
Terpenic Constituents of Essential Oils with Larvicidal Activity against Aedes Aegypti: A QSAR and Docking Molecular Study


 , , , and
, , , and 
Abstract
1. Introduction
2. Results
2.1. Quantitative Structure–Activity Relationship
2.2. DFT Studies
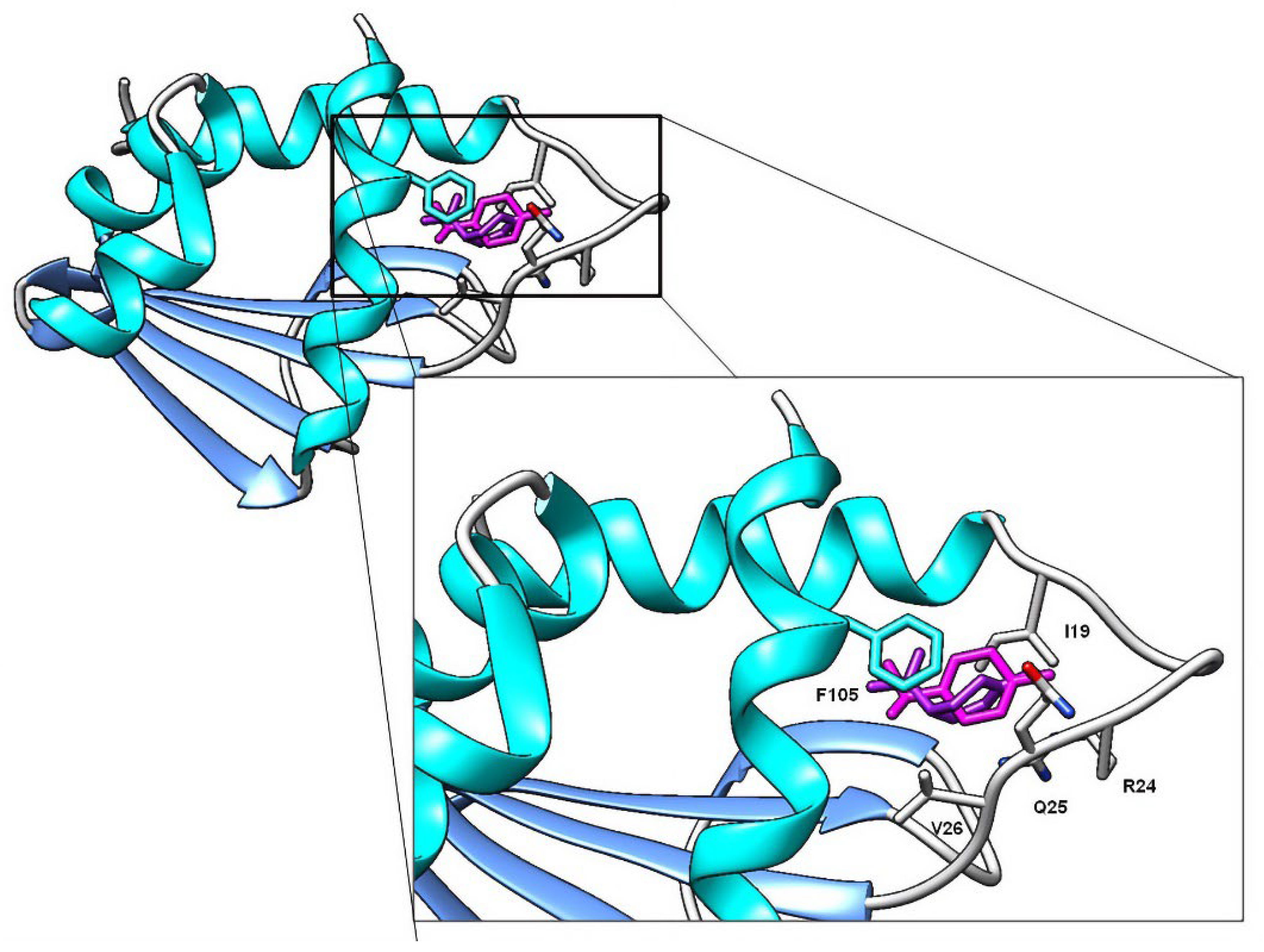
2.3. Molecular Docking Studies
3. Discussion
4. Materials and Methods
4.1. Larvicidal Activities
4.2. In Silico Optimization and Descriptors Calculation
4.3. Structure–Property–Larvicidal Activity Models
4.4. Molecular Docking Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weaver, S.C.; Lecuit, M. Chikungunya virus and the global spread of a mosquito-borne disease. N. Engl. J. Med. 2015, 372, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Murray, N.E.A.; Quam, M.B.; Wilder-Smith, A. Epidemiology of dengue: Past, present, and future prospects. J. Clin. Epidemiol. 2013, 5, 299. [Google Scholar] [CrossRef]
- Tong, M.X.; Hansen, A.; Hanson-Easey, S.; Xiang, J.; Cameron, S.; Liu, Q.; Williams, C. Perceptions of capacity for infectious disease control and prevention to meet the challenges of dengue fever in the face of climate change: A survey among CDC staff in Guangdong Province, China. Environ. Res. 2016, 148, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.P.; Luther, C.; Moo-Llanes, D.; Ramsey, J.M.; Danis-Lozano, R.; Peterson, A.T. Climate change influences on global distributions of dengue and chikungunya virus vectors. Philos. Trans R. Soc. Lond. B Biol. Sci. 2015, 370, 20140135. [Google Scholar] [CrossRef]
- Aguirre-Obando, O.A.; Bona, A.C.D.; Duque, L.J.E.; Navarro-Silva, M.A. Insecticide resistance and genetic variability in natural populations of Aedes (Stegomyia) aegypti (Diptera: Culicidae) from Colombia. Zoologia Curitiba 2015, 32, 14–22. [Google Scholar] [CrossRef]
- Ryan, M.F.; Byrne, O. Plant-insect coevolution and inhibition of acetylcholinesterase. J. Chem. Ecol. 1988, 14, 1965–1975. [Google Scholar] [CrossRef]
- Massebo, F.; Tadesse, M.; Bekele, T.; Balkew, M.; Gebre-Michael, T. Evaluation on larvicidal effects of essential oils of some local plants against Anopheles arabiensis Patton and Aedes aegypti Linnaeus (Diptera, Culicidae) in Ethiopia. Afr. J. Biotech. 2009, 8, 4183–4188. [Google Scholar]
- Andrade-Ochoa, S.; Sánchez-Torres, L.E.; Nevárez-Moorillón, G.V.; Camacho, A.D.; Nogueda-Torres, B. Essential oils and their components as an alternative in the control of mosquito vectors of disease. Biomédica 2017, 37, 224–243. [Google Scholar] [CrossRef]
- Castillo, S.; Pérez-Alfonso, C.O.; Martínez-Romero, D.; Guillén, F.; Serrano, M.; Valero, D. The essential oils thymol and carvacrol applied in the packing lines avoid lemon spoilage and maintain quality during storage. Food Control 2014, 35, 132–136. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils–a review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Kaur, S. Eucalyptus essential oil as a natural pesticide. Forest. Ecol. Manag. 2008, 256, 2166–2174. [Google Scholar] [CrossRef]
- Gillij, Y.G.; Gleiser, R.M.; Zygadlo, J.A. Mosquito repellent activity of essential oils of aromatic plants growing in Argentina. Bioresour. Technol. 2008, 99, 2507–2515. [Google Scholar] [CrossRef]
- Moore, S.J.; Hill, N.; Ruiz, C.; Cameron, M.M. Field evaluation of traditionally used plant-based insect repellents and fumigants against the malaria vector Anopheles darlingi in Riberalta, Bolivian Amazon. J. Med. Entomol. 2007, 44, 624–630. [Google Scholar] [CrossRef]
- Tennyson, S.; Samraj, D.A.; Jeyasundar, D.; Chalieu, K. Larvicidal efficacy of plant oils against the dengue vector Aedes aegypti (L.) (Diptera: Culicidae). Middle-East J. Sci. Res. 2013, 13, 64–68. [Google Scholar] [CrossRef]
- Vera, S.S.; Zambrano, D.F.; Méndez-Sánchez, S.C.; Rodríguez-Sanabria, F.; Stashenko, E.E.; Luna, J.E. Essential oils with insecticidal activity against larvae of Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2014, 113, 2647–2654. [Google Scholar] [CrossRef]
- Lahlou, M. Methods to study the phytochemistry and bioactivity of essential oils. Phytother. Res. 2004, 18, 435–448. [Google Scholar] [CrossRef]
- Shaalan, E.A.S.; Canyon, D.; Younes, M.W.; Abdel-Wahab, H.; Mansour, A.H. A review of botanical phytochemicals with mosquitocidal potential. Environ. Int. 2005, 31, 1149–1166. [Google Scholar] [CrossRef]
- Pavela, R. Acute and synergistic effects of some monoterpenoid essential oil compounds on the house fly (Musca domestica L.). J. Essent. Oil Bear. Pl. 2008, 11, 451–459. [Google Scholar] [CrossRef]
- Koul, O.; Singh, R.; Kaur, B.; Kanda, D. Comparative study on the behavioral response and acute toxicity of some essential oil compounds and their binary mixtures to larvae of Helicoverpa armigera, Spodoptera litura and Chilo partellus. Ind. Crops Prod. 2013, 49, 428–436. [Google Scholar] [CrossRef]
- Hummelbrunner, L.A.; Isman, M.B. Acute, sublethal, anti-feedant, and synergistic effects of monoterpenoid essential oil compounds on the tobacco cutworm, Spodoptera litura (Lep., Noctuidae). J. Agric. Food Chem. 2001, 49, 715–720. [Google Scholar] [CrossRef]
- Lima, T.C.; da Silva, T.K.M.; Silva, F.L.; Barbosa-Filho, J.M.; Marques, M.O.M.; Santos, R.L.C.; de Sousa, D.P. Larvicidal activity of Mentha × villosa Hudson essential oil, rotundifolone and derivatives. Chemosphere 2014, 104, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Kim, J.R.; Wang, M.; Shu, S.; Ahn, Y.J. Larvicidal activity of Conidium monnieri fruit coumarins and structurally related compounds against insecticide-susceptible and insecticide-resistant Culex pipiens pallens and Aedes aegypti. Pest Manag. Sci. 2012, 68, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Lucia, A.; Zerba, E.; Masuh, H. Knockdown and larvicidal activity of six monoterpenes against Aedes aegypti (Diptera: Culicidae) and their structure-activity relationships. Parasitol. Res. 2013, 112, 4267–4272. [Google Scholar] [CrossRef] [PubMed]
- Lomonaco, D.; Santiago, G.M.P.; Ferreira, Y.S.; Arriaga, Â.M.C.; Mazzetto, S.E.; Mele, G.; Vasapollo, G. Study of technical CNSL and its main components as new green larvicides. Green Chem. 2009, 11, 31–33. [Google Scholar] [CrossRef]
- Gao, W.; Siddiqui, M.K.; Imran, M.; Jamil, M.K.; Farahani, M.R. Forgotten topological index of chemical structure in drugs. Saudi Pharm. J. 2016, 24, 258–264. [Google Scholar] [CrossRef]
- Consonni, V.; Todeschini, R.; Pavan, M. Structure/response correlations and similarity/diversity analysis by GETAWAY descriptors. 1. Theory of the novel 3D molecular descriptors. J. Chem. Inf. Comput. Sci. 2002, 42, 682–692. [Google Scholar] [CrossRef]
- Hu, Q.N.; Liang, Y.Z.; Ren, F.L. Molecular graph center, a novel approach to locate the center of a molecule and a new centric index. Theochem-J. Mol. Struct. 2003, 635, 105–113. [Google Scholar] [CrossRef]
- Bonchev, D.; Mekenyan, O.V.A.N.E.S.; Balaban, A.T. Iterative procedure for the generalized graph center in polycyclic graphs. J. Chem. Inf. Comput. Sci. 1989, 29, 91–97. [Google Scholar] [CrossRef]
- Pearson, R.G. Absolute electronegativity and hardness: Application to inorganic chemistry. Inorg. Chem. 1988, 27, 734–740. [Google Scholar] [CrossRef]
- Paluch, G.; Grodnitzky, J.; Bartholomay, L.; Coats, J. Quantitative structure− activity relationship of botanical sesquiterpenes: Spatial and contact repellency to the yellow fever mosquito, Aedes aegypti. J. Agric. Food Chem. 2009, 57, 7618–7625. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Sánchez-Aldana, D.; Chacón-Vargas, K.F.; Rivera-Chavira, B.E.; Sánchez-Torres, L.E.; Camacho, A.D.; Nevárez-Moorillón, G.V. Oviposition deterrent and larvicidal and pupaecidal activity of seven essential oils and their major components against Culex quinquefasciatus Say (Diptera: Culicidae): Synergism–antagonism effects. Insects 2018, 9, 25. [Google Scholar] [CrossRef]
- De Oliveira, A.A.; França, L.P.; Ramos, A.D.S.; Ferreira, J.L.P.; Maria, A.C.B.; Oliveira, K.M.; Araújo, E.S.; da Silv Branches, J.N.; Branches, A.D.S.; Barros, G.d.A.; et al. Larvicidal, adulticidal and repellent activities against Aedes aegypti L. of two commonly used spices, Origanum vulgare L. and Thymus vulgaris L. S. Afr. J. Bot. 2021, 140, 17–24. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Ma, G.; Yang, F.; Zhan, Z.; Guan, L.; Kuang, W.; Wamg, J.; Li, J.; Han, F.; et al. Piperonyl butoxide synergizes the larvicidal activity of Origanum vulgare essential oil and its major constituents against the larvae of Aedes albopictus and Culex pipiens quinquefasciatus. J. Asia Pac. Entomol. 2023, 26, 102025. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Chacón-Vargas, K.F.; Sánchez-Torres, L.E.; Rivera-Chavira, B.E.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Differential antimicrobial effect of essential oils and their main components: Insights based on the cell membrane and external structure. Membranes 2021, 11, 405. [Google Scholar] [CrossRef]
- Youssefi, M.R.; Tabari, M.A.; Esfandiari, A.; Kazemi, S.; Moghadamnia, A.A.; Sut, S.; Maggi, F. Efficacy of two monoterpenoids, carvacrol and thymol, and their combinations against eggs and larvae of the West Nile vector Culex pipiens. Molecules 2019, 24, 1867. [Google Scholar] [CrossRef]
- Silva, V.B.; Travassos, D.L.; Nepel, A.; Barison, A.; Costa, E.V.; Scotti, L.; de Holanda Cavalcanti, S.C. Synthesis and chemometrics of thymol and carvacrol derivatives as larvicides against Aedes aegypti. J. Arthropod. Borne Dis. 2017, 11, 315. [Google Scholar]
- Senra, T.O.S.; Calmon, F.; Zeringóta, V.; Monteiro, C.M.O.; Maturano, R.; da Silva Matos, R.; Daemon, E. Investigation of activity of monoterpenes and phenylpropanoids against immature stages of Amblyomma cajennense and Rhipicephalus sanguineus (Acari: Ixodidae). Parasit Res. 2013, 112, 3471–3476. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Effects of monoterpenoid insecticides on [3H]-TBOB binding in house fly GABA receptor and 36Cl− uptake in American cockroach ventral nerve cord. Pestic. Biochem. Physiol. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Setlur, A.S.; Karunakaran, C.; Pandey, S.; Sarkar, M.; Niranjan, V. Molecular interaction studies of thymol via molecular dynamic simulations and free energy calculations using multi-target approach against Aedes aegypti proteome to decipher its role as mosquito repellent. Mol. Simul. 2022, 49, 1–16. [Google Scholar] [CrossRef]
- Li, Y.; Wu, W.; Jian, R.; Ren, X.; Chen, X.; Hong, W.D.; Sheng, Z. Larvicidal, acetylcholinesterase inhibitory activities of four essential oils and their constituents against Aedes albopictus, and nanoemulsion preparation. Pest Manag. Sci. 2022, 1–11. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Protect. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Zarrad, K.; Hamouda, A.B.; Chaieb, I.; Laarif, A.; Jemâa, J.M.B. Chemical composition, fumigant and anti-acetylcholinesterase activity of the Tunisian Citrus aurantium L. essential oils. Ind. Crops Prod. 2015, 76, 121–127. [Google Scholar] [CrossRef]
- Coats, J.R.; Karr, L.L.; Drewes, C.D. Toxicity and Neurotoxic Effects of Monoterpenoids in Insects and Earthworms. Natural Occurring Pest Bioregulators; American Chemical Society Symposium Series; Hedin, P., Ed.; American Chemical Society: Washington, DC, USA, 1991; pp. 305–316. [Google Scholar]
- Praveena, A.; Sanjayan, K.P. Inhibition of acetylcholinesterase in three insects of economic importance by linalool, a monoterpene phytochemical. In Insect Pest Management, A Current Scenario; Ambrose, D.P., Ed.; Entomology Research Unit, St. Xavier’s College: Palayamkottai, India, 2011; pp. 340–345. [Google Scholar]
- Seo, S.M.; Jung, C.S.; Kang, J.; Lee, H.R.; Kim, S.W.; Hyun, J.; Park, I.K. Larvicidal and acetylcholinesterase inhibitory activities of Apiaceae plant essential oils and their constituents against Aedes albopictus and formulation development. J. Agric. Food Chem. 2015, 63, 9977–9986. [Google Scholar] [CrossRef]
- Yeom, H.J.; Kang, J.S.; Kim, G.H.; Park, I.K. Insecticidal and acetylcholine esterase inhibition activity of Apiaceae plant essential oils and their constituents against adults of German cockroach (Blattella germanica). J. Agric. Food Chem. 2012, 60, 7194–7203. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Ochoa, S.; Correa-Basurto, J.; Rodríguez-Valdez, L.M.; Sánchez-Torres, L.E.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. In vitro and in silico studies of terpenes, terpenoids and related compounds with larvicidal and pupaecidal activity against Culex quinquefasciatus Say (Diptera: Culicidae). Chem. Cent. J. 2018, 12, 53. [Google Scholar] [CrossRef]
- Lan, Q.; Wessely, V. Expression of a sterol carrier protein-x gene in the Yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2004, 13, 519–529. [Google Scholar] [CrossRef]
- Ossendorp, B.C.; Van Heusden, G.P.H.; De Beer, A.L.; Bos, K.; Schouten, G.L.; Wirtz, K.W. Identification of the cDNA clone which encodes the 58-kDa protein containing the amino acid sequence of rat liver non-specific lipid-transfer protein (sterol-carrier protein 2) Homology with rat peroxisomal and mitochondrial 3-oxoacyl-CoA thiolases. Eur. J. Biochem. 1991, 201, 233–239. [Google Scholar] [CrossRef]
- Mannaerts, G.P.; Van Veldhoven, P.P.; Casteels, M. Peroxisomal lipid degradation via β-and α-oxidation in mammals. Cell Biochem. Biophys. 2000, 32, 73–87. [Google Scholar] [CrossRef]
- Antonenkov, V.D.; Van Veldhoven, P.P.; Waelkens, E.; Mannaerts, G.P. Substrate Specificities of 3-Oxoacyl-CoA Thiolase A and Sterol Carrier Protein 2/3-Oxoacyl-CoA Thiolase Purified from Normal Rat Liver Peroxisomes Sterol Carrier Protein 2/3-oxoacyl-CoA Thiolase is involved in the metabolism of 2-Methyl-Branched Fatty Acids and Bile Acid Intermediates. J. Biol. Chem. 1997, 272, 26023–26031. [Google Scholar] [CrossRef]
- Santos, S.R.; Melo, M.A.; Cardoso, A.V.; Santos, R.L.; de Sousa, D.P.; Cavalcanti, S.C. Structure–activity relationships of larvicidal monoterpenes and derivatives against Aedes aegypti Linn. Chemosphere 2011, 84, 150–153. [Google Scholar] [CrossRef]
- Deppmeier, B.J.; Driessen, A.J.; Hehre, T.S.; Hehre, W.J.; Johnson, J.A.; Klunzinger, P.E.; Jianguo, Y. Spartan ‘08, Build 132; Wavefunction Inc.: Irvine, CA, USA, 2009. [Google Scholar]
- Mayo, S.L.; Olafson, B.D.; Goddard, W.A. DREIDING: A generic force field for molecular simulations. J. Phys. Chem. 1990, 94, 8897–8909. [Google Scholar] [CrossRef]
- Stewart, J.J. Optimization of parameters for semiempirical methods II. Applications. J. Comput. Chem. 1989, 10, 221–264. [Google Scholar] [CrossRef]
- Talete S.R.L. Dragon for Windows, Version 5.4; Software for Molecular Descriptor Calculations; Talete S.R.L.: Milano, Italy, 2006.
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revision d. 01; Gaussian, Inc.: Wallingford, CT, USA, 2009.
- Becke, A.D. Becke’s three parameter hybrid method using the LYP correlation functional. J. Chem. Phys. 1993, 98, 5648–5652. [Google Scholar] [CrossRef]
- Lee, C.; Yang, W.; Parr, R.G. Results obtained with the correlation energy density functionals. Phys. Rev. B Condens. Matter Mater. Phys. 1988, 37, 785. [Google Scholar] [CrossRef]
- Takano, Y.; Houk, K.N. Benchmarking the conductor-like polarizable continuum model (CPCM) for aqueous solvation free energies of neutral and ionic organic molecules. J. Chem. Theory Comput. 2005, 1, 70–77. [Google Scholar] [CrossRef]
- Koopmans, T. Über die Zuordnung von Wellenfunktionen und Eigenwerten zu den einzelnen Elektronen eines Atoms. Physica 1934, 1, 104–113. [Google Scholar] [CrossRef]
- Todeschini, R.; Consonni, V.; Mauri, A.; Pavan, M. MobyDigs: Software for regression and classification models by genetic algorithms. Data Handl. Sci. Technol. 2003, 23, 141–167. [Google Scholar] [CrossRef]
- Dyer, D.H.; Lovell, S.; Thoden, J.B.; Holden, H.M.; Rayment, I.; Lan, Q. The structural determination of an insect sterol carrier protein-2 with a ligand-bound C16 fatty acid at 1.35-Å resolution. J. Biol. Chem. 2003, 278, 39085–39091. [Google Scholar] [CrossRef]
- Mori, A.; Lobo, N.F.; de Bruyn, B.; Severson, D.W. Molecular cloning and characterization of the complete acetylcholinesterase gene (Ace1) from the mosquito Aedes aegypti with implications for comparative genome analysis. Insect Biochem. Mol. Biol. 2007, 37, 667–674. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30, S162–S173. [Google Scholar] [CrossRef]
- Kiefer, F.; Arnold, K.; Kunzli, M.; Bordoli, L.; Schwede, T. The SWISSMODEL repository and associated resources. Nucleic Acids Res. 2009, 37, D387–D392. [Google Scholar] [CrossRef] [PubMed]
- Hörnberg, A.; Artursson, E.; Wärme, R.; Pang, Y.P.; Ekström, F. Crystal structures of oxime-bound fenamiphos-acetylcholinesterases: Reactivation involving flipping of the His447 ring to form a reactive Glu334–His447–oxime triad. Biochem. Pharmacol. 2010, 79, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B., III; De Bakker, P.I.; Word, J.M.; Prisant, M.G.; Richardson, D.C. Structure validation by Cα geometry: ϕ, ψ and Cβ deviation. Protein Struct. Funct. Genet. 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Kerwin, S.M. ChemBioOffice Ultra 2010 Suite. J. Am. Chem. Soc. 2010, 132, 2466–2467. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Larvicidal Activity | ||
|---|---|---|---|
| No | Name | LC50 (µg/mL) | Chi ** |
| 1 | 3-Carene | 49.3 (40.3–58.3) | 1.223 |
| 2 | Carvacrol | 8.8 (7.6–10.0) | 1.055 |
| 3 | Carvone epoxide | 217.5 (181.0–253.9) | 1.331 |
| 4 | Carvone | 119.1 (102.1–136.0) | 1.202 |
| 5 | p-Cymene | 86.8 (62.4–71.2) | 1.642 |
| 6 | Geranial | 78.3 (54.9–61.7) | 1.181 |
| 7 | Geraniol | 61.7 (40.6–42.8) | 0.203 |
| 8 | Lima | 1470.9 (1362.13–1579.6) | 3.568 |
| 9 | Hydrodihydrocarvone | 1628.2 (1516.9–1739.5) | 4.152 |
| 10 | Isopulegol | 297.6 (246.0–349.2) | 1.048 |
| 11 | 3-Isopropylpheno | 60.3 (35.7–44.9) | 1.969 |
| 12 | Limonene epoxide | 522.5 (459.2–585.8) | 1.722 |
| 13 | Limonene | 30.5 (27.8–33.1) | 0.326 |
| 14 | Menthol | 404.7(379.1–430.2) | 1.568 |
| 15 | Mentone | 508.9 (482.7–535.0) | 1.247 |
| 16 | Neoisopulegol | 554.6 (506.3–602.9) | 1.051 |
| 17 | Perillaldehyde epoxide | 715.1 (651.4–778.8) | 1.265 |
| 18 | Perillaldehyde | 115.8 (97.9–133.7 | 1.113 |
| 19 | Pulegone epoxide | 1116.2 (999.4–1233) | 0.214 |
| 20 | Pulegone | 188.1 (156.9–219.1) | 1.332 |
| 21 | Rotundifolone | 72.5 (64.4–80.6) | 0.975 |
| 22 | γ-Terpinene | 46.5 (41.1–51.9) | 0.856 |
| 23 | Trans-Dihydrocarvone | 361.3 (331.4–391.2) | 1.323 |
| 24 | Trans-Isopulegone | 538.8 (506.1–571.4) | 1.286 |
| 25 | Thymol | 10.3 (7.9–12.7) | 0.0966 |
| Tx | Temephos | 0.043 (0.041–0.045) | 1.668 |
| QSAR | QPAR | ||||||
|---|---|---|---|---|---|---|---|
| Statistic | Model 2 | Model 3 | Model 4 | Statistic | Model 2 | Model 3 | Model 4 |
| n | 25 | 25 | 25 | n | 25 | 25 | 25 |
| R2 | 88.81 | 87.64 | 87.42 | R2 | 92.87 | 91.4 | 91.28 |
| Q2 | 85.17 | 84.85 | 84.76 | Q2 | 88.57 | 87.44 | 87.29 |
| s | 33.01 | 32.89 | 32.14 | s | 48.01 | 52.77 | 52.91 |
| F | 82.5 | 82.3 | 81.8 | F | 94.1 | 92.8 | 92.8 |
| Descriptor | Contribution | Descriptor | Contribution | ||||
| Nr07 | −197.61 | WC | WC | AlogP | 497.5 | 362.2 | 317.04 |
| nCconj | 224.37 | WC | WC | Pol | −163.49 | −121.95 | −121.87 |
| nArOH | 965.09 | 936.49 | 934.22 | CENT | 8.02 | 4.92 | 4.91 |
| nROR | −254.81 | −317.19 | −379.23 | ELUMO | 61.38 | WC | WC |
| nCO | WC | −372.97 | WC | ƞ | WC | 369.38 | WC |
| nCrt | WC | 258.89 | 242.51 | AMR | WC | WC | −49.2 |
| Intercept | 136.8 | 142.72 | 168.05 | Intercept | 814.32 | 621.2 | 617.04 |
| Compounds | Structural Indices | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| No. | Name | Nr07 | nCrt | nOH | nROR | nR = Ct | nCIR | nCO | nArOH |
| 1 | 3-Carene | 1 | 2 | 0 | 0 | 1 | 3 | 0 | 0 |
| 2 | Carvacrol | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| 3 | (-)-Carvone epoxide | 1 | 2 | 0 | 1 | 1 | 3 | 1 | 0 |
| 4 | (-)-Carvone | 0 | 1 | 0 | 0 | 2 | 1 | 1 | 0 |
| 5 | p-Cymene | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| 6 | Geranial | 0 | 1 | 0 | 0 | 2 | 0 | 1 | 0 |
| 7 | Geraniol | 0 | 1 | 1 | 0 | 2 | 0 | 0 | 0 |
| 8 | Hydrocarvone | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 |
| 9 | Hydrodihydrocarvone | 0 | 2 | 1 | 0 | 0 | 1 | 1 | 0 |
| 10 | Isopulegol | 0 | 2 | 1 | 0 | 1 | 1 | 0 | 0 |
| 11 | 3-Isopropylpheno | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| 12 | (-)-Limonene epoxide | 1 | 2 | 0 | 1 | 1 | 3 | 0 | 0 |
| 13 | (+)-Limonene | 0 | 1 | 0 | 0 | 2 | 1 | 0 | 0 |
| 14 | Menthol | 0 | 2 | 1 | 0 | 0 | 1 | 0 | 0 |
| 15 | Mentone | 0 | 2 | 0 | 0 | 0 | 1 | 1 | 0 |
| 16 | Neoisopulegol | 0 | 2 | 1 | 0 | 1 | 1 | 0 | 0 |
| 17 | Perillaldehyde epoxide | 1 | 2 | 0 | 1 | 1 | 3 | 0 | 0 |
| 18 | (-)-Perillaldehyde | 0 | 1 | 0 | 0 | 2 | 1 | 0 | 0 |
| 19 | Pulegone epoxide | 0 | 3 | 0 | 1 | 0 | 2 | 1 | 0 |
| 20 | (+)-Pulegone | 0 | 1 | 0 | 0 | 2 | 1 | 1 | 0 |
| 21 | Rotundifolone | 1 | 1 | 0 | 1 | 2 | 3 | 1 | 0 |
| 22 | γ-Terpinene | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 0 |
| 23 | Trans-Dihydrocarvone | 0 | 2 | 0 | 0 | 1 | 1 | 1 | 0 |
| 24 | Trans-Isopulegone | 0 | 2 | 0 | 0 | 0 | 1 | 1 | 0 |
| 25 | Thymol | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 |
| Compounds | Molecular Indices | Topological Indices | QM Indices | |||||
|---|---|---|---|---|---|---|---|---|
| No. | Name | AlogP | AMR | Qtot | Pol | CENT | I | ƞ |
| 1 | 3-Carene | 2.873 | 44.722 | 1.861 | 11 | 46 | 8.973 | 6.551 |
| 2 | Carvacrol | 3.243 | 46.984 | 2.999 | 14 | 73 | 8.351 | 6.232 |
| 3 | Carvone epoxide | 1.262 | 45.779 | 2.581 | 17 | 78 | 9.771 | 6.598 |
| 4 | Carvone | 2.361 | 47.174 | 2.495 | 14 | 73 | 9.308 | 6.053 |
| 5 | p-Cymene | 3.51 | 45.29 | 2.631 | 11 | 60 | 8.542 | 6.356 |
| 6 | Geranial | 3.19 | 50.199 | 2.566 | 9 | 102 | 9.05 | 5.701 |
| 7 | Geraniol | 2.934 | 51.182 | 3.367 | 9 | 102 | 8.859 | 6.792 |
| 8 | Hydrocarvone | 1.274 | 49.154 | 3.191 | 16 | 102 | 7.753 | 7.136 |
| 9 | Hydrodihydrocarvone | 1.313 | 48.278 | 3.223 | 16 | 102 | 10.387 | 7.402 |
| 10 | Isopulegol | 2.583 | 47.222 | 2.463 | 14 | 80 | 9.461 | 7.278 |
| 11 | 3-Isopropylpheno | 2.757 | 41.943 | 2.12 | 11 | 80 | 8.523 | 6.254 |
| 12 | Limonene epoxide | 2.269 | 45.239 | 2.033 | 13 | 65 | 9.274 | 7.017 |
| 13 | Limonene | 3.503 | 46.48 | 1.903 | 11 | 60 | 8.746 | 6.824 |
| 14 | Menthol | 2.779 | 47.445 | 3.545 | 14 | 80 | 10.918 | 8.218 |
| 15 | Mentone | 2.597 | 46.52 | 2.575 | 14 | 80 | 10.712 | 7.066 |
| 16 | Neoisopulegol | 2.583 | 47.222 | 2.54 | 14 | 80 | 9.605 | 7.219 |
| 17 | Perillaldehyde epoxide | 1.637 | 45.801 | 2.522 | 16 | 74 | 9.571 | 6.217 |
| 18 | Perillaldehyde | 2.668 | 47.272 | 2.461 | 13 | 71 | 9.442 | 6.019 |
| 19 | Pulegone epoxide | 1.411 | 46.091 | 2.579 | 17 | 104 | 10.826 | 7.238 |
| 20 | Pulegone | 2.752 | 47.129 | 2.448 | 14 | 80 | 9.146 | 6.328 |
| 21 | Rotundifolone | 1.824 | 46.637 | 2.472 | 17 | 86 | 9.392 | 6.095 |
| 22 | γ-Terpinene | 3.449 | 47.553 | 1.784 | 11 | 60 | 7.641 | 5.509 |
| 23 | Trans-Dihydrocarvone | 2.401 | 46.298 | 2.574 | 14 | 73 | 9.491 | 6.656 |
| 24 | Trans-Isopulegone | 2.597 | 46.52 | 2.571 | 14 | 80 | 10.474 | 7.217 |
| 25 | Thymol | 3.243 | 46.98 | 3.045 | 14 | 80 | 8.325 | 6.235 |
| Compounds | Chemical Reactivity Descriptors | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| No. | Name | EHOMO | ELUMO | A | χ | µ | S | GAPE | m |
| 1 | 3-Carene | −8.97 | 4.13 | −4.13 | −2.42 | 2.42 | 0.15 | 13.1 | 0.17 |
| 2 | Carvacrol | − 8.35 | 4.11 | − 4.11 | − 2.11 | 2.11 | 0.16 | 12.46 | 1.67 |
| 3 | Carvone epoxide | −9.77 | 3.42 | −3.42 | −3.17 | 3.17 | 0.15 | 13.19 | 5.59 |
| 4 | Carvone | −9.3 | 2.79 | −2.79 | −3.25 | 3.25 | 0.16 | 12.1 | 3.98 |
| 5 | p-Cymene | − 8.54 | 4.17 | − 4.17 | − 2.18 | 2.18 | 0.15 | 12.71 | 0.05 |
| 6 | Geranial | − 9.05 | 2.3 | − 2.34 | − 3.36 | 3.36 | 0.18 | 11.39 | 4.73 |
| 7 | Geraniol | − 8.85 | 4.72 | − 4.72 | − 2.06 | 2.06 | 0.14 | 13.58 | 2.4 |
| 8 | Hydrocarvone | −7.75 | 6.51 | −6.51 | −0.61 | 0.61 | 0.14 | 14.27 | 3.77 |
| 9 | Hydrodihydrocarvone | −10.38 | 4.41 | −4.41 | −2.98 | 2.98 | 0.13 | 14.8 | 2.52 |
| 10 | Isopulegol | −9.46 | 5.09 | −5.09 | −2.18 | 2.18 | 0.13 | 14.55 | 3.75 |
| 11 | 3-Isopropylpheno | − 8.52 | 3.98 | − 3.98 | − 2.26 | 2.26 | 0.15 | 12.5 | 1.39 |
| 12 | Limonene epoxide | −9.27 | 4.75 | −4.75 | −2.25 | 2.25 | 0.14 | 14.03 | 3.13 |
| 13 | Limonene | −8.74 | 4.9 | −4.9 | −1.92 | 1.92 | 0.14 | 13.64 | 0.58 |
| 14 | Menthol | −10.91 | 5.51 | −5.51 | 2.7 | −2.7 | 0.12 | 15.4 | 2.04 |
| 15 | Mentone | −10.71 | 3.41 | −3.41 | −3.64 | 3.64 | 0.14 | 14.13 | 3.63 |
| 16 | Neoisopulegol | −9.6 | 4.83 | −4.83 | −2.38 | 2.38 | 0.13 | 14.43 | 2.32 |
| 17 | Perillaldehyde epoxide | −9.57 | 2.86 | −2.86 | −3.35 | 3.35 | 0.16 | 12.43 | 3.17 |
| 18 | Perillaldehyde | −9.44 | 2.59 | −2.59 | −3.42 | 3.42 | 0.16 | 12.03 | 3.63 |
| 19 | Pulegone epoxide | −10.82 | 3.65 | −3.65 | −3.58 | 3.58 | 0.13 | 14.47 | 5.68 |
| 20 | Pulegone | −9.14 | 3.51 | −3.51 | −2.81 | 2.81 | 0.15 | 12.65 | 3.55 |
| 21 | Rotundifolone | −9.39 | 2.79 | −2.79 | −3.29 | 3.29 | 0.16 | 12.19 | 3.74 |
| 22 | γ-Terpinene | −7.64 | 3.37 | −3.37 | −2.13 | 2.13 | 0.18 | 11.01 | 0.64 |
| 23 | Trans-Dihydrocarvone | −9.49 | 3.82 | −3.82 | −2.83 | 2.83 | 0.15 | 13.31 | 3.59 |
| 24 | Trans-Isopulegone | −10.47 | 3.96 | −3.96 | −3.25 | 3.25 | 0.13 | 14.43 | 3.5 |
| 25 | Thymol | − 8.32 | 4.14 | − 4.14 | − 2.09 | 2.09 | 0.16 | 12.47 | 1.76 |
| No. | Molecules | Ache | Amino Acid Residues Involved in the Interaction |
|---|---|---|---|
| ΔG (Kcal) | |||
| 9 | Hydrodihydrocarvone | −6.78 | E220 *, N226, V228, H260, A401 *, I413, C414 |
| 8 | Hydrocarvone | −6.68 | L218 *, N226, V227, V228, T254, D259 |
| 25 | Thymol | −6.61 | V196, N226, V228, N399, P400, C414 |
| 2 | Carvacrol | −6.66 | V196, L218, N226, V228, N399, I413 |
| 4 | Carvone | −6.63 | N226, V228, C414, Y461, L462, E485, P488, L486 |
| 20 | Pulegone | −6.61 | N226, V228, I413, C414 *, F457, I458, Y461, L462 |
| 25 | Menthol | −6.61 | L159, N226, V227, V228, A253, D259 *, H260 |
| 15 | Mentone | −6.59 | N226, V227, V271, V228, T254, L255, D256, H260 |
| 16 | Neoisopulegol | −6.59 | V227, I413, C414 *, Y461, L462, F457, I458, E485, L486, V522 |
| 10 | Isopulegol | −6.58 | I413, C414 *, Y461, L462, F457, I458, E485, L486, V522 |
| 23 | Trans-Dihydrocarvone | −6.58 | C414, Y461, L462, E485, L846 |
| 13 | Limonene | −6.55 | C414, Y461, L462, E485, L846, P488, V522 |
| 18 | Perillaldehyde | −6.54 | N226, V227, V271, V228, T254, L255, D256, H260 |
| 22 | γ-Terpinene | −6.54 | N226, V227, V228, I413, C414, Y461 |
| 24 | Trans-Isopulegone | −6.54 | I413, C414 *, Y461, L462, F457, E485, V522 |
| 21 | Rotundifolone | −6.52 | I413, C414 *, Y461, L462, F457, |
| 7 | Geraniol | −6.51 | C414, Y461, L462, E485, P488, L486, V522 |
| 19 | Pulegone epoxide | −6.51 | C414, Y461, L462, E485, L846, P488, V522 |
| 17 | Perillaldehyde epoxide | −6.5 | N226, V227, V271, V228, T254, L255, D256, H260 |
| 6 | Geranial | −6.47 | C414, Y461, L462, E485, P488, L486 |
| 1 | 3-Carene | −6.44 | L159, V228, Y258, D259, H260, V271, S273 |
| 12 | Limonene epoxide | −6.4 | N226, T254, L255, D256, H260, C414 |
| 3 | Carvone epoxide | −6.22 | C414, Y461, L462, E485, L486 |
| 5 | p-Cymene | −6.12 | N226, T254, L255, D256, H260, P400, C414 |
| 11 | 3-Isopropylphenol | −6.12 | N226, A253 *, T254, D259, H260, S273, V271, P400 |
| No. | Molecules | SCP-2 | Amino Acid Residues Involved in the Interaction |
|---|---|---|---|
| ΔG (Kcal) | |||
| 2 | Carvacrol | −6.88 | I19, N23, R24, Q25, V26 *, L48, L102, F105 |
| 25 | Thymol | −6.85 | I19, D20, N23, R24, Q25, V26, L48, F105 |
| 13 | Limonene | −6.83 | I19, R24, Q25, V26, L48, L102, F105 |
| 22 | γ-Terpinene | −6.81 | I19, N23, R24, Q25, V26, L48, L102, F105 |
| 14 | Menthol | −6.69 | I19, R24 *, Q25, V26, Q25 *, F105 |
| 4 | Carvone | −6.62 | R15 *, I19, D20, R24, N23, Q25, V26, F105 |
| 15 | Mentone | −6.51 | R15 *, I19, R24, N23, Q25, V26 |
| 20 | Pulegone | −6.51 | R15, I19, N23, R24 *, Q25, V26 |
| 24 | Trans-Isopulegone | −6.44 | R15 *, I19, D20, R24, N23, Q25, V26, L48 |
| 16 | Neoisopulegol | −6.34 | I19, N23, R24 *, Q25, V26, F105 |
| 21 | Rotundifolone | −6.33 | I19, R24 *, Q25, V26, L48, F105 |
| 10 | Isopulegol | −6.26 | I19, D20, R24, V26 *, L48, F105 |
| 1 | 3-Carene | −6.17 | I19, N23, R24, Q25, V26, F105 |
| 11 | 3-Isopropylphenol | −6.05 | D20, N23, R24, Q25, V26 |
| 12 | Limonene epoxide | −6.03 | I19, D20, R24, N23, Q25, V26, |
| 23 | Trans-Dihydrocarvone | −5.97 | R15 *, I19, D20, R24, N23, Q25, V26 *, F105 |
| 18 | Perillaldehyde | −5.95 | R15, L16, I19, N23, R24 *, Q25, V26, F105 |
| 7 | Geraniol | −5.88 | I19, D20, N23, R24, Q25, V26, L48, L102, F105 |
| 6 | Geranial | −5.76 | I19, D20, N23, R24, Q25, V26 |
| 17 | Perillaldehyde epoxide | −5.57 | R15 *, L16, I19, N23, R24, Q25, V26 |
| 19 | Pulegone epoxide | −5.51 | R15 *, I19, R24, V26 |
| 5 | p-Cymene | −5.32 | D20, N23, R24, Q25, V26 |
| 3 | Carvone epoxide | −5.23 | R15, I19, D20, R24, N23, Q25, V26 |
| 8 | Hydrocarvone | −5.22 | I19, D20 *, R24, N23, Q25, V26, F105 |
| 9 | Hydrodihydrocarvone | −5.21 | R15 *, I19, D20, R24, N23, Q25, V26 *, F105 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Castillo, A.U.; Rodríguez-Valdez, L.M.; Correa-Basurto, J.; Nogueda-Torres, B.; Andrade-Ochoa, S.; Nevárez-Moorillón, G.V. Terpenic Constituents of Essential Oils with Larvicidal Activity against Aedes Aegypti: A QSAR and Docking Molecular Study. Molecules 2023, 28, 2454. https://doi.org/10.3390/molecules28062454
Cruz-Castillo AU, Rodríguez-Valdez LM, Correa-Basurto J, Nogueda-Torres B, Andrade-Ochoa S, Nevárez-Moorillón GV. Terpenic Constituents of Essential Oils with Larvicidal Activity against Aedes Aegypti: A QSAR and Docking Molecular Study. Molecules. 2023; 28(6):2454. https://doi.org/10.3390/molecules28062454
Chicago/Turabian StyleCruz-Castillo, Adrián Ulises, Luz María Rodríguez-Valdez, José Correa-Basurto, Benjamín Nogueda-Torres, Sergio Andrade-Ochoa, and Guadalupe Virginia Nevárez-Moorillón. 2023. "Terpenic Constituents of Essential Oils with Larvicidal Activity against Aedes Aegypti: A QSAR and Docking Molecular Study" Molecules 28, no. 6: 2454. https://doi.org/10.3390/molecules28062454
APA StyleCruz-Castillo, A. U., Rodríguez-Valdez, L. M., Correa-Basurto, J., Nogueda-Torres, B., Andrade-Ochoa, S., & Nevárez-Moorillón, G. V. (2023). Terpenic Constituents of Essential Oils with Larvicidal Activity against Aedes Aegypti: A QSAR and Docking Molecular Study. Molecules, 28(6), 2454. https://doi.org/10.3390/molecules28062454