanoate (PHA) extraction process. Int. J. Biol. Macromol. 2023, 240, 124369. [Google Scholar] [CrossRef]
- Burniol-Figols, A.; Skiadas, I.; Daugaard, A.E.; Gavala, H.N. Polyhydroxyalkanoate (PHA) purification through dilute aqueous ammonia digestion at elevated temperatures. J. Chem. Technol. Biotechnol. 2020, 95, 1519–1532. [Google Scholar] [CrossRef]
- Page, W.; Cornish, A. Growth of Azotobacter vinelandii Uwd in Fish Peptone Medium and Simplified Extraction of Poly-Beta-Hydroxybutyrate. Appl. Environ. Microbiol. 1993, 59, 4236–4244. [Google Scholar] [CrossRef]
- Meneses, L.; Esmail, A.; Matos, M.; Sevrin, C.; Grandfils, C.; Barreiros, S.; Reis, M.A.M.; Freitas, F.; Paiva, A. Subcritical Water as a Pre-Treatment of Mixed Microbial Biomass for the Extraction of Polyhydroxyalkanoates. Bioengineering 2022, 9, 302. [Google Scholar] [CrossRef]
- Rahman, A.; Linton, E.; Hatch, A.D.; Sims, R.C.; Miller, C.D. Secretion of polyhydroxybutyrate in Escherichia coli using a synthetic biological engineering approach. J. Biol. Eng. 2013, 7, 24. [Google Scholar] [CrossRef]
- Sabirova, J.S.; Ferrer, M.; Luensdorf, H.; Wray, V.; Kalscheuer, R.; Steinbuechel, A.; Timmis, K.N.; Golyshin, P.N. Mutation in a “tesB-like” hydroxyacyl-coenzyme A-specific thioesterase gene causes hyperproduction of extracellular polyhydroxyalkanoates by Alcanivorax borkumensis SK2. J. Bacteriol. 2006, 188, 8452–8459. [Google Scholar] [CrossRef]
- Wang, J.; Liu, S.; Huang, J.; Cui, R.; Xu, Y.; Song, Z. Genetic engineering strategies for sustainable polyhydroxyalkanoate (PHA) production from carbon-rich wastes. Environ. Technol. Innov. 2023, 30, 103069. [Google Scholar] [CrossRef]
- Barati, F.; Asgarani, E.; Gharavi, S.; Soudi, M.R. Considerable increase in Poly(3-hydroxybutyrate) production via phbC gene overexpression in Ralstonia eutropha PTCC 1615. BioImpacts 2021, 11, 53–57. [Google Scholar] [CrossRef]
- Kim, S.; Jang, Y.J.; Gong, G.; Lee, S.-M.; Um, Y.; Kim, K.H.; Ko, J.K. Engineering Cupriavidus necator H16 for enhanced lithoautotrophic poly(3-hydroxybutyrate) production from CO2. Microb. Cell. Fact. 2022, 21, 231. [Google Scholar] [CrossRef]
- Lin, L.; Chen, J.; Mitra, R.; Gao, Q.; Cheng, F.; Xu, T.; Zuo, Z.; Xiang, H.; Han, J. Optimising PHBV biopolymer production in haloarchaea via CRISPRi-mediated redirection of carbon flux. Commun. Biol. 2021, 4, 1007. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, G.B.; Kim, H.U.; Park, S.J.; Choi, J. Enhanced production of poly-3-hydroxybutyrate (PHB) by expression of response regulator DR1558 in recombinant Escherichia coli. Int. J. Biol. Macromol. 2019, 131, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zhang, Y.; Yuan, Q.; Liu, Q.; Li, Y.; Wang, Z.; Ma, H.; Chen, T.; Zhao, X. Metabolic engineering of Escherichia coli for poly(3-hydroxybutyrate) production via threonine bypass. Microb. Cell Factories 2015, 14, 185. [Google Scholar] [CrossRef]
- Tadi, S.R.R.; Ravindran, S.D.; Balakrishnan, R.; Sivaprakasam, S. Recombinant production of poly-(3-hydroxybutyrate) by Bacillus megaterium utilizing millet bran and rapeseed meal hydrolysates. Bioresour. Technol. 2021, 326, 124800. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Qiao, G.-Q.; Shuai, B.-W.; Olavarria, K.; Yin, J.; Xiang, R.-J.; Song, K.-N.; Shen, Y.-H.; Guo, Y.; Chen, G.-Q. Engineering NADH/NAD(+) ratio in Halomonas bluephagenesis for enhanced production of polyhydroxyalkanoates (PHA). Metab. Eng. 2018, 49, 275–286. [Google Scholar] [CrossRef]
- Olaya-Abril, A.; Luque-Almagro, V.M.; Manso, I.; Gates, A.J.; Moreno-Vivian, C.; Richardson, D.J.; Roldán, M.D. Poly(3-hydroxybutyrate) hyperproduction by a global nitrogen regulator NtrB mutant strain of Paracoccus denitrificans PD1222. FEMS Microbiol. Lett. 2018, 365, fnx251. [Google Scholar] [CrossRef]
- Hobmeier, K.; Loewe, H.; Liefeldt, S.; Kremling, A.; Pflueger-Grau, K. A Nitrate-Blind, P. putida Strain Boosts PHA Production in a Synthetic Mixed Culture. Front. Bioeng. Biotechnol. 2020, 8, 486. [Google Scholar] [CrossRef]
- Arisaka, S.; Terahara, N.; Oikawa, A.; Osanai, T. Increased polyhydroxybutyrate levels by ntcA overexpression in Synechocystis sp. PCC 6803. Algal. Res. 2019, 41, 101565. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, Y.; Ji, M.; Zhang, X.; Wang, H.; Chen, Y.; Wu, Q.; Chen, G.-Q. Hyperproduction of PHA copolymers containing high fractions of 4-hydroxybutyrate (4HB) by outer membrane-defected Halomonas bluephagenesis grown in bioreactors. Microb. Biotechnol. 2022, 15, 1586–1597. [Google Scholar] [CrossRef]
- Shen, R.; Ning, Z.-Y.; Lan, Y.-X.; Chen, J.-C.; Chen, G.-Q. Manipulation of polyhydroxyalkanoate granular sizes in Halomonas bluephagenesis. Metab. Eng. 2019, 54, 117–126. [Google Scholar] [CrossRef]
- Fukui, T.; Mukoyama, M.; Orita, I.; Nakamura, S. Enhancement of glycerol utilization ability of Ralstonia eutropha H16 for production of polyhydroxyalkanoates. Appl. Microbiol. Biotechnol. 2014, 98, 7559–7568. [Google Scholar] [CrossRef] [PubMed]
- Arikawa, H.; Matsumoto, K.; Fujiki, T. Polyhydroxyalkanoate production from sucrose by Cupriavidus necator strains harboring csc genes from Escherichia coli. W. Appl. Microbiol. Biotechnol. 2017, 101, 7497–7507. [Google Scholar] [CrossRef] [PubMed]
- Huo, G.; Zhu, Y.; Liu, Q.; Tao, R.; Diao, N.; Wang, Z.; Chen, T. Metabolic engineering of an E. coli ndh knockout strain for PHB production from mixed glucose-xylose feedstock. J. Chem. Technol. Biotechnol. 2017, 92, 2739–2745. [Google Scholar] [CrossRef]
- Brojanigo, S.; Gronchi, N.; Cazzorla, T.; Wong, T.S.; Basaglia, M.; Favaro, L.; Casella, S. Engineering Cupriavidus necator DSM 545 for the one-step conversion of starchy waste into polyhydroxyalkanoates. Bioresour. Technol. 2022, 347, 126383. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.E.; Brojanigo, S.; Basaglia, M.; Favaro, L.; Casella, S. Efficient production of polyhydroxybutyrate from slaughterhouse waste using a recombinant strain of Cupriavidus necator DSM 545. Sci. Total Environ. 2021, 794, 148754. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Li, J.; Huang, Z.; Han, X.; Bao, J. Engineering Corynebacterium glutamicum for synthesis of poly(3-hydroxybutyrate) from lignocellulose biomass. Biotechnol. Bioeng. 2022, 119, 1598–1613. [Google Scholar] [CrossRef] [PubMed]
- Chau, T.H.T.; Nguyen, A.D.; Lee, E.Y. Engineering type I methanotrophic bacteria as novel platform for sustainable production of 3-hydroxybutyrate and biodegradable polyhydroxybutyrate from methane and xylose. Bioresour. Technol. 2022, 363, 127898. [Google Scholar] [CrossRef]
- Qin, R.; Zhu, Y.; Ai, M.; Jia, X. Reconstruction and optimization of a Pseudomonas putida-Escherichia coli microbial consortium for mcl-PHA production from lignocellulosic biomass. Front. Bioeng. Biotechnol. 2022, 10, 1023325. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Weng, C.; Peng, X.; Han, Y. Metabolic engineering of Cupriavidus necator H16 for improved chemoautotrophic growth and PHB production under oxygen-limiting conditions. Metab. Eng. 2020, 61, 11–23. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, X.-Y.; Du, H.-T.; Zhang, X.; Ma, Y.-M.; Chen, J.-C.; Ye, J.-W.; Jiang, X.-R.; Chen, G.-Q. Chromosome engineering of the TCA cycle in Halomonas bluephagenesis for production of copolymers of 3-hydroxybutyrate and 3-hydroxyvalerate (PHBV). Metab. Eng. 2019, 54, 69–82. [Google Scholar] [CrossRef]
- Schönheit, P.; Buckel, W.; Martin, W.F. On the Origin of Heterotrophy. Trends. Microbiol. 2016, 24, 12–25. [Google Scholar] [CrossRef] [PubMed]
- De Marco, P. Methylotrophy versus heterotrophy: A misconception. Microbiology 2004, 150, 1606–1607. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Milo, R. The global mass and average rate of rubisco. Proc. Natl. Acad. Sci. USA 2019, 116, 4738–4743. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Oh, M.-K. Strategies for Biosynthesis of C1 Gas-derived Polyhydroxyalkanoates: A review. Bioresour. Technol. 2022, 344, 126307. [Google Scholar] [CrossRef] [PubMed]
- Kamravamanesh, D.; Kovacs, T.; Pflugl, S.; Druzhinina, I.; Kroll, P.; Lackner, M.; Herwig, C. Increased poly-beta-hydroxybutyrate production from carbon dioxide in randomly mutated cells of cyanobacterial strain Synechocystis sp. PCC 6714: Mutant generation and characterization. Bioresour. Technol. 2018, 266, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Tharasirivat, V.; Jantaro, S. Increased Biomass and Polyhydroxybutyrate Production by Synechocystis sp. PCC 6803 Overexpressing RuBisCO Genes. Int. J. Mol. Sci. 2023, 24, 6415. [Google Scholar] [CrossRef] [PubMed]
- Taepucharoen, K.; Tarawat, S.; Puangcharoen, M.; Incharoensakdi, A.; Monshupanee, T. Production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) under photoautotrophy and heterotrophy by non-heterocystous N2-fixing cyanobacterium. Bioresour. Technol. 2017, 239, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Tarawat, S.; Incharoensakdi, A.; Monshupanee, T. Cyanobacterial production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from carbon dioxide or a single organic substrate: Improved polymer elongation with an extremely high 3-hydroxyvalerate mole proportion. J. Appl. Phycol. 2020, 32, 1095–1102. [Google Scholar] [CrossRef]
- Volova, T.G.; Kiselev, E.G.; Shishatskaya, E.I.; Zhila, N.O.; Boyandin, A.N.; Syrvacheva, D.A.; Vinogradova, O.N.; Kalacheva, G.S.; Vasiliev, A.D.; Peterson, I.V. Cell growth and accumulation of polyhydroxyalkanoates from CO2 and H2 of a hydrogen-oxidizing bacterium, Cupriavidus eutrophus B-10646. Bioresour. Technol. 2013, 146, 215–222. [Google Scholar] [CrossRef]
- Nangle, S.N.; Ziesack, M.; Buckley, S.; Trivedi, D.; Loh, D.M.; Nocera, D.G.; Silver, P.A. Valorization of CO2 through lithoautotrophic production of sustainable chemicals in Cupriavidus necator. Metab. Eng. 2020, 62, 207–220. [Google Scholar] [CrossRef]
- Buan, N.R. Methanogens: Pushing the boundaries of biology. Emerg. Top. Life Sci. 2018, 2, 629–646. [Google Scholar] [CrossRef]
- Daniel, S.; Hsu, T.; Dean, S.; Drake, H. Characterization of the H2-Dependent and CO-Dependent Chemolithotrophic Potentials of the Acetogens Clostridium thermoaceticum and Acetogenium kivui. J. Bacteriol. 1990, 172, 4464–4471. [Google Scholar] [CrossRef]
- Petushkova, E.P.; Tsygankov, A.A. Acetate metabolism in the purple non-sulfur bacterium Rhodobacter capsulatus. Biochemistry 2017, 82, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Brady, A.L.; Sharp, C.E.; Grasby, S.E.; Dunfield, P.F. Anaerobic carboxydotrophic bacteria in geothermal springs identified using stable isotope probing. Front. Microbiol. 2015, 6, 897. [Google Scholar] [CrossRef]
- Volbeda, A.; Fontecilla-Camps, J.C. Crystallographic evidence for a CO/CO2 tunnel gating mechanism in the bifunctional carbon monoxide dehydrogenase/acetyl coenzyme A synthase from Moorella thermoacetica. J. Biol. Inorg. Chem. 2004, 9, 525–532. [Google Scholar] [CrossRef]
- Volova, T.; Zhila, N.; Shishatskaya, E. Synthesis of poly(3-hydroxybutyrate) by the autotrophic CO-oxidizing bacterium Seliberia carboxydohydrogena Z-1062. J. Ind Microbiol. Biotechnol. 2015, 42, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Do, Y.S.; Smeenk, J.; Broer, K.M.; Kisting, C.J.; Brown, R.; Heindel, T.J.; Bobik, T.A.; DiSpirito, A.A. Growth of Rhodospirillum rubrum on synthesis gas: Conversion of CO to H2 and poly-beta-hydroxyalkanoate. Biotechnol. Bioeng. 2007, 97, 279–286. [Google Scholar] [CrossRef]
- Lemgruber, R.S.P.; Valgepea, K.; Tappel, R.; Behrendorff, J.B.; Palfreyman, R.W.; Plan, M.; Hodson, M.P.; Simpson, S.D.; Nielsen, L.K.; Kopke, M.; et al. Systems-level engineering and characterisation of Clostridium autoethanogenum through heterologous production of poly-3-hydroxybutyrate (PHB). Metab. Eng. 2019, 53, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Valgepea, K.; Lemgruber, R.S.P.; Meaghan, K.; Palfreyman, R.W.; Abdalla, T.; Heijstra, B.D.; Behrendorff, J.B.; Tappel, R.; Kopke, M.; Simpson, S.D.; et al. Maintenance of ATP Homeostasis Triggers Metabolic Shifts in Gas-Fermenting Acetogens. Cell Syst. 2017, 4, 505–515. [Google Scholar] [CrossRef]
- Khider, M.L.K.; Brautaset, T.; Irla, M. Methane monooxygenases: Central enzymes in methanotrophy with promising biotechnological applications. World J. Microbiol. Biotechnol. 2021, 37, 72. [Google Scholar] [CrossRef]
- Wendlandt, K.D.; Jechorek, M.; Helm, J.; Stottmeister, U. Producing poly-3-hydroxybutyrate with a high molecular mass from methane. J. Biotechnol. 2001, 86, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Salem, R.; Soliman, M.; Fergala, A.; Audette, G.E.; ElDyasti, A. Screening for Methane Utilizing Mixed Communities with High Polyhydroxybutyrate (PHB) Production Capacity Using Different Design Approaches. Polymers 2021, 13, 1579. [Google Scholar] [CrossRef] [PubMed]
- Ghoddosi, F.; Golzar, H.; Yazdian, F.; Khosravi-Darani, K.; Vasheghani-Farahani, E. Effect of carbon sources for PHB production in bubble column bioreactor: Emphasis on improvement of methane uptake. J. Environ. Chem. Eng. 2019, 7, 102978. [Google Scholar] [CrossRef]
- Zuñiga, C.; Morales, M.; Revah, S. Polyhydroxyalkanoates accumulation by Methylobacterium organophilum CZ-2 during methane degradation using citrate or propionate as cosubstrates. Bioresour. Technol. 2013, 129, 686–689. [Google Scholar] [CrossRef] [PubMed]
- López, J.C.; Arnaiz, E.; Merchan, L.; Lebrero, R.; Munoz, R. Biogas-based polyhydroxyalkanoates production by Methylocystis hirsuta: A step further in anaerobic digestion biorefineries. Chem. Eng. J. 2018, 333, 529–536. [Google Scholar] [CrossRef]
- Fergala, A.; AlSayed, A.; Khattab, S.; Ramirez, M.; Eldyasti, A. Development of Methane-Utilizing Mixed Cultures for the Production of Polyhydroxyalkanoates (PHAs) from Anaerobic Digester Sludge. Environ. Sci. Technol. 2018, 52, 12376–12387. [Google Scholar] [CrossRef] [PubMed]
- Myung, J.; Flanagan, J.C.A.; Waymouth, R.M.; Criddle, C.S. Expanding the range of polyhydroxyalkanoates synthesized by methanotrophic bacteria through the utilization of omega-hydroxyalkanoate co-substrates. AMB Express 2017, 7, 118. [Google Scholar] [CrossRef] [PubMed]
- Cal, A.J.; Sikkema, W.D.; Ponce, M.I.; Franqui-Villanueva, D.; Riiff, T.J.; Orts, W.J.; Pieja, A.J.; Lee, C.C. Methanotrophic production of polyhydroxybutyrate-co-hydroxyvalerate with high hydroxyvalerate content. Int. J. Biol. Macromol. 2016, 87, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Olah, G.A. Beyond oil and gas: The methanol economy. Angew. Chem.-Int. Edit. 2005, 44, 2636–2639. [Google Scholar] [CrossRef]
- Mondal, U.; Yadav, G.D. Methanol economy and net zero emissions: Critical analysis of catalytic processes, reactors and technologies. Green Chem. 2021, 23, 8361–8405. [Google Scholar] [CrossRef]
- Bourque, D.; Pomerleau, Y.; Groleau, D. High cell density production of poly-beta-hydroxybutyrate (PHB) from methanol by Methylobacterium extorquens: Production of high-molecular-mass PHB. Appl. Microbiol. Biotechnol. 1995, 44, 367–376. [Google Scholar] [CrossRef]
- Orita, I.; Nishikawa, K.; Nakamura, S.; Fukui, T. Biosynthesis of polyhydroxyalkanoate copolymers from methanol by Methylobacterium extorquens AM1 and the engineered strains under cobalt-deficient conditions. Appl. Microbiol. Biotechnol. 2014, 98, 3715–3725. [Google Scholar] [CrossRef]
- Chang, W.; Yoon, J.; Oh, M.-K. Production of Polyhydroxyalkanoates with the Fermentation of Methylorubrum extorquens Using Formate as a Carbon Substrate. Biotechnol. Bioprocess Eng. 2022, 27, 268–275. [Google Scholar] [CrossRef]
- Stöckl, M.; Harms, S.; Dinges, I.; Dimitrova, S.; Holtmann, D. From CO2 to Bioplastic—Coupling the Electrochemical CO2 Reduction with a Microbial Product Generation by Drop-in Electrolysis. ChemSusChem 2020, 13, 4086–4093. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cao, Y.; Li, F.; Tian, Y.; Song, H. Enzyme-Assisted Microbial Electrosynthesis of Poly(3-hydroxybutyrate) via CO2 Bioreduction by Engineered Ralstonia eutropha. ACS Catal. 2018, 8, 4429–4437. [Google Scholar] [CrossRef]
- Kourilova, X.; Novackova, I.; Koller, M.; Obruca, S. Evaluation of mesophilic Burkholderia sacchari, thermophilic Schlegelella thermodepolymerans and halophilic Halomonas halophila for polyhydroxyalkanoates production on model media mimicking lignocellulose hydrolysates. Bioresour. Technol. 2021, 325, 124704. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-L.; Bhatia, S.K.; Gurav, R.; Choi, T.-R.; Kim, H.J.; Song, H.-S.; Park, J.-Y.; Han, Y.-H.; Lee, S.M.; Park, S.L.; et al. Fructose based hyper production of poly-3-hydroxybutyrate from Halomonas sp. YLGW01 and impact of carbon sources on bacteria morphologies. Int. J. Biol. Macromol. 2020, 154, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.K.; Yoon, J.-J.; Kim, H.-J.; Hong, J.W.; Hong, Y.G.; Song, H.-S.; Moon, Y.-M.; Jeon, J.-M.; Kim, Y.-G.; Yang, Y.-H. Engineering of artificial microbial consortia of Ralstonia eutropha and Bacillus subtilis for poly(3-hydroxybutyrate-co-3-hydroxyvalerate) copolymer production from sugarcane sugar without precursor feeding. Bioresour. Technol. 2018, 257, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Amini, M.; Yousefi-Massumabad, H.; Younesi, H.; Abyar, H.; Bahramifar, N. Production of the polyhydroxyalkanoate biopolymer by Cupriavidus necator using beer brewery wastewater containing maltose as a primary carbon source. J. Environ. Chem. Eng. 2020, 8, 103588. [Google Scholar] [CrossRef]
- Dalsasso, R.R.; Pavan, F.A.; Bordignon, S.E.; Falcao de Aragdo, G.M.; Poletto, P. Polyhydroxybutyrate (PHB) production by Cupriavidus necator from sugarcane vinasse and molasses as mixed substrate. Process. Biochem. 2019, 85, 12–18. [Google Scholar] [CrossRef]
- Lim, S.W.; Kansedo, J.; Tan, I.S.; Tan, Y.H.; Nandong, J.; Lam, M.K.; Ongkudon, C.M. Microbial valorization of oil-based substrates for polyhydroxyalkanoates (PHA) production—Current strategies, status, and perspectives. Process. Biochem. 2023, 130, 715–733. [Google Scholar] [CrossRef]
- Volova, T.; Sapozhnikova, K.; Zhila, N. Cupriavidus necator B-10646 growth and polyhydroxyalkanoates production on different plant oils. Int. J. Biol. Macromol. 2020, 164, 121–130. [Google Scholar] [CrossRef]
- Purama, R.K.; Al-Sabahi, J.N.; Sudesh, K. Evaluation of date seed oil and date molasses as novel carbon sources for the production of poly(3Hydroxybutyrate-co-3Hydroxyhexanoate) by Cupriavidus necator H16 Re 2058/pCB113. Ind. Crop. Prod. 2008, 119, 83–92. [Google Scholar] [CrossRef]
- Santolin, L.; Waldburger, S.; Neubauer, P.; Riedel, S.L. Substrate-Flexible Two-Stage Fed-Batch Cultivations for the Production of the PHA Copolymer P(HB-co-HHx) With Cupriavidus necator Re2058/pCB113. Front. Bioeng. Biotechnol. 2021, 9, 623890. [Google Scholar] [CrossRef]
- Schmid, M.T.; Sykacek, E.; O’Connor, K.; Omann, M.; Mundigler, N.; Neureiter, M. Pilot scale production and evaluation of mechanical and thermal properties of P(3HB) from Bacillus megaterium cultivated on desugarized sugar beet molasses. J. Appl. Polym. Sci. 2022, 139, e51503. [Google Scholar] [CrossRef]
- Tu, W.-L.; Chu, H.-K.; Huang, C.-M.; Chen, C.-H.; Ou, C.-M.; Guo, G.-L. Polyhydroxyalkanoate Production by Cupriavidus necator with Inedible Rice. BioResources 2022, 17, 2202–2213. [Google Scholar] [CrossRef]
- Saratale, R.G.; Cho, S.K.; Saratale, G.D.; Ghodake, G.S.; Bharagava, R.N.; Kim, D.S.; Nair, S.; Shin, H.S. Efficient bioconversion of sugarcane bagasse into polyhydroxybutyrate (PHB) by Lysinibacillus sp. and its characterization. Bioresour. Technol. 2021, 324, 124673. [Google Scholar] [CrossRef]
- Lee, S.M.; Lee, H.-J.; Kim, S.H.; Suh, M.J.; Cho, J.Y.; Ham, S.; Jeon, J.-M.; Yoon, J.-J.; Bhatia, S.K.; Gurav, R.; et al. Screening of the strictly xylose-utilizing Bacillus sp. SM01 for polyhydroxybutyrate and its co-culture with Cupriavidus necator NCIMB 11599 for enhanced production of PHB. Int. J. Biol. Macromol. 2021, 181, 410–417. [Google Scholar] [CrossRef]
- Kalaiyezhini, D.; Ramachandran, K.B. Biosynthesis of Poly-3-Hydroxybutyrate (PHB) from Glycerol by Paracoccus denitrificans in a Batch Bioreactor: Effect of Process Variables. Prep. Biochem. Biotechnol. 2015, 45, 69–83. [Google Scholar] [CrossRef]
- Fauzi, A.H.M.; Chua, A.S.M.; Yoon, L.W.; Nittami, T.; Yeoh, H.K. Enrichment of PHA-accumulators for sustainable PHA production from crude glycerol. Process. Saf. Environ. Protect. 2019, 122, 200–208. [Google Scholar] [CrossRef]
- Tanadchangsaeng, N.; Yu, J. Miscibility of natural polyhydroxyalkanoate blend with controllable material properties. J. Appl. Polym. Sci. 2013, 129, 2004–2016. [Google Scholar] [CrossRef]
- Vijay, R.; Tarika, K. Microbial Production of Polyhydroxyalkanoates (Phas) Using Kitchen Waste as an Inexpensive Carbon Source. Biosci. Biotechnol. Res. Asia 2019, 16, 155–166. [Google Scholar] [CrossRef]
- Das, S.; Majumder, A.; Shukla, V.; Suhazsini, P.; Radha, P. Biosynthesis of Poly(3-hydroxybutyrate) from Cheese Whey by Bacillus megaterium NCIM 5472. J. Polym. Environ. 2018, 26, 4176–4187. [Google Scholar] [CrossRef]
- Loan, T.T.; Trang, D.T.Q.; Huy, P.Q.; Ninh, P.X.; Van Thuoc, D. A fermentation process for the production of poly(3-hydroxybutyrate) using waste cooking oil or waste fish oil as inexpensive carbon substrate. Biotechnol. Rep. 2022, 33, e00700. [Google Scholar] [CrossRef] [PubMed]
- Sangkharak, K.; Khaithongkaeo, P.; Chuaikhunupakarn, T.; Choonut, A.; Prasertsan, P. The production of polyhydroxyalkanoate from waste cooking oil and its application in biofuel production. Biomass Convers Biorefin. 2021, 11, 1651–1664. [Google Scholar] [CrossRef]
- Thinagaran, L.; Sudesh, K. Evaluation of Sludge Palm Oil as Feedstock and Development of Efficient Method for its Utilization to Produce Polyhydroxyalkanoate. Waste Biomass Valorization 2019, 10, 709–720. [Google Scholar] [CrossRef]
- Obruca, S.; Marova, I.; Snajdar, O.; Mravcova, L.; Svoboda, Z. Production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by Cupriavidus necator from waste rapeseed oil using propanol as a precursor of 3-hydroxyvalerate. Biotechnol. Lett. 2010, 32, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Pernicova, I.; Kucera, D.; Nebesarova, J.; Kalina, M.; Novackova, I.; Koller, M.; Obruca, S. Production of polyhydroxyalkanoates on waste frying oil employing selected Halomonas strains. Bioresour. Technol. 2019, 292, 122028. [Google Scholar] [CrossRef] [PubMed]
- Goff, M.; Ward, P.G.; O’Connor, K.E. Improvement of the conversion of polystyrene to polyhydroxyalkanoate through the manipulation of the microbial aspect of the process: A nitrogen feeding strategy for bacterial cells in a stirred tank reactor. J. Biotechnol. 2007, 132, 283–286. [Google Scholar] [CrossRef]
- Merli, G.; Becci, A.; Amato, A.; Beolchini, F. Acetic acid bioproduction: The technological innovation change. Sci. Total. Environ. 2021, 798, 149292. [Google Scholar] [CrossRef]
- Christensen, M.; Jablonski, P.; Altermark, B.; Irgum, K.; Hansen, H. High natural PHA production from acetate in Cobetia sp. MC34 and Cobetia marina DSM 4741(T) and in silico analyses of the genus specific PhaC(2) polymerase variant. Microb. Cell. Fact. 2021, 20, 225. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalez, L.; De Wever, H. Acetic Acid as an Indirect Sink of CO2 for the Synthesis of Polyhydroxyalkanoates (PHA): Comparison with PHA Production Processes Directly Using CO2 as Feedstock. Appl. Sci. 2019, 8, 1416. [Google Scholar] [CrossRef]
- Tao, G.-B.; Tian, L.; Pu, N.; Li, Z.-J. Efficient production of poly-3-hydroxybutyrate from acetate and butyrate by halophilic bacteria Salinivibrio spp. TGB4 and TGB19. Int. J. Biol. Macromol. 2022, 221, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Marang, L.; Jiang, Y.; van Loosdrecht, M.C.M.; Kleerebezem, R. Butyrate as preferred substrate for polyhydroxybutyrate production. Bioresour. Technol. 2013, 142, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-M.; Brigham, C.J.; Kim, Y.-H.; Kim, H.-J.; Yi, D.-H.; Kim, H.; Rha, C.; Sinskey, A.J.; Yang, Y.-H. Biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (P(HB-co-HHx)) from butyrate using engineered Ralstonia eutropha. Appl. Microbiol. Biotechnol. 2014, 98, 5461–5469. [Google Scholar] [CrossRef] [PubMed]
- Zhila, N.O.; Sapozhnikova, K.Y.; Kiselev, E.G.; Nemtsev, I.; Lukyanenko, A.; Shishatskaya, E.; Volova, T.G. Biosynthesis and Properties of a P(3HB-co-3HV-co-4HV) Produced by Cupriavidus necator B-10646. Polymers 2022, 14, 4226. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Jiao, N. Formation of Polyhydroxyalkanoate in Aerobic Anoxygenic Phototrophic Bacteria and Its Relationship to Carbon Source and Light Availability. Appl. Environ. Microbiol. 2011, 77, 7445–7450. [Google Scholar] [CrossRef]
- Koch, M.; Bruckmoser, J.; Scholl, J.; Hauf, W.; Rieger, B.; Forchhammer, K. Maximizing PHB content in Synechocystis sp. PCC 6803: A new metabolic engineering strategy based on the regulator PirC. Microb. Cell Fact. 2020, 19, 231. [Google Scholar] [CrossRef]
- Cortes, O.; Guerra-Blanco, P.; Chairez, I.; Poznyak, T.; Garcia-Pena, E.I. Polymers, the Light at the End of Dark Fermentation: Production of Poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by a Photoheterotrophic Consortium. J. Polym. Environ. 2022, 30, 2392–2404. [Google Scholar] [CrossRef]
- Brandl, H.; Gross, R.; Lenz, R.; Lloyd, R.; Fuller, R. The Accumulation of Poly(3-Hydroxyalkanoates) in Rhodobacter sphaeroides. Arch. Microbiol. 1991, 155, 337–340. [Google Scholar] [CrossRef]
- Nemmour, A.; Inayat, A.; Janajreh, I.; Ghenai, C. Green hydrogen-based E-fuels (E-methane, E-methanol, E-ammonia) to support clean energy transition: A literature review. Int. J. Hydrog. 2023, 48, 29011–29033. [Google Scholar] [CrossRef]
- NIST database of National Institute of Standards and Technology, U.S. Department of Commerce Webbook. Available online: https://webbook.nist.gov/ (accessed on 29 March 2024).
- PubChem Database of National Library of Medicine, Nacional Center for Biotechnology Information. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 29 March 2024).
- Eberle, U.; Felderhoff, M.; Schueth, F. Chemical and Physical Solutions for Hydrogen Storage. Angew. Chem.-Int. Edit. 2009, 48, 6608–6630. [Google Scholar] [CrossRef] [PubMed]
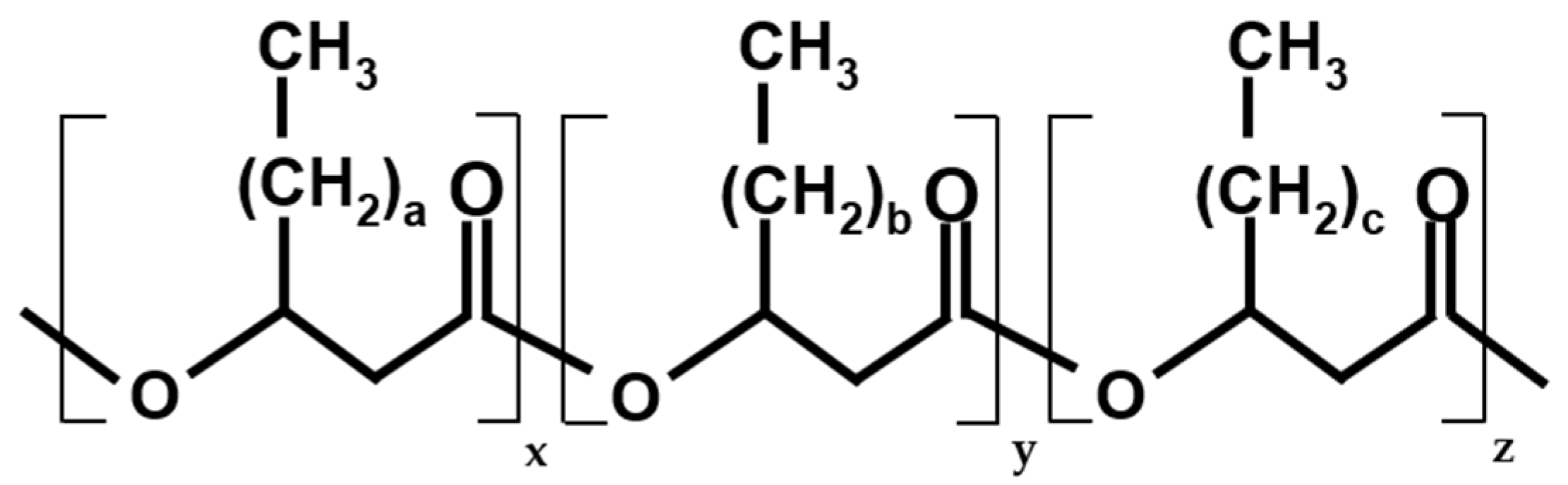
| PHB:PHV:PHH | Tm (°C) | TS (MPa) | EaB (%) | Reference |
|---|---|---|---|---|
| 93:0:17 | 120 | 20 | 850 | [6] |
| 85:0:15 | 115 | 23 | 760 | [6] |
| 69:24:7 | 129, 139 | 20 | 710 | [7] |
| 90:0:10 | 127 | 21 | 400 | [6] |
| 71:26:3 | 131, 143 | 12 | 324 | [8] |
| 80:16:4 | 140, 151 | 12 | 321 | [8] |
| 91:2:7 | 144 | 22 | 312 | [8] |
| 87:9:4 | 142, 153 | 21 | 287 | [8] |
| 66:32:2 | 91, 148 | 5 | 282 | [7] |
| 82:15:3 | 147 | 6 | 84 | [7] |
| 94:3:3 | 129, 144 | 4 | 79 | [7] |
| 14:85:1 | 89 | 14.5 | 78 | [7] |
| 80:20:0 | 170–175 * | 30.8 | 54 | [9] |
| 0:100:0 | 130–135 * | 31.2 | 14 | [9] |
| P4HB 1 | 53 | 104 | 1000 | [10] |
| PHP 2 | 78 | 28.3 | 634 | [11] |
| PP 3 | 170 | 34 | 400 | [12] |
| Source of Energy | Source of Electrons | Source of Carbon | |
|---|---|---|---|
| chemical bond | inorganic compound | oxidized C | |
| chemo- | -litho- | -auto- | -troph |
| photo- | -organo- | -hetero- | |
| electromagnetic radiation | organic compound | reduced C |
| Autotrophy | Methylotrophy | Heterotrophy |
|---|---|---|
| CO2 + sunlight | Methane | Food products |
| CO2 + H2 | Methanol | Food and other industry waste |
| CO | Formate | Hydrocarbons |
| Short carboxylic acids | ||
| Carboxylic acids + sunlight |
| Strain | Growth Source | PHA | PHB | PHV | Other | Reference |
|---|---|---|---|---|---|---|
| Synechocystis sp. PCC 6714 | CO2 + H2O + γ | 37 | [76] | |||
| Synechocystis sp. PCC 6803 | CO2 + H2O + γ | 39 | [77] | |||
| Oscillatoria okeni | CO2 + H2O + γ | 14 | 6 | [78] | ||
| Oscillatoria okeni | CO2 + H2O + γ + acetate | 42 | 7 | [78] | ||
| Anabena sp. | CO2 + H2O + γ | 40 | 5 | [79] | ||
| Anabena sp. | CO2 + H2O + γ + acetate | 14 | 24 | [79] | ||
| C. necator | CO2 + H2 | 85 | [80] | |||
| C. necator | CO2 + H2 | 55 | 20 (PHH) | [81] | ||
| C. necator | CO2 + H2 | 46 | 39 (PHD) | [81] | ||
| C. necator | CO2 + H2 | 42 | 55 (PHO) | [81] | ||
| Seliberia carboxydohydrogena | CO | 63 | [87] | |||
| Clostridrium autoethanogenum | CO | 6 | [89] | |||
| Methylocystis sp. GB 25 | CH4 | 51 | [92] | |||
| Methylophilus + Methylocystis | CH4 | 59 | [93] | |||
| Methylocystis hirsuta | CH4 + methanol, ethanol | 73 | [94] | |||
| Methylobacterium organophilum | CH4 | 60 | 5 | [95] | ||
| Methylobacterium organophilum | CH4 + citrate | 88 | 55 | 35 (PHO), 10 (PHD) | [95] | |
| Methylobacterium organophilum | CH4 + propionate | 60 | 75 | 25 (PHO) | [95] | |
| Methylocystis hirsuta | CH4 + valerate | 54 | 25 | [96] | ||
| Enriched sludge | CH4 + valerate | 52 | 33 | [97] | ||
| Methylocystis parvus | CH4 +4-hydroxybutyrate | 50 | 10 (P4HB) | [98] | ||
| Methylocystis sp. WRRC1 | CH4 + valerate | 60 | 50 | [99] | ||
| Methylobacterium extorquens | methanol | 53 | [102] | |||
| Methylobacterium extorquens | methanol | 44 | [103] | |||
| Methylobacterium extorquens | formate | 43 | [104] | |||
| C. necator | Formate-dropping | 34 | [105] | |||
| Halomonas halophilaHalomonas bluephagenesis | glucose | 85 | [107] | |||
| glucose + acetate | 94 | [56] | ||||
| Halomonas sp. YLGW01 | fructose | 95 | [108] | |||
| Bacillus subtilis + C. necator | sugarcane sugar | 75 | [109] | |||
| C. necator | brewery wastewater | 92 | [110] | |||
| C. necator | hydrolyzed sugarcane molasse | 58 | [111] | |||
| C. necator | crude corn starch | 63 | [65] | |||
| C. necator | palm oil | 72 | [112] | |||
| C. necator | date seed oil | 81 | [114] | |||
| C. necator | fructose + rapeseed oil | 86 | 17 (PHH) | [115] | ||
| Methylobacterium organophilum | citrate | 48 | 35 | 53 | 12 (PHO) | [95] |
| Bacillus megatherium | desugarized sugar beet molasse | 68 | [116] | |||
| C. necator | hydrolyzed inedible rice | 69 | [117] | |||
| Lysinibacillus sp. | hydrolyzed sugarcane bagasse | 62 | [118] | |||
| Bacillus sp. SM01 | xylose form lignocellulose | 62 | [119] | |||
| Paracoccus denitrificans | glycerol | 72 | [120] | |||
| Activated sludge | glycerol | 80 | [121] | |||
| C. necator | glycerol + levulinic acid | 80 | [122] | |||
| Bacillus subtilis | onion peels | 89 | [123] | |||
| Bacillus siamensis | orange peels | 82 | [123] | |||
| Bacillus megatherium | cheese whey | 76 | [124] | |||
| C. necator | slaughterhouse waste fats | 66 | [66] | |||
| C. necator | waste fish oil | 73 | [125] | |||
| Bacillus thermoamylovorans | waste cooking oil | 88 | [126] | |||
| C. necator | sludge palm oil | 74 | 22 (PHO) | [127] | ||
| C. necator | waste rapeseed oil + propanol | 80 | 9 | [125] | ||
| Halomonas hydrothermalis | waste frying oil + valeric acid | 69 | [129] | |||
| Halomonas hydrothermalis | waste frying oil + propanol | 70 | [129] | |||
| Pseudomonas putida | styrene | 42 | [130] | |||
| Cobetia sp. MC34 | acetate | 72 | [132] | |||
| C. necator | acetate | 72 | [133] | |||
| Salinivibrio sp. TGB19 | acetate + butyrate | 89 | [134] | |||
| Plasticicumulans acidovorans | butyrate | 88 | [135] | |||
| C. necator | butyrate | 66 | 13 | [136] | ||
| C. necator | butyrate + 4-valerolactone | 78 | 31 | [137] | ||
| Methylobacterium organophilum | propionate | 37 | 37 | 56 | 7 (PHD) | [95] |
| Dinoroseobacter sp. JL1447 | acetate + γ | 72 | [138] | |||
| Oscillatoria okeni | acetate + γ | 42 | 7 | [78] | ||
| Synechocystic sp. PCC 6803 | acetate + γ | 81 | [139] | |||
| Bacterial consortium | butyrate + acetate + γ | 67 | [140] | |||
| Rhodobacter sphaeroides | acetate + malate + γ | 72 | 2 | [141] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukala, I.; Kučera, I. Natural Polyhydroxyalkanoates—An Overview of Bacterial Production Methods. Molecules 2024, 29, 2293. https://doi.org/10.3390/molecules29102293
Fukala I, Kučera I. Natural Polyhydroxyalkanoates—An Overview of Bacterial Production Methods. Molecules. 2024; 29(10):2293. https://doi.org/10.3390/molecules29102293
Chicago/Turabian StyleFukala, Ivo, and Igor Kučera. 2024. "Natural Polyhydroxyalkanoates—An Overview of Bacterial Production Methods" Molecules 29, no. 10: 2293. https://doi.org/10.3390/molecules29102293
APA StyleFukala, I., & Kučera, I. (2024). Natural Polyhydroxyalkanoates—An Overview of Bacterial Production Methods. Molecules, 29(10), 2293. https://doi.org/10.3390/molecules29102293