
Anti-Inflammatory Activity of Panax notoginseng Flower Saponins Quantified Using LC/MS/MS

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Extraction and Purification of PNFS
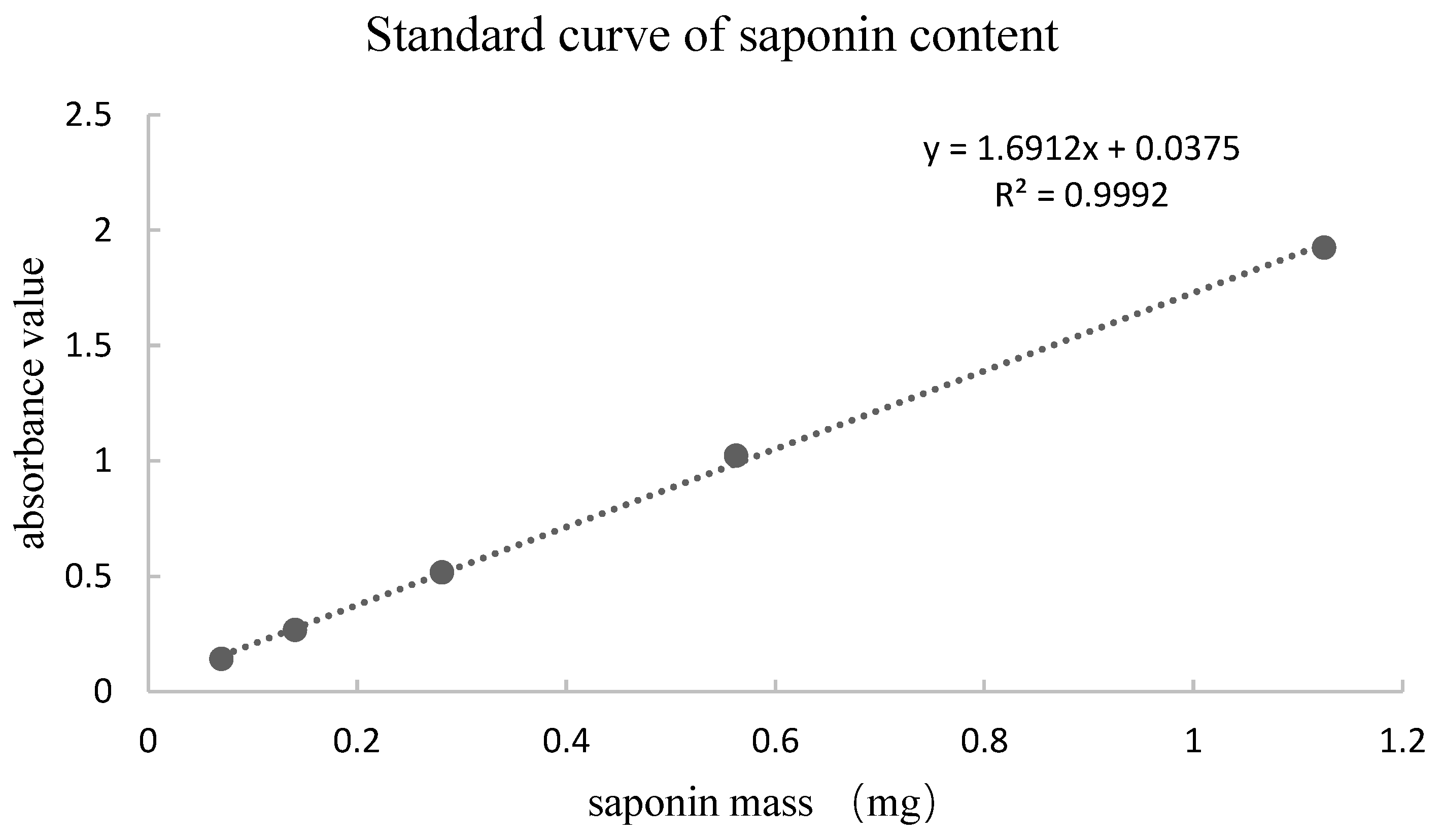
2.3. Measurement of Total Saponin Content
2.4. Measurement of COX-2 Overproduction
2.5. Cell Culture
2.6. Cell Viability
2.7. Measurement of PGE-2, IL-1β, TNF-α, and LL-37 Overproduction by ELISA
2.8. Western Blotting Analysis
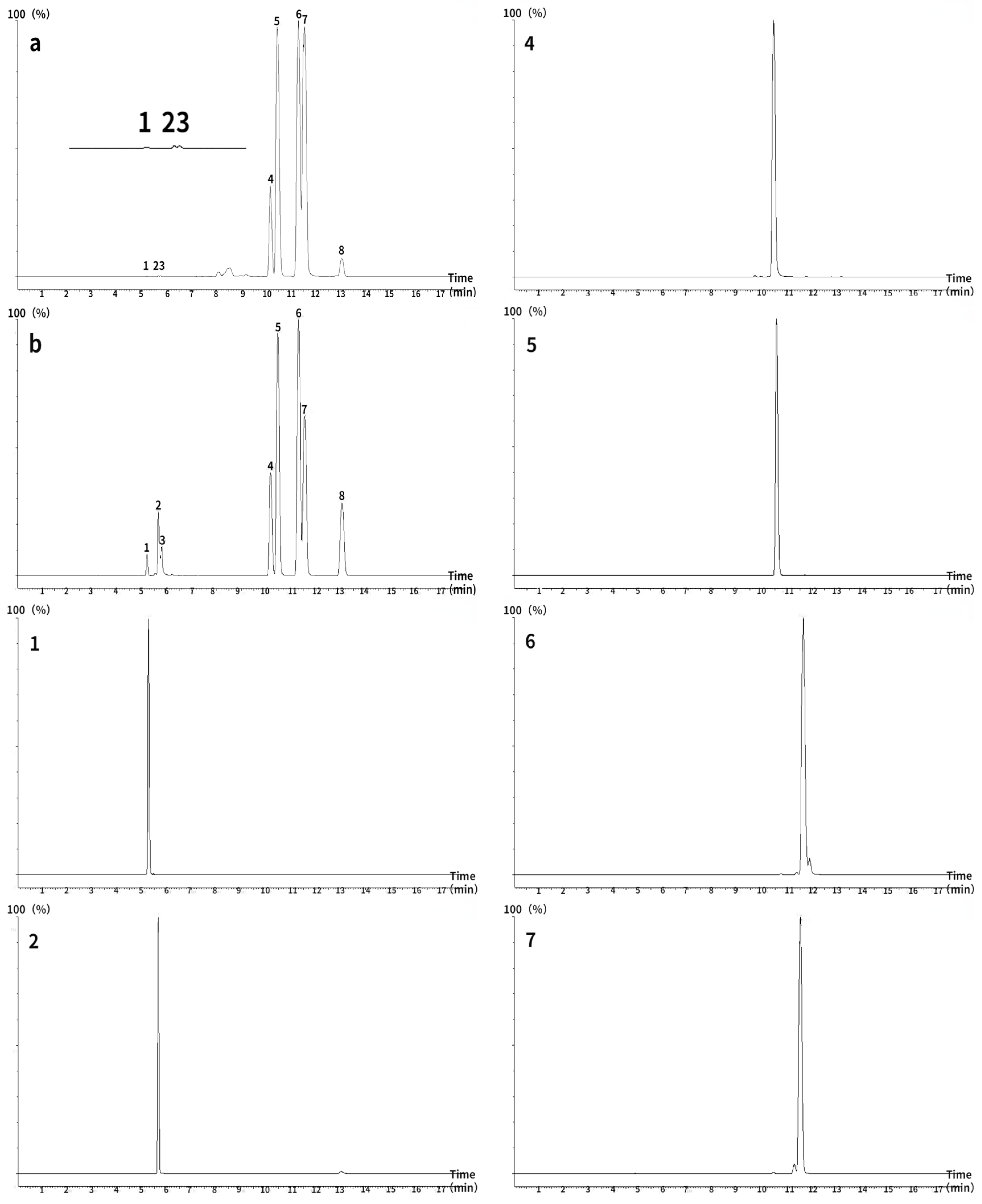
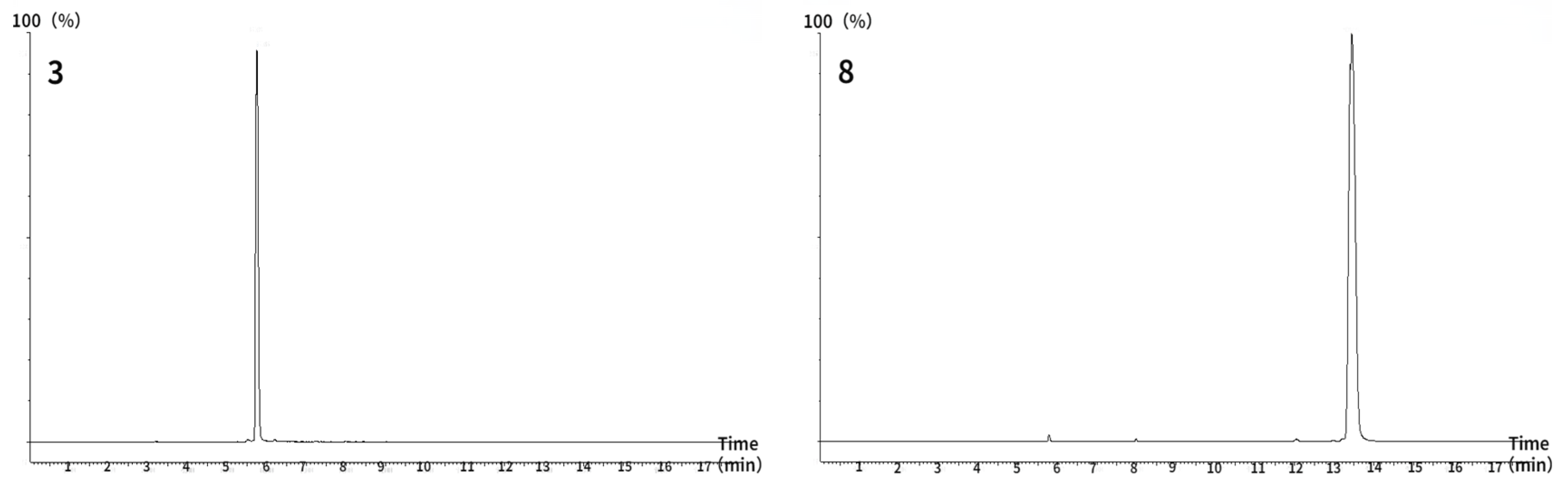
2.9. Quantitation of Saponins in PNFS by UHPLC/MS/MS
2.10. Statistical Analysis
3. Results and Discussion
3.1. Total Saponin Content in 75% Ethanol Extract and Purified Extract (PNFS)
3.2. Effect of PNFS on COX-2
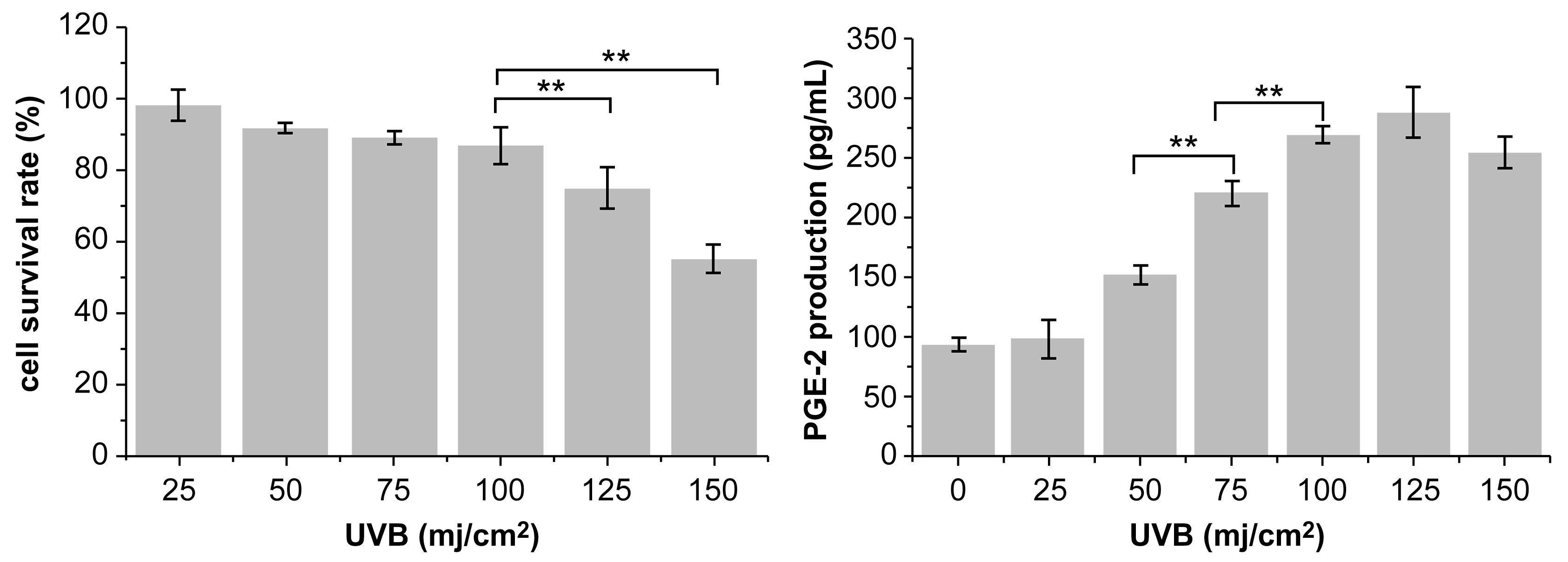
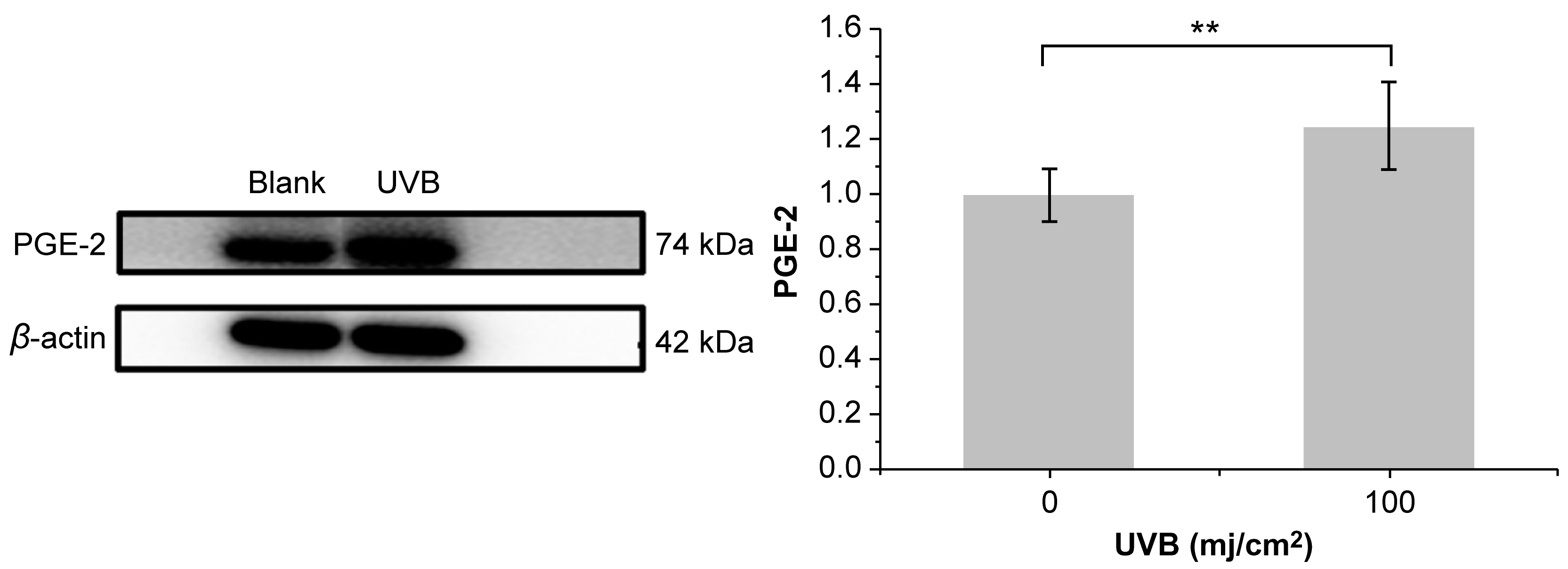
3.3. Cell Viability and Development of the UVB-Irradiation-Induced Inflammation Model
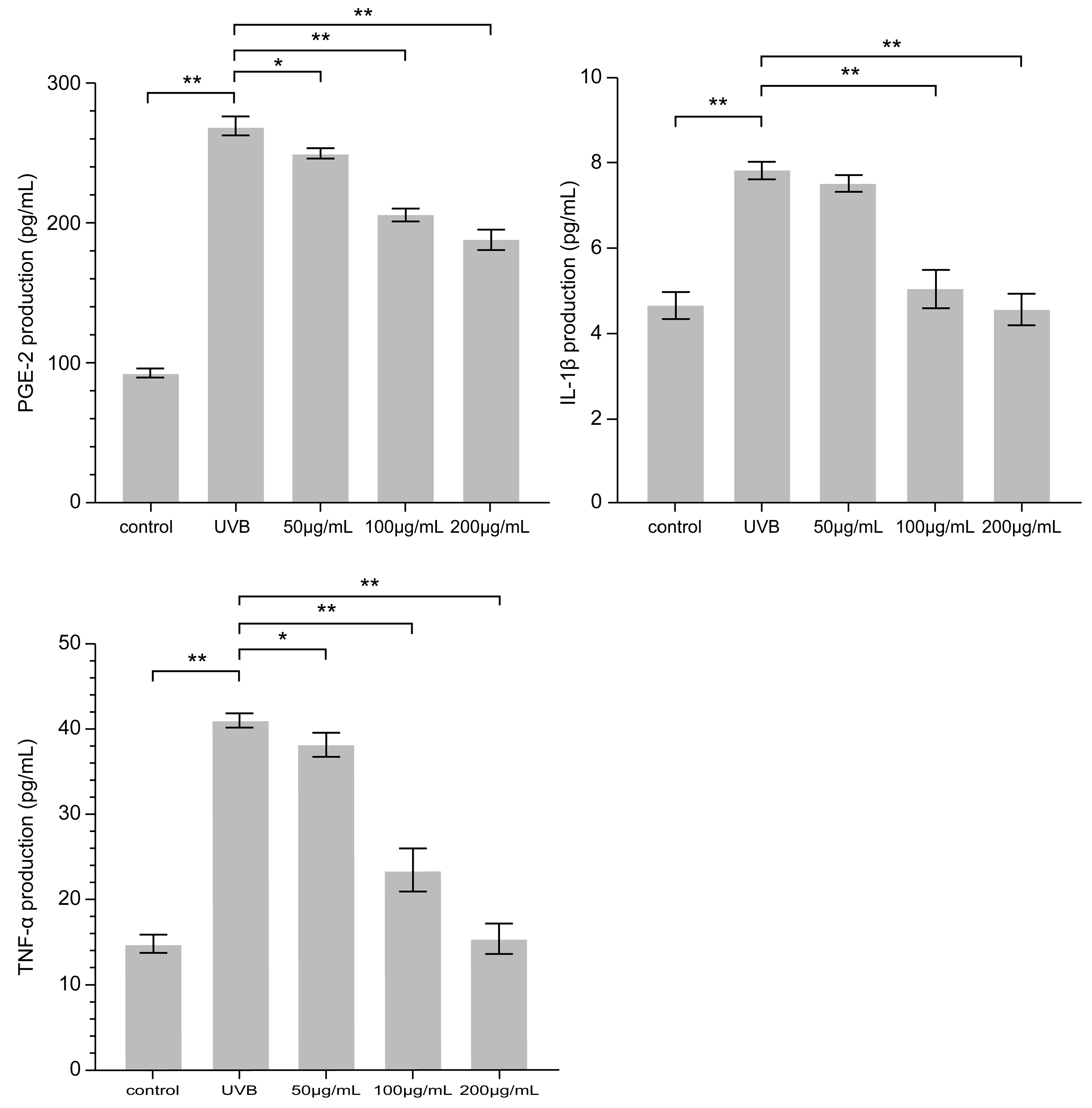
3.4. Effect of PNFS on PGE-2, IL-1β, and TNF-α Determined by ELISA
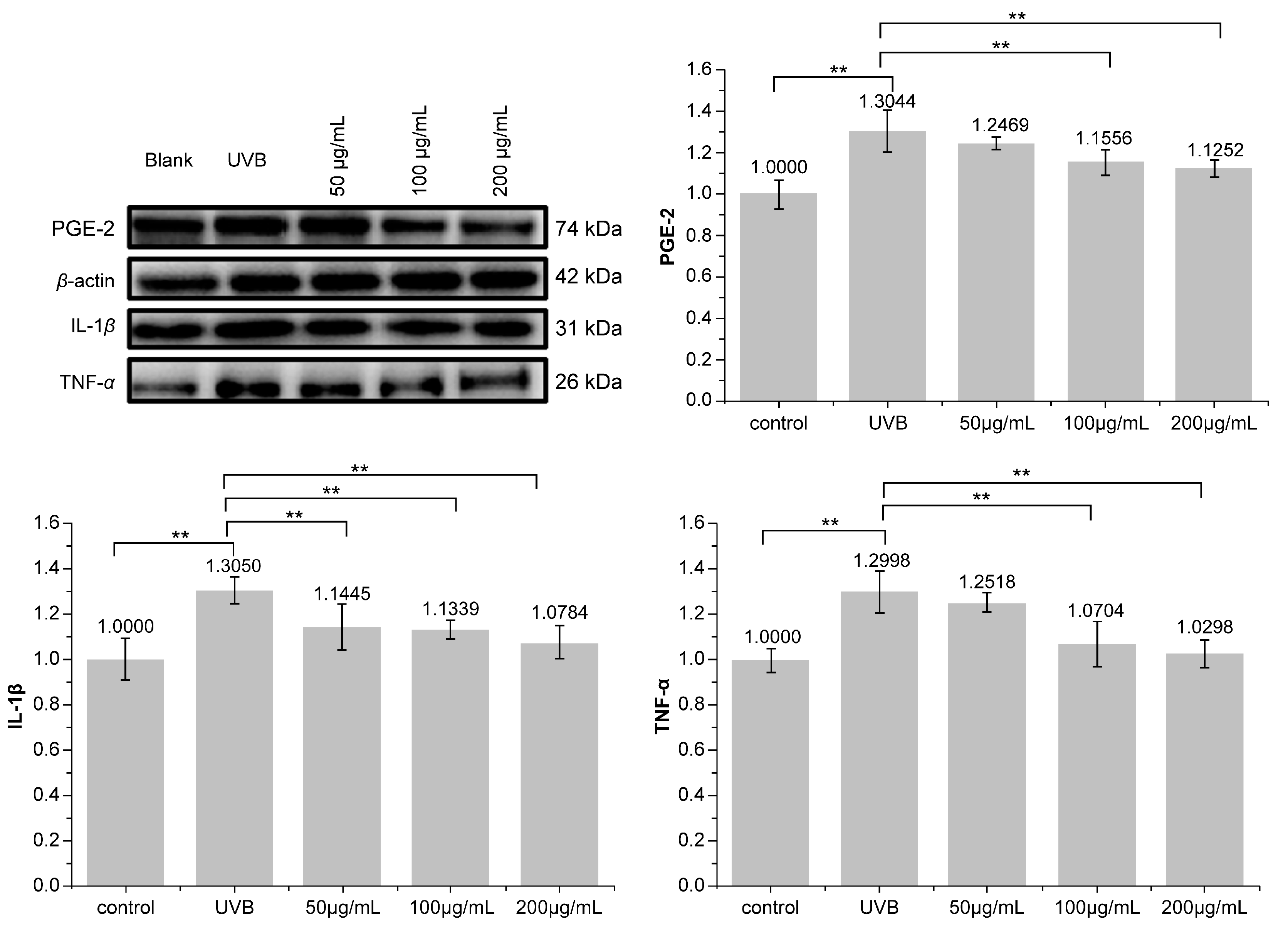
3.5. Effect of PNFS on PGE-2, IL-1β, and TNF-α Determined by Western Blotting
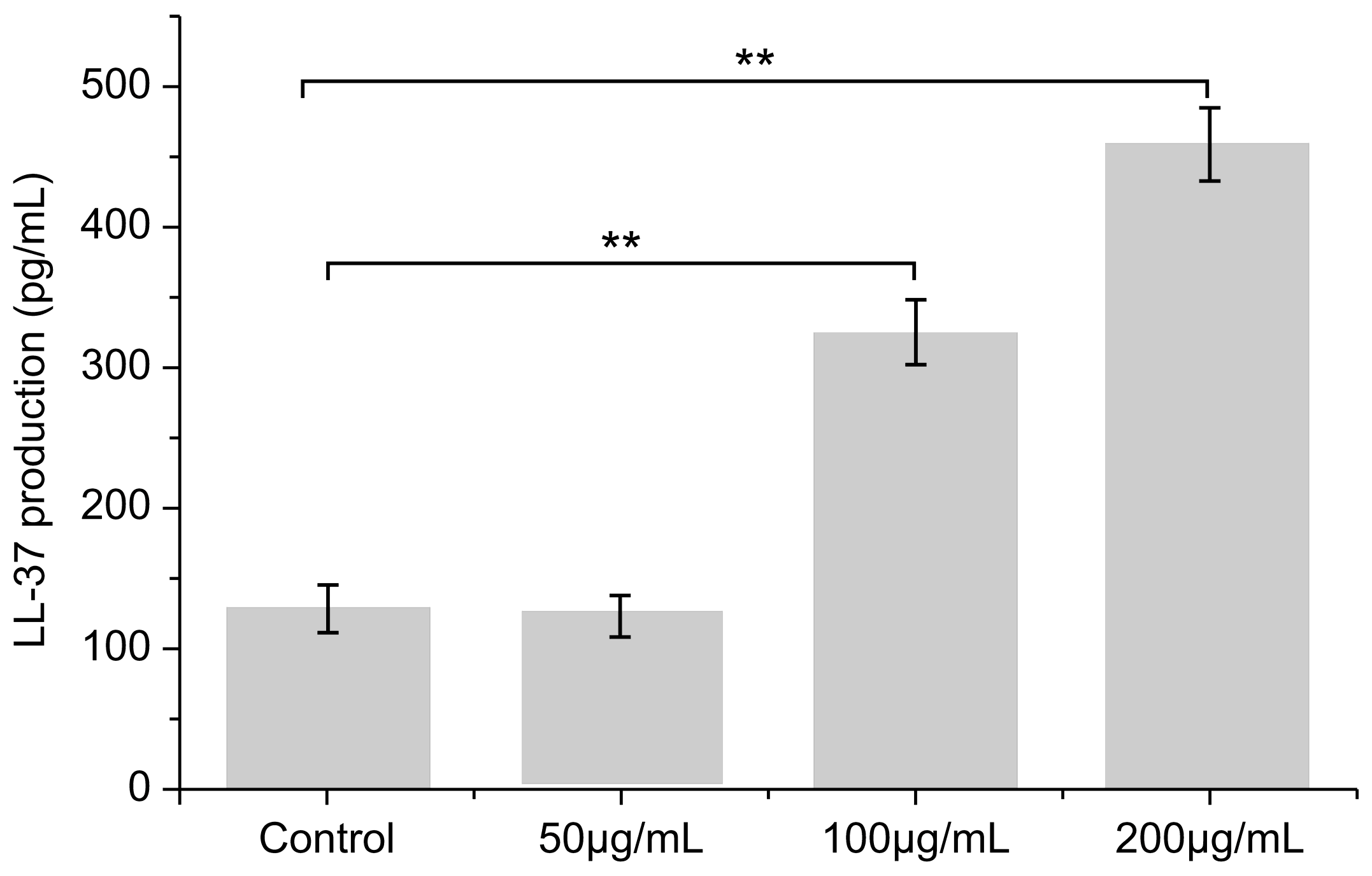
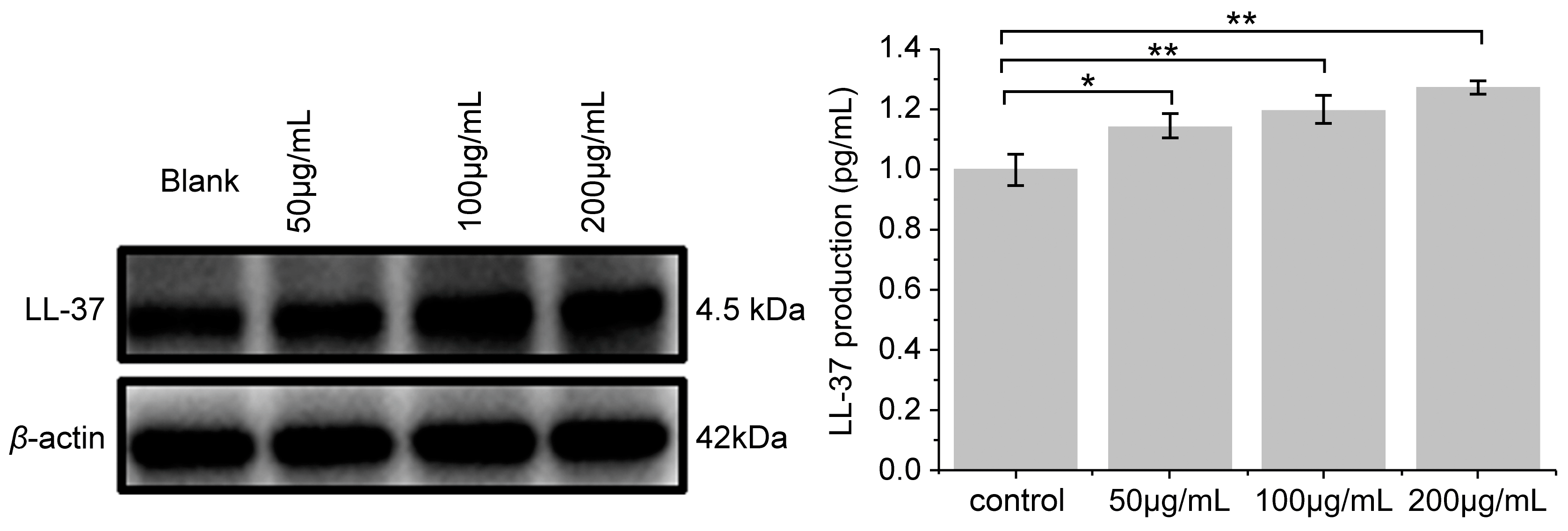
3.6. Effect of PNFS on LL-37 Expression
3.7. Major Saponin Contents in PNFS
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Y.C.; Zhang, T.J.; Guo, H.B.; Xu, J.; Lin, J.; Zhang, H.B.; Han, Y.Q.; Wang, D.Q.; Li, C.Y.; Liu, C.X. Research progress and predictive analysis of quality markers of Panax notoginseng. Chin. Herbal Med. 2021, 9, 2733–2745. [Google Scholar] [CrossRef]
- Gan, C.M.; Chen, H.Y.; Gong, Y.Q. Notoginseng flower the research progress of chemical composition. J. Tradit. Chin. Med. Yunnan 2019, 40, 73–75. [Google Scholar] [CrossRef]
- Peng, X.X.; Zhang, S.H.; Wang, X.L.; Ye, T.; Li, H.; Yan, X.; Wei, X.; Wu, Z.; Hu, J.; Zou, C.; et al. Panax Notoginseng flower saponins (PNFS) inhibit LPS-stimulated NO overproduction and iNOS gene overexpression via the suppression of TLR4-mediated MAPK/NF-kappa B signaling pathways in RAW264. 7 macrophages. Chin Med. 2015, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.F.; Dong, L.L.; Yang, J.; Zhang, L.J.; Xu, J.; Yang, F.; Cheng, R.Y.; Xu, R.; Chen, S.L. Integrated metabolomic and transcriptomic analyses revealed the distribution of saponins in. Acta Pharm. Sin. B. 2018, 8, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chu, S.F.; Zhang, Z.; Chen, N.H. Hepataprotective effects of ginsenoside Rg1—A review. J. Ethnopharmacol. 2017, 206, 178–183. [Google Scholar] [CrossRef]
- Shin, D.; Moon, H.W.; Oh, Y.; Kim, K.; Kim, D.D.; Lim, C.J. Defensive Properties of Ginsenoside Re against UV-B-Induced Oxidative Stress through Up-Regulating Glutathione and Superoxide Dismutase in HaCaT Keratinocytes. Iran. J. Pharm. Res. 2018, 17, 249–260. [Google Scholar]
- Hou, J.G.; Xue, J.J.; Lee, M.; Sung, C.K. Ginsenoside Rd as a potential neuroprotective agent prevents trimethyltin injury. Biomed. Rep. 2017, 6, 435–440. [Google Scholar] [CrossRef]
- Gong, L.; Yin, J.; Zhang, Y.; Huang, R.; Lou, Y.; Jiang, H.; Sun, L.; Jia, J.; Zeng, X. Neuroprotective Mechanisms of Ginsenoside Rb1 in Central Nervous System Diseases. Front. Pharmacol. 2022, 13, 914352. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Zheng, H.; Zhang, R.; Piao, X.; Hu, J.; Zhu, Y.; Wang, Y. Immunomodulatory Effect of Ginsenoside Rb2 Against Cyclophosphamide-Induced Immunosuppression in Mice. Front. Pharmacol. 2022, 13, 927087. [Google Scholar] [CrossRef]
- Yu, T.; Yang, Y.Y.; Kwak, Y.S.; Song, G.G.; Kim, M.Y.; Rhee, M.H.; Cho, J.Y. Ginsenoside Rc from exerts anti-inflammatory activity by targeting TANK-binding kinase 1/interferon regulatory factor-3 and p38/ATF-2. J. Ginseng Res. 2017, 41, 127–133. [Google Scholar] [CrossRef]
- Li, C.Y. Analysis of chemical components and quality control methods of traditional medicinal parts of Panax notoginseng. Sichuan Trad. Chin. Med. 2018, 3, 75–78. [Google Scholar]
- Liu, M.H.; Yang, B.R.; Cheung, W.F.; Yang, K.Y.; Zhou, H.F.; Kwok, J.S.M.; Liu, G.C.; Li, X.F.; Zhong, S.L.; Lee, S.M.Y.; et al. Transcriptome analysis of leaves, roots and flowers of Panax notoginseng identifies genes involved in ginsenoside and alkaloid biosynthesis. BMC Genom. 2015, 16, 265. [Google Scholar] [CrossRef] [PubMed]
- Ivan, A.L.; Campanini, M.Z.; Martinez, R.M.; Ferreira, V.S.; Steffen, V.S.; Vicentini, F.T.; Vilela, F.M.; Martins, F.S.; Zarpelon, A.C.; Cunha, T.M.; et al. Pyrrolidine dithiocarbamate inhibits UVB-induced skin inflammation and oxidative stress in hairless mice and exhibits antioxidant activity in vitro. J. Photochem. Photobiol. B. 2014, 138, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.-H.; Wang, J.-Y.; Chiu, L.-Y.; Huang, Y.-T.; Hung, C.-F.; Wu, N.-L. Spleen tyrosine kinase regulates keratinocyte inflammasome activation and skin inflammation induced by UVB irradiation. Free. Radic. Biol. Med. 2022, 180, 121–133. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, R.; Chen, H.; Zhang, X.; Zhang, Y.; Liu, H.; Li, C.; Chen, Y.; Zeng, Q.; Huang, G. Injectable Supramolecular Hybrid Hydrogel Delivers IL-1β-Stimulated Exosomes to Target Neuroinflammation. ACS Appl. Mater. Interfaces 2023, 15, 5. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, H.; Zhang, Y.; Wang, D.; Zhao, D.; Zhang, J.; Li, M.; Wang, C. Reparative Effects of Dandelion Fermentation Broth on UVB-Induced Skin Inflammation. Clin. Cosmet. Investig. Dermatol. 2022, 15, 471–482. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, J.; Li, M.Y.; Ma, X.J.; Dong, X.; Sun, X.P.; Zhao, D.M.; Zhou, X.M.; Xu, L.H. Research progress of antibacterial peptide LL37 against pathogen infection. Chin. J. Vet. Med. 2020, 2, 63–66. [Google Scholar]
- López-Camarillo, C.; Aréchaga Ocampo, E.; López Casamichana, M.; Pérez-Plasencia, C.; Álvarez-Sánchez, E.; Marchat, L.A. Protein kinases and transcription factors activation in response to UV-radiation of skin: Implications for carcinogenesis. Int. J. Mol. Sci. 2012, 13, 142–172. [Google Scholar] [CrossRef]
- Huiting, P.; Duo-Sheng, L.; Jiao, G. Chemical constituents analysis of Panax notoginseng and its anti-inflammatory mechanism in network pharmacology. Chin. Herb. Med. 2020, 51, 5538–5547. [Google Scholar] [CrossRef]
- Park, S.-H.; Seo, W.; Eun, H.S.; Kim, S.Y.; Jo, E.; Kim, M.-H.; Choi, W.-M.; Lee, J.-H.; Shim, Y.-R.; Cui, C.-H.; et al. Protective effects of ginsenoside F2 on 12-O-tetradecanoylphorbol-13-acetate-induced skin inflammation in mice. Biochem. Biophys. Res. Commun. 2016, 478, 1713–1719. [Google Scholar] [CrossRef]
- Wei, D.; Yang, H.; Zhang, Y.; Zhang, X.; Wang, J.; Wu, X.; Chang, J. Extraction and content determination of total saponins from Panax notoginseng. J. Nankai Univ. 2021, 54, 54–59. [Google Scholar]
- Gui, S.Y.; Zhou, Y.Q. Determination of total ginsenosides in ginseng by colorimetric method. J. Anhui Uni. Trad. Chin. Med. 2003, 4, 51–52. [Google Scholar]
- Pillai-Kastoori, L.; Schutz-Geschwender, A.R.; Harford, J.A. A systematic approach to quantitative Western blot analysis. Anal Biochem. 2020, 593, 113608. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.W.; Cheng, Y.C.; Hung, Y.C.; Lee, C.H.; Fang, J.Y.; Li, W.T.; Wu, Y.R.; Pan, T.L. Red raspberry extract protects the skin against UVB-induced damage with antioxidative and anti-inflammatory properties. Oxid. Med. Cell. Longev. 2019, 2019, 9529676. [Google Scholar] [CrossRef]
- Kun, H. An Enzyme Called Cyclooxygenase UVB Induced the Role of Melanocyte Dendritic Formation and Mechanism Research. Master’s Thesis, The Fourth Military Medical University, Xi’an, China, 2012. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD201301&filename=1012039124.nh (accessed on 26 February 2023).
- Jung, H.W.; Seo, U.K.; Kim, J.H.; Leem, K.H.; Park, Y.K. Flower extract of Panax notoginseng attenuates lipopolysaccharide-induced inflammatory response via blocking of NF-kappaB signaling pathway in murine macrophages. J. Ethnopharmacol. 2009, 122, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Cirone, K.M.; Lahiri, P.; Holani, R.; Tan, Y.L.; Arrazuria, R.; De Buck, J.; Barkema, H.B.; Cobo, E.R. Synthetic cathelicidin LL-37 reduces Mycobacterium avium subsp. paratuberculosis internalization and pro-inflammatory cytokines in macrophages. Cell. Tissue Res. 2020, 379, 207–217. [Google Scholar] [CrossRef]
- Chen, N. Study on the Regulatory Effects of microRNA-203a-5P on IL-6, TNF-α and IL-1β in AD. Ph.D. Thesis, University of Chinese medicine in Guangxi, Guangxi, China, 2019. [Google Scholar]
- Choi, S.W. Epidermis proliferative effect of the Panax ginseng ginsenoside Rb2. Arch. Pharmacol. Res. 2002, 25, 71–76. [Google Scholar]
- Oh, S.J.; Oh, J.; Ryu, I.W.; Kim, K.H.; Lim, C.J. Protective properties of ginsenoside Rb3 against UV-B radiation-induced oxidative stress in HaCaT keratinocytes. Biosci. Biotechnol. Biochem. 2016, 80, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Lim, H.W.; Park, K.H.; Huang, Y.H.; Yoon, J.Y.; Kim, K.H.; Lim, C.J. Ginsenoside Rc protects against UVB-induced photooxidative damage in epidermal keratinocytes. Mol. Med. Rep. 2017, 16, 2907–2914. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Parameter |
|---|---|
| Capillary (kV) | 4 |
| Cone (V) | 100 |
| Desolvation temperature (°C) | 450 |
| Desolvation (L/h) | 700 |
| Cone (L/h) | 30 |
| Ginsenoside | Adduct Formula | Monoisotopic Mass | Parent (m/z) | Daughter (m/z) |
|---|---|---|---|---|
| Rb1 | [M + Na]+ | 1109.31 | 1131.59 | 335.11 |
| Rb2 | [M + Na]+ | 1079.27 | 1101.65 | 335.04 |
| Rb3 | [M + Na]+ | 1079.27 | 1101.59 | 365.08 |
| Rc | [M + Na]+ | 1079.27 | 1101.46 | 334.98 |
| Rd | [M + Na]+ | 947.15 | 969.39 | 789.36 |
| Re | [M + Na]+ | 947.16 | 969.46 | 789.35 |
| Rg1 | [M + Na]+ | 800.49 | 823.50 | 643.32 |
| NG R1 | [M + Na]+ | 933.13 | 955.59 | 203.04 |
| Concentration (μg/mL) | Blank Control | Positive Control | 100 | 200 | 500 | 1000 |
| Inhibition ratio (%) | 22.58 ± 1.22 | 92.01 ± 4.49 | 46.86 ± 1.37 ** | 52.11 ± 5.02 ** | 65.50 ± 4.33 ** | 82.41 ± 3.56 ** |
| Concentration (μg/mL) | 50 | 100 | 200 | 500 | 1000 |
| Cell survival Rate (%) | 97.80 ± 1.89 | 92.14 ± 6.09 | 84.26 ± 3.35 | 83.87 ± 2.47 | 77.64 ± 4.56 |
| Compound | Calibration Curve | r2 | Linear Ranges (μg/mL) | LOQ (ng/mL) | LOD (ng/mL) | Precision RSD (%) | Stability RSD (%) | Repeatability RSD (%) | Recovery (%) |
|---|---|---|---|---|---|---|---|---|---|
| Rb1 | Y = 50.1255x − 3713.21 | 0.9926 | 0.1–100 | 0.004 | 0.002 | 3.45 | 4.22 | 2.51 | 96.21 |
| Rb2 | Y = 53.2646x − 4165.20 | 0.9911 | 0.1–100 | 0.045 | 0.025 | 2.37 | 4.53 | 3.21 | 94.43 |
| Rb3 | Y = 30.9952x − 3740.43 | 0.9999 | 0.1–100 | 0.010 | 0.005 | 3.55 | 4.12 | 3.76 | 103.76 |
| Rc | Y = 37.1653x + 1932.67 | 0.9980 | 0.1–100 | 2 | 0.5 | 1.28 | 3.66 | 4.19 | 95.64 |
| Rd | Y = 7.1643x + 940.04 | 0.9970 | 0.1–100 | 1 | 0.2 | 2.32 | 3.76 | 4.72 | 92.57 |
| Re | Y = 8.5135x + 3545.47 | 0.9984 | 0.1–100 | 1 | 0.5 | 3.97 | 3.42 | 3.11 | 97.49 |
| Rg1 | Y = 3.2125x + 3114.94 | 0.9991 | 0.1–100 | 1.5 | 0.6 | 2.43 | 4.51 | 2.57 | 103.52 |
| NG R1 | Y = 2.9907x + 210.58 | 0.9975 | 0.1–100 | 2 | 0.8 | 3.64 | 3.73 | 3.96 | 106.75 |
| No. | Ginsenoside | PNF (μg/mg) |
|---|---|---|
| 1 | NG R1 | 1.98 ± 0.38 |
| 2 | Re | 1.62 ± 0.36 |
| 3 | Rg1 | 2.78 ± 0.58 |
| 4 | Rb1 | 83.06 ± 1.24 |
| 5 | Rc | 163.66 ± 0.63 |
| 6 | Rb2 | 108.53 ± 1.45 |
| 7 | Rb3 | 223.40 ± 0.22 |
| 8 | Rd | 79.42 ± 0.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Wu, Y.; Ma, W.; Zhang, H.; Meng, X.; Zhang, H.; Guo, M.; Ling, X.; Li, L. Anti-Inflammatory Activity of Panax notoginseng Flower Saponins Quantified Using LC/MS/MS. Molecules 2023, 28, 2416. https://doi.org/10.3390/molecules28052416
Liu J, Wu Y, Ma W, Zhang H, Meng X, Zhang H, Guo M, Ling X, Li L. Anti-Inflammatory Activity of Panax notoginseng Flower Saponins Quantified Using LC/MS/MS. Molecules. 2023; 28(5):2416. https://doi.org/10.3390/molecules28052416
Chicago/Turabian StyleLiu, Junchen, Yuehang Wu, Wenrui Ma, Hongyan Zhang, Xianyao Meng, Huirong Zhang, Miaomiao Guo, Xiao Ling, and Li Li. 2023. "Anti-Inflammatory Activity of Panax notoginseng Flower Saponins Quantified Using LC/MS/MS" Molecules 28, no. 5: 2416. https://doi.org/10.3390/molecules28052416
APA StyleLiu, J., Wu, Y., Ma, W., Zhang, H., Meng, X., Zhang, H., Guo, M., Ling, X., & Li, L. (2023). Anti-Inflammatory Activity of Panax notoginseng Flower Saponins Quantified Using LC/MS/MS. Molecules, 28(5), 2416. https://doi.org/10.3390/molecules28052416